
A gene-family encoding small exported proteins is conserved across Plasmodium genus
10
Molecular & Biochemical Parasitology 126 (2003) 209–218 A gene-family encoding small exported proteins is conserved across Plasmodium genus Cecilia Birago, Veronica Albanesi, Francesco Silvestrini, Leonardo Picci, Elisabetta Pizzi, Pietro Alano, Tomasino Pace, Marta Ponzi ∗ Laboratorio di Biologia Cellulare, Istituto Superiore di Sanità, Viale Regina Elena 299, 00161 Rome, Italy Received 3 June 2002; received in revised form 16 September 2002; accepted 18 October 2002 Abstract A gene-family, named sep, encoding small exported proteins conserved across Plasmodium species has been identified. SEP proteins (13–16 kDa) contain a predicted signal peptide at the NH 2 -terminus, an internal hydrophobic region and a polymorphic, low-complexity region at the carboxy-terminus. One member of the Plasmodium berghei family, Pbsep1, encodes an integral membrane protein expressed along the entire erythrocytic cycle. Immunolocalisation results indicated that PbSEP1 is targeted to the membrane of the parasitophorous vacuole up to the early phases of schizogony, while, in late schizonts, it re-locates in structures within the syncitium. After erythrocyte rupture, PbSEP1 is still detectable in free merozoites thus suggesting its involvement in the early steps of parasite invasion. Seven members of the sep-family in Plasmodium falciparum have been identified. Two of them correspond to previously reported gene sequences included in a family of early transcribed membrane proteins (etramp). Structural, functional and phylogenetic features of the sep family, shown in the present work, supercede this previous classification. PfSEP proteins are exported beyond the parasite membrane and translocated, early after invasion, to the host cell compartment in association with vesicle-like structures. Colocalisation results indicated that PfSEP-specific fluorescence overlaps, at the stage of trophozoite, with that of Pf332, a protein associated with Maurer’s clefts, membranous structures in the cytosol of parasitised red blood cells, most probably involved in trafficking of parasite proteins. The specific signals necessary to direct SEP proteins to the vacuolar membrane in P. berghei or to the host cell compartment in P. falciparum remain to be determined. © 2002 Elsevier Science B.V. All rights reserved. Keywords: Plasmodium falciparum; Plasmodium berghei; Gene-family; Exported proteins 1. Introduction The molecular mechanisms by which Plasmodium para- site gains entry and develops within the red blood cell are the object of extensive investigation as are parasite or host proteins involved in these processes. Early after merozoite invasion, Plasmodium introduces morphological and bio- chemical alterations within the host cell [1]. A second mem- brane, distinct from the plasma membrane, surrounds the parasite, delimiting a new compartment, the parasitophorous Abbreviations: DAPI, 4,6-diamino-2-phenylindole; FITC, fluorescein isothiocyanate; h.p.i., hours post infection; IFA, indirect immunofluores- cence assay; MBCD, methyl--cyclodextrin; PV, parasitophorous vacuole; PVM, parasitophorous vacuolar membrane; TRITC, tetramethylrhodamine isothiocyanate; TVN, tubo-vesicular network Note: Nucleotide sequence data reported in this paper are available in the EMBL database under the accession numbers: AJ420905; AJ420906; AJ420907. ∗ Corresponding author. Tel.: +39-06-49902226; fax: +39-06-49387143. E-mail address: [email protected] (M. Ponzi). vacuole (PV) [2]. Vacuolar membrane (PVM)—the bio- chemical interface between the host cell cytosol and the parasite surface—is connected to the periphery of the red cell through a complex tubo-vesicular network (TVN) most probably implicated in nutrient transport [3]. Plasmodium is able to target protein molecules, including components of the secretory machinery, beyond its plasma membrane [4–6]. Mature erythrocytes are, in fact, terminally differen- tiated cells that are deficient in de novo protein synthesis and lack the machinery for protein trafficking. Exported proteins have been shown to be targeted to the PV/PVM and to different host cell compartments (reviewed in [7,8]). Membranous structures exported to the erythrocyte cytosol and referred to as Maurer’s clefts, seem to play a role in the transport and assembly of parasite proteins targeted to the host cell membrane or cytoskeleton [9,10]. A gene-family encoding small exported proteins (sep), conserved across Plasmodium genus, was characterised in this study. SEP proteins (13–16 kDa) share the presence of a predicted signal peptide at the NH 2 terminus, an internal hy- drophobic region and a polymorphic, low complexity region 0166-6851/02/$ – see front matter © 2002 Elsevier Science B.V. All rights reserved. PII:S0166-6851(02)00275-X
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A gene-family encoding small exported proteins is conserved across Plasmodium genus

Molecular & Biochemical Parasitology 126 (2003) 209–218
A gene-family encoding small exported proteins is conservedacrossPlasmodiumgenus�
Cecilia Birago, Veronica Albanesi, Francesco Silvestrini, Leonardo Picci,Elisabetta Pizzi, Pietro Alano, Tomasino Pace, Marta Ponzi∗
Laboratorio di Biologia Cellulare, Istituto Superiore di Sanità, Viale Regina Elena 299, 00161 Rome, Italy
Received 3 June 2002; received in revised form 16 September 2002; accepted 18 October 2002
Abstract
A gene-family, namedsep, encoding small exported proteins conserved acrossPlasmodiumspecies has been identified. SEP proteins(13–16 kDa) contain a predicted signal peptide at the NH2-terminus, an internal hydrophobic region and a polymorphic, low-complexityregion at the carboxy-terminus. One member of thePlasmodium bergheifamily, Pbsep1, encodes an integral membrane protein expressedalong the entire erythrocytic cycle. Immunolocalisation results indicated that PbSEP1 is targeted to the membrane of the parasitophorousvacuole up to the early phases of schizogony, while, in late schizonts, it re-locates in structures within the syncitium. After erythrocyterupture, PbSEP1 is still detectable in free merozoites thus suggesting its involvement in the early steps of parasite invasion. Seven membersof thesep-family in Plasmodium falciparumhave been identified. Two of them correspond to previously reported gene sequences includedin a family of early transcribed membrane proteins (etramp). Structural, functional and phylogenetic features of thesepfamily, shown inthe present work, supercede this previous classification. PfSEP proteins are exported beyond the parasite membrane and translocated, earlyafter invasion, to the host cell compartment in association with vesicle-like structures. Colocalisation results indicated that PfSEP-specificfluorescence overlaps, at the stage of trophozoite, with that of Pf332, a protein associated with Maurer’s clefts, membranous structures inthe cytosol of parasitised red blood cells, most probably involved in trafficking of parasite proteins. The specific signals necessary to directSEP proteins to the vacuolar membrane inP. bergheior to the host cell compartment inP. falciparumremain to be determined.© 2002 Elsevier Science B.V. All rights reserved.
Keywords: Plasmodium falciparum; Plasmodium berghei; Gene-family; Exported proteins
1. Introduction
The molecular mechanisms by whichPlasmodiumpara-site gains entry and develops within the red blood cell arethe object of extensive investigation as are parasite or hostproteins involved in these processes. Early after merozoiteinvasion, Plasmodiumintroduces morphological and bio-chemical alterations within the host cell[1]. A second mem-brane, distinct from the plasma membrane, surrounds theparasite, delimiting a new compartment, the parasitophorous
Abbreviations:DAPI, 4,6-diamino-2-phenylindole; FITC, fluoresceinisothiocyanate; h.p.i., hours post infection; IFA, indirect immunofluores-cence assay; MBCD, methyl-�-cyclodextrin; PV, parasitophorous vacuole;PVM, parasitophorous vacuolar membrane; TRITC, tetramethylrhodamineisothiocyanate; TVN, tubo-vesicular network
� Note: Nucleotide sequence data reported in this paper are available inthe EMBL database under the accession numbers: AJ420905; AJ420906;AJ420907.
∗ Corresponding author. Tel.:+39-06-49902226;fax: +39-06-49387143.
E-mail address:[email protected] (M. Ponzi).
vacuole (PV) [2]. Vacuolar membrane (PVM)—the bio-chemical interface between the host cell cytosol and theparasite surface—is connected to the periphery of the redcell through a complex tubo-vesicular network (TVN) mostprobably implicated in nutrient transport[3]. Plasmodiumis able to target protein molecules, including componentsof the secretory machinery, beyond its plasma membrane[4–6]. Mature erythrocytes are, in fact, terminally differen-tiated cells that are deficient in de novo protein synthesisand lack the machinery for protein trafficking. Exportedproteins have been shown to be targeted to the PV/PVMand to different host cell compartments (reviewed in[7,8]).Membranous structures exported to the erythrocyte cytosoland referred to as Maurer’s clefts, seem to play a role inthe transport and assembly of parasite proteins targeted tothe host cell membrane or cytoskeleton[9,10].
A gene-family encoding small exported proteins (sep),conserved acrossPlasmodiumgenus, was characterised inthis study. SEP proteins (13–16 kDa) share the presence of apredicted signal peptide at the NH2 terminus, an internal hy-drophobic region and a polymorphic, low complexity region
0166-6851/02/$ – see front matter © 2002 Elsevier Science B.V. All rights reserved.PII: S0166-6851(02)00275-X

210 C. Birago et al. / Molecular & Biochemical Parasitology 126 (2003) 209–218
at the C-terminus. Threesepgenes were originally identifiedin a cDNA library ofPlasmodium berghei, while, six mem-bers of the family inPlasmodium yoelii, one inPlasmodiumvivax and seven inPlasmodium falciparumwere retrievedfrom databases. Pfsep14 and Pfsep11-2 correspond to tworing-specific cDNAs previously identified[11]. These se-quences were clustered by the authors in a gene-family ofearly transcribed membrane proteins (etramp) that includesa third gene encoding aP. falciparumantigen[12], that doesnot exhibit similarities with SEP proteins.
Immunolocalisation studies indicated that members of thesep-family inP. bergheiand inP. falciparumare expressedduring the entire erythrocytic cycle and are exported beyondthe parasite membrane. The product of Pbsep1 is associatedwith the vacuolar membrane up to the early phases of schizo-gony. With the progression of schizont maturation, PbSEP1re-locates to structures within the syncytium and associateswith free merozoites after erythrocyte rupture. Members ofthesep-family in P. falciparum, are, instead, trafficked, earlyafter invasion, to the red blood cell compartment in associa-tion with Maurer’s clefts. Signals and mechanisms involvedin sorting and localisation of these proteins are discussed.
2. Materials and methods
2.1. Parasites
P. bergheiANKA 8417HP[13] and HPE[14] clones weremaintained in Swiss mice. Synchronous infections and pu-rification of blood stages were performed as described[15].P. falciparum3D7 parasites were cultivated under standardconditions[16].
2.2. Isolation of P. berghei sep1-3 genes from genomic andcDNA libraries
Pbsep1 gene was originally identified in a subtractionexperiment as a sequence abundantly transcribed in theP.bergheiANKA clone 8417HP and deleted in a cloned lineof P. bergheiK173 [17]. The complete sequence of the genewas obtained by contig assembly after screening a partialSau3A genomic library of the clone 8417HP[18]. Pbsep2and 3 genes were identified by screening the same genomiclibrary using the coding region of Pbsep1 as a probe. Thetranscribed sequences of the threesepgenes were obtainedfrom a cDNA library of asynchronous blood stages of theclone 8417HP[19].
2.3. Database similarity searches and sequence analysis
Putative SEP proteins inP. falciparum, P. yoeliiandP. vi-vaxwere identified by searching organism specific sequenceBLAST databases available athttp://www.ncbi.nlm.nih.gov/Malaria. Deduced amino acid sequences were re-namedas members of SEP family. Entries and nucleotide se-
quences corresponding to each putative gene are listedbelow: PySEP1 (TIGRc5m1485: 8116–7592 bp), PySEP2(TIGR c5m1153:1083–1553 bp), PySEP3 (TIGRc5m55:20254–19730 bp), PySEP4 (TIGR669Plasmodium), Py-SEP5 (TIGRc5m1076: 1863–1522 bp), PySEP6 (TIGRc5-m2205:1338–919 bp), PvSEP1 (P. vivax GSS: UFL329PvD09:138–524 bp). Entries corresponding toP. fal-ciparum SEP proteins are listed according to PlasmoDBGlimmer prediction: PfSEP14 (chr141.glm 18), PfSEP12(chr121.glm 452) PfSEP11.1 (chr111.glm 46), PfSEP11.2(chr.111.glm 45), PfSEP10 (chr101.glm 31), PfSEP4(chr4 3 glm 21), PfSEP2 corresponds to gene PFB0120won chromosome 2. The data base search was done usingas a query sequence the first 74 aa, corresponding to theN-terminal region conserved among SEP proteins inP.berghei. Multiple alignment of the putative SEP memberswas performed by CLUSTALW (http://www.ebi.ac.uk) andrefined by hand. SignalP and HTMM prediction servers(http://www.cbs.dtu.dk) were used to identify putativesignal-peptides and transmembrane domains respectively.
2.4. RNA extraction and Northern blot analysis
RNA from P. bergheiand P. falciparum parasites wasisolated using Rneasy kit according to the manufacturer’sinstructions (Qiagen). Total RNA size fractionated on 1%agarose under denaturing conditions was transferred toAmersham Hybond-C extra membrane and hybridised un-der standard conditions.
2.5. Primers, PCR probes and sequencing
Primers listed inTable 1were obtained from Roche cus-tom service and used to amplify DNA fragments specificfor P. falciparumandP. berghei sepgenes. PCR amplifica-tions were performed under the following conditions: 1 minat 90◦C, 1 min at 55◦C, and 1 min at 72◦C for 30 cycles.PCR products were cloned in pGEM-T vector (Promega)and sequenced using a Sequenase 2.0 DNA sequencing Kit(Amersham Pharmacia Biotech).
2.6. Immune sera
A polyclonal rabbit antiserum prepared against aKLH-conjugated peptide corresponding to the C-terminalportion of PbSEP1 (ASDNDSVPPS) was obtained fromBiogen custom service. Polyclonal rat antisera were pre-pared against BSA-conjugated peptides (Roche custom ser-vice) corresponding to the C-terminal portions of PfSEP2(KNNKGDNKKGT), PfSEP4 (KESKADKSEEK) and Pf-SEP14 (AKPKNTEVKEP). Sprague Dawley rats were im-munised with a first dose (200�g of conjugated peptides)emulsified in complete Freund’s adjuvant followed by oneboost in incomplete adjuvant and three successive boostswithout adjuvant at 2-week intervals. Blood was finallycollected by cardiac puncture. Mab51/22 directed against

C. Birago et al. / Molecular & Biochemical Parasitology 126 (2003) 209–218 211
Table 1Oligonucleotide primers for PCR amplifications
Gene Primer code Nucleotide sequence (5′ → 3′)
P. bergheiPbsep1 SP1-a CTCAGCATCAGATAATGACA
SP1-b TCTGCTTCAATGTAACATAG
Pbsep2 SP2-a CGAATTTATTGTGTATAAGCSP2-b TACATCATATATATGCGGAG
Pbsep3 SP3-a GCACATATCGCTATCAATTACGTSP3-b CATCTTATTAATTTCATTGCT
P. falciparumPfsep2 PF-MEM 5 AGATTACAATAGCATGGTAGA
PF-MEM 6 GGCTCTGTACATGGTACTTGA
Pfsep4 PF-MEM 1 GCATGAACGTTTTCGTTCCAGGAPF-MEM 2 AAACAGTAGTGGCTACTGCTGTA
Pfsep10 PF-MEM 11 CAGGTAGAAAATTTCAAAGAPF-MEM 12 CCAAAACCTACACCAGCACC
Pfsep11-1 PF-MEM 8 TTGGAGTGGCGGTGATGGTTPF-MEM 15 GCTTTAACACCAGCTGAAAAG
Pfsep11-2 PF-MEM 9 GCCTTGAATTTTATTGCCCCPF-MEM 10 GAGAAATGAACTCCTAAACC
Pfsep12 PF-MEM 13 CTCATTGCTGCCTTATTGGCPF-MEM 14 GGTGAGGTGATCTGGAAGCC
Pfsep14 PF-MEM 3 TTATGTTGCTGCCGGATCTAATAPF-MEM 4 AGCTGTTGTGGCTGGGGCTGG
Pf332 [10] was kindly provided by Dr. Denise Mattei,Institut Pasteur, Paris.
2.7. Parasite subcellular extracts
The cytosolic and particulate fractions of 109 P. bergheiparasites were obtained, in the presence of protease in-hibitors, as described[20]. The particulate fraction was fur-ther extracted for 30 min in ice with either 0.1 M Na2CO3,pH 11.5 or 3 M KCl to solubilise peripheral membrane pro-teins. Following incubations, the insoluble material was sed-imented at 24,000× g for 30 min at 4◦C and directly dis-solved in SDS sample buffer.
2.8. Protein immunodetection
Equal amount ofP. bergheiproteins, as determined byBradford assay (BioRad Protein Assay), were separatedby SDS gel electrophoresis and transferred to nitrocellu-lose filter as described[18] and immunodetected usingPbSEP1-specific rabbit serum (1:200 dilution). The speci-ficity of the serum was checked using preimmune serum.
2.9. Indirect immunofluorescence assay (IFA)
Immunofluorescence assays ofP. falciparuminfected ery-throcytes were performed as described[21] on unfixed blood
smears using immune sera specific for PfSEP 2, 4 and 14(1:200 dilution) and anti-Pf332 Mab 51/22 (1:500 dilution).P. bergheiblood smears were fixed in 2% formaldehyde[22]and incubated with PbSEP1-specific rabbit serum (dilution1:3000). The specificity of the immune sera was checked inparallel experiments using preimmune sera.
2.10. Methyl-β-cyclodextrin treatment
Cultures of synchronousP. bergheiparasites were in-cubated with 10 mM of the cholesterol depleting agentmethyl-�-cyclodextrin (MBCD) at 37◦C for 30 min. Asreported[23], the treatment causes the expulsion from thehost cell of trophozoites freed of their vacuolar membraneallowing an efficient separation of PVM associated com-ponents from parasite plasma membrane. The percentageof parasites released for effect of the treatment was estab-lished by microscopic inspection of Giemsa stained bloodsmears. Control cultures in the absence of MBCD weremaintained in parallel. Indirect immunofluorescences oftreated and untreated parasites were performed as describedabove.
3. Results
3.1. Identification and characterisation of a novelgene-family (sep) in Plasmodium berghei
TheP. bergheigenesep1 was identified in the course of acDNA subtraction experiment[17] and the putative codingregion was used as a probe on molecular karyotypes andtotal DNA restriction patterns. Hybridisation results (notshown) suggested the presence of additional homologoussequences in theP. bergheigenome. The screening with thesame probe of a genomic library[18] and cDNA library pre-pared from asynchronous blood-stage RNA[19] led to theidentification of two additional related genes named Pbsep2and 3. Sequence analysis indicated that the threesepgenesdo not contain introns and share the upstream regulatory anduntranslated sequences. In particular, the upstream regionsof Pbsep1 and 3 exhibit 80% identity within 600 bp fromthe start codon, the identity reaching 100% in the 1200 bpfarther upstream. Pbsep1 and 2 share a 930 bp region up-stream the ATG codon with a 98% identity. In addition, thethree genes share the first 74 amino acids of the putativeproteins (13–16 kDa). The C-terminus and the downstreamuntranslated region are polymorphic. Karyotype analysisusing gene-specific probes (listed inTable 1) indicated thatPbsep1–3 are all located on the two terminal ApaI frag-ments of chromosome 5 (Fig. 1a). The threesepgenes aretranscribed in parasite blood stages both in asexuals andin gametocytes. The size of the corresponding mRNAs, asdetermined by Northern blot analysis (Fig. 1b), is 1.5 kbfor Pbsep1 and 2 and 1.8 kb for Pbsep3. A second fainterband at 2.5 kb was observed in the gametocyte-specific

212 C. Birago et al. / Molecular & Biochemical Parasitology 126 (2003) 209–218
Fig. 1. Chromosome location and transcription pattern of threesepgenes identified inP. berghei(Pbsep1–3). (a) A restriction map of the terminal regionof P. bergheichromosome 5 is drawn to scale, telomeric regions are in black. The three Pbsepgenes are positioned and the direction of transcriptionindicated by arrows. (b) Northern blot of total RNA from asexual stages (A) and purified gametocytes (G) hybridised with probes specific for Pbsep1–3.Molecular mass markers (Novagen) are indicated. (c) Northern blot of total RNAs prepared 5–23 h post invasion (h.p.i.) fromP. bergheiclone 8417HPand hybridised in succession with probes specific for Pbsep1–3 genes.
RNA probed with Pbsep2. Stage-specific RNAs from syn-chronous infections were blotted and hybridised in suc-cession with gene-specific probes. As shown inFig. 1c,Pbsep1 and 2 are transcribed at the ring stage, since 5 hpost infection (h.p.i.), and maintained up to mature sch-izonts (23 h.p.i.); Pbsep3-specific mRNA is transcribed inmature trophozoites (16 h.p.i.) and hardly detectable inthe other stages.
3.2. Sep family is conserved across Plasmodium species
The conserved NH2 portion of deduced PbSEP proteinswas used to explore databases made available byPlas-modiumgenome sequencing projects. Six putative proteinswere retrieved inP. yoelii database (PySEP1–6 inFig. 2).The highly similar adjacent intergenic regions identifiedPySEP1–3 as theP. yoeliicounterparts of PbSEP1–3 respec-tively. Seven protein sequences fromP. falciparumdatabasesand one fromP. vivaxwere also retrieved. Multiple align-ment (Fig. 2) evidentiated features conserved within the SEPfamily: a predicted signal peptide at the N-terminus, an inter-nal hydrophobic region and a polymorphic, low-complexityregion at the C-terminus, characterised by a relativelyhigh abundance of prolines. The position of serine andthreonine residues within the predicted transmembrane
domains suggested the presence of a helix face enrichedin polar residues, as revealed by helical wheel diagrams,suggesting for them a role in promoting helix association[24].
In the case ofP. falciparum, chromosome attribution ofthe sepgenes was confirmed by karyotype analysis usingseven gene-specific probes listed inTable 1(not shown).
Northern blot analysis performed on asynchronousP. falciparum parasites (Fig. 3) indicated that this gene-family is actively transcribed in the human parasite, withthe exception ofPfsep12. In the case ofPfsep2 and Pf-sep10 two separate transcripts were revealed at a shorterexposure.
3.3. Subcellular distribution of PbSEP1
In order to investigate the expression pattern of the proteinencoded by Pbsep1 and the cellular compartment(s) whereit locates, a polyclonal rabbit serum against a synthetic pep-tide corresponding to the C-terminal portion of the deducedprotein was used in Western blot experiments. Equal amountof total cellular proteins, prepared from asynchronousP.bergheiclones 8417HP and HPE were analysed. HPE clonewas used as a negative control, as it harbours a terminal dele-tion of chromosome 5[25] that includes Pbsep1. A band of

C.
Birag
oe
ta
l./Mo
lecu
lar
&B
ioch
em
ical
Pa
rasito
logy
12
6(2
00
3)
20
9–
21
8213
Fig. 2. Multiple alignment of SEP proteins from different species ofPlasmodium. Conserved residues among all aligned proteins are highlighted. Red boxes indicate positively charged residues, greenboxes indicate hydrophobic residues and blue boxes evidentiate polar residues. Putative cleavage sites downstream hypothetical signal peptides are indicated (∇). Predicted transmembrane domain isindicated by arrows. Peptides used to generate antisera are underlined.

214 C. Birago et al. / Molecular & Biochemical Parasitology 126 (2003) 209–218
Fig. 3. Northern blot analysis of theP. falciparum sepgenes. Total RNA from 3D7A clone, asynchronously grown, was run in denaturing agarose gelsin the presence of molecular mass markers Novagen (lane b). Blotted RNA was hybridised with gene-specific probes (lane a).
the expected size of 13 kDa present only in the extract ofthe clone 8417HP (Fig. 4A) identified the product of thePbsep1 gene. The predicted structural features of the SEPfamily suggested the association of these proteins withmembranes. In order to investigate this point, soluble andparticulate protein fractions were prepared from purifiedasynchronous parasites of the clone 8417HP. Immunoblotanalysis revealed that PbSEP1 was always associated to theinsoluble material (not shown). This fraction was furtherextracted in high salt (3 M KCl), or alkali (0.1 M Na2CO3,pH 11) to separate peripheral and integral membrane pro-teins. As shown inFig. 4B, PbSEP1 is not solubilisedby these treatments, which suggests that it is an integralmembrane protein.
Fig. 4. Immunodetection of PbSEP1 with a specific polyclonal rabbit immune serum. (A) Equal amount of total protein extracts from asynchronousP.bergheiclone 8417HP and from the deletion mutant HPE lacking Pbsep1 gene. (B) Particulate fraction from 8417HP parasites extracted with 3 M KClor 0.1 M Na2CO3, pH 11 to separate peripheral (soluble fraction, S) and integral membrane proteins (insoluble fraction, P). Arrowheads indicate theprotein band corresponding to PbSEP1. Molecular mass markers (Novagen) are indicated.
3.4. Immunolocalisation of PbSEP1 in blood-stageparasites and effect of the treatment withcholesterol-depleting agent
In order to identify the subcellular localisation of PbSEP1,indirect immunofluorescence assays were conducted onP.bergheiinfections using the specific rabbit polyclonal serum.The specificity of the anti-PbSEP1 antibody was confirmedusing the deletion mutant clone HPE in control IFA assays(data not shown). In synchronousP. bergheiclone 8417HPanti-PbSEP1 antibodies decorated the periphery of parasitesall along the erythrocytic cycle (Fig. 5a) while no significantfluorescence was detected using preimmune serum. Protru-sions and evaginations were stained in parasites at the ring

C. Birago et al. / Molecular & Biochemical Parasitology 126 (2003) 209–218 215
Fig. 5. Immunolocalisation of PbSEP1 in synchronousP. bergheiinfections. Indirect immunofluorescence assay was performed on formaldehyde fixedparasites with PbSEP1-specific rabbit polyclonal serum (dilution 1:3000) revealed with FITC-conjugated anti-rabbit IgG (panel a). The arrow indicatesspecific fluorescence organised in subcompartments at the parasite surface. Synchronous parasites treated with the cholesterol depleting agent MBCD andsubsequently stained with the specific immune serum are shown in panel b. Parasite nuclei are stained with DAPI. Bars represent 5�m.
and trophozoite stages. PbSEP1-specific fluorescence wasalso observed in subcompartments, in a necklace fashion,particularly evident during trophozoite development. The pe-ripheral distribution of PbSEP1 was maintained during theearly phases of schizogony while, in late schizonts, the im-mune serum decorated structures within the syncytium thatsurrounded the dividing parasites. After erythrocyte rupturePbSEP1 was still detected in free merozoites but the cellu-lar compartment where it locates was not determined due tothe poor resolution of the image.
To further investigate the association of PbSEP1 withthe PVM, experiments were performed with the cholesteroldepleting agent methyl-�-cyclodextrin (MBCD). It was pre-viously reported[23] that MBCD added toP. falciparumcul-tures destabilises the PVM, which contains cholesterol-richmicrodomains. As a consequence, trophozoites were re-leased from the infected cells and the specific fluorescenceof EXP1, a PVM-associated protein, was lost or revealed ina punctuate distribution trailing the parasites. Ring infectederythrocytes were, instead, insensitive to the treatment.When synchronousP. bergheiparasites were treated withMBCD and subsequently stained with anti-PbSEP1 immuneserum, the specific fluorescence was lost or detected in apunctuate distribution in most trophozoites, mature game-tocytes and early schizonts, while rings and late schizontswere not affected by the treatment (Fig. 5b).
3.5. Subcellular localisation of P. falciparum SEP proteins
As shown above, six out of seven Pfsep genes wereactively transcribed during the blood stages of the humanparasite. In order to identify the subcellular localisation ofthe encoded proteins, rat polyclonal antibodies were raised
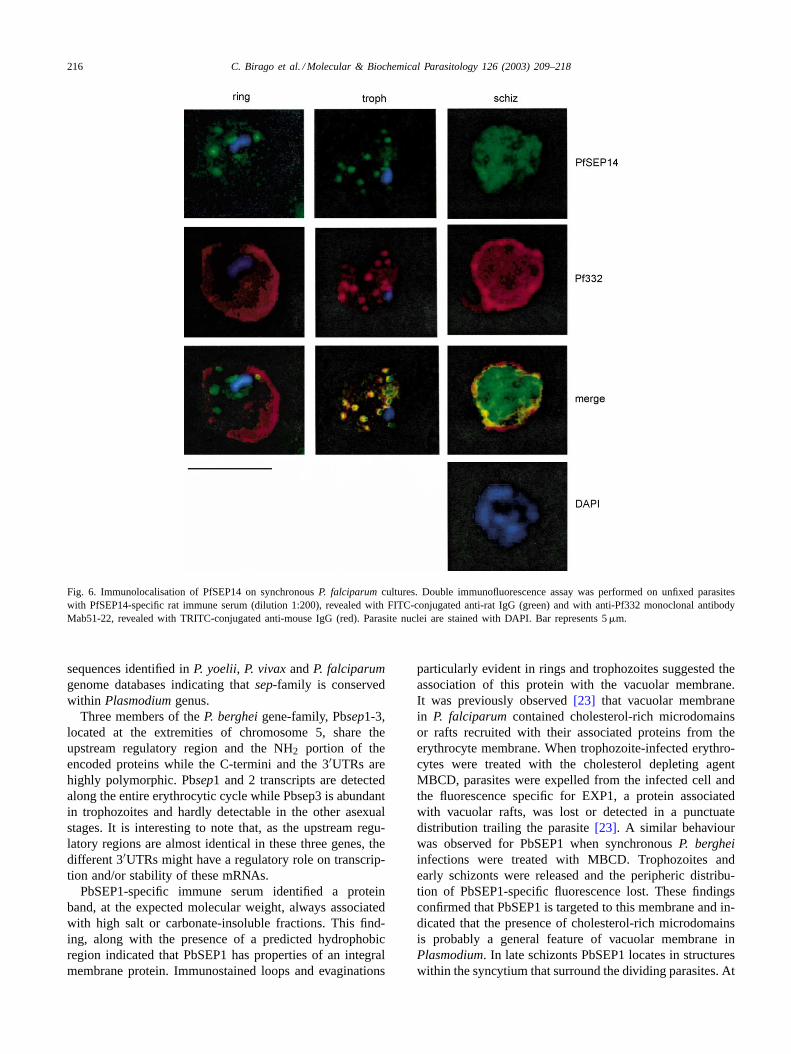
against Pf SEP 4, 11-1 and 14 and tested in immunofluo-rescence assays on synchronous 3D7 parasites. The threeimmune sera gave a similar pattern of fluorescence: at thering and the trophozoite stages vesicle-like structures weredetected within the erythrocytic compartment; with the pro-gression of schizogony the specific-fluorescence appearedmore diffuse and the association with vesicles less evi-dent. In the attempt of identifying the subcellular structuresrecognised by the PfSEP-specific immune sera, colocali-sation experiments were performed with the monoclonalantibody Mab 51-22 raised against Pf332 protein[10]. ThisP. falciparumantigen was found associated with Maurer’sclefts, in the host cell compartment.Fig. 6 shows doublefluorescence assay done with PfSEP14-specific anti serumand anti-Pf332 Mab 51-22 in synchronousP. falciparuminfections. At the stage of ring the distribution of PfSEP14did not coincide with that obtained with Mab51-22 which,at this stage, cross reacts with RESA[10]. A similar pat-tern of fluorescence for both antigens appeared insteadin trophozoites, while at the schizont stage patterns weredistinguishable, in that SEP14-associated fluorescence wasdiffuse within the parasitised cell and did not appear con-centrated at the edges of the schizonts.
4. Discussion
A gene-family encoding small exported proteins (sep) hasbeen identified and characterised inPlasmodium berghei.The encoded proteins (13–16 kDa) contain a predicted signalpeptide at the N-terminus, an internal hydrophobic regionand a polymorphic low complexity region at the C-terminus.These structural features are maintained in related protein
216 C. Birago et al. / Molecular & Biochemical Parasitology 126 (2003) 209–218
Fig. 6. Immunolocalisation of PfSEP14 on synchronousP. falciparumcultures. Double immunofluorescence assay was performed on unfixed parasiteswith PfSEP14-specific rat immune serum (dilution 1:200), revealed with FITC-conjugated anti-rat IgG (green) and with anti-Pf332 monoclonal antibodyMab51-22, revealed with TRITC-conjugated anti-mouse IgG (red). Parasite nuclei are stained with DAPI. Bar represents 5�m.
sequences identified inP. yoelii, P. vivaxandP. falciparumgenome databases indicating thatsep-family is conservedwithin Plasmodiumgenus.
Three members of theP. bergheigene-family, Pbsep1-3,located at the extremities of chromosome 5, share theupstream regulatory region and the NH2 portion of theencoded proteins while the C-termini and the 3′UTRs arehighly polymorphic. Pbsep1 and 2 transcripts are detectedalong the entire erythrocytic cycle while Pbsep3 is abundantin trophozoites and hardly detectable in the other asexualstages. It is interesting to note that, as the upstream regu-latory regions are almost identical in these three genes, thedifferent 3′UTRs might have a regulatory role on transcrip-tion and/or stability of these mRNAs.
PbSEP1-specific immune serum identified a proteinband, at the expected molecular weight, always associatedwith high salt or carbonate-insoluble fractions. This find-ing, along with the presence of a predicted hydrophobicregion indicated that PbSEP1 has properties of an integralmembrane protein. Immunostained loops and evaginations
particularly evident in rings and trophozoites suggested theassociation of this protein with the vacuolar membrane.It was previously observed[23] that vacuolar membranein P. falciparumcontained cholesterol-rich microdomainsor rafts recruited with their associated proteins from theerythrocyte membrane. When trophozoite-infected erythro-cytes were treated with the cholesterol depleting agentMBCD, parasites were expelled from the infected cell andthe fluorescence specific for EXP1, a protein associatedwith vacuolar rafts, was lost or detected in a punctuatedistribution trailing the parasite[23]. A similar behaviourwas observed for PbSEP1 when synchronousP. bergheiinfections were treated with MBCD. Trophozoites andearly schizonts were released and the peripheric distribu-tion of PbSEP1-specific fluorescence lost. These findingsconfirmed that PbSEP1 is targeted to this membrane and in-dicated that the presence of cholesterol-rich microdomainsis probably a general feature of vacuolar membrane inPlasmodium. In late schizonts PbSEP1 locates in structureswithin the syncytium that surround the dividing parasites. At

C. Birago et al. / Molecular & Biochemical Parasitology 126 (2003) 209–218 217
this stage the distribution of the specific fluorescence is notaffected by MBCD treatment. After erythrocyte rupture theprotein was still detected in free merozoites thus suggestingthat PbSEP1 may participate to the initial steps of parasiteinvasion.
This work shows that six out of sevenP. falciparum sepgenes are transcribed during the intraerythrocytic stages. Ex-periments with specific anti sera raised against Pf SEP 4,11-1 and 14 indicate that three members of thesepfamilystart to be expressed early after invasion. This is in agree-ment with a previous work[11] which identified cDNA se-quences corresponding to Pfsep14 and Pfsep11-2 in a set of20 cDNA clones ofP. falciparumspecifically transcribed atthe ring stage. These two early transcribed genes along witha third one[12], which does not exhibit features character-istic of the SEP proteins, were grouped by the authors in agene-family named “early transcribed membrane proteins”(etramp).
A feature conserved within the sep family both in rodentand in human malaria is that the encoded proteins are ex-ported outside the parasite membrane. In the case ofP. fal-ciparum they are probably only transiently associated withthe vacuolar membrane and then rapidly exported to the cy-toplasm of the red blood cell in the form of vesicle-likestructures maintained up to schizogony. A number ofP. fal-ciparumproteins trafficked to the erythrocyte plasma mem-brane or cytoskeleton have been shown to be associated withMaurer’s clefts[9,10,26], membranous structures exportedto the erythrocyte cytosol and most probably involved in pro-tein transport beyond the parasite membrane. Colocalisationexperiments with Pf332, a protein associated with Maurer’sclefts[10] indicated that PfSEP proteins are targeted to thesestructures. It would be interesting to establish whether mem-bers of this protein-family are structural components of thisparasite compartment exported in the host cell.
A contribution to the identification of signals and path-ways underlying protein trafficking withinP. falciparuminfected erythrocytes comes from recent reports in whichsequences of parasite exported proteins fused to reportergenes were expressed into parasites upon transfection. Itwas shown[26] that in the case of the KAHRP (the majorcomponent of the knob structures) the first 60 amino acidsthat include a recessed putative signal peptide, were suffi-cient to direct the green fluorescent protein (GFP) to the PV,but not to the erythrocyte compartment where the protein isnormally translocated. The latter process required sequenceelements within the next 63 amino acids. On the other hand,the N-terminal signal peptide of PfEXP1, a protein associ-ated with the vacuolar membrane, was sufficient to directthe luciferase reporter gene to the erythrocyte compartment[27], as if specific signals were required to retain the proteinin the vacuolar compartment, the transport to the erythro-cyte cytosol representing for this protein the default path-way. Members ofP. bergheiandP. falciparum sep-family,characterised in this work, localise in different cellular com-partments. The conserved protein motifs are probably not
involved in determining protein targeting beyond vacuolarmembrane since PbSEP1 never crosses this compartment,while PfSEP proteins mainly reside in the host cell cyto-plasm. It was suggested[28] that membrane rafts might pro-vide mechanisms to sort between proteins that accumulatein the vacuolar membrane from those that are released at theerythrocyte surface. Transfection experiments are under wayaimed at identifying specific signals directing SEP proteinsin the different cell compartments.
Acknowledgements
We thank Denise Mattei and Tamara Petrucci for helpfuldiscussion and critical reading of the manuscript. We wishto thank the scientists and funding agencies comprising theinternational Malaria Genome Project for making sequencedata from the genome ofP. falciparum(3D7) public priorto publication of the completed sequence. The Sanger Cen-tre (UK) provided sequence for chromosomes 1, 3–9, and13, with financial support from the Wellcome Trust. A con-sortium composed of The Institute for Genome Research,along with the Naval Medical Research Center (USA), se-quenced chromosomes 2, 10, 11 and 14, with support fromNIAID/NIH, the Burroughs Wellcome Fund, and the Depart-ment of Defense. The Stanford Genome Technology Cen-ter (USA) sequenced chromosome 12, with support fromthe Burroughs Wellcome Fund. The Plasmodium GenomeDatabase is a collaborative effort of investigators at the Uni-versity of Pennsylvania (USA) and Monash University (Mel-bourne, Australia), supported by the Burroughs WellcomeFund.
References
[1] Foley M, Tilley L. Protein trafficking in malaria-infected erythrocytes.Int J Parasitol 1998;28:1671–80.
[2] Lingelbach K, Joiner KA. The parasitophorous vacuole membranesurroundingPlasmodiumand Toxoplasma: an unusual compartmentin infected cells. J Cell Sci 1998;111:1467–75.
[3] Haldar K. Intracellular trafficking inPlasmodium-infected erythro-cytes. Curr Opin Microbiol 1998;1:466–71.
[4] Hayashi M, Taniguchi S, Ishizuka Y, et al. A homologue ofN-ethylmaleimide-sensitive factor in the malaria parasitePlasmod-ium falciparum is exported and localized in vesicular structures inthe cytoplasm of infected erythrocytes in the brefeldin A-sensitivepathway. J Biol Chem 2001;276:15249–55.
[5] Albano FR, Berman A, La Greca N, et al. A homologue of Sar1plocalises to a novel trafficking pathway in malaria-infected erythro-cytes. Eur J Cell Biol 1999;78:453–62.
[6] Adisa A, Albano FR, Reeder J, Foley M, Tilley L. Evidence for arole for aPlasmodium falciparumhomologue of Sec31p in the exportof proteins to the surface of malaria parasite-infected erythrocytes.J Cell Sci 2001;114:3377–86.
[7] Albano FR, Foley M, Tilley L. Export of parasite proteins to the ery-throcyte cytoplasm: secretory machinery and traffic signals. NovartisFound Symp 1999;226:157–75.

218 C. Birago et al. / Molecular & Biochemical Parasitology 126 (2003) 209–218
[8] van Dooren GG, Waller RF, Joiner KA, Roos DS, McFadden GI.Traffic jams: protein transport inPlasmodium falciparum. ParasitolToday 2000;16:421–7.
[9] Blisnick T, Morales Betoulle ME, Barale JC, et al. Pfsbp1, a Maurer’scleft Plasmodium falciparumprotein, is associated with the erythro-cyte skeleton. Mol Biochem Parasitol 2000;111:107–21.
[10] Hinterberg K, Scherf A, Gysin J, et al.Plasmodium falciparum:the Pf332 antigen is secreted from the parasite by a brefeldinA-dependent pathway and is translocated to the erythrocyte mem-brane via the Maurer’s clefts. Exp Parasitol 1994;79:279–91.
[11] Spielmann T, Beck HP. Analysis of stage-specific transcription inPlasmodium falciparumreveals a set of genes exclusively transcribedin ring stage parasites. Mol Biochem Parasitol 2000;111:453–8.
[12] Horii T, Bzik DJ, Inselburg J. Characterization of antigen-expressingPlasmodium falciparumcDNA clones that are reactive with parasiteinhibitory antibodies. Mol Biochem Parasitol 1988;30:9–18.
[13] Ponzi M, Janse CJ, Dore E, et al. Generation of chromosome sizepolymorphism during in vivo mitotic multiplication ofPlasmodiumberghei involves both loss and addition of subtelomeric repeat se-quences. Mol Biochem Parasitol 1990;41:73–82.
[14] Janse CJ, Ramesar J, van den Berg FM, Mons B.Plasmodiumberghei: in vivo generation and selection of karyotype mutants andnon-gametocyte producer mutants. Exp Parasitol 1992;74:1–10.
[15] Janse CJ, Mons B, Croon JJ, van der Kaay HJ. Long-term in vitrocultures of Plasmodium bergheiand preliminary observations ongametocytogenesis. Int J Parasitol 1984;14:317–20.
[16] Alano P, Silvestrini F, Roca L. Structure and polymorphism of theupstream region of the pfg27/25 gene, transcriptionally regulated ingametocytogenesis ofPlasmodium falciparum. Mol Biochem Para-sitol 1996;79:207–17.
[17] van Lin LH, Pace T, Janse CJ, et al. Interspecies conservation ofgene order and intron–exon structure in a genomic locus of highgene density and complexity inPlasmodium. Nucleic Acids Res2001;29:2059–68.
[18] Birago C, Pace T, Barca S, Picci L, Ponzi M. A chromatin-asso-ciated protein is encoded in a genomic region highly conserved
in the Plasmodium genus. Mol Biochem Parasitol 1996;80:193–202.
[19] Birago C, Pace T, Picci L, et al. The putative gene for the first enzymeof glutathione biosynthesis inPlasmodium bergheiandPlasmodiumfalciparum. Mol Biochem Parasitol 1999;99:33–40.
[20] Bottius E, Bakhsis N, Scherf A.Plasmodium falciparumtelomerase:de novo telomere addition to telomeric and nontelomeric sequencesand role in chromosome healing. Mol Cell Biol 1998;18:919–25.
[21] Carter R, Graves PM, Creasey A, et al.Plasmodium falciparum: anabundant stage-specific protein expressed during early gametocytedevelopment. Exp Parasitol 1989;69:140–9.
[22] Read M, Sherwin T, Holloway SP, Gull K, Hyde JE. Microtubularorganization visualized by immunofluorescence microscopy duringerythrocytic schizogony inPlasmodium falciparumand investigationof post-translational modifications of parasite tubulin. Parasitology1993;106:223–32.
[23] Lauer S, VanWye J, Harrison T, et al. Vacuolar uptake of hostcomponents, and a role for cholesterol and sphingomyelin in malarialinfection. EMBO J 2000;19:3556–64.
[24] Dawson JP, Weinger JS, Engelman DM. Motifs of serine and thre-onine can drive association of transmembrane helices. J Mol Biol2002;316:799–805.
[25] van Lin LH, Pace T, Janse CJ, Scotti R, Ponzi M. A long range re-striction map of chromosome 5 ofPlasmodium bergheidemonstratesa chromosome specific symmetrical subtelomeric organisation. MolBiochem Parasitol 1997;86:111–5.
[26] Wickham ME, Rug M, Ralph SA, et al. Trafficking and assemblyof the cytoadherence complex inPlasmodium falciparum-infectedhuman erythrocytes. EMBO J 2001;20:5636–49.
[27] Burghaus PA, Lingelbach K. Luciferase, when fused to an N-terminalsignal peptide, is secreted from transfectedPlasmodium falciparumand transported to the cytosol of infected erythrocytes. J Biol Chem2001;276:26838–45.
[28] Haldar K, Samuel BU, Mohandas N, Harrison T, Hiller NL. Transportmechanisms inPlasmodium-infected erythrocytes: lipid rafts and atubovesicular network. Int J Parasitol 2001;31:1393–401.




















