Konijnenburgia, a new genus of the fern family Matoniaceae
12
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Konijnenburgia, a new genus of the fern family Matoniaceae

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Konijnenburgia, a new genus of the fern family Matoniaceae
Jiří Kvaček a,⁎, Jiřina Dašková b
a National Museum, Prague, Václavské nám. 68, 115 79, Praha 1, Czech Republicb Institute of Geology, Academy of Sciences of the Czech Republic, v.v.i., Rozvojová 269, 16500, Praha 6 — Lysolaje, Czech Republic
a b s t r a c ta r t i c l e i n f o
Article history:Received 25 May 2009Received in revised form 5 October 2009Accepted 11 October 2009Available online 23 October 2009
Keywords:KonijnenburgiaNathorstiaMatoniaceaeLate Cretaceous
A new genus Konijnenburgia is introduced for fertile, well preserved ferns of the family Matoniaceae, whichwere previously assigned to the genus Nathorstia Heer. It is based on Konijnenburgia latifolia (Nathorst)comb. nov. from the Upper Cretaceous of Greenland. The lectotype for Konijnenburgia latifolia is designatedand its status is discussed. Konijnenburgia bohemica is described from the Upper Cretaceous, Cenomanian ofthe Czech Republic and compared to Konijnenburgia latifolia and other Cretaceous members of the family.Introduction of the genus Konijnenburgia induced new combinations which are the following: Konijnen-burgia galleyi (Miner) comb. nov. and Konijnenburgia alata (Halle) comb. nov.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
The relict familyMatoniaceae is represented in today's flora by twogenera Matonia R. Brown ex Wallich and Phanerosorus Copel. In theMesozoic however, the family had a much wider distribution (VanKonijnenburg-van Cittert, 1993), occurring in both hemispheres (e.g.Harris, 1980; Givulescu and Popa, 1998; Skog, 2001; Van Konijnen-burg-van Cittert, 2002; Klavins et al., 2004; Nagalingum and Cantrill,2006) and underwent major diversification. Even in Cretaceous timesthe family still showed high diversity and in addition to the generaPhlebopteris Brongniart (1836), Matonia and Delosorus Skog (1988)others also existed, e.g. Monheimia Debey and Ettingshausen (1859)(Herman and Kvaček, 2009) and Nathorstia Heer (1880). The latergenus was however for many years a source of repeated problems andconfusion. Detailed study of the type material of Nathorstia angusti-folia Heer revealed that the type material does not show any detail,consisting of poorly preserved impressions and no other materialfrom the type locality is available (Kvaček and Dašková, 2007). Thelast emendation of Nathorstia by Krassilov (1967) was not based onthe type material of Nathorstia, but on material which Krassilov(1967) assigned to N. pectinata (Göppert) Krassilov. His wellpreserved material comes from the Far East of Russia, from thelocality Lipovci (Krassilov, 1967). The type of N. pectinata howevercomes from the vicinity of Moscow (Göppert in Murchison et al.,1845). The specific homogeneity of the material collected by Krassilov(1967) and the type N. pectinata is very questionable due to poorpreservation of the type.
After a revision of Nathorstia (Kvaček and Dašková, 2007) werealized the necessity for the introduction of a new genus whichwould be established on well preserved material showing clearcharacteristics of the family. In the present paper we designate a newgenus Konijnenburgia for well preserved fossil material previouslyassigned to the genus Nathorstia. Considering the necessity for alsoaccommodating poorly preserved fern material, we emended adiagnosis of Nathorstia as a morphogenus for simple pinnate fernfronds bearing circular sori without other detailed characteristics(Kvaček and Dašková, 2007).
Ferns with large circular indusia and wedge shaped sporangiawere widespread in Cretaceous times and seem to be present in bothhemispheres (Nathorst, 1908; Halle, 1913; Krassilov, 1967 and thepresent paper). Their massive umbrella-like indusia are known fromnumerous bulk macerated samples of charcoalified mesofossils fromboth hemispheres (J. Kvaček, personal observation).
As pointed out already by Kramer and Tryon (1990), reproductiveorgans are important for understanding fern systematics. Thereforedetailed study of the sorus structure in the Matoniaceae as stressed byKlavins et al. (2004) can greatly improve our understanding ofsystematics within the family. In our contribution we elaborated theconcept proposed by Klavins et al. (2004) and combine it withtraditional understanding of evolutionary pathways proposed by VanKonijnenburg-van Cittert (1993). As proved by Klavins et al. (2004)evolution of the indusium must have been completed by the MiddleTriassic and presence or absence of an indusium as proposed byHirmer and Hörhammer (1936) cannot be used as a character.Matoniaceae ferns easily lose their indusia. However, there are othergood characters, associated with the arrangement of sporangia in asorus, which can help us to resolve fossil generawithin the family. Thisimplies another necessary step — designation of morphogenera for
Review of Palaeobotany and Palynology 158 (2010) 308–318
⁎ Corresponding author.E-mail addresses: [email protected] (J. Kvaček), [email protected] (J. Dašková).
0034-6667/$ – see front matter © 2009 Elsevier B.V. All rights reserved.doi:10.1016/j.revpalbo.2009.10.002
Contents lists available at ScienceDirect
Review of Palaeobotany and Palynology
j ourna l homepage: www.e lsev ie r.com/ locate / revpa lbo

Author's personal copy
those fossils preserved as impressions. As we have already suggestedfor Nathorstia, there should be a similar solution for Phlebopteris andfossil Matonia and perhaps also for other genera of the Matoniaceae.However this is out of the scope of the present paper. In agreementwith Klavins et al. (2004) we propose these sporangial characters asthemost taxonomically importantwithin theMatoniaceae: 1. shape, 2.number per sorus, 3. number of layers, 4. type of attachment and 5.orientation of annulus. In the case of annulus at compressed materialone must be careful because its orientation may be altered by collapseof the sporangium.
The form genusMatonisporites had been defined by Couper (1958)for dispersed spores. This genus is characterised by triangular triletespores with long laesure, raised commissures flanked by a margo andinterradial and apical thickenings. Their sculpture is smooth, sporesare thick and generally large.
An extensive survey of the fossil spores of the Matoniaceae wasreported by Van Konijnenburg-van Cittert (1993). She processedeight genera of the fossil Matoniaceae (including older studies focusedon Matoniaceae in situ spores) ranging from the Triassic to theCretaceous: Phlebopteris Brongniart 1836, Selenocarpus Schenk 1867,Matonia R. Brown ex Wallich 1829, Matonidium Schenk 1871, Pia-zopteris Lorch 1967, Nathorstia Heer 1880, Delosorus Skog 1988 andWeichselia Stiehler 1857.
2. Material
The plant material described in this paper comes from two areas:west Greenland and the Czech Republic (Central Europe). It is storedin the National Museum, Prague (NMP) and Naturhistoriska Riksmu-seet Stockholm (NHRM).
The Czech material was collected from The Peruc–KorycanyFormation, which is the basal most part of the Bohemian CretaceousBasin, as defined by Čech et al. (1980) (Fig. 1). The basin is in filled byUpper Cretaceous freshwater, brackish and marine sediments ofCenomanian to Campanian age. Detailed biostratigraphical informa-tion regarding the Bohemian material is based on the pollen spectra(Pacltová, 1977) and dated the Peruc–Korycany Formation as theupper part of the middle Cenomanian.
Thematerial comes from a temporary excavation for the basement ofthe hotel Pramen in Hloubětín–Hutě — a north east suburb of Prague(50°06′34″N, 14°33′20″E). The specimenswere collectedandpartly bulkmacerated in the period 1989–1991. The material was described for thefirst timeby J. Kvaček (1992a,b). Aprofile sectionof the localitywasneveraccessible. Its palaeoenvironment canbe estimated according to theplant
assemblage as swampy forest with prevailing taxodiaceous conifers(Quasisequoia crispa (Velenovský) J. Kvaček, “Sequoia” heterophyllaVelenovský) and angiosperms (Mauldinia bohemica Eklund and J. Kvaček,Grevilleophyllum constans (Velenovský) Velenovský), with an influx ofupland flora. The fragmentary nature of the fossil material and stabilepresence of charcoalified mesofossils (Kvaček and Eklund, 2003; Pátováand Kvaček, 2006; Kvaček and Friis, in press) suggest the matoniaceousferns in the Bohemian Cretaceous Basin grew on uplands perhapsforming fern prairies.
The Greenland material we have studied comes from the westerncoast of Greenland, Nugssuaq Peninsula, Sarqaq, locality Ataneker-dluk, Klyftan 09, where it was collected by the Nordenskiöld'sexpedition in the year 1883.
The Atanikerdluk locality on the Island of Disko shows largeprofiles of the Atane Formation, which belong to the Late Cretaceous,more precisely Late Turonian to Coniacian (Rosenkrantz, 1970). TheAtane Formation represents a 1000–1500 m thick series consisting ofsandstones and shale with coal seams (Pedersen, 1976). The studiedmaterial is preserved as leaf compressions in dark grey brownmudstone. Its precise lithological and stratigraphical position is notknown, because the section was not revisited.
3. Methods
The material was examined using an Olympus SZX 12 binocularmicroscope, Olympus BX 50 and Zeiss Axioskop 2 plus light microscope(with plan-neofluar objectives) and photographed with a microscopecamera, AxioCam MRC and Olympus DP 70. SEM observations wereundertakenusing anSEMCamera SX100 (Institute of Geology, Academyof Sciences of the Czech Republic, v.v.i.), SEM JEOL JSM-6380LV(Institute of Geology and Palaeontology, Faculty of Science, CharlesUniversity, Czech Republic). Macrophotographs were taken with adigital camera Canon 300Dwith a Sigma 1:2.8 macro lens. Images wereadjusted using Adobe Photoshop. 7.0.
Spores in situ were prepared using standard methodology (e. g.Batten, 1999; Collinson, 1999). After sampling fragments of fossilmaterial, hydrofluoric acid (HF, 4%) was applied to dissolve remainingsilica and silicates, followed by maceration in Schultze's solution.Maceration of sori, which were obtained from bulk maceration wascarried out with particular care. The sori were treated in Schultze'ssolution for only a few seconds to remove only leaf or indusium,leaving the sporangia in place. Coalified material was washed in 5%KOH. Part of the extracted spores were mounted in glycerine jelly (2or 3 preparations from one sample) for light microscopy. Part of the
Fig. 1. Location of the Bohemian Cretaceous Basin in Central Europe (left) and location of the fossil site (right). Dark grey area indicates Cretaceous Basin, light area indicatesBohemian Massif (after Uličný et al., 1997).
309J. Kvaček, J. Dašková / Review of Palaeobotany and Palynology 158 (2010) 308–318

Author's personal copy
material was prepared on SEM stubs and observed using standardtechnique (Collinson, 1999).
4. Systematics
Class: PolypodiopsidaOrder: GleichenialesFamily: MatoniaceaeGenus: Konijnenburgia gen. nov.Syn.Nathorstia Heer 1880 pro parteType: Nathorstia latifolia Nathorst 1908, p. 17, pl. 3, figs. 61–76.Etymology: In honour of Johanna H. A. van Konijnenburg-van Cittert,who greatly improved our knowledge of Mesozoic ferns and theirspores.Diagnosis.
Fronds pinnate, pinnule linear lanceolate. Secondary veins leavingthe midrib at right angles. Sori distributed in two rows parallel to themidrib. Each sorus is tightly covered by large indusium. Wedgeshaped, thin-walled sporangia radially arranged. Cells of annulusoriented horizontally — parallel to indusium. Spores ofMatonisporitestype: trilete, triangular in equatorial view, laesurae long withintraradial thickenings and distinct, exina psilate.Discussion. The genus Nathorstia Heer and its status was alreadydiscussed in a separate study (Kvaček and Dašková, 2007). The genusNathorstia is here considered in contrast with Krassilov (1964) andVan Konijnenburg-van Cittert (1993), as a morphogenus accommo-dating impressions of leaves resembling Matoniaceous ferns by theirmorphology, but without preserved details of sori precluding theirdirect identification with the family Matoniaceae.
Another problem is the generic name Reussia Presl in Sternberg(1838), which was used by Göppert (1836) for Reussia pectinataGöppert and mentioned by Krassilov (1967) when he combined itwith Nathorstia pectinata (Göppert) Krassilov. The genus Reussia isbased on the type Reussia scolopedrioides (Brongniart) Presl inSternberg (1838) and should have priority over Nathorstia Heer(1880). Involvement of Reussia may help solve the problem withNathorstia, but the genus Reussia Presl cannot be used because it isnomen rejiciendum (Reussia Endlicher, 1836 nom conserv.). Addition-ally, revising the genus Nathorstia Krassilov (1964) suggested N.pectinata as a type of Nathorstia instead of N. angustifolia. This ishowever against ISBN rules (McNeill et al., 2006).
We came to the conclusion that the only possibility is to describe anew genus bearing characters of “Nathorstia” in the sense of Krassilov(1967) and Van Konijnenburg-van Cittert (1993). The first described,well preserved species of Nathorstia was N. latifolia Nathorst (1908).Its soral anatomy was precisely documented by Nathorst (1908) andspores by Van Konijnenburg-van Cittert (1993). This fern is suggestedto be a type for a newly designed genus, which can accommodate allthose ferns which were assigned to Nathorstia by Van Konijnenburg-van Cittert (1993) — N. pectinata, N. angustifolia, N. alata, N. galley.They are leaf compressions with well preserved sori showing radialarrangement of wedge-shaped sporangia and Matonisporites spores.These spores are similar to several species of dispersed smooth andless cutinized spores of Matonisporites Couper.
The newly introduced genus Konijnenburgia differs from MatoniaR. Brown ex Wallich 1829, Phanerosorus Copeland 1908, Phlebopteris
Brongniart 1836, Matonidium Schenk 1871, Selenocarpus Schenk 1867,Piazopteris Lorch 1967, Delosorus Skog 1988, Weichselia Stiehler 1857,Tomaniopteris Klavins et al. 2004 in having wedge-shaped instead ofovoid sporangia arranged in one layer (Plate II4, III1) and annulusoriented horizontally (Plate V4). It also differs from ParamatoniaMillerand Hickey, 2008 in the absence of details of sori and in situ spores.Sporangia ofKonijnenburgia are thin-walled and usually dorso-ventrallyflattened instead of thick-walled ovoid sporangia as in Matonia,Phanerosorus, Phlebopteris, Tomaniopteris and other representativesof the family. The annulus in Konijnenburgia is oriented horizontally,parallel to the level of the indusium in contrary to other representativesof the family which have the annulus oriented obliquely or vertically.
Konijnenburgia latifolia (Nathorst) comb. nov.
Plate I1–7; Plate II1–4; Plate III 1–6Basionym: Nathorstia latifolia Nathorst 1908, Kunglige Svenska Vetens-kapskademiens Handlingar, Bd. 43, No. 6, p. 17, pl. 3, figs. 61–76
Type locality: Atanekerdluk, Klyftan 09, Sarqaq, Nugssuaq peninsula,Greenland.
Type horizon: Late Cretaceous, Cenomanian.
Lectotype designated here in: NHRS S 108112, Nathorst 1908, pl. 3,fig. 2.
Other material: NHRS S101201, NHRS S101202, NHRS S108104–NHRSS108111, NHRS S108113, NHRS S108114, NHRS S108828–NHRSS108830, NHRS S108836, NHRS S108837, NHRS S108844.Emended diagnosis. Simple pinnate/deeply lobed leaves with leafletshaving sori in regular rows on either side of themidrib. Sterile pinnuleshortly petiolate with constricted bases. Sori large, circular, consistingof wedge-shaped sporangia, attached basally in a single layer, tightlyand completely covered by large indusium.
Description. The lectotype represents a fragment of an entire-marginedpinnula 65 mm long and 11mm wide (Plate I1). From the massivemidven (0.8–1 mm wide), short secondaries emerge at right angles(Plate II1, 3), they reach half the distance between the midrib andlamina margin (Plate II1). Tertiary venation is difficult to observe, it isbelieved to be reticulate. Circular sori 1.5 mm in diameter are arrangedin two rows on both sides of the midrib (Plate I1–3, II2). The sori aredensely arranged. In the specimen No. S 108113 the sori are so densethat their circular outlines are deformed into a rectangular shape (PlateI2). Sori are covered by a large umbrella-like indusium. Sporangia havethin walls and poorly preserved annulus in marginal parts (Plate I7).They are wedge shaped (250×750 μm) and arranged radially in onelayer. Their number is estimated to be about 20–26 per sorus. Othertype material shows sterile bipinnate fronds with petiolate pinnae(Plate II1) 15 mm wide with constricted bases. Some leaf impressionswith sori show clearly the radial arrangement of sporangia (Plate II4).The sporangia consist of numerous smooth spores with well pro-nounced leasurae. They are of Matoniasporites type (Plate III2–6)recalling M. suemegensis Juhász from the Upper Baremian of Hungary(Juhász, 1979). Spores are trilete, laesurae distinct, reaching almost toequator (not more than 80% of spore radius), commissures bordered bya distinct margo, distinctly raised, equatorial contour triangular withslightly convex sides, exine psilate, no perispore preserved, not greatlythickened at apices; mean spore diameter 35 μm (Plate III2–6).
Plate I. Konijnenburgia latifolia (Nathorst) comb. nov., Greenland, Atanekerdluk.
1. Lectotype, fertile pinna showing sori, lectotype, NHRS S108112, Nathorst (1908), pl. 3, fig. 64, ×1.5.2. Fertile pinna showing rectangular sori, NHRS S108113, Nathorst (1908), pl. 3, fig. 65, ×4.3. Two fertile pinnae showing sori, NHRS S101202, ×2.4. Wedge shaped sporangium from the lectotype, NHRS S108112.5. Wedge shaped sporangium from the lectotype, with remains of sporangial wall, NHRS S 108112.6. Wedge shaped sporangium, NHRS S108113.7. Wedge shaped sporangium. Detail of fig. 6, lateral part of sporangium with remains of several thicker cells of annulus (arrowed), NHRS S108113.
310 J. Kvaček, J. Dašková / Review of Palaeobotany and Palynology 158 (2010) 308–318

Author's personal copy
311J. Kvaček, J. Dašková / Review of Palaeobotany and Palynology 158 (2010) 308–318

Author's personal copy
Discussion. As already stated by Nathorst (1908) and others, lastlyby Van Konijnenburg-van Cittert (1993), Konijnenburgia latifoliashows diagnostic characters of the Matoniaceae including frondstructure, arrangement of sori and type of spores. This species isknown only from the Cretaceous of Greenland. It differs from K.bohemica in having less robust sori, which when maceratedseparated into individual sporangia. Konijnenburgia latifolia differsfurther from K. bohemica in having a higher number (usually 20) ofsporangia per sorus. Konijnenburgia latifolia also differs from Konij-nenburgia alata (Halle) comb. nov. from the Lower Cretaceous ofArgentina (Halle, 1913) in having a higher number (20–26) ofsporangia per sorus, petiolate pinnae and non-winged rachis. K.latifolia differs from Konijnenburgia galleyi (Miner) comb. nov. fromthe Cenomanian of Kansas, USA (Miner, 1935) in a higher number ofsporangia per sorus (up to 16) and spores having raised laesuraewith interradial thickenings. For detailed comparisons see tab. 1 and
tab. 2. As already pointed out by Nathorst (1908), Drynaria fasciaBayer from the Cenomanian of the Czech Republic (Bayer, 1899)shows a very similar arrangement of sori. Due to its poorpreservation and lack of any anatomical details, particularly shapeof sporangia, the taxon is left in the genus Nathorstia under thename Nathorstia fascia (Bayer) Nathorst. Distinctions between K.latifolia and other genera were mentioned in the discussion of thegenus.
Konijnenburgia bohemica sp. nov.
Plate IV1–5; Plate V1–6; Plate VI1–6Syn:2006 Nathorstia cf. fascia (Bayer) Nathorst; Pátová et J. Kvaček, Areport of ferns of the Bohemian Cenomanian…, p. 91, pl. 1, Figs. 1–10
Etymology: after the name of the country — Bohemia.
Plate II. Konijnenburgia latifolia (Nathorst) comb. nov., Greenland, Atanekerdluk.
1. Fragment of sterile bipinnate leaf, NHRS S108104, Nathorst (1908), pl. 3, fig. 61, ×1.2.2. Fertile pinna impression bearing two rows of sori, detail of lectotype, NHRS S108112, ×11.3. Detail of Fig. 1 — sterile bipinnate leaf, NHRS S108104. ×2.2.4. Two sori showing radially arranged sporangia , NHRS S108114, Nathorst (1908), pl. 3, fig. 68, ×28.
Plate III. Konijnenburgia latifolia (Nathorst) comb. nov., Greenland, Atanekerdluk.
1. LM photo of radially arranged sori, coll. NHRS, Nathorst (1908), pl. 3, fig. 67, 72.2. SEM detail showing tightly packed spores, NHRS S101202.3. SEM photo of group of spores, NHRS S101202.4. SEM photo of group of spores, NHRS S101202.5. SEM photo of spore, proximal surface with trilete mark and interradial thickenings, NHRS S101202.6. SEM photo of distal surface of spore, NHRS S101202.
312 J. Kvaček, J. Dašková / Review of Palaeobotany and Palynology 158 (2010) 308–318

Author's personal copy
313J. Kvaček, J. Dašková / Review of Palaeobotany and Palynology 158 (2010) 308–318

Author's personal copy
Type locality: Hloubětín-Hutě, north east suburb of Prague.
Type horizon: Peruc–Korycany Formation, Cenomanian, LateCretaceous.
Holotype designated here: F3600a,b.
Other material: NMP F 2927a,b, F 2928a–e, F 3566–F 3575, F 3601–F 3605.
Diagnosis. Simple pinnate decurrent leaves with pinnulae having soriin regular rows on both sides of the alate midrib. Sori large, circular,consisting of wedge-shaped, thin-walled sporangia attached basallyin single layer, tightly and completely covered by large indusium.Spores with interradial thickenings.Description. The holotype represents a fragment of isolated lanceolateentire-margined pinnule (25 mm long and 6 mm broad) (Plate IV1,4). It shows two rows of sori on the abaxial surface, running parallel to
Plate IV. Konijnenburgia bohemica gen. et sp. nov., Praha, Hloubětín-Hutě.
1. Holotype, fragment of simple pinnate leaf, NMP F 3600a, ×1.5.2. Fragment of pinna showing arrangement of sori, NMP F 2697a, ×4.3. Fragment of simple pinnate leaf, NMP F 3601, ×1.4. Deatail of holotype, pinna showing arrangement of sori, NMP F 3600b., ×8.5. Fragment of simple pinnate leaf, NMP F 3602, ×4.
314 J. Kvaček, J. Dašková / Review of Palaeobotany and Palynology 158 (2010) 308–318

Author's personal copy
Plate V. Konijnenburgia bohemica gen. et sp. nov., Praha, Hloubětín-Hutě.
1. Isolated sorus covered by indusium, NMP F 2928a.2. Deatil of biseriate indusium, NMP F 2928b.3. Detail of biseriate margin of indusium, detail of fig. 1, NMP F 2928a.4. Detail of wedge shaped sporangium, note annalus running in horizontal direction , parallel to indusium, NMP F 3567.5. Portion of sporangium, Detail of Fig. 2, NMP F 2928b.6. Sporangial wall with thicker cells of annulus, NMP F 3567.
315J. Kvaček, J. Dašková / Review of Palaeobotany and Palynology 158 (2010) 308–318
Author's personal copy
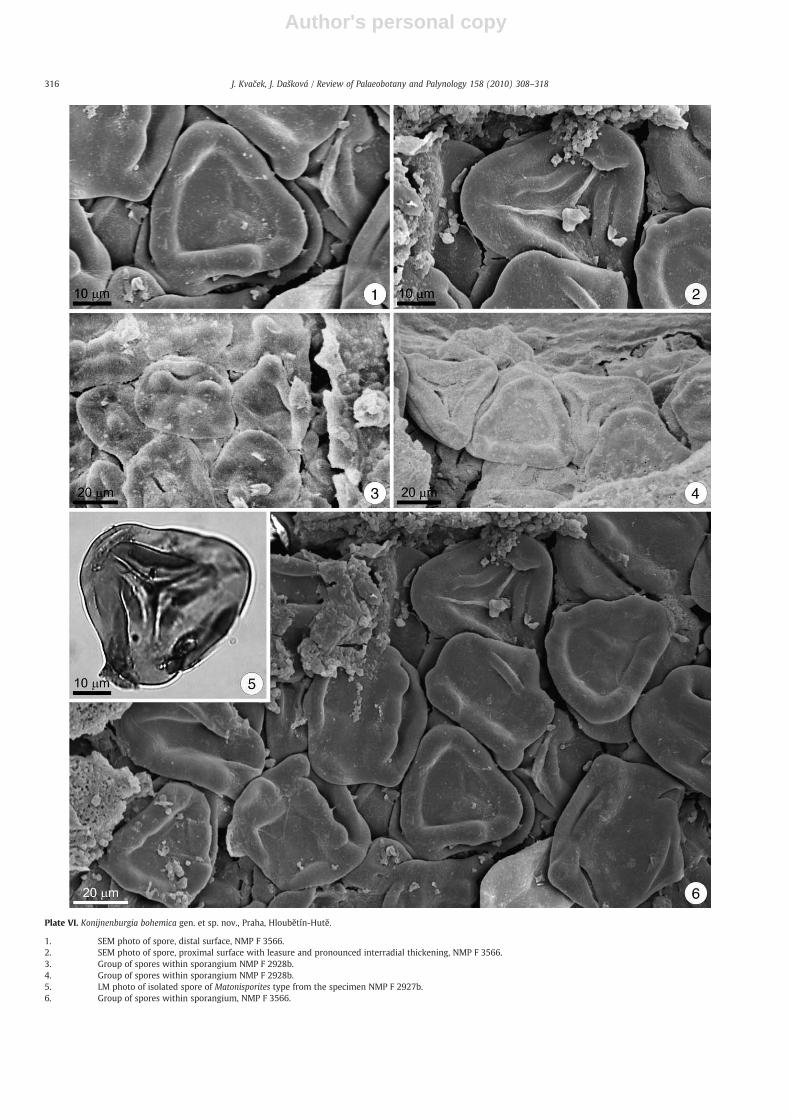
Plate VI. Konijnenburgia bohemica gen. et sp. nov., Praha, Hloubětín-Hutě.
1. SEM photo of spore, distal surface, NMP F 3566.2. SEM photo of spore, proximal surface with leasure and pronounced interradial thickening, NMP F 3566.3. Group of spores within sporangium NMP F 2928b.4. Group of spores within sporangium NMP F 2928b.5. LM photo of isolated spore of Matonisporites type from the specimen NMP F 2927b.6. Group of spores within sporangium, NMP F 3566.
316 J. Kvaček, J. Dašková / Review of Palaeobotany and Palynology 158 (2010) 308–318

Author's personal copy
the pinnule midvein. Each sorus is tightly covered by a massivecircular persistent indusium, 1.5–2 mm in diameter (Plate V1, 3). Theindusium is flat and relatively smooth, showing delicate radialstriation and biseriate margin (Plate V3). Distance between two soriis 2–2.5 mm (Plate IV1, 2, 4, 5). A sorus consists of 12–14 radiallyarranged wedge-shaped sporangia 500 μm long. Spores of Matoni-sporites type are trilete, triangular in equatorial outline, with slightlyconvex sides, leasurae raised, extending 75% of the spore radius;interradial thickenings well pronounced (Plate VI2); equatorialthickening (crassitude) present (Plate VI1, 5); exine psilate, noperispore preserved, thickness of the exospore 1.5 μm. Sporesdiameter in equatorial view ranges from 45 μm to 52 μm (meandiameter 47 μm) (Plate VI1–6). In some cases the compacted sorioccur isolated in bulk macerated samples.Discussion. Konijnenburgia bohemica is similar to K. latifolia. It differs incharacteristics of the sorus, which is in the case of K. bohemica coveredby a very thick indusium. A bulk macerated sorus of K. bohemica neverseparates into individual sporangia as in the case of K. latifolia. It alsodiffers in a number of sporangia per sorus anddetails of vegetative parts.For detailed comparison of K. latifolia and K. latifolia see the Table 1.Nathorstia fascia from Vyšehořovice was described by Bayer (1899)originally under the generic name Drynaria (Bory) J. Smith which is amember of the family Polypodiaceae. Revision of the material studiedby Bayer (1899), particularly type and arrangement of sori did notconfirm the previously suggested affinity to the Polypodiaceae andargue for the Matoniaceae (see Van Konijnenburg-van Cittert, 1993).However, N. fascia is preserved as a leaf impression and therefore itcannot be included to the genus Konijnenburgia gen. n. Due to lack ofany detailed information about sporangia and spores, Nathorstia fasciais left separately in the morphogenus genus designed for suchimperfectly preserved fern fronds.K. bohemica differs from Konijnenburgia alata (Halle) comb. nov. inhaving a slightly lower number of sporangia (12–14) and havinglarger spores with intraradial thickenings. K. bohemica differs fromKonijnenburgia galleyi (Miner) comb. nov. in having larger size soriand having larger spores with raised laesurae. K. galleyi very muchresembles the material described by Krassilov (1967) from the FarEast of Russia under the name Nathorstia pectinata (Göppert)Krassilov. For detailed comparisons see Table 1 and Table 2.
Spores Matonisporites found in situ in K. bohemica show similarthickenings as known in dispersed sporesMatonisporites phlebopter-oides Couper (1958). Matoniaceous spores are quite rare in dispersedsporomorph spectra of the Bohemian Cretaceous Basin and areusually identified as Matonisporites sp. (Svobodová personalcommunication).
5. Results
In the present paper we propose a new genus Konijnenburgia as amember of the family Matoniaceae. It shows the following mostimportant characteristics: wedge shaped sporangia in one layer with ahorizontally arranged annulus. Based on previously published data,we suggest that species of Nathorstia alata, Halle and N. galleyi(Miller) Van Konijnenburg-van Cittert should also be included in thenew genus Konijnenburgia. As a result of this assumption we proposethe following new combinations:Konijnenburgia galleyi (Minner) comb.nov., Basionym: Laccopteris galleyiMinner 1935, Contributions Mus. Palaeont. p. 288, pl. 1, figs. 1–4.Konijnenburgia alata (Halle) comb. nov., Basionym: Nathorstia alataHalle 1913 Kungl. Svensk. Vetensk Handl. 51, 3, p. 20, pl. 1, figs. 1–9.
Acknowledgements
We thank Marcela Svobodová for consultations on Matonisporitesdispersed spores and Thomas Denk andOve Johansson (NaturhistoriskaRiksmuseet, Stockholm) for their help with the collections. This workwas supported by the Grant Agency of ASCR, v.v.i. (project No. IAA304070701), Ministry of Culture of the Czech Republic (project DE06P04OMG009) and by the Academy of Sciences of the Czech Republic(Z 3013 0516). The paper profited from constructive comments of twoanonymous referees.
References
Batten, D.J., 1999. Small palynomorphs. In: Jones, T.P., Rowe, N.P. (Eds.), Fossil plantsand spores. The Geological Society, London.
Bayer, E., 1899. Einige neue Pflanzen der Perucer Kreideschichten in Bohmen.Sitzungsber. Kgl. Böhm. Ges. Wiss., Math-nat. Cl. 30, 1–51.
Brongniart, A.T., 1828–1838. Histoire des végétaux fosilles, ou recherches botaniques etgeologiques sur les végétaux renfermés dans les diverses couches du globe. vol. I, 1(1828): 1 – 80 pp.; vol. I, 2 ( 1828): 81–136 pp.; vol. I, 3 ( 1829): 137–168 pp.; vol. I,4 (1830): 169–208 pp.; vol. I, 5 (1831): 209–248 pp.; vol I, 6 (1831): 249–264 pp.;vol. I, 7 (1833): 265–288 pp.; vol. I, 8 (1834): 289–312 pp.; vol. I, 9 (1834): 313–336 pp.; vol. I, 10 (1836): 337–368 pp.; vol. I, 11 (1837): 369–416 pp.; vol. I, 12(1837): 417–488 pp.; vol. II, 13 (1837): 1–24 pp.; vol. II, 14 (1838): 25–56 pp.; vol.II, 15 (1838): 57–72 pp. G. Dufour et D´Ocagne, Paris.
Čech, S., Klein, V., Kříž, J., Valečka, J., 1980. Revision of the Upper Cretaceous stratigraphyof the Bohemian Cretaceous Basin. Věst. Ústř. úst. geol. 55 (5), 277–296.
Collinson, M.E., 1999. Scanning electron microscopy of megafossils and mesofossils. In:Jones, T.P., Rowe, N.P. (Eds.), Fossil plants and spores: modern techniques.Geological Society, London.
Copeland, E.B., 1908. New genera and species of Bornean ferns. Philipp. J. Sci., Sect. C 3,343–351.
Couper, R.A., 1958. British Mesozoic microspores and pollen grains. Palaeontogr.Bohem. 103, 75–79.
Table 1Comparison of species of Konijnenburgia.
Diameter of sorus Number of sporangia Pinnae decurrent Pinnae petiolate Midrib alate
Konijnenburgia latifolia 1.5 mm 18–24 No Yes NoKonijnenburgia alata 1–1.5 mm 12–15 Yes No YesKonijnenburgia galleyi 1.3–1.7 mm 11–16 Unknown Unknown UnknownNathorstia pectinata sensu Krassilov (1967) 2 mm 12–15 No No UnknownKonijnenburgia bohemica 2 mm 12–14 Yes No Yes
Table 2Comparison of matoniaceous spores: Konijnenburgia latifolia, K. alata, K. gelleyi, Nathorstia pactinata sensu Krassilov (1967) (according to Van Konijnenburg-van Cittert, 1993) and K.bohemica.
Equatorial diameter Laesurae raised Length of radii Interradial thickenings
Konijnenburgia latifolia 38 Yes 75% YesKonijnenburgia alata 30 Yes 90% NoKonijnenburgia galleyi 28 No 90% NoNathorstia pectinata sensu Krassilov (1967) 30 Yes 90% Probably yesKonijnenburgia bohemica 43 Yes 75% Yes
317J. Kvaček, J. Dašková / Review of Palaeobotany and Palynology 158 (2010) 308–318

Author's personal copy
Debey, M.H., Ettingshausen, C., 1859a. Die urweltlichen Acrobryen des Kreidegebirgesvon Aachen und Maastricht. Denkschr. Österreich. Akad. Wiss., Math. — nat. Kl. 17,183–248.
Endlicher, S.L. 1836–41. Genera plantarum secundum ordines naturales disposita. Part1–2 (1836): 1–160 pp.; part 3–5 (1837): 161– 400 pp.; part 6–8 (1838): 401–640 pp.; part 9–12 (1839): 641–960 pp.; part 13–17 (1840): 961–1360 pp.; part 18(1841): 1360–1483 pp. Fr. Beck, Wien.
Givulescu, R., Popa, M.E., 1998. Aninopteris formosa n. gen, n. sp., a new LiassicMatoniaceous genus and species from Anina, Banat, Romania. Rev. Palaeobot.Palynol. 104, 51–66.
Göppert, H.R., 1836. Die fossilen Farrenkräuter (Systema filicium fossilium). Nova ActaLeopold. 17, 1–486.
Halle, T.G., 1913. Some Mesozoic plant-bearing deposits in Patagonia and Tierra delFuego and their floras. K. Sven. Vetensk.akad. Handl. 51 (3), 1–58.
Harris, T.M., 1980. The Yorkshire Jurassic fern Phlebopteris braunii (Goeppert) and itsreference to Matonia. 5R. Br. Bull. Br. Mus. Nat. Hist (Geol.) 33, 295–311.
Heer, O., 1880. Nachträge zur fossilen Flora Grönlands. K. Sven. Vetensk.akad. Handl. 18(2), 1–17.
Herman, A.B., Kvaček, J., 2009. Late Cretaceous Grünbach Flora of Austria. Naturhistor-isches Museum, Wien, 219 pp.
Hirmer, M., Hörhammer, L., 1936. Morphologie, Systematik und geographischeVerbreitung der fossilen und rezenten Matoniaceen. Palaeontogr. Bohem. 81, 1–66.
Juhász, M., 1979. Dispersed Matoniaceae spores from the Hungarian Lower and MiddleCretaceous sediments. Acta Biol. Szeged 23 (1–2), 33–47.
Klavins, S.D., Taylor, T.N., Taylor, E.L., 2004. Matoniaceous ferns (Gleicheniales) fromthe Middle Triassic of Antarctica. J. Paleontol. 78 (1), 211–217.
Kramer, K.U., Tryon, R.M., 1990. Introduction to the treatment of pteridophytes. In:Kubitzki, K. (Ed.), The Families and Genera of Vascular Plants, volume I,Pteridophytes and Gymnosperms. Springer Verlag, Berlin.
Krassilov, V.A., 1964. On the genus Nathorstia (ferns). Paleontol. J. (Moscow) 2,115–120 (in Russian).
Krassilov, V.A., 1967. Early Cretaceous flora of southern Primorya and its significance forthe stratigraphy. Nauka, Moscow. 248 pp.
Kvaček, J., 1992a. Leaf and fruit compressions from the Bohemian Cenomanian. In:Kovar-Eder, J. (Ed.), Palaeovegetational development in Europe and regionsrelevant to its palaeofloristic evolution, Proceedings of the Pan-EuropeanPalaeobotanical Conference, Vienna, 19–23 September 1991. Museum of NaturalHistory, Vienna, pp. 301–304.
Kvaček, J., 1992b. A new Cenomanian floral locality from Praha [Nová lokalitacenomanské flóry v Praze]. J. Nat. Mus. (Prague), Nat. Hist. Ser., 158 (1–4), 42.
Kvaček, J., Dašková, J., 2007. Revision of the type material to the genus Nathorstia Heer.J. Nat. Mus. (Prague), Nat. Hist. Ser., 176, 117–123.
Kvaček, J., Eklund, H., 2003. A report on newly recovered reproductive structures fromthe Cenomanian of Bohemia (Central Europe). Int. J. Plant Sci. 164 (6), 1021–1039.
Kvaček, J., Friis, E.M., in press. Zlatkocarpus gen. nov, a new angiosperm reproductivestructure with monocolpate-reticulate pollen from the Late Cretaceous (Cenoma-nian) of the Czech Republic. Grana.
Lorch, J., 1967. Jurassic flora of Makhtesh Ramon, Israel. Isr. J. Botan. 16, 131–155.McNeill, J., Barrie, F.R., Burdet, H.M., Demoulin, V., Hawksworth, D.L., Marhold, K.,
Nicolson, D.H., Prado, J., Silva, P.C., Skog, J.E., Wiersema, J.H., Turland, N.J., 2006.
International code of botanical nomenclature (Vienna code). Gantner, Ruggell,Liechtenstein, p. 568.
Miller, I.M., Hickey, L.J., 2008. The fossil flora of the Winthrop Formation (Albian–EarlyCreatceous) of Washington State, USA. Part I: Bryophyta and Pteridophytina. Bull.Peabody Museum Nat. Hist. 49 (2), 135–180.
Miner, E.L., 1935. A new Laccopteris from the Cretaceous of Kansas. Contrib. Mus.Palaeontol., Univ. Mich. 4 (17), 287–290.
Murchison, R.I., de Verneuil, E., de Keyserling, A., 1845. Géologie de la Russie d'Europe etdes montagnes de l'Oural. Vol. II. 3e partie. Paléontologie (Reliure inconnue).J. Murray, Londres, Bertrand, Paris, p. 512.
Nagalingum, N.S., Cantrill, D.J., 2006. Early Cretaceous Gleicheniaceae and Matoniaceae(Gleicheniales) from Alexander Island, Antarctica. Rev. Palaeobot. Palynol. 138,73–93.
Nathorst, A.G., 1908. Über Nathorstia Heer. Palaeobotanical Mitteilungen 5. K. Sven.Vetensk.akad. Handl. 43 (6), 14–19.
Pacltová, B., 1977. Cretaceous angiosperms of Bohemia — Central Europe. Bot. Rev. 43(1), 128–142.
Pátová, R., Kvaček, J., 2006. A report of ferns of the Bohemian Cenomanian and theirspores in situ. Acta Univ. Carol., Geol. 47 (1–4), 91–94.
Pedersen, K.R., 1976. Fossil floras of Greenland. In: Escher, A., Watt, W.S. (Eds.), Geologyof Greenland. The Geological Survey of Greenland, Odense.
Rosenkrantz, A., 1970. Marine Upper Cretaceous and lower-most Tertiary deposits inWest Greenland. Meddel. Dansk Geol. Foren. 19, 406–453.
Schenk, A., 1867. Die fossile Flora der Grenzschichten des Keupers und Lias in Franken.Kreidel, Wiesbaden. 232 pp.
Schenk, A., 1871. Die fossile flora der nordwestdeutschen Wealdenformation. TheodorFischer, Cassel. 62 pp.
Skog, J.E., 1988. Reassignment of Aspidium heterophyllum to a new genus in the familyMatoniaceae. Am. J. Bot. 75, 1120–1129.
Skog, J.E., 2001. Biogeographic distribution of Mesozoic ferns related to modern fernfamilies. Brittonia 53, 236–269.
Sternberg, K.M., 1820–1838. Versuch einer geognostisch-botanischen Darstellung derFlora der Vorwelt. — vol. I, 1(1820): 1–24 pp. F. Fleischer, Leipzig; vol. I, 2 (1821):1–33 pp. F. Fleischer, Leipzig; vol. I, 3 (1823): 1–39 pp. Ernst Brenck's Wittwe,Regensburg; vol. I, 4 (1825): 1–48 pp. tent. I–XLII, Ernst Brenck's Wittwe,Regensburg; vol. II, 5/6 (1833): 1–80 pp. Johann Spurny, Prag; vol. II, 7/8 (1838)(C.B. Presl and A.J. Corda coauthors): 81–220 pp. Gotlieb Hässe Söhne, Prag.
Stiehler, A.W., 1857. Beiträge zur Kenntniss der vorweltlichen Flora des Kreidegebirgesim Harz. Palaeontographica 5 (47–70), 71–80.
Uličný, D., Kvaček, J., Svobodová, M., Špičáková, L., 1997. High-frequence sea-levelfluctuations and plant habitats in Cenomanian fluvial to estuarine seccessions:Pecínov Querry, Bohemia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 136, 165–197.
Van Konijnenburg-van Cittert, J.H.A., 1993. A review of theMatoniaceae based on in situspores. Rev. Palaeobot. Palynol. 78, 235–267.
Van Konijnenburg-van Cittert, J.H.A., 2002. Ecology of some Jurassic ferns in Eurasia.Rev. Palaeobot. Palynol. 199, 113–124.
Wallich, N., 1829. Plantae Asiaticae rariores: or, Descriptions and figures of a selectnumber of unpublished East Indian plants, vol. 1 (1). Treuttel and Würtz, London.84 pp.
318 J. Kvaček, J. Dašková / Review of Palaeobotany and Palynology 158 (2010) 308–318




















