
International Portfolio Diversification: A Malaysian Perspective

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofitpublishers, academic institutions, research libraries, and research funders in the common goal of maximizing access tocritical research.
Diversification of mammals from the Miocene of SpainAuthor(s): M. Soledad Domingo , Catherine Badgley , Beatriz Azanza , DanielDeMiguel , and M. Teresa AlberdiSource: Paleobiology, 40(2):196-220. 2014.Published By: The Paleontological SocietyDOI: http://dx.doi.org/10.1666/13043URL: http://www.bioone.org/doi/full/10.1666/13043
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated contentindicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.

Diversification of mammals from the Miocene of Spain
M. Soledad Domingo, Catherine Badgley, Beatriz Azanza, Daniel DeMiguel, and M. Teresa Alberdi
Abstract.—The mammalian fossil record of Spain is long and taxonomically well resolved, offering themost complete record of faunal change for the Neogene of Europe. We evaluated changes indiversification, composition, trophic structure, and size structure of large mammals over the middleand late Miocene with methods applied to this record for the first time, including ordination of fossillocalities to improve temporal resolution and estimation of confidence intervals on taxa temporalranges. By contrast, analysis within the traditional Mammal Neogene (MN) biochronology obscuresimportant aspects of diversification. We used inferred temporal ranges of species and evaluated percapita rates of origination, extinction, diversification, and turnover over 0.5-Myr time intervals.
Three periods of significant faunal change occurred between 12.0 and 5.5 Ma: (1) From 12.0 to 10.5Ma, elevated origination rates led to an increase in diversity without significant change in ecologicalstructure. Immigrants and geographic-range shifts of species to lower latitudes during an interval ofglobal cooling contributed to these faunal changes. (2) From 9.5 to 7.5 Ma, high extinction ratesfollowed by high origination rates coincided with significant changes in taxonomic composition andecological structure. These changes represent the Vallesian Crisis, with replacement of a fauna of forestaffinities (with frugivores and browsers) by a fauna of open woodlands (with grazers and mixedfeeders). (3) From 6.5 to 5.5 Ma, high extinction rates reduced diversity without substantial changes inecological structure, and large mammal faunas became highly endemic across the northernMediterranean region. This interval includes the Messinian Salinity Crisis, the desiccation of theMediterranean basin. Extinction may have been caused by geographic isolation and aridification, withevolution of endemic lineages giving rise to new species in the early Pliocene. These distinctmacroevolutionary patterns of faunal change correspond to different geographic scales of inferredclimatic and tectonic drivers.
M. Soledad Domingo. Museum of Paleontology, University of Michigan, Ann Arbor, Michigan 48109, U.S.A.E-mail: [email protected]. Present address: Departamento de Paleontologıa, Facultad de CC.Geologicas, Universidad Complutense de Madrid, Madrid 28040, Spain
Catherine Badgley. Department of Ecology and Evolutionary Biology, University of Michigan, Ann Arbor,Michigan 48109, U.S.A. E-mail: [email protected]
Beatriz Azanza. Departamento de Ciencias de la Tierra, Facultad de Ciencias, Instituto Universitario deInvestigacion en Ciencias Ambientales de Aragon, Universidad de Zaragoza, Zaragoza 50009, Spain. E-mail:[email protected]
Daniel DeMiguel. Institut Catala de Paleontologia Miquel Crusafont, Universitat Autonoma de Barcelona,08193 Cerdanyola del Valles, Barcelona, Spain. E-mail: [email protected]
M. Teresa Alberdi. Departamento de Paleobiologıa, Museo Nacional de Ciencias Naturales-CSIC, Madrid 28006,Spain. E-mail: [email protected]
Accepted: 14 November 2013Published online: 14 February 2014Supplemental material deposited at Dryad: doi 10.5061/dryad.4kp4r
Introduction
Long, dense mammalian fossil records areunique windows for investigating the dynam-ics of biotic assemblages in the past and forevaluating potential drivers of biotic change(e.g., Maas et al. 1995; Barry et al. 2002; vander Meulen et al. 2005; van Dam et al. 2006;Badgley et al. 2008; Finarelli and Badgley2010). For the European record, the Neogenebasins of Spain contain the highest density ofmammalian fossil sites in the continent(Agustı et al. 2001). The completeness of the
Neogene mammalian fossil record of the
Iberian Peninsula has been estimated as 77%
at the species level and 91% at the generic
level (Alba et al. 2001). The record of Neogene
mammals from Spain is not only rich and
continuous but also well resolved taxonomi-
cally, making it possible to obtain higher
taxonomic and temporal resolution of mam-
malian evolution through time than in other
areas of Europe. These qualities have motivat-
ed several studies of faunal dynamics focusing
on the Spanish record of large mammals (e.g.,
� 2014 The Paleontological Society. All rights reserved. 0094-8373/14/4002-0004/$1.00
Paleobiology, 40(2), 2014, pp. 196–220DOI: 10.1666/13043

Alberdi et al. 1997; Morales et al. 1999; Azanzaet al. 2000) and small mammals (e.g., Daams etal. 1999; van der Meulen et al. 2005; van Damet al. 2006; Casanovas-Vilar and Agustı 2007;Gomez-Cano et al. 2013), or both (e.g., Agustıet al. 1999a, 2003, 2006; van Dam et al. 2001;Hernandez-Fernandez et al. 2006; Casanovas-Vilar et al. 2010) over different Neogeneintervals.
For the European Neogene record, changesin mammalian faunas through time havetraditionally been analyzed by placing local-ities chronologically into Mammal Neogene(MN) units (e.g., Agustı et al. 2001; Fortelius etal. 2006; Costeur and Legendre 2008; Eronenet al. 2009, 2012; Casanovas-Vilar et al. 2010).The MN system is the most widely usedbiochronological framework for correlatingcontinental European fossil sites (Mein 1975);it relies primarily on faunal associations andfirst and last appearances of mammalianlineages. The MN system contains 17 units ofdifferent durations, some as long as 3.0 Myrand some as short as 0.5 Myr. The differentunit durations pose a problem for analyses ofdiversity because longer units are expected tocontain more species than shorter units.Furthermore, since the MN units are in partdefined by first and last appearances, origina-tion and extinction rates are expected to behigh in all of the MN units (Casanovas-Vilar etal. 2010).
Although some Iberian basins contain longrecords of Neogene mammalian fossil locali-ties (e.g., the Teruel, Calatayud-Daroca, andValles-Penedes Basins), a comprehensivescheme for the Spanish mammal record canbe achieved only by integrating biochrono-logical data from different basins into acommon framework. Establishing stratigraph-ic, and thus temporal, relationships amongsites from different Iberian basins is difficultbecause many fossil localities lack radiometricor paleomagnetic age estimates and becausefaunas of the interior basins exhibit somedifferences from those of the northeasternbasins. In the context of these uncertainties,the MN timescale has become the primarybiochronological system for the Spanish Neo-gene mammalian record. Nevertheless, cur-rent techniques based on the relative sequence
of lineage appearances permit ordination andage estimation for fossil localities with hightaxonomic resolution and lead to more con-strained estimates of faunal change throughtime. Maximum Likelihood Appearance EventOrdination (Alroy 2000) has proven capable ofordinating sequences of fossiliferous sites withinternal consistency. These methods have beensuccessfully applied to the Spanish Neogenerecord as an alternative to the MN system(e.g., Azanza et al. 2000; Hernandez Fernan-dez et al. 2004; Domingo et al. 2007; GomezCano et al. 2011).
Once fossil localities and lineages are wellresolved in age and sequence, an even morereliable view of the tempo and mode of faunalchange can be gained by estimating theuncertainty in the duration of individuallineages. The observed stratigraphic rangesof species typically underestimates their actualduration to an extent that depends on thecompleteness of the fossil record (Signor andLipps 1982). Various methods have beendeveloped to estimate the actual longevitiesof fossil taxa (Paul 1982; Koch 1987; Straussand Sadler 1989; Marshall 1990, 1994, 1997,2010 and references therein; Barry et al. 2002).To the best of our knowledge, they have notbeen used in analyses of the Spanish Neogenerecord of large mammals.
Our analysis of the mammalian recordcovers the interval from 12.0 to 5.5 Ma (latemiddle Miocene to latest Miocene, corre-sponding approximately to MN7/8 toMN13). By this time, global temperatureswere cooling as Antarctic ice sheets expanded,after the warm period known as the MiddleMiocene Climatic Optimum (Zachos et al.2001). The Mediterranean area also experi-enced more regional episodes, including theVallesian Crisis and the Messinian SalinityCrisis. The Vallesian Crisis refers to a set ofextinctions that affected mainly the mammalfaunas of Western Europe and coincided withincreasing seasonality of precipitation (and oftemperature in near-coastal basins from theeastern Iberian Peninsula) and a shift fromhumid, forest vegetation to drier woodland(Agustı and Moya-Sola 1990; Agustı et al.1999a, 2003; Agustı and Anton 2002; Casano-vas-Vilar et al. 2005). This change occurred
MIOCENE MAMMAL DIVERSIFICATION 197

between 9.72 and 9.23–9.02 Ma and wasassociated with the cooling phase of the Mi7oxygen isotope stage, dated at ca. 9.6 to 8.8 Ma(Agustı et al. 2003; van Dam et al. 2006). TheMessinian Salinity Crisis refers to the desicca-tion of the Mediterranean Sea and formationof thick evaporite deposits (Hsu et al. 1973).Northeastward movement of the African plateduring the late Cenozoic caused progressiveclosure of the Betic and Rifian marine corri-dors that connected the Mediterranean Seaand the Atlantic Ocean, leading to theisolation of the Mediterranean Basin and theeventual drying of a sea that already had anegative hydrologic budget (evaporation .
precipitation þ river discharge), as does themodern Mediterranean (Mariotti et al. 2002;Rouchy and Caruso 2006). Falling sea levelplayed a role as well (Krijgsman et al. 1999;Rouchy and Caruso 2006). The MessinianSalinity Crisis began at 5.96 Ma and completeisolation lasted from 5.59 Ma to 5.33 Ma(Krijgsman et al. 1999). Surprisingly, despitethe disappearance of a vast body of water, nodrastic changes in regional vegetation orclimatic conditions occurred on the IberianPeninsula, according to paleoenvironmentalstudies based on pollen records and stableisotopes of C and O from mammalian enamel(e.g., Fauquette et al. 2006; Jimenez-Moreno etal. 2010; Domingo et al. 2013).
Our goals in this study are to evaluatepatterns of change in the diversity, composi-tion, and ecological structure of mammalianfaunas in relation to the history of tectonics,climate, and vegetation of the Iberian regionduring the late middle Miocene to lateMiocene. We focus on the record of largemammals (.1 kg in estimated body weight).Large and small mammals have often beenanalyzed separately due to the differentrecovery techniques (surface collection andexcavation vs. screen washing, respectively)and because ecological relationships andgeographic ranges differ according to bodysize. Our approach utilizes ordination of fossillocalities into a common biochronologicalframework and estimation of confidenceintervals for the temporal duration of species.We analyze per capita rates of origination,extinction, diversification, and turnover, as
well as changes in faunal composition andtrophic and size structure for 0.5-Myr inter-vals. In addition, we compare large mammaldiversity based on inferred ranges withdiversity based on observed ranges binnedinto 0.5-Myr intervals and with diversityusing the MN system.
Materials and Methods
Fossil Data
We utilized a database of large mammalfossil sites from Spain spanning the latemiddle Miocene to the early Pleistocene, i.e.,from MN7/8 to MN17, approximately from12.5 to 2.5 Ma (updated from Domingo et al.2007). To construct the database, we compiledfaunal lists from the literature; we refinedtaxonomic discrepancies with advice fromspecialists in the mammalian groups ofinterest or the researchers working at specificfossil sites. Because these sources contain themost current information, we used the taxo-nomic lists in our database instead of those inthe New and Old Worlds (NOW) database(Fortelius 2012). The faunal database used inthis study is available upon request to thecorresponding author.
Although our analysis interval extends from12.0 to 5.5 Ma (from ca. MN7/8 to MN13), theinclusion of younger localities was necessaryto provide directionality for the temporalordination of fossil sites. For the ordinationmethod, we included radiometric or paleo-magnetic age estimates whenever they wereavailable for the localities; the time period inthis study was adjusted for this requirement.The fossil sites are located in 17 Cenozoicbasins in Spain (Fig. 1).
The database contains a total of 93 faunallists, 72 of them from the study interval.Under the term ‘‘large mammal,’’ we includedspecies in the orders Artiodactyla, Perissodac-tyla, Hyracoidea, Proboscidea, Primates, andCarnivora. The total number of species in ouranalysis interval is 155 (Table 1, Supplemen-tary Table 1). Taxa identified as cf. or aff. wereconsidered to belong to their nominal species.The species level of taxonomic resolution forlarge mammals reflects the long paleontolog-ical tradition in Spain and was our preferred
198 M. SOLEDAD DOMINGO ET AL.

taxonomic level of diversity analysis. Occur-rence data from localities lying outside theanalysis interval were used in order to avoidedge effects, i.e., overestimating the number oforiginations at the beginning and the numberof extinctions at the end of the interval.
Each species was assigned to a trophiccategory and a size category. We used dentalmicrowear and mesowear analyses (DeMiguel2009; DeMiguel et al. 2011), as well asadditional information from the NOW data-base (Fortelius 2012) to assign species to atrophic category. Trophic determinations werefurther refined by consulting the literature foreach of the major taxonomic groups and theadvice of specialists (L. Domingo personalcommunication 2011, 2012; J. Morales personalcommunication 2011). Species were assignedto one of seven trophic categories: frugivore,browser, mixed feeder, grazer, omnivore,carnivore, and invertebrate faunivore. TheNOW database and additional data compiledby us were used to assign each species to oneof seven body-size categories, following thesize classification of Hernandez Fernandez etal. (2006): (A) 0.1–1 kg, (B) 1–10 kg, (C) 10–45kg, (D) 45–90 kg, (E) 90–180 kg, (F) 180–360kg, and (G) . 360 kg.
Analytical Methods
Ordination and Age Estimates of Fossil Sites.—We used Maximum Likelihood AppearanceEvent Ordination (ML AEO) to assign ageestimates to the fossil sites (Alroy 2000).Species at localities from MN14 to MN17 wereused to polarize the sequence. For somebasins, stratigraphic relationships for some ofthe fossil sites are known and this informationwas included in the ML AEO analysis. Thismethod ordinates fossil sites by using nestedsubsets of appearances; to obtain a numericalage for each locality, the ordination must becalibrated using age estimates for somelocalities based on paleomagnetic or radio-metric methods. Fourteen localities includedin this study have been paleomagneticallydated (Supplementary Table 2; see Supple-mentary Text for an explanation of the ageestimate of the Venta del Moro locality). Weutilized the geomagnetic timescale of Grad-stein et al. (2004). In order to date the entiresequence of fossil sites, we performed aregression of the 14 independent ages on theircorresponding ordination position (Supple-mentary Table 3). Two paleomagneticallydated fossil sites, Librilla and El Rincon 1,were omitted from the regression analysis asoutliers because they decreased the quality ofthe fit (Domingo et al. 2007). A quadraticregression was preferred to a linear one,because a nested F-test showed that thequadratic variable (mean ordination position2)was significant in the regression model. Also,the standard error of the quadratic regressionwas lower than the standard error of the linearmodel (0.351 and 0.489 Myr, respectively). Theregression coefficients were significant at p ,
0.01 and the regression model had an R2 of
FIGURE 1. Cenozoic basins (in gray) from the IberianPeninsula. Basins containing fossil sites used in this study:1 ¼ Valles–Penedes Basin, 2 ¼ Alicante North PrebeticBasin, 3 ¼ Valencia Basin, 4 ¼ Cabriel Basin, 5 ¼ JucarBasin, 6¼ Tagus Basin, 7¼Guadix–Baza Basin, 8¼ TeruelBasin, 9 ¼ Calatayud-Daroca Basin, 10 ¼ Sarrion–MijaresBasins, 11¼Duero Basin, 12¼ Ebro Basin, 13¼CalatravaBasin, 14 ¼ Fortuna Basin, 15 ¼ Granada Basin, 16 ¼Cerdanya Basin, and 17¼ Seu d’Urgell Basin.
TABLE 1. Number of large-mammal species before andafter applying criteria for inclusion in the diversityanalyses. CI, confidence intervals; IR, inferred ranges.
Number
Species present in the study interval 155Species found in only one fossil site
(¼ singletons) 62Species with CI .1 Myr (excluded from IR
approach) 13Species with CI �1 Myr (retained in IR
approach) 80
MIOCENE MAMMAL DIVERSIFICATION 199

0.99 (Supplementary Table 3). The ML AEOprocedure reduced the database from 72 to 57localities because faunal lists that are subsetsof longer lists and those that had fewer thantwo taxa were considered non-informative forordination and, consequently, were eliminatedfrom the analysis. Discarding these fossil sitesdid not result in the deletion of any species,and the total number of occurrences (species 3
sites) was 523 for the analysis interval.Once the localities were assigned a numer-
ical age estimate (Supplementary Table 4),they were parsed into time bins of 0.5 Myrover the analysis interval. We chose 0.5-Myrtime units so that fossil sites occurred in alltime bins (minimum number of fossil sites¼ 2;maximum number of fossil sites¼ 6; median¼4) and, because our knowledge of age rela-tionships did not warrant finer time resolu-tion. The time bin from 12.0 to 11.5 Maincludes fossil sites and species dated from11.99 to 11.50 Ma, the time bin from 11.5 to11.0 Ma includes fossil sites and species datedfrom 11.49 to 11.00 Ma, and so on. We refer totime bins by their midpoint age; for example,the time bin 12.0 to 11.5 Ma is denoted as timebin 11.75 Ma. After the ML AEO wasperformed, a few fossil localities were placedin MN units different from those traditionallyassigned to those sites (Supplementary Fig. 1).
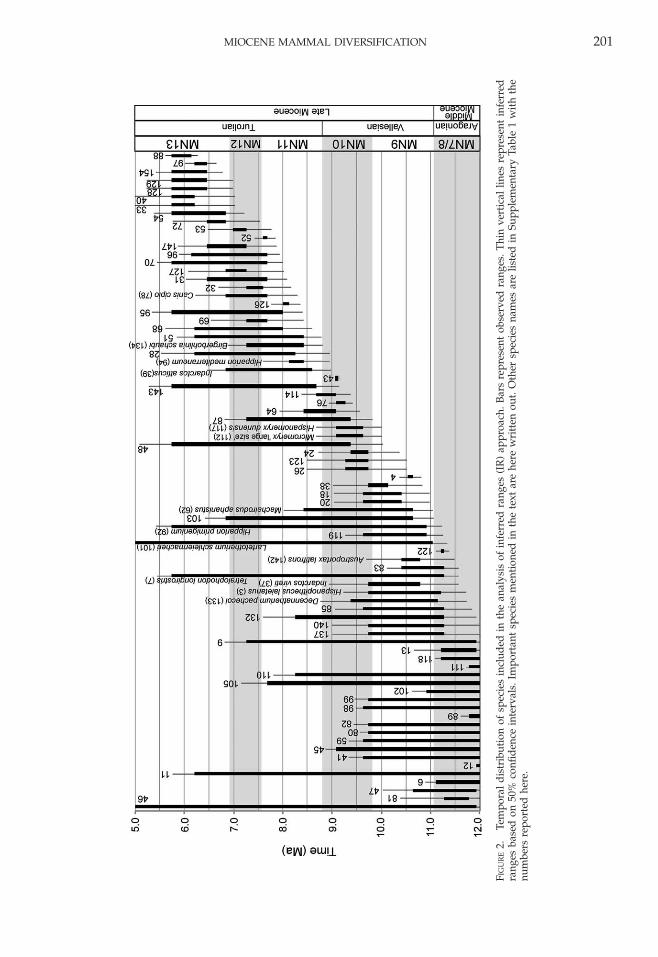
Estimation of Confidence Intervals.—The agesof the fossil sites estimated by ML AEO wereused to date the stratigraphic ranges ofmammal species. The age of the oldest fossilsite at which a species appears was consideredto be its observed first appearance (OFA) andthe age of the youngest fossil site at which thatspecies appears was considered its observedlast appearance (OLA).
Based on the duration of the observedtemporal range of each species and on thenumber of horizons documented for eachspecies, 50% confidence intervals on the OFAand OLA were estimated following the meth-od of Marshall (1990) (Fig. 2, SupplementaryTable 5). To employ this method, we tested thedata for two assumptions from Marshall(1990): (1) that the gap size between horizonswas not correlated with stratigraphic position(assessed with a Spearman rank test), and (2)that the distribution of gaps was random
(assessed with a Kolmogorov-Smirnov test).Fossil sites with the same age estimate andfrom the same basin were considered to occurin a single horizon.
For species with either their OFA or OLAwithin the study interval, we estimated theconfidence interval on either the base or thetop of their observed temporal range with theequation:
a ¼ ð1� CÞ�1=ðH�1Þ � 1; ð1Þ
in which a is the confidence interval expressedas a proportion of the total observed strati-graphic range, C is the confidence level (50%in our study), and H is the number offossiliferous horizons (¼ fossil sites, with thecaveat above).
For species that have both the OFA andOLA within the study interval, the end-pointswere estimated jointly. In this case, the valueof a is obtained from the equation:
C ¼ 1� 2ð1þ aÞ�ðH�1Þ þ ð1þ 2aÞ�ðH�1Þ: ð2Þ
Once the values of a are known, then theInferred First Appearance (IFA) and theInferred Last Appearance (ILA) are calculatedas follows:
IFA ¼ OFA� aðOLA�OFAÞ; ð3aÞ
ILA ¼ OLAþ aðOLA�OFAÞ: ð3bÞ
Confidence intervals cannot be estimatedfor species found in only one horizon or fossilsite (i.e., singletons). The number of singletonsin our database was 62 (Table 1).
After estimating the 50% confidence inter-vals, observed and inferred first and lastappearances were compared. We evaluatedthe tradeoff between having a large sample ofspecies to analyze versus the magnitude of theconfidence interval. Large confidence intervals(.1.0 Myr) usually corresponded to taxafound in only two or three fossil horizonsthat occur far apart in time. For these taxa, the0.50 probability of finding them before andafter their observed temporal ranges was solarge that, in our opinion, incorporating theirIFA and ILA contributed more noise thanvaluable information to our analysis. Weselected only species with a difference of �1
200 M. SOLEDAD DOMINGO ET AL.
FIG
UR
E2.
Tem
po
ral
dis
trib
uti
on
of
spec
ies
incl
ud
edin
the
anal
ysi
so
fin
ferr
edra
ng
es(I
R)
app
roac
h.
Bar
sre
pre
sen
to
bse
rved
ran
ges
.T
hin
ver
tica
lli
nes
rep
rese
nt
infe
rred
ran
ges
bas
edo
n50
%co
nfi
den
cein
terv
als.
Imp
ort
ant
spec
ies
men
tio
ned
inth
ete
xtar
eh
ere
wri
tten
ou
t.O
ther
spec
ies
nam
esar
eli
sted
inS
up
ple
men
tary
Tab
le1
wit
hth
en
um
ber
sre
po
rted
her
e.
MIOCENE MAMMAL DIVERSIFICATION 201

Myr between their observed and inferred firstand last appearances for further analysis. Thetotal number of such species was 80 (Table 1,Fig. 2, Supplementary Table 5), correspondingto 86% of the initial number of species aftersingletons were excluded.
We used 50% confidence intervals in ourstudy as a compromise between a reasonableconfidence level and the number of speciesretained in the analysis for different levels ofconfidence. For example, the number ofspecies maintained in the analysis had weused 80% confidence intervals would havebeen only 46 (i.e., only ~50% of the total non-singletons species). The overall results of ouranalysis with 50% confidence intervals werequalitatively similar to those obtained with80% confidence intervals.
Diversity Metrics.—For each time interval,we determined four variables following themethod of Foote (2000): (1) NFL, the number oftaxa confined to the time interval of interest(in our study, these species are considered tobe single-bin species; note that singletons,which are species found in only one horizon,are included in this category as well), (2) NbL,the number of taxa that cross the bottom(older) boundary only, (3) NFt, the number oftaxa that cross the top (younger) boundaryonly, and (4) Nbt, the number of taxa that crossboth boundaries. Unless otherwise indicated,Nbt includes taxa that are recorded before,during, and after the interval of interest aswell as range-through taxa that are inferred tobe present in the focal interval because theyoccur in the preceding and succeeding inter-vals (Supplementary Table 6). We estimatedspecies diversity as the total number of speciesin these four categories for each time bin.
We calculated per capita origination rate,p(i), and extinction rate, q(i), using equationsfrom Foote (2000):
pðiÞ ¼ �ln�
NbtðiÞ=NtðiÞ�=Dt; ð4Þ
qðiÞ ¼ �ln�
NbtðiÞ=NbðiÞ�=Dt; ð5Þ
where Nt (i)¼NFt (i)þNbt (i); Nb (i)¼NbL (i)þNbt (i); and Dt ¼ interval length. Dt can beeliminated from the equations when all time
intervals have equal duration. In this study,origination and extinction refers not only tospeciation and true extinction but also togeographic-range extensions and contractions.
Single-bin taxa are particularly sensitive tointerval length and to the quality of preserva-tion (their number is greater in long, inten-sively sampled time intervals), which can giverise to spurious values of diversity metrics.Foote’s (2000) diversity metrics standardizerates according to changes in standing diver-sity and omit the sampling effects associatedwith single-bin species. Even though single-bin species were excluded, sampling intensitycan still exert an influence on origination andextinction rates. We evaluated the quality ofthe fossil record throughout the study interval,using Foote’s equation for assessing thepreservation rate:
rðiÞ ¼ �ln�
1�Nbt;sampðiÞ=NbtðiÞ�=Dt; ð6Þ
where Nbt,samp is the number of speciespresent before, during and after the timeinterval of interest and Nbt is the same numberplus range-through species (Foote 2000). Then,correlation tests (Pearson’s r or Kendall’s s,depending on the distribution properties ofthe sample) were performed on the firstdifferences to determine whether changes inp(i) and q(i) were related to changes in r(i). Theuse of first differences emphasizes whetherchanges in preservation rate influence changesin diversity metrics.
For comparative purposes, we reporteddiversity metrics for three versions of theSpanish large mammal record that differbased on the timescale used and the temporalranges of the taxa: (1) the IR (inferred range)version, based on the IFA and ILA of speciesanalyzed in 0.5-Myr time bins (Fig. 2); (2) theMN version, based on the traditional alloca-tion of fossil sites to MN units (SupplementaryFig. 2); and (3) the OR (observed range)version, based on the OFA and OLA of speciesanalyzed in 0.5-Myr time bins (SupplementaryFig. 3).
As a measure of net change in diversitythrough time, we calculated per-lineage netdiversification following the procedure ofFinarelli and Badgley (2010):
202 M. SOLEDAD DOMINGO ET AL.

dðiÞ ¼��
NtðiÞ �NbðiÞ�=NbtðiÞ
�=Dt: ð7Þ
Net diversification represents the differencebetween the number of originations andextinctions in a given time bin, scaled to thediversity passing through that time bin.Periods of significantly high diversificationrate have either large origination rates or largeextinction rates but not both, because thatwould result in d(i) ’ 0.
The turnover rate accounts for the totalamount of faunal change for each interval andis expressed as
tðiÞ ¼��
NbLðiÞ þNFtðiÞ�=NbtðiÞ
�=Dt: ð8Þ
Turnover rate compares the number ofspecies that pass through both upper andlower boundaries of the interval (Nbt, which isthe portion of fauna that remains static) withthe number of species that do not passthrough (NFt and NbL, the portion of the faunathat changes) (Badgley and Finarelli 2013).Significant t(i) can result from (1) significantlylarge p(i) and low to moderate q(i), in whichcase new species join the previously existingtaxa without significant displacement or ex-tinction of the resident taxa; (2) low tomoderate p(i) and significantly large q(i), inwhich case the resident fauna experienceshigh levels of disappearances without thearrival of new immigrants or without specia-tion in situ; or (3) significantly large p(i) andsignificantly large q(i), which represent thereplacement of most of the existing species bynew species.
The significance of the diversity metrics wasevaluated by bootstrapping the temporaldurations of taxa with replacement 500 timesand obtaining new estimates for p(i), q(i), d(i),and t(i) across all time intervals. For eachmetric, we defined confidence intervals as twostandard deviations of the mean of thebootstrap distribution. The observed diversitymetric was considered significant when theconfidence interval of the bootstrap distribu-tion did not include 0 (Finarelli and Badgley2010).
We assessed changes in taxonomic compo-sition, trophic structure, and size structureover time following the multinomial likeli-
hood approach of Finarelli and Badgley(2010). In this approach, the proportionalrepresentation of each family or ecologicalcategory in relation to total diversity within atime bin was compared with proportionsbased on the previous time bin. For each timeinterval i, the log-likelihood of the multinomi-al distribution is given by
LnLðiÞ ¼X
j
ajlnðpjÞ; ð9Þ
where aj is the count of species in family (orecological category) j, and p is j’s proportion oftotal diversity in time interval i for each family(or ecological category) (Edwards 1992). TheLnL for interval i is the sum over all j families(or ecological categories). Next, we calculatedLnL using counts for interval i but proportionsfor interval i–1. The difference between thesetwo LnL values increases as faunal propor-tions change to a greater extent from one timeinterval to the next. Values of LnL(i) �LnL(i�1) greater than 2 were indicative ofsignificant change in faunal proportions be-tween time intervals (Edwards 1992). Thefamilies (or ecological categories) with highLnL differences between adjacent intervals ofsignificant change indicated which taxa orecological categories were the major contribu-tors to faunal change.
Finally, we performed principal componentanalysis (PCA) of the number of species perfamily or ecological category in each time binin order to identify time intervals that grouptogether or differ strongly in terms of taxo-nomic composition, trophic structure, or sizestructure. We used raw data (species counts)and utilized the covariance matrix for each ofthree PCAs (based on taxonomic composition,trophic structure, or size structure). Theeigenvalues, scores, and loadings of principalcomponents 1 (PC1) and 2 (PC2) are reportedfor each analysis. When the PCA produced aclear grouping of time intervals, we assessedthe significance of groups with an analysis ofsimilarity test (ANOSIM) using the Bray-Curtis similarity index. The PCAs and ANO-SIM were performed with PAST version 2.10(Hammer and Harper 2006).
MIOCENE MAMMAL DIVERSIFICATION 203

Results
Species Diversity
For the inferred ranges of taxa with 50%confidence intervals (IR approach), speciesdiversity changed little between 12.0 and 5.5Ma (Fig. 3C). Between 11.0 and 9.0 Ma,diversity increased somewhat, then decreasedand remained stable until the time bin at 6.25Ma, when it declined to the lowest level overthe analysis interval. For the IR approach,species diversity was generally lower than inthe MN approach or the OR approach,because IR diversity was based on a reduceddata set. For the MN approach, if all specieswere included, species richness peaked inMN9 (Fig. 3A, Supplementary Table 6); ifsingle-bin species were removed from theanalysis, then richness in MN9 decreasednotably and became comparable to the rich-ness in MN7/8. A similar trend occurred with
the OR approach: if all species were counted,then a conspicuous peak occurred at the 9.75Ma time bin, but that peak decreased if single-bin species were excluded (a small peakpersisted) (Fig. 3B, Supplementary Table 6).When single-bin species were excluded, boththe MN approach and the OR approachshowed higher species diversity in the firstpart of the analysis interval and a decrease inthe second half.
Origination, Extinction and Preservation Rates
In the IR approach, the highest originationrate, p(i), occurred at time bin 11.75 Ma (Figs.3F, 4A). A significant p(i) also occurred duringfive additional intervals in the early andmiddle portions of the analysis interval (Fig.4A). The p(i) trend of the IR approach differedfrom those in both the MN and OR approach-es (Fig. 3), which had higher p(i) at MN9 andMN12 and at the 11.25, 9.75, 7.75 and 6.25 Ma
FIGURE 3. Species diversity and per capita rates of origination, p(i), and extinction, q(i), for each of three approaches. A,Diversity for the MN approach. B, Diversity for the OR approach. C, Diversity for the IR approach. D, Origination andextinction rates for the MN approach. E, Origination and extinction rates for the OR approach. F, Origination andextinction rates for the IR approach. Species diversity for all species refers to the total number of species (NFLþNbLþNFtþNbt) in each time bin. For the estimation of non-single-bin species diversity, we excluded the NFL term. All-speciesdiversity and non-single-bin species diversity are identical for the IR approach.
204 M. SOLEDAD DOMINGO ET AL.

FIGURE 4. Diversity metrics using the IR approach. A, Per capita rate of origination, p(i), and extinction, q(i). B, Netdiversification rate, d(i). C, Turnover rate, t(i). Asterisks denote significant values as inferred from the bootstrappingroutine applied to the diversity metrics. Bootstrapped confidence intervals for p(i), q(i), d(i) and t(i) are 0.160, 0.185, 0.348,and 0.394, respectively.
MIOCENE MAMMAL DIVERSIFICATION 205

time bins, respectively. For the IR approach,the highest extinction rates, q(i), occurredduring two intervals at 9.25 Ma and 5.75 Ma;adjacent time bins at 8.75 Ma and 6.25 Ma also
recorded significant q(i). Although shifted intime, q(i) also exhibited two major peaks forthe MN and OR data sets, in MN9 and MN13
(Fig. 3D) and in time bins 9.75 Ma and 5.75 Ma(Fig. 3E), respectively.
For the MN data set, changes in p(i) werecorrelated with changes in preservation rate,
r(i) (Table 2). For the OR approach, changes inq(i) were correlated with changes in preserva-tion rate and changes in p(i) were marginally
correlated with r(i). The fact that originationand extinction rates covary with the quality ofthe record makes the biological interpretationof patterns difficult under these two approach-
es.
It was not possible to determine r(i) for theIR approach. When the confidence intervals
are estimated, the distinction between range-through taxa and taxa actually sampled in atime interval—the terms in the r(i) formula—disappears. The basis for estimating the
preservation rate lies in detecting whetherOFAs and OLAs (and therefore, the origina-tions and extinctions) in a sequence could bethe result of sampling bias. If the correlation is
significant, then it is probable that originationsand extinctions are overestimated in sometime bins and underestimated in others. The
use of confidence intervals addresses thisproblem to a large degree. This study andprevious studies (e.g., Casanovas-Vilar et al.2010) demonstrate that the MN approach and
the OR approach are potentially biased by thepreservation of the record; therefore wefocused on the IR approach for the remaininganalyses.
Diversification and Turnover Rates
Significant diversification rates, d(i), oc-curred during the time intervals 9.25 Ma and5.75 Ma (Fig. 4B); both episodes resulted froman elevated number of extinctions coupledwith low levels of origination. Significantturnover, t(i), occurred during six time inter-vals at the beginning, middle, and end of theanalysis interval (Fig. 4C). These intervals canbe grouped into three periods of significantturnover separated by periods of relativestability. At 11.75 Ma, 11.8% of the totalturnover throughout the sequence occurred,mainly due to originations. From 9.5 to 7.5Ma, 44.3% of the total turnover occurred infour successive time intervals, as a conse-quence of both originations and extinctions.This was a prolonged period of faunal changewhen extinctions were greater in the first twointervals, whereas originations were greater inthe two succeeding time bins. The interval5.75 Ma contains 10.1% of the total turnover,mainly due to extinctions. Overall, these threetime periods (six 0.5-Myr intervals) include66.2% of the total turnover.
Changes in Taxonomic Composition andEcological Structure
By the likelihood method of comparingadjacent time intervals, the taxonomic compo-sition of large mammals exhibited threeintervals of significant change. At 10.75 Ma,change was dominated by an increase in thenumber of mustelids (Fig. 5A,B). The greatestamount of change occurred at time interval8.75 Ma as a consequence of changes inproportions of several families. At timeinterval 8.25 Ma, change was dominated byan increase in the number of cervids.
The PCA of taxonomic composition, basedon the number of species in each family per
TABLE 2. Correlation tests of first differences of p(i) vs. r(i) and q(i) vs. r(i) for the MN approach and the OR approach.We used Pearson’s r when samples were normally distributed; otherwise, we used Kendall’s s. p , 0.05, significantlycorrelated; p . 0.05, not significant.
MN approach OR approach
p(i) vs. r(i) q(i) vs. r(i) p(i) vs. r(i) q(i) vs. r(i)
Pearson’s r – 0.805 0.534 0.591Kendall’s s 1.000 – – –p 0.042 0.195 0.074 0.043
206 M. SOLEDAD DOMINGO ET AL.

FIGURE 5. Faunal composition at the family level through time. A, Family-level faunal composition expressed asproportions in each time bin. B, Log-likelihood ratios of faunal composition comparing adjacent 0.5-Myr time bins.Likelihood differences greater than 2 indicate significant change in faunal composition for that time interval compared tothe previous one. C, PCA of faunal composition, biplot for the two first principal components (PC1 and PC2 explained78.77% of the variance). PC1 separates the time bins into two groups: one that includes time intervals from 11.75 to 9.25Ma (squares) and another from 8.75 to 5.75 Ma (circles). 11.75 Ma represents the time bin from 12.0 to 11.5 Ma, 11.25 Marepresents the time bin from 11.5 to 11.0, and so on. Families are indicated for loadings greater thanþ0.15 or less than�0.15 on PC1 or PC2. The loadings are given in Supplementary Table 7.
MIOCENE MAMMAL DIVERSIFICATION 207

time interval, separated the sequence into twodistinctive periods (Fig. 5C). PC1, with 63.95%of the variance, separated time intervals intotwo groups that differed significantly infaunal composition (ANOSIM test, Bray-Cur-tis index: R ¼ 0.97, p , 0.01). One clusterincluded intervals from 11.75 to 9.25 Ma,when faunas contained more suids, rhinocer-oses, and hominids among the herbivores andamphicyonids and barbourofelids among thecarnivores. The other cluster included inter-vals from 8.75 to 5.75 Ma, when faunas hadmore equids, felids, and hyaenids (Fig. 5C,Supplementary Table 7). PC2, with 14.83% ofthe variance, mainly contrasted faunas withhigh versus low numbers of musk deer(Moschidae), with time bins 9.25 and 8.75Ma recording the highest number of moschidspecies.
In terms of trophic structure, the likelihoodmethod showed a significant change at 8.75Ma compared to the previous interval (Fig.6A,B). This change was mainly the conse-quence of an increase in the number ofinvertebrate faunivores and mixed feeders. Inthe PCA, PC1, with 77.10% of the variance,again separated time intervals into two dis-tinct clusters (ANOSIM test, Bray-Curtis in-dex: R¼ 0.84, p , 0.01). Time bins from 11.75to 9.25 Ma contained a large proportion ofbrowsing species and some frugivores amongthe herbivores, whereas time bins from 8.75 to5.75 Ma featured an increase in the proportionof grazers, fewer browsers, and absence offrugivores (Fig. 6C, Supplementary Table 8).PC2, with 14.78% of the variance, contrastedtime bins with higher versus lower propor-tions of carnivores.
In terms of size structure, the only period ofsignificant change occurred at 7.75 Ma by thelikelihood analysis, as a consequence of anincrease in the number of species with a bodymass of 10–45 kg (Fig. 7A,B). The PCA did notcluster time intervals as a function of their age(Fig. 7C), although the four youngest intervalslie at one end of the axis. PC1, with 53.91% ofthe variance, contrasted time bins with ahigher proportion of species in size categoryB (1–10 kg) against time bins with morespecies in size category C (10–45 kg) (Fig.7C, Supplementary Table 9). PC2, with 24.34%
of the variance, differentiated time bins withhigher and lower proportions of mammals insize categories 10–45 kg and .360 kg.
Discussion
Our results revealed differences among thethree approaches to binning diversity by timeinterval and, for the IR approach, highlightedthree time intervals of important change in thediversity and ecological structure of largemammals.
Diversity per time interval fluctuated be-tween about 20 and 40 species (excludingsingle-bin species) for the MN, OR, and IRdata sets (Fig. 3). For both MN and OR datasets (Fig. 3A,B), diversity was greater in timeintervals older than 9.5 Ma and lower there-after. For the IR data set (Fig 3C), diversityremained high between 11.0 and 9.0 Ma, thendeclined slightly thereafter. The long, unevenduration of MN units has the effect ofobscuring changes in diversity relative to theOR and IR approaches and lacks the temporalresolution of diversification metrics at finertime scales (Fig. 3). The IR approach dampenschanges in diversity among time intervals(because of the reduced data set) but stillreveals significant changes in diversification.
For both the MN and OR approaches,origination and extinction rates were correl-ated with the preservation rate, implying thatdiversity changes could be at least partly anartifact of changes in the quality of the record.For example, the peak in species diversity inMN9 is probably a consequence of the higherfrequency of rich fossil localities compared toMN10 (Supplementary Table 4) (Casanovas-Vilar et al. 2012). Specifically, one localityaccounts for much of the diversity in MN9: thelocality of Can Llobateres (Valles-PenedesBasin) contains 34 (non-singleton) large mam-mal species. When single-bin species wereremoved from the MN analysis, the MN9 peakdisappeared but it is likely that the subsequentdecrease in species diversity still reflected thechange in the preservation rate.
The OR data set also shows the influence ofchanges in the quality of the record. The MNboundaries were those of Agustı et al. (2001)recalculated with the 2004 geomagnetic time-scale (Gradstein et al. 2004), with the conse-
208 M. SOLEDAD DOMINGO ET AL.

FIGURE 6. Trophic structure through time. A, Proportion of species in each trophic category expressed as percentages. B,Log-likelihood ratios of trophic categories comparing adjacent 0.5-Myr time bins. Likelihood differences greater than 2indicate significant change in trophic categories for that time interval compared to the previous one. C, PCA of trophicstructure, biplot for the two first principal components (PC1 and PC2 explained 91.88% of the variance). PC1 separatesthe sequence in two groups: one that includes time intervals from 11.75 to 9.25 Ma (squares) and another from 8.75 to5.75 Ma (circles). The loadings are given in Supplementary Table 8.
MIOCENE MAMMAL DIVERSIFICATION 209

FIGURE 7. Body-size structure through time. A, Proportion of species in each size category expressed as percentages. B,Log-likelihood ratios of body-size categories comparing adjacent 0.5-Myr time bins. Likelihood differences greater than 2indicate significant change in body-size categories for that time interval compared to the previous one. C, PCA of body-size structure, biplot for the two first principal components (PC1 and PC2 explained 78.25% of the variance). Notemporal pattern is evident. The loadings are given in Supplementary Table 9.
210 M. SOLEDAD DOMINGO ET AL.

quence that some localities in time bin 9.75Ma, which belonged to MN9, fall in the MN10time span in the OR approach. Also, aspreviously indicated, the ordination of local-ities through the ML AEO method hasresulted in the shift of a few localities fromone MN unit to the preceding or succeedingone (Supplementary Fig. 1). Such was the casefor Can Llobateres and Los Valles deFuentiduena, two very rich localities thatproduced a peak in diversity at 9.75 Ma,followed by an apparent decrease in richness,which was further enhanced by the lowerquality of the record in the subsequent timebin (Fig. 3B, Supplementary Table 4).
In contrast, the use of confidence intervalson the observed temporal ranges of taxadiminished the fluctuations associated withvariable preservation and offers a morerealistic view of the true appearances anddisappearances of taxa. In the IR approach,the high number of originations and extinc-tions at the beginning and end of the analysisinterval (Fig. 4A) raises the question aboutwhether the data set is influenced by edgeeffects. We evaluated occurrence data fromoutside the analysis interval to avoid sucheffects. When confidence intervals fell outsidethe analysis interval, we counted those taxa asspecies that were present in those time bins,but not as appearances at the beginning of thesequence or disappearances at the end. Forthese reasons, the patterns obtained with theIR approach should not be the consequence ofedge effects. Nevertheless, we did not estimateconfidence intervals for species disappearingbefore or appearing after the analysis interval,and thus extinctions at the beginning andoriginations at the end of the sequence mightbe underestimated (see below).
Three Intervals of Significant Faunal Change
The diversity metrics highlighted three timeintervals when two or more of the metricshave significant values (Fig. 4). The firstinterval of significant faunal changes encom-passes the first three time bins, between 12.0and 10.5 Ma. Origination rates (p(i)) weresignificant during each 0.5-Myr time bin, andthe turnover rate (t(i)) was significant at timebin 11.75 Ma. Although extinction rates (q(i))
were nonsignificant over this interval, therewere enough extinctions that net diversifica-tion (d(i)) values were also nonsignificant. Wedid not estimate confidence intervals forspecies disappearing just before 12.0 Ma, anddoing so might have yielded higher extinctionrates than reported here. Nevertheless, webelieve that would not be the case: there arenot major declines in the number of speciesfrom older fossil sites (73% of the speciespresent in MN6 were present in succeedingtime intervals). Therefore, the estimation ofconfidence intervals for pre-12.0 Ma taxashould not have added a significantly highnumber of disappearances to the beginning ofour analysis interval.
The IFAs of 22 large mammal species fellinto this first interval of significant faunalchange (Supplementary Table 5) and diversityincreased. Faunal composition changed sig-nificantly at time bin 10.75 Ma (Fig. 5B) withan increase in the proportion of mustelids.Neither trophic structure nor size structurechanged significantly during this interval(Figs. 6A,B, 7A,B).
Among the species with IFAs during thisperiod of high origination rates, severallineages are inferred to have appeared in theIberian Peninsula earlier than reported inother studies. Confidence intervals extendedthe temporal range of the hominoid Hispano-pithecus from 11.1–9.5 Ma (this is the observedrange; Casanovas-Vilar et al. 2011) to 11.7–9.2Ma (this is the inferred range), therebyimplying that this genus (including the speciesH. laietanus and H. crusafonti) first appeared bythe late Aragonian instead of the earlyVallesian (Fig. 2, Supplementary Table 5).Other notable taxa that are inferred to haveappeared in the Iberian Peninsula earlier thanreported elsewhere include the ursid Indarctosvireti, the rhinoceros Lartetotherium schleierma-cheri, and the bovid Austroportax latifrons. TheOFAs of these three species have traditionallybeen documented in the Vallesian but ouranalysis places the IFAs of these species in thelate Aragonian (Fig. 2, Supplementary Table5).
The first hipparionine horse documented inthe Iberian Peninsula is Hipparion primigenium(Hippotherium primigenium sensu Bernor et al.
MIOCENE MAMMAL DIVERSIFICATION 211

1996, 1997), with an IFA of 11.24 Ma (Fig. 2,Supplementary Table 5). Hipparionine horsesdispersed into Eurasia from North Americafollowing the reestablishment of the Beringland bridge at around 12.0 Ma, when growthof the Antarctic Ice Sheet led to a drop in sealevel (Agustı and Anton 2002). Based on OFAs,Domingo et al. (2007) suggested that the age of11.1 Ma proposed by Agustı et al. (2001) for theentrance of hipparionine horses in the IberianPeninsula was too old. However, according tothe 50% confidence intervals, the IFA is evenolder than that proposed by Agustı et al.(2001). The elevated number of appearancesduring this period corresponds to an intervalof mobility of mammals over much of theworld. Several mammalian lineages crossedthe Bering land bridge and made their firstappearances in Eurasia (although they did notnecessarily reach western Europe) or NorthAmerica, including Hipparion, the carnivoransParataxidea and Barbourofelis, and the gelocidruminant Pseudoceras (Tedford et al. 2004).Other taxa that appeared during this periodin the Iberian Peninsula were immigrants fromAsia (the giraffid Decennatherium pachecoi andthe machairodontine Machairodus aphanistus)or Africa (the proboscidean Tetralophodon long-irostris). Most of the remaining species thatappeared in the Iberian Peninsula were alreadypresent in the European record, so it is likelythat many of these appearances resulted fromgeographic range shifts of taxa to lowerlatitudes in response to global cooling.
Significant turnover driven mainly by highorigination rates agrees with the assessment ofAgustı and Anton (2002) that the immigranttaxa joined the resident faunas of the IberianPeninsula without displacement of potentialcompetitors, as shown by the low levels ofextinction, increase in large mammal diversity,and stable ecological structure during thisinterval.
The second interval of significant faunalchange occurred between 9.5 and 7.5 Ma.Significant origination and extinction rates aswell as change in composition and faunalstructure characterize this interval. Origina-tion rates were significant during the time binsof 8.75, 8.25, and 7.75 Ma, whereas extinctionrates were significant at 9.25 Ma and 8.75 Ma
(Fig. 4A). Diversification was significant at9.25 Ma, and turnover was significant overthis entire period (Fig. 4B,C).
The significant extinction rates in our studycoincide with the period of significant extinc-tions detected during MN10 by Casanovas-Vilar et al. (2010) for large and small mam-mals, although our study offers more detailedinformation on the timing of change. Incontrast to our results, these authors did notdetect significant originations between 9.0 and7.5 Ma for large mammals. The differencebetween their results and ours may be due totheir use of the MN timescale and their use ofgenera instead of species as their unit of faunalanalysis.
The use of 50% confidence intervals sug-gests that some of the species that werethought to have become extinct in the Valle-sian survived until the Turolian. These speciesinclude the musk deer Micromeryx ‘‘large size’’and Hispanomeryx duriensis and the giraffidDecennatherium pachecoi (Fig. 2, Supplementa-ry Table 5). Also, species that contributed tosignificant origination rates with IFAs earlierthan previously thought (i.e., the IFA lies inthe late Vallesian instead of the Turolian)include the ursid Indarctos atticus, the equidHipparion mediterraneum, and the giraffidBirgerbohlinia schaubi. The earliest canids inthe Iberian Peninsula were reported to appearin MN12 with Canis cipio. The confidenceintervals suggest that the first appearance ofCanis cipio happened earlier, around 8.3 Ma(Fig. 2, Supplementary Table 5).
Significant extinctions between 9.5 and 8.5Ma coincided with a set of extinctions docu-mented in previous work and named the‘‘Vallesian Crisis’’ (Agustı et al. 1999a; For-telius et al. 2006; Casanovas-Vilar et al. 2010).An important difference from previous studiesin our analysis is that extinction remainedsignificantly high at the beginning of MN11(Fig. 3F). Thus, faunal change occurred over aprolonged period when high rates of extinc-tion were followed by high rates of originationuntil 7.5 Ma; i.e., originations were elevateduntil roughly the end of MN11 (Fig. 3F). Highlevels of origination were detected in MN12 inprevious studies based on the MN system(e.g., Casanovas-Vilar et al. 2010) but were not
212 M. SOLEDAD DOMINGO ET AL.

detected in the IR approach (Fig. 3D,F). Thehigh origination rates were probably anartifact of a low preservation rate duringMN11 followed by a high preservation ratein MN12.
The significant diversification and turnoverrates between 9.5 and 7.5 Ma marked a periodof substantial change in faunal compositionand structure. Significant changes in log-likelihood ratios of taxonomic compositionoccurred during the time bin at 8.75 Ma, as theconsequence of shifts in the proportions ofseveral families (e.g., increase in the Equidae,Hyaenidae, and Felidae, and decrease in theSuidae and Rhinocerotidae), and at 8.25 Madriven by an increase in the Cervidae (Fig.5A,B). A significant change in trophic struc-ture at 8.75 Ma (Fig. 6A,B) resulted from anincrease in the number of invertebrate fauni-vores and mixed feeders compared to theprevious time bin (a decrease of browsing andomnivorous species marginally contributed tothis change in trophic structure as well). Forbody-size structure (Fig. 7A,B), the onlyinterval of significant change occurred at 7.75Ma and resulted from an increase of species incategory C (10–45 kg), mainly represented atthis time by hyaenids, felids, and the bovidGazella deperdita.
Change in faunal composition was alsorevealed in the PCAs of taxonomic composi-tion and trophic structure (Figs. 5C, 6C). Inboth analyses, PC1 contrasted faunas fromolder time intervals (11.75 to 9.25 Ma) withfaunas from younger time intervals (8.75 to5.75 Ma). In faunas from time intervals 11.75to 9.25 Ma, there was greater representation ofherbivores associated with more forestedenvironments, such as the hominoids, rhino-cerotids, and suids that are mainly classifiedas frugivores, browsers, and omnivores.Equids were important components of herbi-vore faunas in younger time intervals andmost of these species were grazers (accordingto the NOW database; Fortelius 2012). Bothgrazers and mixed feeders increased as thebrowsing species diminished over time. In theIberian Peninsula, the increase in grazers wasnot related to the expansion of C4 grasses, asoccurred in several other regions of the worldbetween 8 and 3 Ma (e.g., Cerling et al. 1997;
Stromberg 2011). Isotopic analyses of herbi-vore tooth enamel from the Iberian Peninsulahave demonstrated no significant consump-tion of C4 vegetation by any of the speciesincluded in the analysis (Domingo et al. 2013).As in regions of Mediterranean vegetationtoday, C3 vegetation has always been domi-nant in the Iberian Peninsula. In the carnivoreguild, older faunas were rich in amphicyonidsand barbourofelids (Fig. 5); then, between 9.5and 9.0 Ma, these two families vanished fromthe Iberian Peninsula. Younger faunas showedan increase in felids and hyaenids and thearrival of the first canids.
This second period of faunal change in-volved substantial replacement of lineagesrather than extinction and speciation withinresident lineages. During this period, 70% ofthe large mammal genera of those species thatwent extinct, as well as the families Amphi-cyonidae, Barbourofelidae, and Hominidae,disappeared from the Iberian Peninsula. Mostof the new species that appeared in the IberianPeninsula in this second period of changewere already present in eastern Europe or infossil sites from Asia (mainly Turkey, Iran, andChina) and expanded from there into westernEurope (Agustı and Anton 2002; Casanovas-Vilar et al. 2010) or had their first appearancein contemporaneous localities from these areasand the Iberian Peninsula.
In the Iberian Peninsula, this substantialfaunal change corresponded to changingclimatic conditions and increased seasonalitytoward drier, more open habitats (Agustı et al.1999a; Fortelius et al. 2006; Casanovas-Vilar etal. 2010). Macrofloral remains from the Valles-Penedes Basin document the replacement ofevergreen subtropical woodland by moreseasonal, deciduous woodland (Agustı et al.2003; Marmi et al. 2012). Stable isotopeanalyses of herbivore enamel from differentbasins of the central Iberian Peninsula show,by this time, an increase in d13C values,suggesting a shift toward more open wood-land as well as a decrease in mean annualprecipitation of ~200 mm/yr (Domingo et al.2013). In their study of rodent faunas from theIberian Peninsula, Casanovas-Vilar andAgustı (2007) highlighted that in the interiorbasins increasing seasonality occurred as the
MIOCENE MAMMAL DIVERSIFICATION 213

lengthening of the dry season, whereas innear-coastal basins from eastern Iberia, sea-sonality in precipitation was accompanied byan increase in seasonality in temperature. Inthis context, species that relied on the presenceof fruits throughout year, such as the homi-noids, declined and became extinct becausefruits were not present during the drier (andcooler) seasons developed at this time.
The extent of the Vallesian Crisis in otherregions of Europe is still under debate. InCentral Europe, this episode seems to extenduntil the end of MN10, when the amphicyo-nids and hominids disappeared (Franzen andStorch 1999). MN9 localities are rare in theGreek fossil record, making it difficult toassess the importance of this episode in thisarea (Bonis and Koufos 1999). Fortelius et al.(2006) indicated that in eastern Europe speciesrichness doubled during this period, whereasLunkka et al. (1999) documented a gradualdecrease in mammal diversity in the SinapFormation of Turkey. Casanovas-Vilar et al.(2005) interpreted the Vallesian Crisis asaffecting only southwestern Europe.
The third period of significant faunalchange occurred from 6.5 Ma to 5.5 Ma.Significant rates of extinction in time intervals6.25 Ma and 5.75 Ma coincided with loworigination rates (Fig. 4A), resulting in thelowest species diversity for the entire studyinterval. A significant negative diversificationrate and turnover rate occurred in interval5.75 Ma (Fig. 4B,C). Although the time intervalof 5.25 Ma was not included in this analysis,many (10) disappearances occurred in thistime period as well (Supplementary Table 5).This part of the analysis interval correspondsto the end of the Miocene, coinciding with theMessinian Salinity Crisis, which began at 5.96Ma (Krijgsman et al. 1999). Origination ratesbetween 6.5 and 5.5 Ma may be underesti-mated, because we did not determine confi-dence intervals for species with OFAs after 5.5Ma. Inferred ranges for those taxa could haveresulted in higher origination rates during thisthird period of faunal change, especiallybecause 87.5% of the species present in thesubsequent time interval (MN14; early Plio-cene) were new. Log-likelihood analyses oftaxonomic composition, trophic structure, and
body size indicated no significant changesduring this time (Figs. 5–7). Extinction affectedall trophic groups in the herbivore guild: threebrowsers, one mixed-feeder, and two grazersdisappeared during this period. Our study(Fig. 3D) and previous studies (Casanovas-Vilar et al. 2010) also detected high levels ofextinction and low diversity for the largemammals of the Iberian Peninsula duringMN13, which corresponds approximately tothe period of significant extinctions recog-nized here.
Agustı and Anton (2002) suggested thatduring the Messinian Salinity Crisis, thedisappearance of marine barriers promotedthe dispersal of mammals between NorthAfrica and southern Europe, when the firsthippopotamuses (genus Hexaprotodon), andmacaques (genus Macaca) of African originappeared in the Iberian Peninsula. Camels(genus Paracamelus) have an American originbut could have arrived to the Iberian Penin-sula via North Africa at the same time.However, the inferred range calculated forHexaprotodon crusafonti (IFA ¼ 6.78 Ma; Fig. 2,Supplementary Table 5) suggests that thisspecies was present in the Iberian Peninsulaalmost 1 Myr before the onset of the MessinianSalinity Crisis. Macaca and Paracamelus werenot included in our analysis but their OFAs insouthwestern Europe are dated at 6.1 Ma and6.23 Ma, respectively (van der Made et al.2006; Gibert et al. 2013). Agustı et al. (2006)and Gibert et al. (2013) have suggested thatthese dispersals were enabled by a significantrestriction of the Mediterranean-Atlantic con-nection and the presence of ephemeral landbridges between southwestern Europe andAfrica before the first episode of evaporitedeposition. Gibert et al. (2013) suggested thatthe falling sea level was the main driver forthese connections. Only the three taxa men-tioned above are commonly cited as cases ofspecies reaching the Iberian Peninsula fromNorth Africa at this time, suggesting that thenumber of large mammals dispersing fromAfrica to the Iberian Peninsula was limited.Moreover, their appearances pre-dated theonset of the Messinian Salinity Crisis. Duringthis period of interconnected landmasses, it issurprising that so little interchange of large
214 M. SOLEDAD DOMINGO ET AL.

mammals occurred across the desiccatedMediterranean Basin. The sparse or halophyticvegetation or the persistence of large evapo-ritic lakes may have inhibited the dispersal ofmammals, as may have the steep slopes anddepth of the basin (more than 4 km in someareas; Wust 1961).
Another feature of the late Miocene mam-malian faunas from the northern Mediterra-nean region is their regional endemism.Analysis of faunal similarity between thenortheastern (Greece, Anatolia, Caucasus/Iran) and northwestern (Iberian Peninsula)regions of the Mediterranean shows very lowlevels of similarity during MN13 (Costeur andLegendre 2008; Eronen et al. 2009; Casanovas-Vilar et al. 2010; the latter two studiescompared faunas at the generic level, whichis further evidence of the strong dissimilarityof mammal assemblages from this timeperiod) that extended until MN14 (earlyPliocene). In contrast, mammalian faunasfrom the previous time period (MN12)showed high similarity between these tworegions. This change suggests that strongbarriers to dispersal were established betweenthe northeastern and northwestern Mediterra-nean regions at the end of the Miocene.Stronger latitudinal temperature gradients incombination with increasing aridity in the lateMiocene (Pound et al. 2012) could haveenhanced climatic barriers along the southernmargin of the Pyrenees and Alps and restrict-ed many species to ecosystems in the Medi-terranean peninsulas. In addition, fromaround 5.0 Ma (with the timing slightly earlieror later for different parts of the Alps) until thepresent day, accelerated erosion rates in theSwiss and Western Alps imply rapid uplift(Kuhlemann 2007). By this time, moderate torapid uplift elevated the Carpathian, Dinaride,and Apennine ranges and also parts of theEuropean platform (e.g., Ardennes, Sudetes).This accelerated uplift may have furtherobstructed the dispersal of mammalian taxaacross the northern Mediterranean region.During the convergence of the Eurasian andAfrican plates, mountains of northwesternAfrica (Atlas, Rif, Tell ranges) and southernSpain (Betic range) also experienced upliftthroughout the Miocene (Gomez et al. 2000),
creating potential barriers to large mammalsdispersing between Africa and the IberianPeninsula. In fact, the historical biogeographyof modern taxa suggests that the AtlasMountains constituted an important barrierduring the middle to late Miocene, withseveral cases of vicariant speciation northand south of these mountains (e.g., Brown etal. 2002).
The Messinian Salinity Crisis resulted fromtectonic events that did not fundamentallyalter the terrestrial environmental conditions.Paleoenvironmental analyses based on pollen,stable isotopes of C and O from mammalianherbivore tooth enamel, and rodent assem-blages indicate stable vegetation and climaticconditions in terrestrial ecosystems during theMessinian Salinity Crisis in the northwesternMediterranean (e.g., Fauquette et al. 2006;Costeur et al. 2007; Jimenez-Moreno et al.2010; Domingo et al. 2013). The lack of anenvironmental response to the disappearanceof such a massive body of water and theprevalence of C3 vegetation in southwesternEurope led Matson and Fox (2010) to suggestthat the ‘‘Mediterranean climate,’’ in whichprecipitation occurs mainly during the winter,was already present by the latest Miocene andthat precipitation in the Iberian Peninsulawould have not depended upon the Mediter-ranean Sea but, as occurs today, had itsprimary source in the Atlantic Ocean.
It seems, therefore, that the elevated rates ofextinction observed during this third timeperiod were not the consequence of localenvironmental vegetation or climatic changesassociated with the Messinian Salinity Crisis.Rather, it is feasible that the elevated extinc-tion rates were the response of the largemammal fauna to geographic isolation of theIberian Peninsula, the intensification of cli-matic gradients across the Mediterraneanregion, and the progressive aridification thatbegan at around 9.0 Ma and culminatedduring the Messinian Salinity Crisis. Thetrophic structure of herbivorous mammalsindicates a progressive increase in openhabitats from intervals 8.75 Ma to 5.75 Ma,as indicated by the increasing proportion ofgrazing species and decreasing proportion ofbrowsing species (Fig. 6). This trend is
MIOCENE MAMMAL DIVERSIFICATION 215

consistent with the global cooling and dryingtrends during the late Miocene that are alsoevident in the increased hypsodonty of Eur-asian ungulates (Eronen et al. 2009, 2012) andin the progressive replacement of warm-temperate evergreen broadleaf and mixed-forest biomes by cooler and drier biomes(Pound et al. 2012).
Finally, the transition from the latest Mio-cene to the early Pliocene seems to have been aperiod of faunal replacement, because morethan 87% of the early Pliocene species from theIberian Peninsula were new (Azanza et al.2000; Agustı et al. 2001). The same patterncharacterizes faunas of the northeastern Med-iterranean, where the Pikermian chronofauna,dominated by open-country mammals, de-clined at the Miocene/Pliocene boundary(Eronen et al. 2009). Nevertheless, it is notablethat, analyzed at the generic level, faunas fromSpain had fairly high continuity, with up to68% of the late Miocene genera persisting intothe early Pliocene. In terms of the geographicorigin of the new Pliocene species from theIberian Peninsula, 75% made their first ap-pearance in the Iberian Peninsula or simulta-neously in the Iberian Peninsula and France orItaly (Supplementary Table 10). Thus, it seemsthat the appearance of many of the IberianPliocene species resulted from the evolution ofMiocene ancestors in this region. The appear-ance of new species has been associated withhumid conditions that developed at thebeginning of the Pliocene and culminated inthe mid-Pliocene (Pliocene Warm Period) as aconsequence of restricted ocean circulationbetween the Pacific and the Atlantic oceansin response to the shoaling of the CentralAmerican seaway (Haug et al. 2001). Asignificant shift toward lower d13C values inthe enamel of Iberian herbivores and anincrease in mean annual precipitation of~400 mm/yr between the latest Miocene andearly Pliocene (MN13 to MN14) also reflectedvegetation growing under the more humidconditions of the Pliocene (Domingo et al.2013).
Each of these periods of diversification hada unique pattern of macroevolutionary changeand a different set of inferred causes. The firstperiod, with high origination rates and in-
crease in regional diversity but no change introphic structure or size structure, involvedarrival and establishment of immigrants(some of them long-distance immigrants), inthe absence of major barriers to dispersal.Intercontinental dispersal and global cooling,rather than regional environmental changeswithin the Iberian Peninsula, are the inferreddrivers of this interval of change. The secondperiod involved significant rates of change inall of the diversity metrics, as well as changesin trophic structure and size structure. Newlyappearing lineages were in part immigrantsfrom eastern Europe and Asia, and substantialchanges in regional climate and vegetationwere evident from multiple lines of evidence.The third period of diversification featuredhigh extinction rates and decrease in regionaldiversity without significant change in taxo-nomic composition or ecological structure.This period coincided with strong regionalendemism and evidence of tectonic barriers todispersal, as well as longer-term intensifica-tion of climatic gradients across the world. It isintriguing that the most profound period offaunal change (the second one in this study)was associated with clear indications ofchange within the terrestrial ecosystems ofthe Iberian Peninsula, whereas the other twoperiods were associated with tectonic andenvironmental drivers outside the IberianPeninsula.
Conclusion
The high levels of completeness and taxo-nomic resolution of the Spanish Miocene largemammal fossil record allowed us to analyzethe tempo and mode of faunal changes in thecontext of global and regional climatic andtectonic history. With ordination and ageestimation of fossil sites, we established afiner chronology of faunal change than by theMN timescale. The fact that observed tempo-ral ranges of fossil taxa typically underesti-mate their actual duration was addressed withthe estimation of confidence intervals and thecorresponding inferred temporal ranges. Adirect consequence of using confidence inter-vals was that the sequence and timing ofappearances and disappearances of specieschanged and the inferred ranges of some
216 M. SOLEDAD DOMINGO ET AL.

species departed from the traditional view.Ideally, confidence intervals should be imple-mented in the fossil record of other regions inorder to construct more accurate and compa-rable rates of origination and extinction, andgeographic range shifts of mammals withinand between continents.
Using this new framework, we analyzedchanges in species diversity, taxonomic com-position, and ecological structure for the largemammals of Spain between 12.0 to 5.5 Ma.Our results highlight three time intervalswhen diversity metrics have significant valuesseparated by periods when diversity remainedstable.
1. From 12.0 to 10.5 Ma, faunal change wasdriven mainly by elevated originationrates. This was a period of mobility of thelarge mammals related to a global coolingpulse. Extinction rates were low anddiversity increased. Immigrants becameestablished in the Iberian Peninsula with-out significant displacement of the residentlarge mammal species or guilds.
2. From 9.5 to 7.5 Ma, faunal change wascharacterized by high extinction ratesfollowed by high origination rates. Thebeginning of this interval represents theVallesian Crisis. In contrast to previousstudies, our analysis implied that theperiod of faunal change was prolongedwith sequential episodes of extinction andorigination. Substantial turnover in faunalcomposition and ecological structure andthe disappearance of genera and evenentire families indicate substantial replace-ment of the fauna. This turnover washighlighted by the clear separation offaunas into two time intervals: an olderfauna (from intervals 11.75 to 9.25 Ma) witha higher proportion of species from moreforested environments, including higherproportions of frugivores and browsers,and a younger fauna (from intervals 8.75 to5.75 Ma) with increasing proportions ofgrazing species in open woodland.
3. From 6.5 to 5.5 Ma, high extinction rates,especially at interval 5.75 Ma, led tosignificant negative diversification andhigh turnover rates and the lowest diver-
sity of the entire analysis interval. Al-though origination rates were very low,they were probably underestimated, and,in fact, could have been elevated. Althoughthis period of change coincided with theMessinian Salinity Crisis, marine desicca-tion was apparently not the driver of faunalchange. In fact, the Messinian SalinityCrisis did not significantly modify theterrestrial environmental conditions of theIberian Peninsula. During this time, theIberian mammal fauna became highlyendemic, possibly as a consequence ofrapid Alpine uplift that imposed barriersto dispersal across the MediterraneanBasin. We suggest that the high extinctionrates observed at this time were a responseto geographic isolation due to tectonicbarriers and intensification of latitudinalclimatic gradients and aridification thatwere underway since 9.0 Ma. In summary,both marine desiccation and faunal chan-ges, were consequence of the same regionaltectonic and climatic episodes, but there isno causal relationships between them. Theelevated number of late Miocene disap-pearances and Pliocene appearances repre-sent evolutionary turnover in situ, assuggested by the high continuity of generaand endemism into the early Pliocene.
Acknowledgments
We thank J. Morales and L. Domingo forproviding information about the taxonomiccomposition, trophic structure, and size struc-ture of the species included in this study. Wethank J. Finarelli for sharing his bootstraproutine. Paleobiology Editor Bruce MacFad-den and two anonymous reviewers providedconstructive comments on the manuscript.This study was funded by the SpanishMinisterio de Economıa y Competitividad(MINECO) project CGL2010-19116/BOS toMTA. M.S.D. was supported by postdoctoralfellowships from the Fundacion Espanola parala Ciencia y la Tecnologıa (FECYT-MINECO),from a National Science Foundation grant(EAR-0957992) to C.B. and from MINECOproject CGL2009-09000. D.D.M. acknowledgesgrant JCI-2011-11697 (MINECO).
MIOCENE MAMMAL DIVERSIFICATION 217

Literature Cited
Agustı, J., and M. Anton. 2002. Mammoths, sabertooths, and
hominids: 65 million years of mammalian evolution in Europe.
Columbia University Press, New York.
Agustı, J., and S. Moya-Sola. 1990. Mammal extinctions in the
Vallesian (Upper Miocene). Lecture Notes in Earth Science
30:425–432.
Agustı, J., L. Cabrera, M. Garces, and M. Llenas. 1999a. Mammal
turnover and global climate change in the late Miocene
terrestrial record of the Valles-Penedes Basin (NE Spain). Pp.
397–412 in Agustı et al. 1999b.
Agustı, J., L. Rook, and P. Andrews, eds. 1999b. The evolution of
Neogene terrestrial ecosystems in Europe. Cambridge Univer-
sity Press, Cambridge.
Agustı, J., L. Cabrera, M. Garces, W. Krijgsman, O. Oms, and J. M.
Pares. 2001. A calibrated mammal scale for the Neogene of
Western Europe: state of the art. Earth-Science Reviews 52:247–
260.
Agustı, J., A. Sanz de Siria, and M. Garces. 2003. Explaining the
end of the hominoid experiment in Europe. Journal of Human
Evolution 45:145–153.
Agustı, J., M. Garces, and W. Krijgsman. 2006. Evidence for
African-Iberian exchanges during the Messinian in the Spanish
mammalian record. Palaeogeography, Palaeoclimatology, Palae-
oecology 238:5–14.
Alba, D. M., J. Agustı, and S. Moya-Sola. 2001. Completeness of
the mammalian fossil record in the Iberian Neogene. Paleobi-
ology 27:79–83.
Alberdi, M. T., B. Azanza, E. Cerdeno, and J. L. Prado. 1997.
Similarity relationship between mammal faunas and biochro-
nology from Latest Miocene to Pleistocene in the Western
Mediterranean area. Eclogae Geologicae Helvetiae 90:115–132.
Alroy, J. 2000. New methods for quantifying macroevolutionary
patterns and processes. Paleobiology 26:707–733.
Azanza, B., M. T. Alberdi, and J. L. Prado. 2000. Large mammal
turnover pulses correlated with latest Neogene glacial trends in
the northwestern Mediterranean region. In M. B. Hart, ed.
Climates: Past and Present. Geological Society, London (Special
Publication 181):161–170.
Badgley, C., and J. Finarelli. 2013. Diversity dynamics of mammals
in relation to tectonic and climatic history: comparison of three
Neogene records from North America. Paleobiology 39:373–399.
Badgley, C., J. C. Barry, M. E. Morgan, S. V. Nelson, A. K.
Behrensmeyer, T. E. Cerling, and D. Pilbeam. 2008. Ecological
changes in Miocene mammalian record show impact of
prolonged climatic forcing. Proceedings of the National Acad-
emy of Sciences USA 105:12145–12149.
Barry, J. C., M. E. Morgan, L. J. Flynn, D. Pilbeam, A. K.
Behrensmeyer, S. M. Raza, I. A. Khan, C. Badgley, J. Hicks, and
J. Kelley. 2002. Faunal and environmental change in the late
Miocene Siwaliks of northern Pakistan. Paleobiology Memoir 3.
Paleobiology 28(Suppl. to No. 2):1–71.
Bernor, R. L., G. D. Koufos, M. O. Woodburne, and M. Fortelius.
1996. The evolutionary history and biochronology of European
and Southwest Asian Late Miocene and Pliocene Hipparionine
horses. Pp. 137–154 in R. L. Bernor, V. Fahlbusch and H. W.
Mittman, eds. The Evolution of Western Eurasian Neogene
mammal faunas. Columbia University Press, New York.
Bernor, R. L., H. Tobien, L.-A. Hayek, and H.-W. Mittmann. 1997.
The Howenegg Hipparionine horses: systematics, stratigraphy,
taphonomy and paleoenvironmental context. Andrias 10:1–230.
Bonis, L. de, and G. D. Koufos. 1999. The Miocene large mammal
succession in Greece. Pp. 205–237 in Agustı et al. 1999b.
Brown, R. P., N. M. Suarez, and J. Pestano. 2002. The Atlas
Mountains as a biogeographical divide in North-West Africa:
evidence from mtDNA evolution in the Agamid lizard Agama
impalearis. Molecular Phylogenetics and Evolution 24:324–332.
Casanovas-Vilar, I., and J. Agustı. 2007. Ecogeographical stability
and climate forcing in the Late Miocene (Vallesian) rodent
record of Spain. Palaeogeography, Palaeoclimatology, Palae-
oecology 248:169–189.
Casanovas-Vilar, I., S. Moya-Sola, J. Agustı, and M. Kohler. 2005.
The geography of a faunal turnover: tracking the Vallesian
Crisis. Pp. 247–301 in A. T. Elewa, ed. Migration of organisms:
climate, geography, ecology. Springer, Heidelberg.
Casanovas-Vilar, I., I. Garcıa-Paredes, D. M. Alba, L. W. van den
Hoek Ostende, and S. Moya-Sola. 2010. The European Far West:
Miocene mammal isolation, diversity and turnover in the
Iberian Peninsula. Journal of Biogeography 37:1079–1093.
Casanovas-Vilar, I., D. M. Alba, M. Garces, J. M. Robles, and S.
Moya-Sola. 2011. Updated chronology for the Miocene homi-
noid radiation in Western Eurasia. Proceedings of the National
Academy of Sciences USA 108:5554–5559.
Casanovas-Vilar, I., L. W. van den Hoek Ostende, M. Furio, and A.
Madern. 2012. Patterns as pretty as can be: the range and extent
of the Vallesian Crisis (Late Miocene) in the Valles-Penedes
Basin (Catalonia, Spain). Journal of Vertebrate Paleontology
32(Suppl.):75.
Cerling, T. E., J. M. Harris, B. J. MacFadden, M. G. Leakey, J.
Quade, V. Eisenmann, and J. R. Ehleringer. 1997. Global
vegetation change through the Miocene/Pliocene boundary.
Nature 389:153–158.
Costeur, L., and S. Legendre. 2008. Spatial and temporal variation
in European Neogene large mammals diversity. Palaeogeogra-
phy, Palaeoclimatology, Palaeoecology 261:127–144.
Costeur, L., S. Montuire, S. Legendre, and O. Maridet. 2007. The
Messinian Event: what happened to the peri-Mediterranean
mammalian communities and local climate? Geobios 40:423–
431.
Daams, R., A. J. van der Meulen, M. A. Alvarez Sierra, P. Pelaez-
Campomanes, and W. Krijgsman. 1999. Aragonian stratigraphy
reconsidered, and a re-evaluation of the middle Miocene
mammal biochronology in Europe. Earth and Planetary Science
Letters 165:287–294.
DeMiguel, D. 2009. Morfologıa funcional y biomecanica de la
denticion en rumiantes (Mammalia, Artiodactyla). Aplicacion
del desgaste dentario en la reconstruccion paleoambiental del
Mioceno de la Cordillera Iberica. Ph.D. thesis. Universidad de
Zaragoza, Zaragoza, Spain.
DeMiguel, D., B. Azanza, and J. Morales. 2011. Paleoenvironments
and paleoclimate of the Middle Miocene of central Spain: a
reconstruction from dental wear of ruminants. Palaeogeogra-
phy, Palaeoclimatology, Palaeoecology 302:452–463.
Domingo, L., P. L. Koch, M. Hernandez Fernandez, D. L. Fox, M. S.
Domingo, and M. T. Alberdi. 2013. Late Neogene and early
Quaternary paleoenvironmental and paleoclimatic conditions in
Southwestern Europe: isotopic analyses on mammalian taxa.
PLoS ONE 8(5): e63739. doi:10.1371/journal.pone.0063739.
Domingo, M. S., M. T. Alberdi, and B. Azanza. 2007. A new
quantitative biochronological ordination for the Upper Neogene
mammalian localities of Spain. Palaeogeography, Palaeoclima-
tology, Palaeoecology 255:361–376.
Edwards, A. W. F. 1992. Likelihood, expanded ed. Johns Hopkins
University Press, Baltimore.
Eronen, J. T., M. M. Ataabadi, A. Micheels, A. Karme, R. L. Bernor,
and M. Fortelius. 2009. Distribution history and climatic
controls of the Late Miocene Pikermian chronofauna. Proceed-
ings of the National Academy of Sciences USA 106:11,867–
11,871.
Eronen, J. T., M. Fortelius, A. Micheels, F. T. Portmann, K.
Puolamaki, and C. M. Janis. 2012. Neogene aridification of the
Northern Hemisphere. Geology 40:823–826.
218 M. SOLEDAD DOMINGO ET AL.

Fauquette, S., J.-P. Suc, A. Bertini, S.-M. Popescu, S. Warny, N. B.
Taoufiq, M.-J. Perez Villa, H. Chikhi, N. Feddi, D. Sullaby, G.
Clauzon, and J. Ferrier. 2006. How much did climate force the
Messinian Salinity Crisis? Quantified climatic conditions from
pollen records in the Mediterranean Region. Palaeogeography,
Palaeoclimatology, Palaeoecology 238:281–301.
Finarelli, J. A., and C. Badgley. 2010. Diversity dynamics of
Miocene mammals in relation to the history of tectonism and
climate. Proceedings of the Royal Society of London B 277:2721–
2726.
Foote, M. 2000. Origination and extinction components of
taxonomic diversity: general problems. Paleobiology 26:74–102.
Fortelius, M., coord. 2012. New and Old Worlds database of fossil
mammals (NOW). University of Helsinki. http://www.helsinki.
fi/science/now/ (accessed January 2012).
Fortelius, M., J. Eronen, L. Liu, D. Pushkina, A. Tesakov, I.
Vislobokova, and Z. Zhang. 2006. Late Miocene and Pliocene
large land mammals and climatic changes in Eurasia. Palae-
ogeography, Palaeoclimatology, Palaeoecology 238:219–227.
Franzen, J. L., and G. Storch. 1999. Late Miocene mammals from
Central Europe. Pp. 165–190 in Agustı et al. 1999b.
Gibert, L., G. R. Scott, P. Montoya, F. J., Ruiz-Sanchez, J. Morales,
L. Luque, J. Abella, and M. Lerıa. 2013. Evidence for an African-
Iberian mammal dispersal during the pre-evaporitic Messinian.
Geology 41:691–694.
Gomez, F., W. Beauchamp, and M. Barazangi. 2000. Role of the
Atlas Mountains (northwest Africa) within the African-Eurasian
plate-boundary zone. Geology 28:775–778.
Gomez Cano, A. R., M. Hernandez Fernandez, and M. A. Alvarez-
Sierra. 2011. Biogeographic provincialism in rodent faunas from
the Iberoccitanian Region (southwestern Europe) generates
severe diachrony within the Mammalian Neogene (MN)
biochronologic scale during the Late Miocene. Palaeogeography,
Palaeoclimatology, Palaeoecology 307:193–204.
Gomez Cano, A. R., J. L. Cantalapiedra, A. Mesa, A. Moreno
Bofarull, and M. Hernandez Fernandez. 2013. Global climate
changes drive ecological specialization of mammal faunas:
trends in rodent assemblages from the Iberian Plio-Pleistocene.
BMC Evolutionary Biology 13:94.
Gradstein, F., J. Ogg, and A. Smith. 2004. A geologic time scale
2004. Cambridge University Press, Cambridge.
Hammer, Ø., and D. Harper. 2006. Paleontological data analysis.
Blackwell, Malden, Mass.
Haug, G. H., R. Tiedemann, R. Zahn, and A. C. Ravelo. 2001. Role
of Panama uplift on oceanic freshwater balance. Geology
29:207–210.
Hernandez Fernandez, M., B. Azanza, and M. A. Alvarez Sierra.
2004. Iberian Plio-Pleistocene biochronology: micromammalian
evidence for MNs and ELMAs calibration in southwestern
Europe. Journal of Quaternary Science 19:605–616.
Hernandez Fernandez, M., M. T. Alberdi, B. Azanza, P. Montoya,
J. Morales, M. Nieto, and P. Pelaez-Campomanes. 2006.
Identification problems of arid environments in the Neogene-
Quaternary mammal record of Spain. Journal of Arid Environ-
ments 66:585–608.
Hsu, K. J., W. B. F. Ryan, and M. B. Cita. 1973. Late Miocene
desiccation of the Mediterranean. Nature 242:240–244.
Jimenez-Moreno, G., S. Fauquette, and J.-P. Suc. 2010. Miocene to
Pliocene vegetation reconstruction and climate estimates in the
Iberian Peninsula from pollen data. Review of Paleobotany and
Palynology 162:403–415.
Koch, C. F. 1987. Prediction of sample size effects on the measured
temporal and geographic distribution patterns of species.
Paleobiology 13:100–107.
Krijgsman, W., F. J. Hilgen, I. Raffi, F. J. Sierro, and D. S. Wilson.
1999. Chronology, causes and progression of the Messinian
Salinity Crisis. Nature 400:652–655.
Kuhlemann, J. 2007. Paleogeographic and paleotopographic
evolution of the Swiss and Eastern Alps since the Oligocene.
Global and Planetary Change 58:224–236.
Lunkka, J. P., M. Fortelius, J. Kappelman, and S. Sen. 1999.
Chronology and mammal faunas of the Miocene Sinap
Formation, Turkey. Pp. 238–264 in Agustı et al. 1999b.
Maas, M. C., M. R. L. Anthony, P. D. Gingerich, G. Gunnell, and D.
W. Krause. 1995. Mammalian generic diversity and turnover in
the Late Paleocene and Early Eocene of the Bighorn and Crazy
Mountains Basins, Wyoming and Montana (USA). Palaeogeog-
raphy, Palaeoclimatology, Palaeoecology 115:181–207.
Mariotti, A., M. V. Struglia, N. Zeng, and K.-M. Lau. 2002. The
hydrological cycle in the Mediterranean Region and implica-
tions for the water budget of the Mediterranean Sea. Journal of
Climate 15:1674–1690.
Marmi, J., I. Casanovas-Vilar, J. M. Robles, S. Moya-Sola, and D.
Alba. 2012. The paleoenvironment of Hispanopithecus laietanus as
revealed by paleobotanical evidence from the Late Miocene of
Can Llobateres 1 (Catalonia, Spain). Journal of Human
Evolution 62:412–423.
Marshall, C. R. 1990. Confidence intervals on stratigraphic ranges.
Paleobiology 16:1–10.
———. 1994. Confidence intervals on stratigraphic ranges: partial
relaxation of the assumption of randomly distributed fossil
horizons. Paleobiology 20:459–469.
———. 1997. Confidence intervals on stratigraphic ranges with
nonrandom distributions of fossil horizons. Paleobiology
23:165–173.
———. 2010. Using confidence intervals to quantify the uncer-
tainty on the end-points of stratigraphic ranges. In J. Alroy and
G. Hunt, eds. Quantitative methods in paleobiology. Paleonto-
logical Society Papers 16:291–316.
Matson, S. D., and D. L. Fox. 2010. Stable isotopic evidence for
terrestrial latitudinal climate gradients in the Late Miocene of
the Iberian Peninsula. Palaeogeography, Palaeoclimatology,
Palaeoecology 287:28–44.
Mein, P. 1975. Resultats du groupe de travail des vertebres:
biozonation du Neogene mediterraneen a partir des
mammiferes. Pp. 77–81 in J. Senes, ed. Report on activity of
the Regional Committee on Mediterranean Neogene Stratigra-
phy, Bratislava.
Morales, J., M. Nieto, M. Kohler, and S. Moya-Sola. 1999. Large
mammals from the Vallesian of Spain. Pp. 113–126 in Agustı et
al. 1999.
Paul, C. R. C. 1982. The adequacy of the fossil record. In K. A.
Joysey and A. E. Friday, eds. Problems of phylogenetic
reconstruction. Systematics Association Special Volume 21:75–
117. Academic Press, London.
Pound, M. J., A. M. Haywood, U. Salzmann, and J. B. Riding. 2012.
Global vegetation dynamics and latitudinal temperature gradi-
ents during the Mid to Late Miocene (15.97–5.33 Ma). Earth-
Science Reviews 112:1–22.
Rouchy, J. M., and A. Caruso. 2006. The Messinian salinity crisis in
the Mediterranean basin: a reassessment of the data and an
integrated scenario. Sedimentary Geology 188–189:35–67.
Signor, P. W., and J. H. Lipps. 1982. Sampling bias, gradual
extinction patterns and catastrophes in the fossil record. In L. T.
Silver and P. H. Schultz, eds. Geological implications of impacts
of large asteroids and comets on the Earth. Geological Society of
America Special Paper 190:291–296.
Strauss, D., and P. M. Sadler. 1989. Classical confidence intervals
and Bayesian probability estimates for the ends of local taxon
ranges. Mathematical Geology 21:411–427.
Stromberg, C. A. E. 2011. Evolution of grasses and grassland
ecosystems. Annual Review of Earth and Planetary Sciences
39:517–544.
MIOCENE MAMMAL DIVERSIFICATION 219

Tedford, R. H., L. B. Albright III, A. D. Barnosky, I. Ferrusquia-
Villafranca, R. M. Hunt Jr., J. E. Storer, C. C. Swisher III, M. R.
Voorhies, S. D. Webb, and D. P. Whistler. 2004. Mammalian
biochronology of the Arikareean through Hemphillian interval
(Late Oligocene through Early Pliocene epochs). Pp. 169–231 in
M. O. Woodburne, ed. Columbia University Press, New York.
van Dam, J. A., L. Alcala, A. Alonso Zarza, J. P. Calvo, M. Garces,
and W. Krijgsman. 2001. The Upper Miocene mammal record
from the Teruel-Alfambra region (Spain). The MN system and
continental stage/age concepts discussed. Journal of Vertebrate
Paleontology 21:367–385.
van Dam, J. A., H. A. Aziz, M. A. Alvarez Sierra, F. J. Hilgen, L. W.
van den Hoek Ostende, L. J. Lourens, P. Mein, A. J. van der
Meulen, and P. Pelaez-Campomanes. 2006. Long-period astro-
nomical forcing of mammal turnover. Nature 443:687–691.
van der Made, J., J. Morales, and P. Montoya. 2006. Late Miocene
turnover in the Spanish mammal record in relation to palae-
oclimate and the Messinian Salinity Crisis. Palaeogeography,
Palaeoclimatology, Palaeoecology 238:228–246.
van der Meulen, A. J., P. Pelaez-Campomanes, and S. A. Levin.
2005. Age structure, residents, and transients of Miocene rodent
communities. American Naturalist 165:108–125.
Wust, G. 1961. On the vertical circulation of the Mediterranean
Sea. Journal of Geophysical Research 66:3261–3271.
Zachos, J., M. Pagani, L. Sloan, E. Thomas, and K. Billups. 2001.
Trends, rhythms, and aberrations in global climate 65 Ma to
present. Science 292:686–693.
220 M. SOLEDAD DOMINGO ET AL.
Copyright © 2022 FDOKUMEN



















