
Leucine Supplementation Accelerates Connective Tissue Repair of Injured Tibialis Anterior Muscle
Upload
independentCategory
view
1download
0
DNA AND CELL BIOLOGYVolume 25, Number 6, 2006© Mary Ann Liebert, Inc.Pp. 331–339
trnp: A Conserved Mammalian Gene Encoding a NuclearProtein That Accelerates Cell-Cycle Progression
MARINA VOLPE, SALLY SHPUNGIN, CHANY BARBI, GALYA ABRHAM, HANNA MALOVANI, RON WIDES, and URI NIR
ABSTRACT
We herein describe a novel protein encoded by a single exon in a single-copy conserved mammalian gene.This protein, termed TMF regulated nuclear protein (TRNP), was identified in a yeast “two-hybrid” screenin which the “BC box” containing protein-TMF/ARA160 served as a bait. TRNP is a basic protein which ac-cumulates in an insoluble nuclear fraction in mammalian cells. It is 227 aa long in humans and chimps and223 aa long in mice. Enforced expression of TRNP in cells that do not express this protein significantly in-creased their proliferation rate by enhancing their cell-cycle progression from the G0/G1 to the S phase. Likeanother proliferation promoting factor, Stat3, TRNP was directed to proteasomal degradation by TMF/ARA160. Thus, the trnp gene encodes a novel mammalian conserved nuclear protein that can accelerate cell-cycle progression and is regulated by TMF/ARA160.
331
INTRODUCTION
MAMMALIAN REGULATORY CASCADES are multicomponentialand complex systems that direct the specific response of
different cell types to external and intracellular cues. Identifi-cation of novel cellular regulators and exploration of their cellular functions should further our understanding of the reg-ulation of both novel and known signaling pathways. Charac-terization of a new regulatory factor could be achieved by thefunctional analysis of genetic information pertinent to a novelgene. Alternatively, a previously unknown regulator can beidentified through its binding to a known cellular partner(Rigaut et al., 1999; Knuesel et al., 2003). We have recentlyidentified a Golgi resident protein (Mori and Kato, 2002; Frid-mann-Sirkis et al., 2004) termed TATA Element ModulatoryFactor (TMF/ARA160) (Garcia et al., 1992) that under definedstress conditions is dispersed in myoblastic cells and thereafterrecruits the growth promoting transcription factor Stat3, to pro-teasomal degradation (Perry et al., 2004). While screening forproteins that can interact with TMF/ARA160 in a yeast two-hybrid system (Schwartz et al., 1998), we isolated a cDNA en-coding a previously uncharacterized mouse protein. Here, weshow that this protein accumulates in the nuclei of mammaliancells and can be directed to proteasomal degradation by TMF/
ARA160, thus endowing it the name TMF Regulated NuclearProtein (TRNP). TRNP is highly conserved in mammals, andis encoded by one exon of a single-copy gene in all sequencedmammalian genomes. Interestingly, TRNP was found to facil-itate cell-cycle progression in mammalian cells.
MATERIALS AND METHODS
Bioinformatic sources and analyses
A trnp mouse cDNA was uncovered in a two-hybrid screenfor TMF/ARA160 interactors. Blast, Fasta, and Smith-Water-man searches with trnp gene and protein sequences were car-ried out against public protein, EST, cDNA, and genome data-bases to uncover proteins, messages, genes, homologs, andhomologous domains. Genome database searches were carriedout against protein models, gene models, genome assemblies,and where necessary, genomic sequence trace databases. Mul-tialignments were performed using Clustal and GCG-Pileup al-gorithms. Analysis of protein structures, domains, and motifsincluded the following sites and algorithms: ELM (EukaryoteLinear Motif resource); SMART (HMM based Simple Modu-lar Architecture Research Tool); and ExPaSy (Expert ProteinAnalysis System).
Faculty of Life Sciences, Bar-Ilan University, Ramat-Gan, Israel.

Cell lines
Murine myoblastic C2C12, human MCF-7, and HEK293cells were grown in Dulbecco’s modified Eagle’s medium(DMEM) (Gibco, Gaithersburg, MD). Growth medium wassupplemented with 10% fetal calf serum (FCS) (Beit Haemek,Israel). For inhibition of proteasome activity cells were sub-jected to 50 �M of the proteasome inhibitor MG-132 for 5 h.
Transfection of cells
C2C12 cells (25 � 105) or 5 � 106 MCF-7 or 1 � 106 293cells were plated in 10-cm plates, 24 h before transfection. Thecells were transfected using the LipofectAMINE reagent, ac-cording to the manufacturer’s instructions (Invitrogen, SanDiego, CA). Each transfection was performed by mixing 12 �gDNA with 32 �l of LipofectAMINE and 1 ml serum-free Opti-MEM (Gibco) medium. The mixture was incubated for 25 minat room temperature, and was then diluted in 5 ml of serum-free medium. Diluted DNA suspensions were overlaid onto thecells which were then incubated for 6.5 h at 37°C in a CO2 in-cubator. After incubation, 5 ml Opti-MEM supplemented with10% FCS was added and the resulting mixture was replaced 15h later with 10 ml fresh DMEM containing 10% FCS. For sta-ble transfection, the cells were selected with 0.5 mg/ml G418(Sigma, St. Louis, MO) for 2 weeks. For analyzing the effectof TMF/ARA160 on TRNP, C2C12 cells were cotransfectedwith 10 �g DNA vector encoding TMF/ARA160 and 5 �gDNA vector-encoding TRNP.
Construction of plasmids
The pCMV–c-Myc–TRNP expression plasmid was con-structed by amplifying a murine TRNP clone (MGC Gene ser-vice—6491759) with a forward primer: 5� TTA GAA TTCTGC CGG GCT GCC GAA TCA 3�, which bears at it’s 5� enda EcoR1 site, and a reverse primer, 5� TTT AAC TCG AGTCAG CGC TGG GGG GAG G 3�, which bears an Xho1 site atits 3� end. The amplified fragment was cloned into an EcoR1–Xho1 restricted pCMV–c-Myc vector (BD Clontech, Palo Alto,CA). The same fragment was cloned into an EcoR1–Xho1 re-stricted pcDNA3 vector (Invitrogen) for the pcDNA3–TRNPexpression plasmid.
For the construction of a bacterial TRNP expression vector,cDNA encoding the mouse TRNP was flanked by BamHI andHindIII sites, and was inserted between the BamHI and HindIIIsites in the pQE30 bacterial expression vector (Qiagen Inc.,Chatsworth, CA). In this vector, the expressed protein is precededat its N-terminus by six histidine residues (His–TRNP), and couldthus be purified by immobilized metal affinity chromatography(IMAC). The pcDNA3 HA-TMF/ARA160 expression vector hadbeen described before (Schwartz et al. 1998).
Preparation of anti-TRNP antibodies
6xHis–TRNP was expressed in Esherichia coli JM109 cellsand was then purified by IMAC, using TALON-metal affinityresin (Clontech), which consists of Sepharose–cobalt beads. Pu-rified TRNP (500 �g) was injected subcutaneously into 3-month-old female rabbits. This was repeated two more times at21-day-intervals, and the presence of anti-TRNP antibodies inthe obtained sera was determined using Western blot analysis.
Subcellular fractionation
Subcellular fractionation was carried out basically as de-scribed before (Mendez and Stillman, 2000).
C2C12 cells were harvested and resuspended (2 � 107
cells/ml) in buffer A (10 mM HEPES pH 7.9, 10 mM KCl, 1.5mM MgCl2, 0.34 M sucrose, 10% glycerol, 1 mM DTT, 5 �g/mlaprotinin, 5 �g/ml leupeptin, and 0.5 �g/ml pepstatin A). Tri-ton X-100 was added (0.1% final), and the cells were incubatedfor 5 min on ice (TCE-total cell extract). Nuclei were collectedin the pellet (P) by low-speed centrifugation (4 min, 1300 � g,at 4°C). The supernatant (S) was further clarified by high-speedcentrifugation (15 min, 20,000 � g, at 4°C) (CS-cytoplasmicsupernatant) to remove cell debris and insoluble aggregates,which themselves were discarded as cytoplasmic pellet. Nuclei(pellet) were washed once in buffer A, and were then lysed inbuffer B (3 mM EDTA, 0.2 mM EGTA, 1 mM DTT, and pro-tease inhibitors as described above). Lysates were centrifuged(4 min, 1700 � g, 4°C) and supernatants (NS—nuclear super-natant) were collected. Insoluble chromatin was washed oncein buffer B, and was centrifuged again under the same condi-tions. The final chromatin pellet (NP—nuclear pellet) was re-suspended in SDS-sample buffer and sonicated for 15 sec in aMisonix Sonicator3000 using a microtip at 25% amplitude. Torelease chromatin-bound proteins by nuclease treatment, cellnuclei (P) were resuspended in buffer A plus 1 mM CaCl2 and0.2 U of micrococcal nuclease (Sigma). After 1 min incubationat 37°C, in the presence or absence of 0.5 M NaCl, the nucle-ase reaction was stopped by the addition of 1 mM EGTA. Nu-clei were collected by low-speed centrifugation and were lysedaccording to the chromatin isolation protocol described above.
Western blot analysis
Whole-cell proteins from cell lines and organs were extractedin lysis buffer: 20 mM Tris pH 7.5, 150 mM NaCl, 1 mM EDTA,1% Nonidet P-40, 0.5% sodium deoxycholate, and 2 mMNa2VO4, and were incubated with agitation for 40 min at 4°C.The proteins were cleared by centrifugation at 14,000 � g for 30min at 4°C. Protein concentration was determined by Bradfordanalysis. For Western blot analysis, 30 �g of protein from eachsample was resolved by 12% or 14% SDS-PAGE. Electroblottedproteins were detected using polyclonal anti-TRNP antibodies,anti-c-Myc antibodies (Zymed Laboratories, San Francisco, CA),anti-HA antibodies (Covance), anti-�-actin antibodies (Sigma), oranti-H2B antibodies (Santa Cruz Biotechnology, Santa Cruz, CA).Immunoabsorbed antibodies were visualized using chemilumi-nescence reaction (Pierce, Rockford, IL).
Immunocytochemical analysis
Immunocytochemistry was carried out essentially as de-scribed before (Ben-Dor et al., 1999; Priel-Halachmi et al.,2000). C2C12 cells were exposed to 1:100 anti-c-Myc (ZymedLaboratories) or anti-HA (Covance) antibodies. Mouse mono-clonal antibodies were visualized with fluorescein isothio-cyanate conjugated donkey antimouse secondary antibodies(Jackson Laboratories, Bar Harbor, ME). Nuclei were visual-ized by staining the DNA with 0.05 mg/ml propidium iodide.Bound fluorophors were detected with a Bio-Rad (Hercules,CA) MRC 1024 upright confocal microscope with a krypton–
VOLPE ET AL.332

FIG. 1. trnp is a conserved mammalian gene with a single encoding exon. (A) The human and mouse trnp genes each producean invariantly spliced approximately 1.7-kb two-exon message from an approximately 7-kb nascent message. The entire codingregion resides within the first exon of each message. Exons, introns, and coding regions are diagrammed according to the pro-vided key. Genome position is indicated by cytogenetic locus (4 D3 for mouse and 1p36.11 for human), and by position on thechromosomes’ sequence axes (with provided scales). (B) Multialignment of five mammalian TRNP protein sequences: humanprotein gi: 61966741; mouse protein gi: 38078875; and chimpanzee, dog, and cow protein models derived through sequence sim-ilarities to genome and expressed sequences of these three organisms. Amino acid identities shown in black boxes, similaritiesin shades of gray. (C) A tree indicating phylogenetic distances between TRNP protein core sequences for seven mammalianspecies, as determined by ClustalW.
argon ion laser. Confocal microscope image analysis was per-formed using Bio-Rad software, and figures were compiled us-ing the Laser Sharp 3.0 software package.
Quantitation of viable cells
Relative number of viable cells in a given culture was de-termined by either counting Trypan blue excluding cells or byapplying the MTT assay. Cultures were assayed for the pro-duction of formazan through the reduction of MTT (3-[4,5-di-methylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide). Five-thousand MCF-7 cells per well were plated in 96-well plates.Twelve and 120 h after plating, growth medium was removedand cells were incubated for 2 h with 100 �l of MTT solution(5 mg/ml MTT dissolved in PBS) at 37°C. Cells were then lysedwith 50% N,N-dimethyl formamide and the levels of the for-mazan reduction product were determined by measuring ab-sorbance at 570 nm in a computer-controlled microplate ana-lyzer.
Flow cytometry analysis
Cells were rinsed twice with cold phosphate-buffered saline(PBS), and were then harvested after treatment with trypsin(Beit Haemek). Suspended cells were washed twice in 10-mlcold PBS and collected at 500 � g for 5 min. The pellet wasresuspended in 200 �l of cold PBS and cells were stained inthe dark with 200 �l propidium iodide (PI) solution (20 mMTris pH 8, 1 mM NaCl, 0.1% (v/v) NP-40, 1.4 mg/ml RNaseA, 0.005 mg/ml propidium iodide) for 30 min at 37°C. The to-tal cellular DNA content was determined using a Becton Dick-inson flow cytometer (FACSCalibur), Multiple software andModFit LT software.
BrdU incorporation assay
The cells were washed twice with cold PBS, and were fixedfor 30 min with cold ethanol (70%) in ice. Fixed cells were in-cubated for 30 min in 2 M HCl 0.5% Triton X-100. The cellswere centrifuged and resuspended in 0.1 M Na2B4O7*10 H2O,pH 8.5. The cells were spun down and resuspended in block-ing solution (1% bovine serum albumin in PBS 0.5% Tween)for 30 min at room temperature in the presence of 1:50 dilutedanti-BrdU monoclonal antibody (Becton Dickinson, Fullerton,CA). Cells were centrifuged again, and were then incubated un-der the same conditions with 1:100 fluorescein isothiocyanate(FITC)-conjugated goat antimouse antibody (Jackson Labora-tories). The cells were centrifuged and stained in the dark with5 �g/ml PI solution. The cellular DNA content was determinedusing a Becton Dickinson flow cytometer (FACSCalibur) andMultiple software.
Semiquantitative RT-PCR analysis
Whole-cell RNA was extracted from organs using TRIReagent (Molecular Research Center, ICN, Aurora, OH) fol-lowing the manufacture’s instructions. Two micrograms of to-tal RNA was reverse-transcribed using the ThermoScript™ RT-PCR System (Invitrogen) with oligo dT primer and was thenamplified with specific nucleotides derived from murine trnpmRNA: forward primer 5� ACG GAC GGA CGG ATC CA 3�and reverse primer 5� CAG ACC CTT GCC ACT GTA A 3�.The selected primers are derived from separated exons (1 and2), which flank an intron of the trnp gene. PCR was performedunder linear conditions, at which 25 cycles were found to beoptimal for quantitative comparison of the TRNP mRNA lev-els. The expected 245-bp long PCR product is specific to themature TRNP mRNA, and can be differentiated from a con-tamination of genomic DNA PCR products. 18S ribosomalRNA primer pairs (Quantum RNA, Ambion, Austin, TX) were
VOLPE ET AL.334
FIG. 2. Expression of the trnp in mice tissues. (A) Whole-cell RNA was prepared from mice tissues, and samples weresubjected to semiquantitative RT-PCR analysis (upper panel).18S ribosomal RNA served as an internal control (lower panel).(B) Whole-cell lysates were prepared from mice tissues. Pro-teins were resolved in SDS-PAGE and were then reacted with anti-TRNP or antiactin antibodies using a Western blotanalysis.
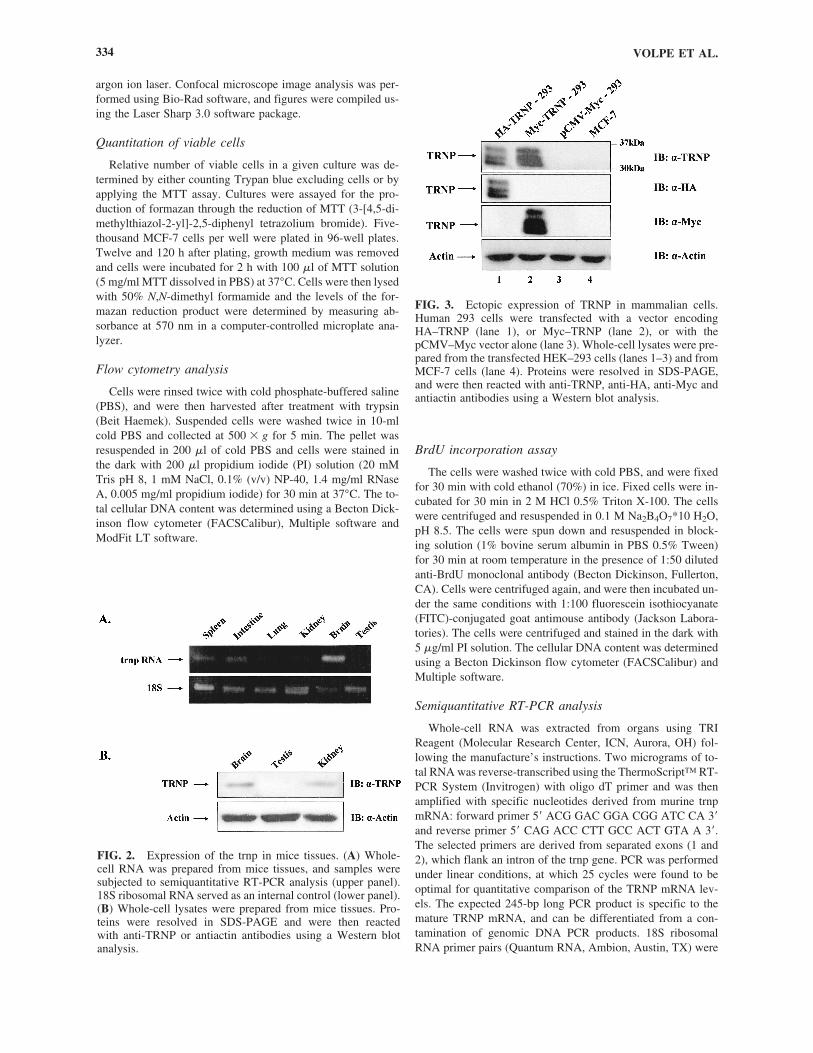
FIG. 3. Ectopic expression of TRNP in mammalian cells. Human 293 cells were transfected with a vector encodingHA–TRNP (lane 1), or Myc–TRNP (lane 2), or with thepCMV–Myc vector alone (lane 3). Whole-cell lysates were pre-pared from the transfected HEK–293 cells (lanes 1–3) and fromMCF-7 cells (lane 4). Proteins were resolved in SDS-PAGE,and were then reacted with anti-TRNP, anti-HA, anti-Myc andantiactin antibodies using a Western blot analysis.

used to yield a 489-bp fragment derived from the 18S rRNA asan internal control. PCR products were separated on a 1.4%agarose gel, and were visualized with ethidium bromide.
RESULTS
trnp encodes a conserved mammalian protein from asingle genomic exon
While characterizing proteins that could interact withTMF/ARA160 (Perry et al., 2004) in a yeast two-hybrid screen-ing system (Schwartz et al., 1998), we isolated a murine cDNAcorresponding to a 1607 bp RIKEN full-length cDNA, with a223-aa reading frame (2300002D11Rik, BC049168.1; [TheFANTOM Consortium and The RIKEN Genome ExplorationResearch Group Phase I & II Team, 2002]). A corresponding1745-bp cDNA exists in humans, encoding a 227-aa proteinwith 90% sequence similarity to that of the mouse (LOC388610,BC069216.1). This gene, which we refer to as trnp (see below),is a single-copy gene per haplotype in each of these two species.
The genes reside at chromosome 4 D3 in the mouse, and atchromosome 1p36.11 in humans, as loci: 2300002D11Rik–NCBI Refseq XM_149530.5; and LOC388610–NCBI RefseqNM_001013642.1; respectively (Fig. 1A). Both orthologs aretwo exon genes which encode their protein products from thefirst exon only (Fig. 1A). Several dozen EST records for thesespecies, and others suggest that the genes do not undergo al-ternative splicing.
Comprehensive searches of genomic and expressed sequencedatabases revealed a single ortholog for every sequenced mam-malian genome. This included all placental mammals plus themarsupial opossum, all of which have a single clearly conservedortholog (Fig. 1B), with no additional genes of any significantsimilarity. Phylogenetic trees’ structures support the existenceof a single, never duplicated, ancestral gene for all of the trnporthologs (Fig. 1C). In contrast, intensive and exhaustivesearches of other sequenced vertebrate genomes revealed atmost very fragmentary and distant domain similarities. Nonepresent a compelling case for homologous genes or related pro-teins. This included analyses of an amniote (chicken), an am-phibian (Xenopus tropicalis), and several fish genomes. Hence,
A NOVEL CONSERVED MAMMALIAN GENE 335
FIG. 4. Subcellular distribution of TRNP. C2C12 cells were transfected with a pCMV–Myc vector alone (A–C), or with thepCMV–Myc–TRNP plasmid (D–F). Forty-eight hours posttransfection the cells were fixed and costained with anti-Myc anti-bodies (B, E), and with propidium iodide for nuclei visualization (A, D). The photographs represent 1-�m confocal laser sec-tions. The original magnification of cells was �600. (G) C2C12 cells were subjected to subcellular fractionation. Protein lysateswere prepared from the total cell extract (TCE, lane 1), from the cytoplasm (lane 2), and from the nuclear fraction (lane 3) ac-cording to the protocol indicated in Materials and Methods and in scheme 5A. Nuclei pellets (P) were dissolved in a SDS-sam-ple buffer followed by sonication. Proteins from all fractions were resolved in SDS-PAGE and were then reacted with anti-TRNPor antihistone H2B antibody in a Western blot analysis.

this gene appears to have arisen exclusively in the mammalianlineage.
The multialignment indicated that the first 20 aa, and the Cterminal half represent the highly conserved regions of TRNPproteins. Protein sequence analysis did not convincingly revealthe presence of any known motifs or domains. However, thepresence of low-complexity amino acid clusters are identified,two of which are present in all mammalian orthologs. A pro-line rich region extends from Pro20 to Pro54 in the murine pro-tein, and in similar positions in other orthologs, while displacedin opossum (Fig. 1B). A glycine/alanine-rich region extendsfrom Gly72 to Gly94 in the mouse protein, and in all placentalorthologs (Fig. 1B).
Expression of the trnp gene in mice tissues
To analyze the expression pattern of the trnp gene, whole-cell RNA was prepared from mice tissues, and the relative lev-els of the trnp RNA were determined using RT-PCR analysis.trnp RNA was detected in all tissues analyzed. However, therelative level of the trnp RNA varied among the different tis-sues, with the highest level detected in the brain (Fig. 2A). Thiscoincides with profiles obtained using DNA array analysis inhuman tissues, which indicated relatively high trnp RNA lev-els in the brain (Yanai et al., 2005). To follow the accumula-tion of the TRNP protein in different tissues and cell types,polyclonal antibodies were raised against the TRNP protein.When these anti-TRNP antibodies were reacted with murine tis-sue extracts a major band of approximately 36 kDa was de-tected in brain and kidney but not in testes lysates (Fig. 2B).This coincides with the trnp RNA expression profiles in theseorgans (Fig. 2A). To verify the specificity of the anti-TRNP an-tibodies, HEK293 human kidney epithelial cells were trans-fected with an expression vector encoding TRNP fused eitherto a Myc or HA tag. Both chimeric proteins reacted with theantibodies, giving rise to heterogeneous populations of bandswhich extended from 30 to 36 kDa (Fig. 3). This could reflectextensive posttranslational modifications that TRNP undergoesin the transfected cells. Interestingly, no endogenous TRNP pro-tein could be detected in the 293 cells, nor was this protein de-tected in breast carcinoma MCF-7 cells (Fig. 3, lanes 3 and 4).However, TRNP was detected, although in relatively low lev-els, in C2C12 myoblastic cells and in fibroblastic cells (Fig. 4Gand data not shown).
TRNP is a nuclear protein
To gain a better understanding of the functional role ofTRNP, we analyzed the subcellular localization of TRNP. Theprotein was fused to a Myc tag, and was ectopically expressedin C2C12 myoblastic cells. This enabled us to specifically uti-lize monoclonal anti-Myc antibodies in an immunocytochemi-cal assay. Cells were fixed 48 h posttransfection, and were thensubjected to immunocytochemical analysis. TRNP accumulatedexclusively in the nuclei of the transfected cell. However, it wasnot evenly distributed in the nucleus, and it was excluded fromdefined nuclear spots. Some of these spots were intensivelystained with propidium iodide, and may represent highly con-densed chromatin (Amirand et al., 1998) (Fig. 4A–F). Similardistribution profiles were obtained when PC3 cells stably ex-pressing an HA-tagged TRNP were subjected to an indirect im-
munocytochemical analysis, using anti-HA monoclonal anti-bodies (data not shown). Like the exogenous protein, the en-dogenous TRNP was also mainly detected in the nuclear frac-tion of C2C12 cells (Fig. 4G). However, it should be stressedthat no endogenous TRNP could be detected in enriched nu-clear fractions prepared from MCF-7 cells.
To further characterize the nuclear accumulation of TRNP,a biochemical, marker validated fractionation assay was ap-plied to the transfected C2C12 cells (Fig. 5A). TRNP was ab-sent from the cytosolic fraction and was constantly associ-ated with an insoluble nuclear fraction that also containedhistones (Fig. 5B). The inclusion of TRNP in a nuclear pre-cipitate could reflect the association of this protein with chro-
VOLPE ET AL.336
FIG. 5. TRNP is associated with insoluble nuclear fraction.(A) Subcellular fractionation scheme (for details, see Materialsand Methods). (B) C2C12 cells were transfected with thepCMV–Myc–TRNP expression vector. Forty-eight hours post-transfection cells were subjected to biochemical fractionationaccording to the scheme in A. Protein lysates were preparedfrom the total cell extract (TCE), soluble cytoplasmic fraction(CS), solubilized nuclear proteins fraction—the nucleoplasm(NS)—and from chromatin–nuclear matrix-bound fraction(NP). Nuclei were either treated or not treated with micrococ-cal nuclease. (C) In addition to other treatments, nuclei wereincubated with 0.5 M NaCl. Protein extracts were then preparedfrom all fractions. (D) SDS (0.5%) was added to the nuclei ly-sis buffer, and the nuclear lysates were divided into NS and NPfractions. Proteins from all fractions were resolved in SDS-PAGE, and were reacted with anti-Myc (upper panel) or anti-H2B antibody (lower panel) in a Western blot analysis. Theprofiles represent one out of three independent experiments thatgave similar results.

matin structures. Treatment of nuclei with micrococcal en-donuclease led to the release of histones to the nucleoplasm,but did not affect the association of TRNP with the insolu-ble nuclear fraction (Fig. 5B). However, combining nucleasetreatment with a 0.5 M NaCl wash led to a partial release ofTRNP (Fig. 5C). An almost complete dissolution of the re-cipitated TRNP was achieved upon exposure of nucleasetreated nuclei to 0.5% SDS (Fig. 5D). Hence, TRNP is asso-ciated with insoluble nuclear fraction.
TRNP accelerates the proliferation rate of MCF-7 cells
To explore the cellular roles of TRNP the protein was ec-topically expressed in cells that do not accumulate it at de-tectable levels. MCF-7 breast carcinoma cells were stabelytransfected with a vector directing the expression of a HA-tagged TRNP under the control of a CMV promoter (HA–TRNPcells). In parallel, cells were transfected with the expressionvector alone (pcDNA3 cells) (Fig. 6A). MCF-7 cells ectopi-cally expressing the HA–TRNP protein exhibited a higher pro-liferation rate than the cells transfected with vector alone. In-terestingly, this difference between the two cell populations wasprofoundly seen only after 5 days in culture (Fig. 6B). UsingMTT analysis, we confirmed the differences between the num-ber of cells in the two cultures. After seeding identical numberof cells and propagating the HA–TRNP expressing and thepcDNA3 cells for 5 days in culture, the number of theHA–TRNP cells was 2–2.5-fold higher than the number of thepcDNA3 cells (Fig. 6C). To examine whether TRNP affects thecell-cycle progression profile of the transfected cells, bothHA–TRNP and pcDNA3 cells were subjected to flow-cytome-
try analysis. This revealed a significant twofold increase in thepercentage of HA–TRNP cells that reside in the S phase witha concomitant decrease in the percentage of G0/G1 cells (Fig.7A). BrdU incorporation analysis corroborated the approxi-mately twofold increase in the percentage of HA–TRNP cellsthat reside in the S phase (Fig. 7B). Interestingly, the percent-age of cells which reside in the G2/M phase was also alteredby TRNP (Fig. 7A), raising the possibility that TRNP could af-fect the G2-M transition as well.
These results suggest that TRNP accelerates the prolifera-tion rate of mammalian cells by enhancing their progressionthrough key cell-cycle transition points.
TMF/ARA160 directs the proteasomal degradation of TRNP
The interaction between TMF/ARA160 and TRNP in a yeast“two-hybrid” system suggested a functional link between thetwo proteins. TMF/ARA160 is a “BC box” containing proteinwhich was shown to direct the proteasomal degradation of Stat3 (Perry et al., 2004). We therefore examined whether TMF/ARA160 can drive the proteasomal degradation of TRNP.C2C12 cells were cotransfected with vectors encodingTMF/ARA160 and TRNP in the absence or presence of a pro-teasome inhibitor. While coexpression of ectopic TMF/ARA-160 led to the proteasomal degradation of TRNP, subjection ofthe cells to the proteasome inhibitor MG-132 prevented this ef-fect (Fig. 8). It should be noted that the proteasome inhibitorelevated the overall level of the ectopic TRNP (Fig. 8, lanes 3and 4), most probably by also affecting the endogenousTMF/ARA160. Hence, TMF/ARA160 can direct the proteaso-mal degradation of TRNP.
A NOVEL CONSERVED MAMMALIAN GENE 337
FIG. 6. TRNP increases the proliferation rate of MCF-7 cells. (A) MCF-7 cellswere stably transfected with the pcDNA3–HA–TRNP expression vector (lane 1)or with the pcDNA3 vector alone (lane 2). Whole-cell lysates were resolved inSDS-PAGE, and were then reacted with anti-HA (upper panel), anti-TRNP (mid-dle panel), or with antiactin antibody (lower panel), in a Western blot analysis.(B) MCF-7 cells stably expressing HA–TRNP, or transfected with the expressionvector alone were seeded and grown in culture. The number of viable cells wascounted each day using trypan blue vital stain. (C) MCF-7 cells stably express-ing HA–TRNP, or transfected with the expression vector alone were seeded andgrown in culture and were subjected to MTT analysis, after 12 h or 120 h.

DISCUSSION
As the coding sequences of the human genome are being re-vealed, novel genes are being identified. This necessitates theascription of functional roles to the newly discovered genes.We describe a novel single copy gene which is located at hu-
man chromosome 1p36.11 and at mouse chromosome 4 D3.The murine gene, previously notated as 2300002D11Rik, con-tains a presumed 223 aa long open reading frame in a singleexon. We have isolated a cDNA corresponding to the2300002D11Rik gene from a mouse embryonic library, whichencodes a 223 aa protein, and have named it TRNP. TRNP ishighly conserved in mammals, and this high homology (76%)ranges from placental to marsupial species. Interestingly, no ob-vious ortholog of TRNP could be identified in other vertebrates,suggesting that TRNP represents a relatively “recent” evolu-tionary function that has evolved in mammals.
The protein sequence of TRNP contains several low com-plexity aa stretches. A proline rich region extends from Pro20
to Pro54 at the N-terminal portion of TRNP, and it most prob-ably imposes an extended N-terminal structure on the protein.This proline-rich stretch may also mediate the interaction ofTRNP with other cellular proteins (Khursigara et al., 2005).Adjacent to the proline-rich region resides a glycine/alanine-rich region which, in the murine TRNP, extends from Gly72 toGly94. A similar glycine/alanine-rich domain was found to beessential for the trans-activation function of the Oct-6 tran-scription factor (Meijer et al., 1992).
Enforced expression of the trnp cDNA in mammalian cellsled to the accumulation of an ectopic protein whose size rangedfrom 30 to 36 kDa. This indicates the subjection of the exoge-nous TRNP to extensive posttranslational modification. Simi-lar heterogeneity of the endogenous TRNP protein was alsoseen in xenografts of PC3 prostate carcinoma cells (data not
VOLPE ET AL.338
FIG. 7. TRNP increases the percentage of cells re-siding in the S phase. (A) MCF-7 cells stably trans-fected with the pcDNA3–HA–TRNP expression vec-tor or with the pcDNA3 vector alone, were subjectedto flow-cytometry analysis. Distribution of cells be-tween the different populations is shown. (B) Thetwo cell lines were subjected to BrdU incorporationanalysis and the percentage of cells incorporatingBrdU was determined. The profiles represent one outof three independent experiments that gave similarresults.
FIG. 8. TMF/ARA160 directs the proteasomal degradation ofTRNP. C2C12 cells were cotransfected with the pCMV–Myc–TRNP expression vector together the pcMV–Myc expression vec-tor (lanes 1 and 3), or with the pcDNA–HA–TMF/ARA160 expression vector (lanes 2 and 4). Cells were either left untreatedor were subjected for 5 h to 50 �M of the proteasome inhibitorMG 132. Whole-cell protein lysates were resolved in SDS-PAGEand then reacted with anti-Myc, anti-HA, and antiactin antibodiesusing Western blot analysis. These results represent one out of threeindependent experiments that gave similar results.

shown). Hence, this heterogeneity of TRNP might be linked tospecific growth conditions.
TRNP accumulates mainly in the nucleus of cells. However,amino acids sequence analysis did not reveal the presence of anuclear localization sequence in TRNP. TRNP may thus be co-transported to the nucleus together with another nuclear protein(Soutoglou et al., 2005). Alternatively, TRNP might enter thenucleus by free diffusion, due to its relatively low molecularweight mass (Kurooka and Yokota, 2005).
Careful inspection of immunocytochemical staining imagesindicated the exclusion of TRNP from some intensively stainedpropidium iodide nuclear spots, which most probably representcondensed chromatin structures (Amirand et al., 1998). How-ever, TRNP was not present in the nucleoplasm (NS) but wasconstantly associated with an insoluble nuclear fraction. TRNPwas partially released from the precipitated fraction by 0.5 MNaCl and micrococcal nuclease treatment. The protein was al-most completely dissolved in ionic detergent-like SDS. The saltand nuclease-dependent release of TRNP from the precipitatednuclear fraction could indicate its tight association with chro-matin structures. However, since TRNP is excluded from someintensely condensed chromatin, it might be associated with eu-chromatic, transcriptionally active chromatin. In accordancewith the possible association of TRNP with chromatin, this pro-tein is highly basic protein with a pI of 11.9 (calculated usingthe European Molecular Biology Open Software Suite).
Ectopic expression of TRNP in MCF-7 breast carcinomacells which do not express this protein enhanced their transi-tion from the G0/G1 to the S phase of the cell cycle. This ledmost probably to the accelerated proliferation rate of the trans-fected cells. Thus, TRNP is a nuclear protein that can affectcell-cycle progression in mammalian cells.
TRNP was isolated through its association with TMF/ARA160 in a yeast “two-hybrid” screening system. This sug-gested a functional interaction between these two proteins. In-deed, we showed in the current work that TMF/ARA160 candirect the proteasomal degradation of TRNP. TMF/ARA160was also found to drive the proteasomal degradation of anotherproliferation promoting protein-Stat3 (Perry et al., 2004). Thiscoincides with a presumed tumor suppressive activity of TMF/ARA160 (Perry et al., 2004).
Further experiments should reveal whether the TRNP func-tion is unique to certain cell types in mammals, and whetheranother protein fulfills this function in nonmammalians.
ACKNOWLEDGMENTS
We thank Ms. S. Victor for typing this manuscript. This workwas supported by grants from the Horowitz Foundation andfrom the Prostate Cancer Foundation, UK.
REFERENCES
AMIRAND, C., VIARI, A., BALLINI, J.P., REZAEI, H., BEAUJEAN,N., JULLIEN, D., KAS, E., and DEBEY, P. (1998). Three distinctsub-nuclear populations of HMG-I protein of different properties re-vealed by co-localization image analysis. J. Cell Sci 111, 3551–3561.
BEN-DOR, I., BERN, O., TENNENBAUM, T., and NIR, U. (1999).Cell cycle-dependent nuclear accumulation of the p94fer tyrosine ki-nase is regulated by its NH2 terminus and is affected by kinase do-main integrity and ATP binding. Cell Growth Differ. 10, 113–129.
FRIDMANN-SIRKIS, Y., SINIOSSOGLOU, S., and PELHAM, H.R.(2004). TMF is a golgin that binds Rab6 and influences Golgi mor-phology. BMC Cell Biol. 5, 18.
GARCIA, J.A., OU, S.H., WU, F., LUSIS, A.J., SPARKES, R.S., andGAYNOR, R.B. (1992). Cloning and chromosomal mapping of a hu-man immunodeficiency virus 1 “TATA” element modulatory factor.Proc. Natl. Acad. Sci. USA 89, 9372–9376.
KHURSIGARA, C.M., DE CRESCENZO, G., PAWELEK, P.D., andCOULTON, J.W. (2005). Deletion of the proline-rich region of TonBdisrupts formation of a 2:1 complex with FhuA, an outer membranereceptor of Escherichia coli. Protein Sci. 14, 1266–1273.
KNUESEL, M., WAN, Y., XIAO, Z., HOLINGER, E., LOWE, N.,WANG, W., and LIU, X. (2003). Identification of novel protein–pro-tein interactions using a versatile Mammalian tandem affinity pu-rification expression system. Mol. Cell. Proteomics 2, 1225–1233.
KUROOKA, H., and YOKOTA, Y. (2005). Nucleo-cytoplasmic shut-tling of Id2, a negative regulator of basic helix-loop-helix transcrip-tion factors. J. Biol. Chem. 280, 4313–4320.
MEIJER, D., GRAUS, A., and GROSVELD, G. (1992). Mapping thetransactivation domain of the Oct-6 POU transcription factor. Nu-cleic Acids Res. 20, 2241–2247.
MENDEZ, J., and STILLMAN, B. (2000). Chromatin association ofhuman origin recognition complex, cdc6, and minichromosomemaintenance proteins during the cell cycle: Assembly of prereplica-tion complexes in late mitosis. Mol. Cell. Biol. 20, 8602–8612.
MORI, K., and KATO, H. (2002). A putative nuclear receptor coactivator(TMF/ARA160) associates with hbrm/hSNF2 alpha and BRG-1/hSNF2beta and localizes in the Golgi apparatus. FEBS Lett. 520, 127–132.
PERRY, E., TSRUYA, R., LEVITSKY, P., POMP, O., TALLER, M.,WEISBERG, S., PARRIS, W., KULKARNI, S., MALOVANI, H.,PAWSON, T., et al. (2004). TMF/ARA160 is a BC-box-containing pro-tein that mediates the degradation of Stat3. Oncogene 23, 8908–8919.
PRIEL-HALACHMI, S., BEN-DOR, I., SHPUNGIN, S., TENNEN-BAUM, T., MOLAVANI, H., BACHRACH, M., SALZBERG, S.,and NIR, U. (2000). FER kinase activation of Stat3 is determined bythe N-terminal sequence. J. Biol. Chem. 275, 28902–28910.
RIGAUT, G., SHEVCHENKO, A., RUTZ, B., WILM, M., MANN,M., and SERAPHIN, B. (1999). A generic protein purificationmethod for protein complex characterization and proteome explo-ration. Nat. Biotechnol. 17, 1030–1032.
SCHWARTZ, Y., BEN-DOR, I., NAVON, A., MOTRO, B., and NIR, U.(1998). Tyrosine phosphorylation of the TATA element modulatory fac-tor by the FER nuclear tyrosine kinases. FEBS Lett. 434, 339–345.
SOUTOGLOU, E., DEMENY, M.A., SCHEER, E., FIENGA, G., SAS-SONE-CORSI, P., and TORA, L. (2005). The nuclear import ofTAF10 is regulated by one of its three histone fold domain-contain-ing interaction partners. Mol. Cell. Biol. 25, 4092–4104.
THE FANTOM CONSORTIUM AND THE RIKEN GENOME EX-PLORATION RESEARCH GROUP PHASE I & II TEAM. (2002).Analysis of the mouse transcriptome based on functional annotationof 60,770 full-length cDNAs. Nature 420, 563–573.
YANAI, I., BENJAMIN, H., SHMOISH, M., CHALIFA-CASPI, V.,SHKLAR, M., OPHIR, R., BAR-EVEN, A., HORN-SABAN, S.,SAFRAN, M., DOMANY, E., et al. (2005). Genome-wide midrangetranscription profiles reveal expression level relationships in humantissue specification. Bioinformatics 21, 650–659.
Address reprint requests to:Uri Nir, Ph.D.
Faculty of Life SciencesBar-Ilan UniversityRamat-Gan 52900
Israel
E-mail: [email protected]
Received for publication December 29, 2005; received in re-vised form January 25, 2006; accepted March 6, 2006.
A NOVEL CONSERVED MAMMALIAN GENE 339
Copyright © 2022 FDOKUMEN




















