
A new genus of the family Jaculinidae (Cheilostomata, Bryozoa) from the Miocene of the tropical...
15
98 Accepted by D. Gordon: 11 Jun. 2014; published: 16 Jul. 2014 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2014 Magnolia Press Zootaxa 3838 (1): 098–112 www.mapress.com/zootaxa/ Article http://dx.doi.org/10.11646/zootaxa.3838.1.5 http://zoobank.org/urn:lsid:zoobank.org:pub:458183F5-78CF-4E00-AEA3-8C4477CF829B A new genus of the family Jaculinidae (Cheilostomata, Bryozoa) from the Mio- cene of the tropical western Atlantic KAMIL ZÁGORŠEK 1* , LAÍS V. RAMALHO 2 , BJÖRN BERNING 3 & VLADIMIR DE ARAÚJO TÁVORA 4 1 Central European Institute of Technology, Brno University of Technology, Technicka 10, CZ-616 00 Brno, Czech Republic [email protected] 2 Universidade Federal de Rio Grande – FURG, Instituto Oceanográfico - Laboratório de Oceanografia Geológica, Av. Itália, s/n, Carreiros, Rio Grande, Rio Grande do Sul, Brazil. E-mail: [email protected] 3 Oberösterreichisches Landesmuseum, Geowissenschaftliche Sammlungen, Welser Str. 20, 4060 Leonding, Austria. E-mail: [email protected] 4 Laboratório de Paleontologia, Faculdade de Geologia, Instituto de Geociências, Universidade Federal do Pará, Caixa Postal 8608, Belém-PA, Brazil. E-mail: [email protected] * Corresponding author Abstract Pirabasoporella gen. nov. is introduced for three new bryozoan species from the Early Miocene of the tropical western Atlantic. The genus is placed in the family Jaculinidae Zabala, a peculiar group of cheilostome bryozoans characterised by reticulate colonies formed by uni- or biserial branches that are connected by kenozooidal struts. This colonial morphology superficially resembles colonies of the Paleozoic order Fenestrata (Stenolaemata) and some Recent Cyclostomata. As jaculinid colonies are anchored to soft sediments via rhizoids, however, they differ in life habit from Paleozoic and modern fenestrate colonies, which are firmly attached to stable substrata by an encrusting base. The three new species are Pirabasoporella atalaiaensis n. sp. from the Brazilian Pirabas Formation, Pirabasoporella baitoae n. sp. from the Baitoa Formation (Dominican Republic), and Pirabasoporella chipolae n. sp. from the Floridan Chipola Formation. Their presence in the Early Miocene western Atlantic represents the earliest record of Jaculinidae, and suggests that the origin of the family, the only living species of which are known from the eastern Atlantic and Mediterranean Sea, extends well into the Paleogene. The Jaculinidae is here transferred from the lepraliomorph superfamily Schizoporelloidea Jullien to the umbonulomorph Lepralielloidea Vigneaux owing to the partly umbonuloid frontal shield and non-schizoporelloid ovicell. Key words: new genus and species, Pirabas Formation, Jaculinidae, palaeoenvironment, palaeobiogeography, systematics Introduction During investigations of the Cenozoic Pirabas Formation (Pará State, Brazil), numerous specimens were found that closely resembled species of the little-known, Pleistocene to Recent family Jaculinidae Zabala, 1986 from the northeastern Atlantic and Mediterranean Sea. Jaculinid species form reticulate colonies composed of uni- or biserial, subparallel branches that are connected by kenozooidal struts. Whereas the reteporiform colonies of modern ascophoran cheilostomes, such as those of the genus Reteporella Busk, 1884, connect neighbouring branches via trabeculae that consist of autozooids, species of Jaculinidae superficially rather resemble fenestrate colonies of Paleozoic stenolaemates, in which the connecting struts are formed by kenozooids. In contrast to these species, whose reticulate colonies are attached to stable substrata by an encrusting base, recent jaculinids are anchored to soft bottoms via rhizoids. The facies of the Pirabas Formation, in which the present specimens were found, also indicates this type of environment. The screening of museum collections comprising coeval bryozoan faunas from similar paleoenvironments revealed two additional species from the Caribbean region. As the western Atlantic taxa differ in a number of morphological traits from the NE Atlantic and Mediterranean representatives, a new jaculinid genus and three new species are introduced here.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of A new genus of the family Jaculinidae (Cheilostomata, Bryozoa) from the Miocene of the tropical...

ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2014 Magnolia Press
Zootaxa 3838 (1): 098–112
www.mapress.com/zootaxa/Article
http://dx.doi.org/10.11646/zootaxa.3838.1.5
http://zoobank.org/urn:lsid:zoobank.org:pub:458183F5-78CF-4E00-AEA3-8C4477CF829B
A new genus of the family Jaculinidae (Cheilostomata, Bryozoa) from the Mio-
cene of the tropical western Atlantic
KAMIL ZÁGORŠEK1*, LAÍS V. RAMALHO2, BJÖRN BERNING3 & VLADIMIR DE ARAÚJO TÁVORA4
1Central European Institute of Technology, Brno University of Technology, Technicka 10, CZ-616 00 Brno, Czech Republic
[email protected] Federal de Rio Grande – FURG, Instituto Oceanográfico - Laboratório de Oceanografia Geológica, Av. Itália, s/n,
Carreiros, Rio Grande, Rio Grande do Sul, Brazil. E-mail: [email protected]österreichisches Landesmuseum, Geowissenschaftliche Sammlungen, Welser Str. 20, 4060 Leonding, Austria.
E-mail: [email protected]ório de Paleontologia, Faculdade de Geologia, Instituto de Geociências, Universidade Federal do Pará, Caixa Postal 8608,
Belém-PA, Brazil. E-mail: [email protected]
* Corresponding author
Abstract
Pirabasoporella gen. nov. is introduced for three new bryozoan species from the Early Miocene of the tropical western
Atlantic. The genus is placed in the family Jaculinidae Zabala, a peculiar group of cheilostome bryozoans characterised
by reticulate colonies formed by uni- or biserial branches that are connected by kenozooidal struts. This colonial
morphology superficially resembles colonies of the Paleozoic order Fenestrata (Stenolaemata) and some Recent
Cyclostomata. As jaculinid colonies are anchored to soft sediments via rhizoids, however, they differ in life habit from
Paleozoic and modern fenestrate colonies, which are firmly attached to stable substrata by an encrusting base.
The three new species are Pirabasoporella atalaiaensis n. sp. from the Brazilian Pirabas Formation,
Pirabasoporella baitoae n. sp. from the Baitoa Formation (Dominican Republic), and Pirabasoporella chipolae n. sp.
from the Floridan Chipola Formation. Their presence in the Early Miocene western Atlantic represents the earliest record
of Jaculinidae, and suggests that the origin of the family, the only living species of which are known from the eastern
Atlantic and Mediterranean Sea, extends well into the Paleogene.
The Jaculinidae is here transferred from the lepraliomorph superfamily Schizoporelloidea Jullien to the
umbonulomorph Lepralielloidea Vigneaux owing to the partly umbonuloid frontal shield and non-schizoporelloid ovicell.
Key words: new genus and species, Pirabas Formation, Jaculinidae, palaeoenvironment, palaeobiogeography,
systematics
Introduction
During investigations of the Cenozoic Pirabas Formation (Pará State, Brazil), numerous specimens were found that closely resembled species of the little-known, Pleistocene to Recent family Jaculinidae Zabala, 1986 from the northeastern Atlantic and Mediterranean Sea. Jaculinid species form reticulate colonies composed of uni- or biserial, subparallel branches that are connected by kenozooidal struts. Whereas the reteporiform colonies of modern ascophoran cheilostomes, such as those of the genus Reteporella Busk, 1884, connect neighbouring branches via trabeculae that consist of autozooids, species of Jaculinidae superficially rather resemble fenestrate colonies of Paleozoic stenolaemates, in which the connecting struts are formed by kenozooids.
In contrast to these species, whose reticulate colonies are attached to stable substrata by an encrusting base, recent jaculinids are anchored to soft bottoms via rhizoids. The facies of the Pirabas Formation, in which the present specimens were found, also indicates this type of environment. The screening of museum collections comprising coeval bryozoan faunas from similar paleoenvironments revealed two additional species from the Caribbean region. As the western Atlantic taxa differ in a number of morphological traits from the NE Atlantic and Mediterranean representatives, a new jaculinid genus and three new species are introduced here.
98 Accepted by D. Gordon: 11 Jun. 2014; published: 16 Jul. 2014

FIGURE 1. Sketch of the studied Brazilian localities .FIGURE 2. Stratigraphic section at Salinopolis.
Material and methods
Four sections of the Pirabas Formation (Pará State, northern Brazil) were studied, all of which are Miocene in age (Fig. 1). The most speciose bryozoan fauna of all studied sections was found at Atalaia Beach near Salinópolis city (0°36’5” S, 47°18’48” W). The section comprises a sequence of fossiliferous stratified calcarenites, massive calcarenites, calcirudites, calcilutites and mudstones, as well as fossiliferous calcareous sandstones (Góes et al.
1990) cropping out on the sandy beach. The samples were taken from the silty layer just below the calcirudite (Fig. 2).
Section B-17 Mine is situated north of the city of Capanema (1°2’47” S, 47°9’26” W) in an active quarry (Távora et al. 2009). The stratigraphic section is composed of alternations of grey carbonate lithofacies and calcareous sandstones, and the samples were taken from clay intercalations within the limestone sequences in the upper part of the section.
The Fortaleza Island section (0°41’43” S, 47°10’23” W) includes stratified calcarenites, calcirudites, clays and calcareous sandstones (Góes et al. 1990). We sampled the calcirudite and the partly weathered stratified calcarenite, which contained the fewest number of bryozoan species in the association (Távora & Fernandes 1994; Távora et al. 2004).
The Aricuru section (0°42’10” S, 47°30’43” W) is characterized by argillaceous calcirudite interbedded with (sandy) marls. The upper part of the fine-grained sandstone yielded bryozoans (Távora & Fernandes 1999).
Altogether, a total of 43 samples of about 1-2 kg each from all four sections were studied: 28 samples from Atalaia Beach, 16 samples from B-17 Mine, four samples from Fortaleza Island, and five samples from Aricuru. Specimens of Pirabasoporella gen. nov. were found in 34 samples: Atalaia beach yielded specimens from 22 samples, B-17 Mine from 11 samples, Aricuru from one sample and none from Fortaleza Island.
The samples were washed in soda and sieved as described by Zágoršek et al. (2012), using sieves with 0.063, 0.9, 1.2 and 2 mm mesh diameter, at the Laboratório de Oceanografia Geológica, Federal University of Rio Grande (FURG), and Laboratório de Paleontologia, Federal University of Pará.
The samples from Brazil are stored in the paleontological collection of the Geoscience Museum of the Federal
Zootaxa 3838 (1) © 2014 Magnolia Press · 99A NEW GENUS OF JACULINIDAE FROM MIOCENE OF BRAZIL

University of Pará (MG-UFPA). The material from the Lower Miocene of Florida (Chipola Formation) and material from Dominican Republic (Baitoa Formation) is housed at the Department of Paleobiology, Natural History Museum, Smithsonian Institution, Washington, D.C. (USNM).
Geological setting
The Pirabas Formation (Maury 1925) occurs discontinuously in the Pará, Maranhão and Piauí states of northern Brazil, cropping out along coastal cliffs and road cuts and in mines. This unit predominantly consists of limestone that was deposited under warm, shallow marine conditions with normal salinity. The facies architecture within individual units revealed a dominantly progradational domain, reflected by the upsection increase in relatively more proximal facies associations that contain barrier islands (Távora et al. 2009).
The fossiliferous content is rich and diverse, comprising a majority of skeletized animal phyla with a predominance of molluscs, bryozoans and echinoids. The paleobiotic content confirms the affinity of the Pirabas Formation with other synchronous lithostratigraphic units of Central America, northern South America (Venezuela and Colombia) and the southern United States, and is included in the Caribbean Biogeographic Province (Woodring 1966, 1974, 1978; Távora et al. 2004).
The gastropod Orthaulax pugnax and the foraminifera Globorotalia opima were initially used to define the age of the Pirabas Formation as Late Oligocene to Early Miocene (Maury 1925; Ferreira 1967; Ferreira et al. 1981). Later, the occurrence of species of the genus Globorotalia (G. kugleri) and specially Globigerinoides (G. primordius, G. altiaperturus, G. trilobus) and Globoquadrina (G. dehiscens), allowed specifying the age of deposition as Early Miocene (Fernandes 1988; Fernandes and Távora 1990; Távora and Fernandes 1999). This is confirmed by ostracods: Pseudoceratina droogeri, Paracytheridea tschoppi, Pellucistoma magniventra, Costa
variabilocostata, Aurila amygdala, Cytherura elongata, Hermanites hornibrooki, H. tschoppi, Cushmanidea
howei, Munseyella muehlemanni and Quadracythere brachypigaia, as well as by the bryozoan genus Cupuladria, the first appearance datum of which is also Lower Miocene (Barbosa 1971; Ramos et al. 2004).
The Baitoa Formation forms up to 75 m of thick sequences in the Cibao Valley, northern Dominican Republic. The dominant sediments are conglomerates with intercalated silts, shales and fine conglomerates (Saunders et al. 1986). Based on ostracods and molluscs, the formation is late Early to early Middle Miocene in age (Saunders et al. 1986).
The Chipola Formation is exposed in the Florida Panhandle (Scott 2001). According to geochronologic investigations using Sr isotopes (Bryant et al. 1992; Jones et al. 1993) the age is estimated at 18.3–18.9 Ma, thus Early Miocene (Bryant et al. 1992). The Chipola Formation is composed of a repetition of clays, sandy clays to sands, and carbonate layers with abundant shell beds (Scott 2001).
Measurements are given in µm unless stated otherwise. Abbreviations used are:
AAL, AAW abfrontal avicularian length and widthAD aperture diameter (if apertural length and width have almost the same value)AL, AW autozooidal length and widthBW branch widthFL, FW fenestrule length and widthOL, OW ovicell length and widthRPL, RPW rhizoid pore-plate length and widthSAL, SAW suboral avicularian length and widthSD strut diameterSL, SW spine length and widthSPL, SPW strut pore-plate length and width# number of measurements.
ZÁGORŠEK ET AL.100 · Zootaxa 3838 (1) © 2014 Magnolia Press

Systematic account
The terminology follows that of Hayward and Ryland (1974). Bryozoan classification is based on that in Bock & Gordon (2013).
Superfamily Lepralielloidea Vigneaux, 1949
Family Jaculinidae Zabala, 1986
Genus Pirabasoporella gen. nov.
Type species. Pirabasoporella atalaiaensis n. sp.
Other species included. Pirabasoporella baitoae n. sp. and Pirabasoporella chipolae n. sp.
Etymology. Alluding to the type formation (Pirabas Formation) of Miocene strata in Brazil (Maury 1925).Diagnosis. Colony rigid, dichotomously branching, reticulate, subparallel neighbouring branches connected
by tubular horizontal struts; branches with autozooids arranged in two alternating longitudinal series and opening on one side only, colony attached by rhizoids. Autozooids with a pseudoporous, mixed umbonuloid and lepralioid frontal shield, orifice with proximal pseudosinus, condyles indistinct if present, oral spines present or absent. Struts formed by hollow kenozooids budded from and communicating with large pore plates at the distolateral and proximolateral walls of two zooids on either side. Adventitious avicularia of two types: suboral (always present) and frontal or abfrontal (occasionally present). Ovicell terminal, hyperstomial with uncalcified ectooecium and a pitted endooecial surface, not closed by the operculum in life. The dorsal (abfrontal) side of each zooid with a large circular foramen that is sealed by a multiporous pore plate and from which the rhizoid is produced.
Remarks. Owing to its peculiar reticulate colony growth, with kenozooidal struts connecting neighbouring branches, and because of strong similarities in orifice and ovicell shape as well as its mode of attachment via abfrontal rhizoids, Pirabasoporella gen. nov. is classified in the family Jaculinidae Zabala, 1986. This poorly known family, which is presently being revised by Rosso and Berning (in prep.), today occurs in the bathyal eastern Atlantic and Mediterranean Sea. Recent species assigned to the eponymous genus Jaculina Jullien in Jullien and Calvet, 1903 include the genotype J. blanchardi Jullien in Jullien and Calvet, 1903 as well as J. parallelata
(Waters, 1895) and J. tessellata Hayward, 1979. However, the Mediterranean fossil species Vibraculina conti
Neviani, 1895, which is the genotype of Vibraculina Neviani, 1895 and which is currently provisionally classified in the family Vicidae Gordon, 1988 (D.P. Gordon, pers. comm. 2013), can probably be regarded as congeneric. Vibraculina is thus likely to be a senior synonym of Jaculina, and Pirabasoporella would be the only other genus in the Jaculinidae.
Besides the kenozooidal struts that connect neighbouring branches, which are composed of one or two series of autozooids, species hitherto classified with the Jaculinidae are characterized by hyperstomial ovicells with a smooth or deeply pitted surface, by a sinusoidal orifice with or without oral spines, by the presence of adventitious avicularia, by abfrontal pore plates from which rhizoids are produced, and by frontal shields that are imperforate except for a single marginal row of pores. Everything else being principally the same, the major morphological difference justifying the introduction of the new jaculinid genus Pirabasoporella is the pseudoporous frontal shield in these western Atlantic species (see Discussion). Moreover, the mostly oval abfrontal rhizoidal pore plates, as well as their communication pores, are distinctly larger and situated towards the zooid centre, whereas those in the eastern Atlantic/Mediterranean taxa are comparatively small, round, and positioned directly at the distal zooidal margin. Adventitious (spatulate) avicularia on the frontal and abfrontal zooidal surface, in addition to the small suboral one, are also known only in Pirabasoporella.
The Jaculinidae has been classified in the lepraliomorph superfamily Schizoporelloidea Jullien, 1883 (D.P. Gordon, pers. comm. 2013). The discovery of an umbonuloid component in the frontal shield in Pirabasoporella
(Fig. 4F, G), and the fact that the jaculinid ovicell is not of the schizoporelloid type, suggests placing the family among umbonulomorph taxa. For lack of a better alternative, we here classify the Jaculinidae within superfamily Lepralielloidea. The jaculinid ooecium consists of a mostly membranous ectooecium, only the most basal part of which is calcified, and an entirely calcified and imperforate endooecium. The endooecial surface is smooth
Zootaxa 3838 (1) © 2014 Magnolia Press · 101A NEW GENUS OF JACULINIDAE FROM MIOCENE OF BRAZIL

(Jaculina blanchardi), or more or less deeply but superficially pitted (Jaculina parallelata, Pirabasoporella
atalaiaensis n. sp.). A similar type of ovicell is found in the Petraliidae Levinsen, 1909 and Petraliellidae Harmer, 1957, which are also grouped in the Schizoporelloidea at present. Moreover, orificial structure, including the presence of a suboral avicularium, and the formation of reticulate colonies in some petraliid species (see below) argue for a closer relationship among these taxa. However, it is unknown to us whether the Petraliidae and Petraliellidae have a lepralioid or (partly) umbonuloid frontal shield. A thorough phylogenetic analysis will therefore be dealt with in a future revision of the Jaculinidae (Rosso & Berning, in prep.).
Similarities between the reticulate colonies of jaculinid species and those of other bryozoan taxa are only superficial and not indicative of any phylogenetic relationship. Fenestrate, or reteporiform, colonial growth habits have repeatedly evolved in a number of unrelated taxa (McKinney & Jackson 1989). Although differing significantly in origin and structure, kenozooidal struts also characterize the well-known Paleozoic order Fenestrata as well as some Recent cyclostome bryozoan genera such as Fenestulipora Taylor & Gordon, 1997. In contrast, the connections between branches in all modern Cheilostomata with reteporiform colonies (termed trabeculae) are composed of autozooids. Reticulate colonies sensu lato appear in distantly related cheilostome genera such as, among others, Retiflustra Levinsen, 1909 (Flustridae), Smittipora Jullien, 1882 (Onychocellidae), Petralia
MacGillivray, 1869 (Petraliidae), Galeopsis Jullien in Jullien and Calvet 1903 (Celleporidae), Reteporella Busk, 1884 (Phidoloporidae), or Julianca Gordon, 1989 (incertae sedis).
Apart from species of Retiflustra, the colony bases in all other above-mentioned taxa are attached to more or less hard, stable substrata, and are heavily calcified in order to provide maximum resistance to wave action for these erect, rigid colonies. As a colony base or excessive ontogenetic frontal thickening of proximal branches have never been observed in any jaculinid species, it is likely that early astogeny does not involve fixation to a stable substratum. Instead, in Recent species the cuticular rhizoids, which derive from the large abfrontal pore plates, have a relatively large diameter and very thin walls, and are presumably the only means of attachment on soft substrata in Jaculinidae (see Discussion). As a consequence of these differences in habitat and attachment mode, we here refrain from classifying these rooted reticulate colonies in the Jaculinidae as ‘fenestrate’ or ‘reteporiform’.
Pirabasoporella atalaiaensis n. sp.
(Figs 3–4)
Etymology. Alluding to the type locality at Atalaia Beach (Salinopolis city, Pará State).Material examined. Holotype: MG-UFPA 6200-I, a colony fragment from the type locality at Atalaia Beach
(Salinopolis city, Pará State); type stratum, Pirabas Formation, Miocene. Paratype: 10 separate colony fragments labelled as MG-UFPA 6201-I to 6211-I. Additional material: More than 100 colony fragments, MG-UFPA 6212-I to 6300-I from sections Atalaia, B-17 Capanema and Aricuru.
Diagnosis. Pirabasoporella with relatively large zooids and few pores in the frontal shield. Oral spines absent. Adventitious avicularia of two types: a small suboral one associated with an umbo, and a larger one occasionally developing on the frontal shield during ontogeny. Ovicell terminal, hyperstomial with uncalcified ectooecium and a pitted endooecial surface, not closed by the operculum. Abfrontal rhizoidal foramen relatively small and proximally situated.
Description. Colony rigid, reticulate with bifurcating branches connected by kenozooidal struts at regular intervals, branches growing more or less parallel to each other (Figs. 3A-C). The struts usually developing at every second autozooid, jointly budded laterally from the distolateral and proximolateral corners of two zooids (Fig. 3B), slightly curved or straight, thick but hollow with microstructures indicating the same origin as autozooidal skeleton, surface smooth or with faint ridges and a row of slit-like areolar pores near each branch contact (Fig. 3D), struts delimiting elongate rounded-rectangular to lozenge-shaped fenestrules indented by slightly bulging peristomial rim of the interjacent zooids.
Autozooids rectangular but slightly curved, arranged in two alternating longitudinal series initially separated by thin raised ridges that gradually thicken during ontogeny (Fig. 3F), frontal shield mixed umbonuloid and lepralioid, the short umbonuloid part restricted to the distal shield (Fig. 4F, G), the interior surface smooth and often with concentric growth lines, the lepralioid frontal shield perforated by 15–25 pores, some pores not penetrating the shield entirely (Fig. 4 E, F). Primary orifice immersed, ill-defined, secondary aperture not well preserved, relatively large, suborbicular with a variably broad U-shaped pseudosinus (Fig. 3E), surrounded by a short, wide peristome that becomes thickened during ontogeny, proximolaterally carrying two protuberances, the prominent one towards the neighbouring zooid produced by a suboral avicularium (Fig. 3D, E).
ZÁGORŠEK ET AL.102 · Zootaxa 3838 (1) © 2014 Magnolia Press
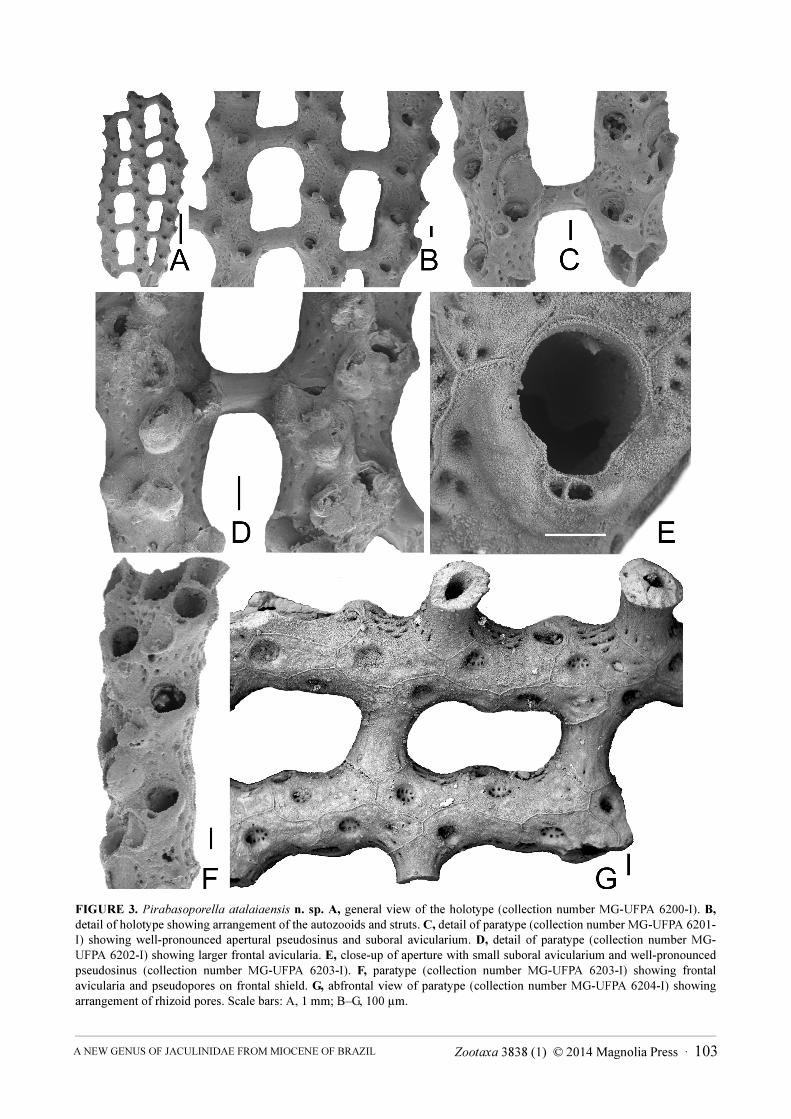
FIGURE 3. Pirabasoporella atalaiaensis n. sp. A, general view of the holotype (collection number MG-UFPA 6200-I). B,
detail of holotype showing arrangement of the autozooids and struts. C, detail of paratype (collection number MG-UFPA 6201-I) showing well-pronounced apertural pseudosinus and suboral avicularium. D, detail of paratype (collection number MG-UFPA 6202-I) showing larger frontal avicularia. E, close-up of aperture with small suboral avicularium and well-pronounced pseudosinus (collection number MG-UFPA 6203-I). F, paratype (collection number MG-UFPA 6203-I) showing frontal avicularia and pseudopores on frontal shield. G, abfrontal view of paratype (collection number MG-UFPA 6204-I) showing arrangement of rhizoid pores. Scale bars: A, 1 mm; B–G, 100 µm.
Zootaxa 3838 (1) © 2014 Magnolia Press · 103A NEW GENUS OF JACULINIDAE FROM MIOCENE OF BRAZIL

FIGURE 4. Pirabasoporella atalaiaensis n. sp. A, two well-developed ovicells facing the same fenestrule (collection number MG-UFPA 6205-I). B, four ovicells, showing the thickened imperforate lip around the ooecial opening (collection number MG-UFPA 6206-I). C, close-up of ovicell showing its shape and contact with the distal autozooid (collection number MG-UFPA 6207-I). D, close-up of ovicell showing opening and contact with the distal autozooid (collection number same as A). E,
interior view showing communication pores of rhizoidal pore plates (collection number MG-UFPA 6208-I). F, interior view showing orifices and pores in proximal frontal shield (collection number MG-UFPA 6209-I). G, close-up of the interior of an autozooid showing the umbonuloid part of the frontal shield arrow (collection number same as F). Scale bars all 100 µm.
Avicularia polymorphic, of two different types: a single, small, suboral avicularium on each autozooid (Fig. 3E), cystid slightly raised, situated proximolateral to aperture, rostrum subtriangular, directing laterally to proximolaterally, pivotal bar complete. Large adventitious frontal avicularia forming anywhere on the frontal shield in elder zooids (Fig. 3D); rostrum elongate-triangular, directed proximolaterally.
Ovicells large, terminal, globular but flattened frontally, hyperstomial, recumbent on proximal part of distal
ZÁGORŠEK ET AL.104 · Zootaxa 3838 (1) © 2014 Magnolia Press

zooid (Fig. 4A–D); ectooecium uncalcified, endooecium densely sculptured with numerous tiny pits or pores apart from a broad, smooth, prominent rim surrounding ovicell opening that arches above aperture (Fig. 4D), not closed by the operculum of the maternal zooid in life (Fig. 4B).
Abfrontal side of colony smooth, zooidal borders marked by thin raised ridges (Fig. 3G), each zooid with large, deep, round to elongate rhizoid pore, occasionally surrounded by a thick prominent rim during ontogeny, sealed at its base by a multiporous pore plate with the pores situated along the margin (Fig. 4E). An additional pore of the same type present at distal edge of lateral walls of zooids that do not produce a kenozooidal strut.
Ancestrula not observed.
TABLE 1. Measurements for Pirabasoporella atalaiaensis n. sp.
Remarks. Although more than 100 colony fragments of Pirabasoporella atalaiaensis n. sp. were found, all of them are very small, which complicates assessing the presence and extent of morphological change during ontogeny within a colony, and distinguishing it from interspecific differences. The fragments do vary to quite an extent with respect to strut thickness, presence/absence of large frontal avicularia, and size of the frontal shield pores, for example (compare Figs 3B, 3D, 4A). Morphometric analyses show overlapping ranges of strut thickness in the various fragments, with thin struts appearing in colonies that can be regarded as ontogenetically young colonies (Fig. 4A), and thick struts in older colonies (Fig. 3B). Large frontal avicularia were exclusively found in colonies that were composed of older or even ephebic (gerontic) zooids, some orifices of which were sealed off by frontal calcification (Fig. 3D, zooid at lower left). Thus, we consider intercolonial morphological variability to be the result of ontogenetic change, not of interspecific differences. Frontal thickening of zooids and an increase in strut diameter is likely to be explained by the necessity to reinforce an expanding colony.
Pirabasoporella baitoae n. sp.
(Figure 5)
Etymology. Alluding to the Early Miocene type Baitoa Formation in the Dominican Republic. Material examined. Holotype: USNM 595120, the colony fragment depicted in Fig. 5A, B, from the type
locality at Rio Yaque del Norte; type stratum Baitoa Formation, Early Miocene, (sensu Saunders et al. 1986), Dominican Republic. Paratypes: USNM 595121 to 595125, 5 separate colony fragments. Additional material: More than 100 colony fragments in USNM, numbers NMB 17285–17289 and NMB 16935, NMB 17184 and Olsson 179.
Diagnosis. Pirabasoporella with relatively few frontal-shield pores that become progressively closed during ontogeny, surface often nodular. Apertural pseudosinus relatively wide. Abfrontal rhizoidal pores large and encircled by an elevated rim, strut pores very large. Adventitious avicularia present on abfrontal side of some zooids; apart from the suboral avicularium additional frontal avicularia are lacking. Ovicells unknown.
Description. Colony rigid, reticulate, with bifurcating branches connected by kenozooidal struts, branches growing more or less parallel to each other (Fig. 5A, C); struts irregularly developed, jointly budded laterally from strut pore-plate, rarely one strut budded from two adjacent plates (Fig. 5G), slightly curved or straight, surface smooth or with faint ridges and a row of slit-like areolar pores near each branch contact. No fenestrules preserved.
Autozooids elongate-pentagonal, widest at about mid-distance, arranged in two alternating longitudinal series, separated by thin raised ridges that are especially prominent on the abfrontal side, the lepralioid part of the frontal shield perforated by up to 40 pores, many of which are closed during ontogeny while the lateral areolar pores remain open, surface occasionally nodular; umbonuloid part confined to distalmost frontal shield. Interzooidal communication via multiporous pore plates. Primary orifice immersed and ill-defined, with inconspicuous condyles, secondary orifice relatively large (Fig. 5B), suborbicular, about as wide as long, with a broadly U-shaped pseudosinus comprising 1/5 of total orifice length, formed by two extremely short pointed lateral denticles.
BW SD FL FW AL AW AD OL OW
mean 470 176 870 462 577 223 143 266 255
min 411 100 649 250 543 177 120 228 216
max 516 300 1021 638 631 24 163 286 271
# 40 55 17 17 20 20 20 6 6
Zootaxa 3838 (1) © 2014 Magnolia Press · 105A NEW GENUS OF JACULINIDAE FROM MIOCENE OF BRAZIL

FIGURE 5. Pirabasoporella baitoae n. sp. A, holotype, general view (collection number USNM 595120). B, detail of aperture of holotype, with suboral avicularium. C, paratype (collection number USNM 595121) showing late ontogenetic stage with several closed frontal pores. D, paratype (collection number USNM 595122) with partly preserved struts. E, strut initiated from two different pore plates (i.e. zooids) on one side of branch while fusing towards, and connecting with, a single pore plate on the opposite branch (collection number USNM 595123). F, paratype (collection number USNM 595124) showing strut-producing pore plates (right margin), rhizoid-producing pore plates (left margin and central part), and small oval avicularia in the bifurcation and on zooids in the branch centre (arrow). G, lateral view of paratype (collection number USNM 595125) showing three round, strut-producing pore plates (black arrows on two of them), two oval rhizoid pore plates (centre), and two small avicularia (white arrows). Scale bars all 100 µm.
ZÁGORŠEK ET AL.106 · Zootaxa 3838 (1) © 2014 Magnolia Press

Two types of avicularia present: each autozooid on frontal side with a single, small, suboral avicularium situated directly proximolateral to pseudosinus (Fig. 5B), oblique to frontal plane, rising distally, rostrum semielliptical, directed laterally towards median line, cystid raised distally to produce a suboral umbo, crossbar complete, without columella. Some zooids have an additional, slightly larger (see Table 2 for measurements) adventitious avicularium on abfrontal side, situated proximal to rhizoidal pore on a somewhat elevated cystid, elongate- oval with complete crossbar (Fig. 5F, G)
Abfrontal side of colony relatively smooth to distinctly nodular, zooidal borders marked by prominent ridges with a deep central suture, each zooid with an extremely large oval to circular rhizoid pore (up to 268 µm diameter) positioned relatively distal, surrounded by a thick prominent rim and sealed at its base by a multiporous pore plate with the large pores situated along the margin (Fig. 5F). Rhizoid pores of a similar type, but prominently oval (Fig 5G), present at distal edge of lateral walls of zooids that do not produce a kenozooidal strut, the pores in these usually smaller, not limited to pore-plate margin.
Ovicells and ancestrula not observed.
TABLE 2. Measurements for Pirabasoporella baitoae n. sp.
Remarks. Pirabasoporella baitoae n. sp. differs from P. atalaiaensis n. sp. and P. chipolae n. sp. in having generally fewer pores in the frontal shield (which occasionally displays a nodular surface), a broader apertural pseudosinus and the occasional presence of abfrontal avicularia while large frontal avicularia are absent. Further differences from P. atalaiaensis are the shorter but broader autozooids, and distinctly larger abfrontal rhizoidal pore plates that are circled by an elevated rim. In contrast to P. chipolae, P. baitoae does not have oral spines.
Among the approximately 100 colony fragments studied, none contained ovicells or indications of their presence. Insofar as P. atalaiaensis, like all other jaculinid species, produces hyperstomial ovicells, we do not interpret their lack in P. baitoae as an indication of internal brooding but perhaps reflecting a response or adaptation to environmental conditions, in which asexual colony growth may prevail over sexual reproduction. A similar strategy is found in Metrarabdotos, for example, in which several hundred autozooids may be formed for every gonozooid (Cheetham et al. 2001). However, we have no evidence (e.g. repair of broken zooids) that propagation in P. baitoae took place predominantly via fragmentation of colonies. On the other hand, neither ancestrulae nor early astogenetic colonies have ever been reported in any living or fossil jaculinid.
In one instance (Fig. 5E) a connecting strut was observed that originated from two different pore plates (i.e. zooids) on one side of the branch while fusing towards, and connecting to, a single pore plate on the opposite branch.
Pirabasoporella chipolae n. sp.
(Figure 6)
Etymology. Alluding to the type Early Miocene Chipola Formation in Florida.Material examined. Holotype: USNM 595126, the colony fragment depicted in Fig. 6A, from the type
locality at Farley Creek, Florida; type stratum, Chipola Formation, Early Miocene (sensu Saunders et al. 1986). Paratypes: USNM 595127 to 595131, 5 separate colony fragments. Additional studied material: More than 20 colony fragments in USNM, collection numbers TU LOC. 824, Scolaro collection; plus three fragments on a slide, NHMUK 1968.1.6.7, TU loc.825, leg. R.J. Scolaro.
Diagnosis. Pirabasoporella with numerous frontal-shield pores, a pair of oral spines, large spatulate avicularia occurring on lateral side of some later ontogenetic zooids, and deeply immersed rhizoidal as well as strut pore plates with numerous small communication pores.
BW SD AL AW AD SPL SDW PRL RPW AAL SAL
mean 458 183 500 236 151 245 233 186 97 93 57
min 403 153 443 200 142 181 179 128 85 64 44
max 514 202 543 28 158 302 268 219 113 113 81
# 37 18 32 32 64 12 12 24 24 6 5
Zootaxa 3838 (1) © 2014 Magnolia Press · 107A NEW GENUS OF JACULINIDAE FROM MIOCENE OF BRAZIL

FIGURE 6. Pirabasoporella chipolae n. sp. A, holotype (collection number USNM 595126) showing a triangular fenestrule above a branch bifurcation. B, detail of aperture of paratype (collection number USNM 595127) showing oral spines and the characteristic pseudosinus formed by two centrally directing lateral denticles. C, paratype (collection number USNM 595127) showing irregular growth with small fenestrules. D, paratype (collection number USNM 595128) showing well-calcified frontal shield with suboral avicularia. E, lateral view of paratype (collection number USNM 595129) showing strut-producing pore plates with a large spatulate avicularium (arrow). F, Abfrontal side of paratype (collection number USNM 595130) with large, deep, round to oval rhizoid-producing pores of variable size. G, abfrontal view of paratype (collection number USNM 595131) showing rhizoid-producing pores and two struts initiated from two different pore plates fusing with each other. Scale bars all 100 µm.
Description. Colony rigid, ramose, reticulate with bifurcating branches connected by kenozooidal struts at irregular intervals (Fig 6A), branches growing more or less parallel to each other. Struts usually developing chaotically, jointly budded laterally from the strut pore plate, slightly curved or straight, surface smooth or with faint ridges and a row of slit-like areolar pores near each branch contact, struts delimiting rounded-rectangular (Fig. 6A) to irregularly oval fenestrules (Fig. 6C).
Autozooids subrectangular, arranged in two alternating longitudinal series, not separated by thin raised ridges, but with prominent raised ridges on abfrontal side. Lepralioid part of frontal shield perforated by up to 30 pseudopores, many of which are closed during ontogeny. Primary orifice immersed and ill-defined, condyles
ZÁGORŠEK ET AL.108 · Zootaxa 3838 (1) © 2014 Magnolia Press

indistinct if present; secondary orifice relatively large, oval, longer than wide, with a broadly oval, wide pseudosinus comprising ¼ total orifice length, formed by two short, acute lateral denticles directed medially, the round or oval bases of two large spines situated directly at each lateral orifice margin (Fig. 6B).
Avicularia polymorphic: a single, small, suboral avicularium on each autozooid, situated directly proximal to pseudosinus cystid raised distally, oblique to frontal plane, rostrum semielliptical, directed laterally, crossbar complete, without columella; in ontogenetically older zooids a larger spatulate avicularium (Fig. 6E) rarely develops proximolateral to orifice towards branch axis, its rostrum curved and directed proximolaterally, distally entirely filled by a flat smooth shelf, crossbar not observed.
Abfrontal side of colony relatively smooth (Fig. 6E, F), zooidal borders marked by prominent ridges with deep median suture, each zooid with large, deep, round to oval rhizoid pore of very variable size (36–380 µm diameter; see Fig. 6F) positioned close to distal margin, usually surrounded by thick prominent rim during ontogeny, sealed at its base by a multiporous pore plate with the pores situated along the margin. An additional pore plate of a similar type present at distal edge of lateral walls of zooids that do not produce a kenozooidal strut.
Ovicells and ancestrula not observed.
TABLE 3. Measurements for Pirabasoporella chipolae n. sp.
Remarks. Pirabasoporella chipolae n. sp. differs from P. atalaiaensis n. sp. and P. baitoae n. sp. in having oral spines, occasional spatulate frontal avicularia and deeply immersed rhizoidal pore plates with relatively small communication pores. While oral spines are lacking from P. atalaiaensis and P. baitoae and most other jaculinid species, they are present in a few Jaculina species, e.g. J. tessellata, thus their presence in P. chipolae is not extremely unusual.
Preservation of some of the specimens is excellent, and the secondary orifice typical of jaculinids is most apparent in this species (e.g. Fig. 6B). The proximal pseudosinus is formed by two distinct lateral denticles (sensu
Berning et al. 2014) directed medially.As in P. baitoae, a connecting strut was observed in P. chipolae that originated from two different pore plates
on one side of the branch while fusing towards the opposite branch (Fig. 6G). Also, ovicells were not found in P.
chipolae. Few specimens were studied, however, so their absence is not surprising.
Discussion
The main difference between fossil western Atlantic Pirabasoporella and eastern Atlantic and Mediterranean Jaculinidae is the perforate frontal shield in the former. Recent species have only a single row of lateral areolar pores (BB, pers. observ.), whereas, in Pirabasoporella, at least the proximomedian part of the frontal shield is perforated and the latero-abfrontal areas contain more pores. Porous and imperforate frontal shields are not unusual in sister genera [e.g. Pourtalesella/Buffonellaria (cf. Berning & Kuklinski 2008) or Therenia/Herentia (cf. Berning et al. 2008)], while, according to Gordon (2000), the porous frontal shield is considered to be the derived character. Early Miocene Pirabasoporella with its perforate frontal shield clearly predates the Pleistocene to Recent eastern Atlantic/Mediterranean clade, and may counter this hypothesis. However, most Jaculinidae seem to occur at bathyal depths (but see below), the sediments of which have only a sparse fossil record. It is thus likely that the present temporal sequence of occurrence is caused by a sampling artefact. Moreover, the presence of jaculinid clades on both sides of the Atlantic argues for a considerable history of the group. Their adaptation to soft sediments, and their reproduction via short-lived trochophore larvae, makes it unlikely that species have crossed the ocean by rafting on floating objects or via their larval stage, respectively. Dispersal via abyssal basins seems equally unlikely given the precipitation of calcitic skeletons in jaculinids. The most probable scenario is an early
BW SD FL FW AL AW AD SPL SDW RPL RPW AAL AAW SAL SAW SL SW
mean 543 212 641 339 530 257 178 388 353 117 75 289 81 67 41 49 27
min 423 196 610 248 491 197 151 351 324 54 36 281 78 52 34 34 22
max 594 22 673 438 566 299 210 401 380 206 103 296 85 78 52 60 33
# 17 8 3 3 24 24 44 9 9 10 10 2 2 13 13 10 10
Zootaxa 3838 (1) © 2014 Magnolia Press · 109A NEW GENUS OF JACULINIDAE FROM MIOCENE OF BRAZIL

Paleogene origin of the Jaculinidae when the continental shelves of the Atlantic were positioned considerably closer together. All fossil and Recent jaculinid species have been found in tropical to warm-temperate regions, while there are no records of Recent Jaculinidae from the western Atlantic to date.
The abfrontal rhizoidal pore plates and their communication pores in Pirabasoporella are distinctly larger than in the eastern Atlantic/Mediterranean taxa, suggesting that the rhizoids had a much greater diameter. Also, every zooid contains a pore plate, providing for a dense spacing of rhizoids. In Recent Jaculinidae, which occur at ~200–1250 m depth (J. blanchardi 230–980 m; J. dichotoma 1250 m; J. tessellata 460–1000 m), the relatively thin rhizoids are usually more widely spaced (BB, pers. observ.). The depositional environment of the respective formations in which the three Pirabasoporella species were found suggests that the water depth was comparatively shallow, perhaps inner-shelf (Távora & Fernandes 1994), and that stronger anchoring may have been needed to withstand higher current speeds. As no living jaculinid colonies have been observed in situ, the position of colonies relative to the seafloor is unclear and cannot be resolved with the fragmented material studied during this work.
Acknowledgements
We thank the Departamento Nacional de Produção Mineral (DNPM) for permission to sample the material in Brazil, and “Cimentos do Brasil S.A. - Cibrasa” for authorization to collect at the B-17 Mine. We are grateful to Ana Paula Correa and Claudio N. Lamarão (Instituto de Geociências, Laboratório de Microscopia Eletrônica de Varredura, UFPA) and Amanda G. Veiga (Museu Nacional, UFRJ) for SEM pictures, to ANP/Petrobrás for financing and developing the Centro de Microscopia Eletrônica de Varredura do Museu Nacional/UFRJ (ANPETRO 8808). Dennis Gordon (NIWA) is thanked for suggesting links between the Jaculinidae and Petraliidae/Petraliellidae. BB acknowledges support from the SYNTHESYS Project (http://www.synthesys.info/), which is financed by the European Community Research Infrastructure Action under the FP7 “Capacities” Program, and which allowed study of Jaculina-types at the Muséum National d'Histoire Naturelle, Paris (FR-TAF-1902), and the Natural History Museum, London (GB-TAF-3347).
Our thanks go also to Dr. JoAnn Sanner from Smithsonian Institution, Department of Paleobiology who provided us the specimens from Dominican Republic and Florida. The research has been supported by the Grant agency of the Czech Republic, project number 205/09/0103: "Shallow water ecosystems from the Middle Miocene of the Central Paratethys: Succession and interactions between inorganic and organic elements of the ecosystems."
References
Arnold, P. (1987) Observations on living colonies of Retiflustra spp. (Cheilostomata: Anasca) from the central Queensland shelf, Australia. Cahiers de Biologie Marine, 28, 147–157.
Barbosa, M.M. (1971) Cupuladria canariensis (Busk) no Brasil. Boletim do Museu Nacional, Rio de Janeiro, 35, 1–6.Berning, B. (2013) New and little-known Cheilostomata (Bryozoa, Gymnolaemata) from the NE Atlantic. European Journal of
Taxonomy, 44, 1–25. http://dx.doi.org/10.5852/ejt.2013.44
Berning, B. & Kuklinski, P. (2008) North-east Atlantic and Mediterranean species of the genus Buffonellaria (Bryozoa, Cheilostomata): implications for biodiversity and biogeography. Zoological Journal of the Linnean Society, 152, 537–566. http://dx.doi.org/10.1111/j.1096-3642.2007.00379.x
Berning, B., Tilbrook, K.J. & Rosso, A. (2008) Revision of the north-eastern Atlantic and Mediterranean species of the genera Herentia and Therenia (Bryozoa: Cheilostomata). Journal of Natural History, 42, 1509–1547.
Berning, B., Tilbrook, K.J. & Ostrovsky, A.N. (2014) What, if anything, is a lyrula? In: Rosso, A., Wyse Jackson, P.N. & Porter, J. (Eds.), Bryozoan Studies 2013. Studi Trentini di Scienze Naturali, Acta Geologica, 94, 21–28.
Bock, P.E. & Gordon, D.P. (2013) Phylum Bryozoa Ehrenberg, 1831. In: Zhang, Z.-Q. (Ed.), Animal Biodiversity: An outline of higher-level classification and survey of taxonomic richness (Addenda 2013). Zootaxa, 3703 (1), 67–74. http://dx.doi.org/10.11646/zootaxa.3703.1.14
Bryant, J.D., Macfadden, B.J. & Mueller, P.A. (1992) Improved chronologic resolution of the Hawthorn and Alum Bluff Groups in northern Florida: Implications for Miocene chronostratigraphy. Geological Society of America Bulletin, 104, 208–218. http://dx.doi.org/10.1130/0016-7606(1992)104<0208:icroth>2.3.co;2
ZÁGORŠEK ET AL.110 · Zootaxa 3838 (1) © 2014 Magnolia Press

Busk, G. (1884) Report on the Polyzoa. Part I. The Cheilostomata. Report of the Scientific Results of the Voyage of H.M.S. Challenger during the years 1873–76, 10, 1–216.
Cheetham, A.H., Jackson, J.B.C. & Sanner, J. (2001) Evolutionary significance of sexual and asexual modes of propagation in Neogene species of the bryozoan Metrarabdotos in tropical America. Journal of Paleontology, 75, 564–577. http://dx.doi.org/10.1666/0022-3360(2001)075<0564:esosaa>2.0.co;2
Cook, P.L. (1981) The potential of minute bryozoan colonies in the analysis of deep sea sediments. Cahiers de Biologie
Marine, 22, 89–106.Cook, P.L. & Chimonides, P.J. (1981) Morphology and systematics of some rooted cheilostome Bryozoa. Journal of Natural
History, 15, 97–134. http://dx.doi.org/10.1080/00222938100770081
Fernandes, J.M.G. (1988) Bioestratigrafia da Formação Pirabas, Estado do Pará. In: Congresso Brasileiro De Geologia, 35, Rio
de Janeiro, Anais. Vol. 6. Sociedade Brasileira de Geologia, Rio de Janeiro, pp. 2376–2382. Fernandes, J.M.G. & Távora, V.A. (1990) Estudo dos foraminíferos da Formação Pirabas procedentes do furo CB-UFPA-P1
(85), município de Capanema, Estado do Pará. In: Congresso Brasileiro De Geologia, 33, Natal, Anais. Sociedade Brasileira de Geologia Press, Natal, pp. 470–475.
Ferreira, C.S. (1967) Contribuição à Paleontologia do Estado do Pará. O gênero Orthaulax Gabb, 1872 na Formação Pirabas.
In: Simpósio Sobre A Biota Amazônica. Atas Press, Belém, pp. 169–181. Ferreira, C.S., Vicalvi, M.A. & Macedo, A.C.M. (1981) Nota sobre a sequência sedimentar ao sul do rio Guamá, Estado do
Pará. Evidências do Oligo-Mioceno marinho, através dos resultados preliminares da sondagem feita em Vila Mãe do Rio (“48”), BR-010, Município de Irituia. Anais da Academia Brasileira de Ciências, Rio de Janeiro, 53, 208–209.
Góes, A.M., Rossetti, D.F., Nogueira, A.C. & Toledo, P.M. (1990) Modelo deposicional preliminar da Formação Pirabas no nordeste do Pará. Boletim do Museu Paraense Emílio Goeldi, Série Ciências da Terra, 2, 3–15.
Gordon, D.P. (1988) The bryozoan families Sclerodomidae, Bifaxariidae, and Urceoliporidae and a novel type of frontal wall. New Zealand Journal of Zoology, 15, 249–290. http://dx.doi.org/10.1080/03014223.1988.10422619
Gordon, D.P. (1989) The marine fauna of New Zealand: Bryozoa: Gymnolaemata (Cheilostomatida Ascophorina) from the western South Island continental shelf and slope. New Zealand Oceanographic Institute Memoirs, 97, 1–158.
Gordon, D.P. (2000) Towards a phylogeny of cheilostomes—morphological models of frontal wall/shield evolution. In: Herrera Cubilla, A. & Jackson, J.B.C. (Eds.), Proceedings of the 11th International Bryozoology Association Conference, 1998,
Panama City. Smithsonian Tropical Research Institute, Balboa, pp. 17–37.Harmer, S.F. (1957) The Polyzoa of the Siboga Expedition, Part IV. Cheilostomata, Ascophora. II. Siboga Expeditie, 28d,
641–1147.Hayward, P.J. (1979) Deep water Bryozoa from the coasts of Spain and Portugal. Cahiers de Biologie Marine, 20, 59–75.Hayward, P.J. & Ryland, J.S. (1979) British Ascophoran Bryozoans. Synopses of the British Fauna, n.s., 14, 1–306.Hirose, M. (2011) Orientation and righting behavior of the sand-dwelling bryozoan Conescharellina catella. Invertebrate
Biology, 130, 282–290. http://dx.doi.org/10.1111/j.1744-7410.2011.00237.x
Jones, D.S., Mueller, P.A., Hodell, D.A. & Stanley, L.A. (1993) 87Sr/86Sr geochronology of Oligocene and Miocene marine strata in Florida. Geological Survey Special Publication, 37, 15–26.
Jullien, J. (1882) Note sur une nouvelle division des Bryozoaires Cheilostomiens. Bulletin de la Société Zoologique de France,6, 271–285.
Jullien, J. & Calvet, L. (1903) Bryozoaires provenant des campagnes de l'Hirondelle (1886-1888). Résultats des Campagnes
Scientifiques du Prince de Monaco, 23, 1–188.Levinsen, G.M.R. (1909) Morphological and Systematic Studies on the Cheilostomatous Bryozoa. Nationale Forfatteres Forlag,
Copenhagen, 431 pp.Marcus, E. (1938) Briozoarios marinhos brasileiros II. Boletim da Faculdade de Filosofia, Ciências e Letras da Universidade
de São Paulo, Zoologia, 2, 1–196.Maury, C.J. (1925) Fósseis terciários do Brasil com descrição de novas formas cretáceas. Monografia do Serviço Geológico e
Mineralógico do Brasil, Rio de Janeiro, 4, 1–665.MacGillivray, P.H. (1869) Descriptions of some new genera and species of Australian Polyzoa; to which is added a list of
species found in Victoria. Transactions and Proceedings of the Royal Society of Victoria, 9, 126–148.McKinney, F.K. & Jackson, J.B.C. (1989) Bryozoan Evolution. Unwin Hyman, Boston, 238 pp.Neviani, A. (1895) Nuove genere e nuove specie di Briozoi fossili. Rivista Italiana di Paleontologia e Stratigrafia, 1, 82–84.Ramalho, L.V., Muricy, G. & Taylor, P.D. (2009) Cyclostomata (Bryozoa, Stenolaemata) from Rio de Janeiro State, Brazil.
Zootaxa, 2057, 32–52.Ramos, M.I.F., Távora, V.A., Pinheiro, M.P. & Baia, N.B. (2004) Microfósseis. In: Rossetti, D.F. & Góes, A.M. (Eds.), O
Neógeno da Amazônia Oriental. Editora do Museu Paraense Emílio Goeldi, Coleção Friederich Katzer, Belém, pp. 93–107.
Saunders, J.B., Jung, P. & Biju-Duval, B. (1986) Neogene palaeontology of the northern Dominican Republic, 1, Field surveys, lithology, environment, and age. Bulletins of American Paleontology, 89 (323), 1–79.
Scolaro, R.J. (1968) Paleoecologic interpretation of some Florida Miocene Bryozoa. Atti della Società italiana di scienze
Zootaxa 3838 (1) © 2014 Magnolia Press · 111A NEW GENUS OF JACULINIDAE FROM MIOCENE OF BRAZIL

naturali, e del Museo civico di storia naturale. Milano, 108, 174–177.Scott, T.M., Campbell, K.M., Rupert, F.R., Arthur, J.D., Missimer, T.M., Lloyd, J.M., Yon, W. & Duncan, J.G. (2001) Geologic
Map of the State of Florida. Florida Geological Survey Open File Report, 80, 1–99.Tavener-Smith, R. (1969) Skeletal structure and growth in the Fenestellidae (Bryozoa). Palaeontology, 12, 281–309. Tavener-Smith, R. (1971) Polypora stenostoma: A Carboniferous bryozoan with cheilostomatous features. Palaeontology, 14,
178–187. Távora, V.A. & Fernandes, J.M.G. (1994) Uma fáunula de briozoários da Formação Pirabas (Mioceno Inferior), Estado do Pará.
Acta Geológica Leopoldensia, 17 (39/1), 145–156. Távora, V.A. & Fernandes, J.M.G. (1999) Estudo de los foraminíferos de la Formación Pirabas (Mioceno Inferior), Estado do
Pará, Brasil y su correlación com faunas del Caribe. Revista Geológica de América Central, 22, 59–70. Távora, V.A., Imbeloni, E.F.F., Cacela, A.S.M. & Baia, N.B. (2004) Paleoinvertebrados. In: Rossetti, D.F. & Góes, A.M. (Eds.),
O Neógeno da Amazônia Oriental. Editora do Museu Paraense Emílio Goeldi, Coleção Friederich Katzer, Belém, pp. 111–131.
Távora, V.A., Silveira, E.S.F. & Milhomem Neto, J.M. (2009) Mina B-17, Capanema, PA, Expressivo registro de uma paleolaguna do Cenozóico brasileiro. In: Winge, M., Schobbenhaus, C., Souza, C.R.G, Fernandes, A.C.S., Berbert-Born, M., Queiroz, E.T. & Campos, D.A. (Eds.), Sítios Geológicos e Paleontológicos do Brasil. CPRM-SIGEP, Brasília, 2, 363–370.
Taylor, P.D. & Gordon, D.P. (1997) Fenestulipora, gen. nov., an unusual cyclostome bryozoan from New Zealand and Indonesia. Invertebrate Taxonomy, 11, 689–703. http://dx.doi.org/10.1071/it96015
Vigneaux, M. (1949) Révision des Bryozoaires Néogènes du bassin d'Aquitaine et essai de classification. Mémoires de la
Société Géologique de France, 28, 1–155.Waters, A.W. (1895) On Mediterranean and New Zealand Reteporae and fenestrate Bryozoa. Journal of the Linnean Society
(Zoology), 25, 255–271. http://dx.doi.org/10.1111/j.1096-3642.1895.tb00389.x
Woodring, W.P. (1966) The Panama land bridge as a sea barrier. American Philosophy Society Proceedings, Washington, 110, 425–433.
Woodring, W.P. (1974) Affinities of Miocene marine molluscan faunas on Pacific side of Central America. Publicaciones
Geológicas del Instituto Centroamericano de Investigación y Tecnología Industrial, 4, 179–188. Woodring, W.P. (1978) Distribution of Tertiary marine molluscan faunas in southern Central America. Instituto Geológico
Universidad Autonome, 101, 153–166.Zabala, M. (1986) Fauna dels Briozous dels Països Catalans. Institut d'Estudis Catalans, Arxius de la Secció de Ciéncies 84,
Barcelona, 836 pp. Zágoršek, K., Nehyba, S., Tomanová Petrová, P., Hladilová, Š., Bitner, M.A., Doláková, N., Hrabovský, J. & Jašková, V.
(2012) Local catastrophe caused by tephra input near Přemyslovice (Moravia, Czech Republic) during the Middle Miocene. Geological Quarterly, 56, 269–284. http://dx.doi.org/10.7306/gq.1021
ZÁGORŠEK ET AL.112 · Zootaxa 3838 (1) © 2014 Magnolia Press




















