
Involvement of the telencephalon in spaced-trial avoidance learning in the goldfish ( Carassius...
8
Involvement of the telencephalon in spaced-trial avoidance learning in the goldfish (Carassius auratus) Manuel Portavella a, * , Cosme Salas a , Juan P. Vargas a,b , Mauricio R. Papini c a Laboratorio de Psicobiologı ´a, Departamento de Psicologı ´a Experimental, Facultad de Psicologı ´a, Universidad de Sevilla, c/ Camilo Jose ´ Cela, s/n 41018, Sevilla, Spain b Department of Psychology, Bowling Green State University, Bowling Green, OH 43403, USA c Department of Psychology, Texas Christian University, Box 298920, Fort Worth, TX 76129, USA Received 22 January 2003; received in revised form 28 May 2003; accepted 19 June 2003 Abstract Goldfish (Carassius auratus) received escape– avoidance training in a shuttle-response situation at a rate of a single trial per day. Widely spaced training evaluates the ability of a discriminative stimulus to control an avoidance response in the absence of stimulus carry-over effects from prior recent trials. In Experiment 1, master goldfish exhibited significantly faster avoidance learning than yoked controls. The results suggest that the shuttle response was instrumentally acquired. Experiment 2 demonstrated a significant deficit in the acquisition of avoidance behavior following ablation of the telencephalon. The implications of spaced-trial, telencephalon-dependent avoidance learning, as demonstrated in these experiments for the first time, are discussed in the context of comparative research on instrumental learning in goldfish. These results provide further support for the hypothesis that the fish telencephalon contains an emotional system that is critical for fear conditioning. D 2003 Elsevier Inc. All rights reserved. Keywords: Avoidance learning; Emotion; Fear; Fish; Memory systems; Spaced-trial training; Telencephalon 1. Introduction Learning theorists have traditionally assumed that medi- ational states induced by aversive reinforcements such as electric shock (i.e., fear) and the surprising omission of appetitive reinforcers such as food (i.e., frustration) affect behavior via similar mechanisms [1–4]. Gray [2] refers to this as the fear = frustration hypothesis. For example, partial food reinforcement and continuous food reinforcement mixed with a partial shock punishment schedule both in- crease resistance to extinction in rats relative to just contin- uous food reinforcement [5]. In fact, many of the behavioral (e.g., aggressive behavior) and physiological consequences (e.g., pituitary–adrenal activation) of aversive events are similar to those that occur following surprising reward omissions (for a review, see Ref. [6]). Such similarities suggest that at least some of the mechanisms underlying the conditioning of fear and frustration should covary in any given species. Teleost fish provide an opportunity to explore the fear = frustration hypothesis in a comparative context. On the one hand, there is good evidence that avoidance learning is based on the acquisition of a mediational state of fear in the goldfish. For example, prior Pavlovian pairings of a warning stimulus (WS) with shock facilitate subsequent avoidance acquisition relative to groups pre- exposed to escape conditioning, shock only, and naive controls [7]. Off-avoidance baseline, Pavlovian training also endows the WS with the power to facilitate avoidance responding trained separately [8]. Following Mowrer [3], Zhuikov et al. [9] hypothesized that the Pavlovian contin- gency (either in purely Pavlovian situations or that em- bedded in the avoidance situation) leads to the acquisition of an internal state of fear. Conditioned fear, in turn, contributes to the development of the instrumental stimu- lus – response association because of a reduction in fear that follows the shuttle response. The model of Zhuikov et al. [9] provides reasonably good quantitative predictions of avoidance learning in goldfish under a variety of condi- 0031-9384/$ – see front matter D 2003 Elsevier Inc. All rights reserved. doi:10.1016/S0031-9384(03)00208-7 * Corresponding author. Tel.: +34-954-55-44-01; fax: +34-954-55-17- 84. E-mail address: [email protected] (M. Portavella). Physiology & Behavior 80 (2003) 49 – 56
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Involvement of the telencephalon in spaced-trial avoidance learning in the goldfish ( Carassius...

Physiology & Behavior 80 (2003) 49–56
Involvement of the telencephalon in spaced-trial avoidance learning
in the goldfish (Carassius auratus)
Manuel Portavellaa,*, Cosme Salasa, Juan P. Vargasa,b, Mauricio R. Papinic
aLaboratorio de Psicobiologıa, Departamento de Psicologıa Experimental, Facultad de Psicologıa, Universidad de Sevilla,
c/ Camilo Jose Cela, s/n 41018, Sevilla, SpainbDepartment of Psychology, Bowling Green State University, Bowling Green, OH 43403, USA
cDepartment of Psychology, Texas Christian University, Box 298920, Fort Worth, TX 76129, USA
Received 22 January 2003; received in revised form 28 May 2003; accepted 19 June 2003
Abstract
Goldfish (Carassius auratus) received escape–avoidance training in a shuttle-response situation at a rate of a single trial per day. Widely
spaced training evaluates the ability of a discriminative stimulus to control an avoidance response in the absence of stimulus carry-over effects
from prior recent trials. In Experiment 1, master goldfish exhibited significantly faster avoidance learning than yoked controls. The results
suggest that the shuttle response was instrumentally acquired. Experiment 2 demonstrated a significant deficit in the acquisition of avoidance
behavior following ablation of the telencephalon. The implications of spaced-trial, telencephalon-dependent avoidance learning, as
demonstrated in these experiments for the first time, are discussed in the context of comparative research on instrumental learning in goldfish.
These results provide further support for the hypothesis that the fish telencephalon contains an emotional system that is critical for fear
conditioning.
D 2003 Elsevier Inc. All rights reserved.
Keywords: Avoidance learning; Emotion; Fear; Fish; Memory systems; Spaced-trial training; Telencephalon
1. Introduction
Learning theorists have traditionally assumed that medi-
ational states induced by aversive reinforcements such as
electric shock (i.e., fear) and the surprising omission of
appetitive reinforcers such as food (i.e., frustration) affect
behavior via similar mechanisms [1–4]. Gray [2] refers to
this as the fear = frustration hypothesis. For example, partial
food reinforcement and continuous food reinforcement
mixed with a partial shock punishment schedule both in-
crease resistance to extinction in rats relative to just contin-
uous food reinforcement [5]. In fact, many of the behavioral
(e.g., aggressive behavior) and physiological consequences
(e.g., pituitary–adrenal activation) of aversive events are
similar to those that occur following surprising reward
omissions (for a review, see Ref. [6]). Such similarities
suggest that at least some of the mechanisms underlying the
0031-9384/$ – see front matter D 2003 Elsevier Inc. All rights reserved.
doi:10.1016/S0031-9384(03)00208-7
* Corresponding author. Tel.: +34-954-55-44-01; fax: +34-954-55-17-
84.
E-mail address: [email protected] (M. Portavella).
conditioning of fear and frustration should covary in any
given species.
Teleost fish provide an opportunity to explore the
fear = frustration hypothesis in a comparative context. On
the one hand, there is good evidence that avoidance
learning is based on the acquisition of a mediational state
of fear in the goldfish. For example, prior Pavlovian
pairings of a warning stimulus (WS) with shock facilitate
subsequent avoidance acquisition relative to groups pre-
exposed to escape conditioning, shock only, and naive
controls [7]. Off-avoidance baseline, Pavlovian training
also endows the WS with the power to facilitate avoidance
responding trained separately [8]. Following Mowrer [3],
Zhuikov et al. [9] hypothesized that the Pavlovian contin-
gency (either in purely Pavlovian situations or that em-
bedded in the avoidance situation) leads to the acquisition
of an internal state of fear. Conditioned fear, in turn,
contributes to the development of the instrumental stimu-
lus–response association because of a reduction in fear
that follows the shuttle response. The model of Zhuikov et
al. [9] provides reasonably good quantitative predictions of
avoidance learning in goldfish under a variety of condi-

M. Portavella et al. / Physiology & Behavior 80 (2003) 49–5650
tions. Furthermore, the complete ablation of the telenceph-
alon impairs avoidance learning in goldfish [8,10–14], a
result analogous to that observed in mammals with lesions
in various telencephalic structures, particularly the amyg-
dala [15–18]. However, the use of relatively massed
conditions of training (i.e., multiple trials per session) in
the cited experiments implies that the animal is responding
to the WS after having recently experienced electric shocks
and performed the shuttle response. Such salient events
may facilitate the acquisition of fear in their own right,
rendering ambiguous the theoretical claim that fear is
induced by the WS. Spaced-trial procedures (e.g., one trial
per day) can contribute to determine whether the WS can
elicit fear in the absence of carry-over effects from
previous trials.
On the other hand, the fear = frustration hypothesis cannot
explain the results of a variety of appetitive experiments
involving surprising reward omissions. A wide range of
experiments with goldfish and other nonmammalian verte-
brates, trained under widely spaced conditions (e.g., one trial
per day), have thus far failed to provide any evidence that
their instrumental behavior can be modulated by the associa-
tive reinstatement of prior frustrative outcomes [19–21].
Instead, the results of such experiments can be understood
in terms of a simple learning rule according to which
reinforcement increases the strength of the instrumental
response, whereas nonreinforcement decreases it. It seems
possible, therefore, that just as conditioned frustration fails to
control widely spaced instrumental performance in goldfish,
and a similarly spaced protocol of avoidance training may fail
to induce a conditioned response of fear. By contrast, widely
spaced acquisition of avoidance behavior would suggest a
dissociation between the basic underlying mechanisms of
appetitive (i.e., frustration) and aversive (i.e., fear) learning in
the goldfish [1–4].
The following experiments were designed to answer the
following questions. First, can goldfish learn an avoidance
response in a spaced-trial situation? Second, is such spaced-
trial avoidance learning dependent on the integrity of the
telencephalon?
2. Experiment 1: Master–yoked acquisition
The goal of Experiment 1 was to validate the spaced-
training procedure and to demonstrate the instrumental
nature of avoidance learning under such conditions using
a master–yoked design. Master–yoked pairs of goldfish
received one daily trial (one trial per day, one trial per
session) of shuttle-avoidance training. A visual cue (the WS)
was turned on in the tanks of both master and yoked
animals. A shuttle response by the master animal (swim-
ming over a barrier) during the presentation of the WS
turned it off and prevented the shock from being adminis-
tered in both the master’s and yoked’s tanks. Failure to
respond by the master led to the administration of an electric
shock also in both tanks. A similar master–yoked study
involving multiple trials per session demonstrated that the
shuttle response is acquired to a higher level by master
animals experiencing an instrumental contingency than by
yoked animals exposed to purely Pavlovian contingencies
[14].
2.1. Materials and methods
2.1.1. Subjects
Sixteen experimentally naive goldfish (Carassius aura-
tus), purchased from a local aquarium in Seville, served as
subjects. Animals were 9–12 cm in body length and lived in
pairs in tanks measuring 42 cm in length, 26 cm in width,
and 15 cm in height. Living tanks were cleaned every other
day and the water was kept at a constant temperature of 21
jC. Water was continuously aerated and filtered. The
aquarium room was subject to a 14:10-h light:dark cycle
(light from 07:00 to 21:00 h). Three pellets of Tetra-pond
fish were provided daily, at least 60 min after the training
session ended.
2.1.2. Apparatus
Goldfish received avoidance training in conventional
shuttle tanks. Two pairs of identical tanks were used, built
entirely with glass plates and with identical internal dimen-
sions (50� 14� 25 cm). Each box was filled with continu-
ously aerated water to a depth of 9.5 cm. In the center of each
box, there was a trapezoidal barrier measuring 10 cm at the
top, 18 cm at its base, and 7.5 cm in height. A shuttle
response required that the fish swam over the barrier by
tilting its body on one side. This response was recorded by
photocells located at each side of the barrier and 1.5 cm
above the barrier. The end glass plates on each extreme were
translucent and allowed for the presentation of a green light
(10 W) used as the WS. The lateral walls of the tank were
covered with metallic plates connected to a conventional
stimulator. Electric shocks (0.39 V ac/cm, 50 Hz sinusoidal
stimulation) were delivered through these plates by a stim-
ulator. A computer controlled all events and recorded the
response latency, that is, the time from the turning on of the
warning visual stimulus to either the occurrence of the
shuttle response, or the termination of the trial.
2.1.3. Training procedure
Prior to the training period, there were three sessions of
habituation to the shuttle tank in which no WS or shocks
were delivered. In the first session, animals were placed in
the shuttle tank during 30 min with the water level set at 6 cm
above the barrier. The following two sessions were 5 min
long and the water level was dropped to 2 cm above the
barrier. Animals were then randomly assigned to one of two
groups, master or yoked (n = 8), and administered one
training trial per day (for 40 training days).
During training, animals were transferred in pairs from
their home tanks to the training tanks. Each particular shuttle
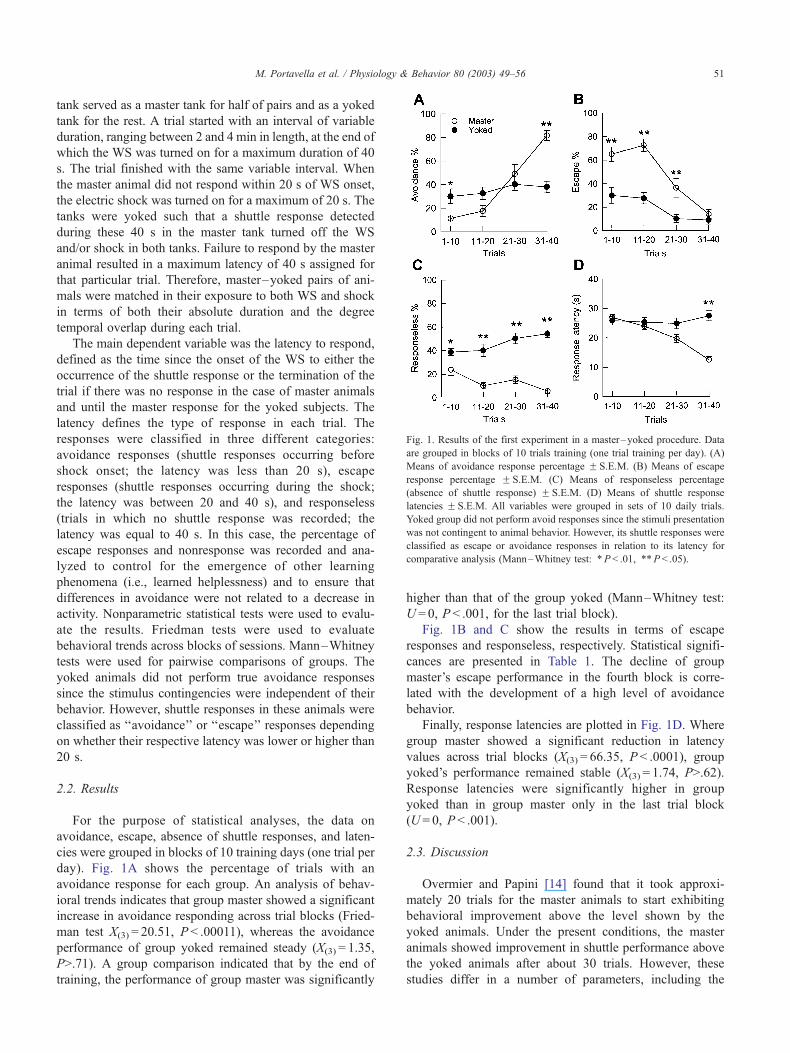
Fig. 1. Results of the first experiment in a master–yoked procedure. Data
are grouped in blocks of 10 trials training (one trial training per day). (A)
Means of avoidance response percentage F S.E.M. (B) Means of escape
response percentage F S.E.M. (C) Means of responseless percentage
(absence of shuttle response) F S.E.M. (D) Means of shuttle response
latencies F S.E.M. All variables were grouped in sets of 10 daily trials.
Yoked group did not perform avoid responses since the stimuli presentation
was not contingent to animal behavior. However, its shuttle responses were
classified as escape or avoidance responses in relation to its latency for
comparative analysis (Mann–Whitney test: *P< .01, **P < .05).
M. Portavella et al. / Physiology & Behavior 80 (2003) 49–56 51
tank served as a master tank for half of pairs and as a yoked
tank for the rest. A trial started with an interval of variable
duration, ranging between 2 and 4 min in length, at the end of
which the WS was turned on for a maximum duration of 40
s. The trial finished with the same variable interval. When
the master animal did not respond within 20 s of WS onset,
the electric shock was turned on for a maximum of 20 s. The
tanks were yoked such that a shuttle response detected
during these 40 s in the master tank turned off the WS
and/or shock in both tanks. Failure to respond by the master
animal resulted in a maximum latency of 40 s assigned for
that particular trial. Therefore, master–yoked pairs of ani-
mals were matched in their exposure to both WS and shock
in terms of both their absolute duration and the degree
temporal overlap during each trial.
The main dependent variable was the latency to respond,
defined as the time since the onset of the WS to either the
occurrence of the shuttle response or the termination of the
trial if there was no response in the case of master animals
and until the master response for the yoked subjects. The
latency defines the type of response in each trial. The
responses were classified in three different categories:
avoidance responses (shuttle responses occurring before
shock onset; the latency was less than 20 s), escape
responses (shuttle responses occurring during the shock;
the latency was between 20 and 40 s), and responseless
(trials in which no shuttle response was recorded; the
latency was equal to 40 s. In this case, the percentage of
escape responses and nonresponse was recorded and ana-
lyzed to control for the emergence of other learning
phenomena (i.e., learned helplessness) and to ensure that
differences in avoidance were not related to a decrease in
activity. Nonparametric statistical tests were used to evalu-
ate the results. Friedman tests were used to evaluate
behavioral trends across blocks of sessions. Mann–Whitney
tests were used for pairwise comparisons of groups. The
yoked animals did not perform true avoidance responses
since the stimulus contingencies were independent of their
behavior. However, shuttle responses in these animals were
classified as ‘‘avoidance’’ or ‘‘escape’’ responses depending
on whether their respective latency was lower or higher than
20 s.
2.2. Results
For the purpose of statistical analyses, the data on
avoidance, escape, absence of shuttle responses, and laten-
cies were grouped in blocks of 10 training days (one trial per
day). Fig. 1A shows the percentage of trials with an
avoidance response for each group. An analysis of behav-
ioral trends indicates that group master showed a significant
increase in avoidance responding across trial blocks (Fried-
man test X(3) = 20.51, P < .00011), whereas the avoidance
performance of group yoked remained steady (X(3) = 1.35,
P>.71). A group comparison indicated that by the end of
training, the performance of group master was significantly
higher than that of the group yoked (Mann–Whitney test:
U = 0, P < .001, for the last trial block).
Fig. 1B and C show the results in terms of escape
responses and responseless, respectively. Statistical signifi-
cances are presented in Table 1. The decline of group
master’s escape performance in the fourth block is corre-
lated with the development of a high level of avoidance
behavior.
Finally, response latencies are plotted in Fig. 1D. Where
group master showed a significant reduction in latency
values across trial blocks (X(3) = 66.35, P < .0001), group
yoked’s performance remained stable (X(3) = 1.74, P>.62).
Response latencies were significantly higher in group
yoked than in group master only in the last trial block
(U = 0, P < .001).
2.3. Discussion
Overmier and Papini [14] found that it took approxi-
mately 20 trials for the master animals to start exhibiting
behavioral improvement above the level shown by the
yoked animals. Under the present conditions, the master
animals showed improvement in shuttle performance above
the yoked animals after about 30 trials. However, these
studies differ in a number of parameters, including the

Table 1
Experiment 1. Master–yoked acquisition
Group master versus yoked (Mann–Whitney test analyses)
Response kind Trial blocks
1–10 11–20 21–30 31–40
Escape (%) U= 5;
P=.004
U = 0;
P=.001
U= 7;
P=.007
U = 22.5;
P=.29
Responseless (%) U= 11;
P=.024
U = 1;
P=.001
U= 0.5;
P=.001
U = 0;
P=.001
Statistical analysis of differences of escape and responseless percentage
between group master and yoked.
M. Portavella et al. / Physiology & Behavior 80 (2003) 49–5652
maximum length of the WS (10 vs. 40 s in the present
experiment) and, of course, the administration of multiple
trials per session versus a single trial per session. In a single-
trial situation, the behavior is relatively uncontaminated by
carry-over effects from prior trials, including the WS, the
electric shock, response-elicited cues, and more extensive
exposure to the training context. Some, or even all, of these
cues may facilitate memory retrieval during the course of
acquisition and thus account for relatively better perfor-
mance [22]. The evidence presented here suggests that
goldfish can reactivate whatever information is necessary
for performing the avoidance response (e.g., mediational
state of fear [8]) essentially on the basis of the discrimina-
tive stimulus.
3. Experiment 2: Bilateral telencephalic ablation
If this spaced-trial instance of avoidance learning was
dependent on the same mechanisms that operate under
massed-trial conditions, then it should be vulnerable to the
bilateral ablation of the telencephalon [12]. In Experiment 2,
the space-trial performance of goldfish with telencephalic
lesions was compared to that of sham-operated and intact
controls. In addition, a group exposed only to the WS
provided a nonshocked baseline condition to determine the
extent to which the lesions impaired spontaneous levels of
shuttling behavior.
3.1. Materials and method
3.1.1. Subject and apparatus
Twenty-three experimentally naive goldfish served as
subjects in the present experiment. These animals were
between 9 and 12 cm in body length. They were obtained
from the same local source and maintained as described in the
previous experiment. The same living tanks and shuttle
avoidance tanks described before were also used in the
present experiment.
3.1.2. Surgical and histological procedures
Animals were anesthetized by immersion in a solution of
tricaine methanosulfonate (1:20,000). When movement had
ceased, the animal was placed in the surgical chamber where
it remained partially bathed in water with a constant
concentration of anesthetic. Lateral holders that maintained
the body still during surgery fixed the animal into position.
An adjustable tube was inserted into the animal’s mouth and
connected to a pump that provided a constant flow of water.
The concentration of the anesthetic in the water was kept at
1:20,000 during surgery.
The dorsal skin and skull were carefully removed and the
underlying fatty tissue removed by aspiration. The exposed
telencephalon was aspirated with a micropipette connected
to a manual vacuum system. Surgery was performed under
visual inspection by means of a binocular microscope.
Following the ablation, the piece of skull was replaced in
its original position and fixed with cyanocrilate glue. The
fish was returned to its home tank for a recovery period of 5
days. Sham operations were performed exactly as described,
except that the nervous tissue was not injured.
At the end of the experiment, fish with telencephalic
ablations and sham operations were deeply anesthetized
(1:5000) and perfused with 50 ml of 0.9% saline solution,
followed by 125 ml of fixative solution (10% formalin in
phosphate buffer 0.1 M, pH 7.4). The brains were removed
from the skull, inspected for the preliminary evaluation of
the ablation, and cut in 50 Am thick transversal sections for
histological analysis.
3.1.3. Training procedure
There were four groups in the present experiment. The
animals in the group ablation (n = 8) received a bilateral
ablation of the telencephalon before the start of the training,
whereas those in group sham (n = 4) received a sham
operation. The goldfish assigned to group intact (n = 4) were
not subjected to any surgery. Finally, the goldfish assigned to
group S-only (n = 8) were also intact but received different
training These animals were exposed to sessions that were
similar in every respect to the avoidance sessions of the other
goldfish, except no shock was ever delivered. This condition
was introduced to assess the extent to which the change in
illumination per se was reinforcing to these animals, as well
as to provide a no shock baseline to evaluate the extent of the
lesion effect. All training parameters were the same as in
Experiment 1. All animals were trained for 30 days (one trial
per day). The same dependent variables described in Exper-
iment 1 were recorded. Friedman tests were computed to
analyze behavioral trends across trial blocks. At each trial
block, overall group comparisons were based on the Krus-
kal–Wallis test and pairwise comparisons on the Mann–
Whitney test.
3.2. Results
One goldfish in group ablation died in the course of the
experiment, leaving the group with an n = 7. Optical inspec-
tion of the ablated brains indicated that telencephalic abla-
tions were complete (Fig. 2). The perfusion and brain
extraction were carried out 40 days after surgery (5 postsur-

Fig. 2. Nissl stained of sections of sham (A) and telencephalic (C). Level of brain sections brains (B). Abbreviations: Cer, cerebellum; Dc, dorsocentral telen-
cephalon; Dd, dorsodorsal telencephalon; Dl, dorsolateral telencephalon; Dm, dorsomedial telencephalon; Olf T, olfactory tracts; ON, optic nerves; Op tec, optic
tectum; PO, preoptic nucleus; PP, periventricular preoptic nucleus; Tel, telencephalon; VN, vagal nucleus; LH, lateral hypothalamus; V, ventral telencephalon.
Table 2
Experiment 2. Bilateral telencephalic ablation
Animals Avoidance (%)
Trials: 1–10 Trials: 11–20 Trials: 21–30
Sham 1 30 80 80
Sham 2 50 80 80
Sham 3 10 70 90
Sham 4 30 40 100
Intact 1 50 90 90
Intact 2 60 70 100
Intact 3 30 10 100
Intact 4 50 80 70
Individual data of avoidance percentage from sham and intact animals of
group control.
M. Portavella et al. / Physiology & Behavior 80 (2003) 49–56 53
gery days, 30 training days, and 4–6 posttraining days). The
ablated brains presented the aspect of a swelled cylinder on
the optic nerves. This cylinder composed principally by
diencephalon is the product of a probable process of regen-
eration and fusion of two hemispheres. The good perfusion of
this brain part is difficult.
The histological analysis showed no damage to the
preoptic area and optic tracts. The optic tectum was also
spared. None of the sham animals exhibited any evidence of
damage to the telencephalon or optic tectum.
In this experiment, also, the percentage of escape re-
sponses and nonresponse were recorded and analyzed to
control the effects of the telencephalon ablation on the motor
capabilities of operated animals and to guarantee that the
differences in avoidance levels were not related with this
possible interference.
No statistically significant differences were observed
between sham and intact animals in responseless (Usz 3,
Ps=.13), escape (Usz 4, Ps>.23), and avoidance responses
(Mann–Whitney test: Usz 3, Ps>.13). Individual data of
avoidance percentages are showed in Table 2. Differences
were found only in the second block of latency values (first:
U = 2, P>.08; second: U = 0, P < .021; third: U = 2, P>.08).
The sham and intact groups showed similar learning levels
in avoidance, escape, and responseless; therefore, sham and
intact data are pooled into group control (n = 8) for statistical
analyses.
The main results of this experiment are presented in Fig.
3. Analyses of the behavioral trends exhibited by each of
these groups in each dependent variable yielded the follow-
ing results. The avoidance performance of these groups is
presented in Fig. 3A. Group control showed a significant
and progressive increase in avoidance behavior across trial
blocks (Friedman test: X(2) = 10.18, P < .007). In contrast,
groups ablation and S-only showed a low level of avoidance
responses in all trial blocks, but whereas the former group
was steady (X(2) = 0.5, P>.77), the latter exhibited a signif-
icant increase across trial blocks (X(2) = 6.44, P < .041).
Groups were also compared in terms of their avoidance
performance at the level of each trial block. Significant
group differences were found in all trial blocks (Kruskal–
Wallis tests: Xs(2)z 9.28, Ps < .01). Pairwise comparisons
indicated that the performance of group control was signif-
icantly higher than that of group ablation (Mann–Whitney
test: UsV 6, Ps < .01) and that of group S-only at each trial
block (Mann–Whitney test: UsV 10.5, Ps < .05). There
were no differences between groups ablation and S-only
(Mann–Whitney test: Usz 18.5, Ps>.2).
The escape performance of all the groups is plotted in
Fig. 3B and the trials without response in Fig. 3C. Statistical
significances are shown in Table 3.
Finally, the average response latencies for each group are
presented in Fig. 3D. In terms of behavioral trends, group

Table 3
Experiment 2. Bilateral telencephalic ablation
Response kind Trial blocks
1–10 11–20 21–30
All groups (Kruskal–Wallis test analyses)
Escape (%) X(2) = 15.52;
P=.0
X(2) = 9.46;
P=.007
X(2) = 12.9;
P=.002
Responseless (%) X(2) = 15.42;
P=.0
X(2) = 16.51;
P=.0
X(2) = 18.76;
P=.0
Group S-only versus control (Mann–Whitney test analyses)
Escape (%) U = 0.5;
P=.001
U = 29;
P=.746
U= 19.5;
P=.178
Responseless (%) U = 0;
P=.001
U = 0;
P=.001
U= 0;
P=.001
Group S-only versus telencephalic (Mann–Whitney test analyses)
Escape (%) U = 1.5;
P=.001
U = 4;
P=.005
U= 5;
P=.007
Responseless (%) U = 0;
P=.001
U = 0;
P=.001
U= 0;
P=.001
Group control versus telencephalic (Mann–Whitney test analyses)
Escape (%) U = 14.5;
P=.108
U = 5.5;
P=.008
U= 0.5;
P=.001
Responseless (%) U = 26.5;
P=.856
U = 24.5;
P=.629
U= 20;
P=.117
M. Portavella et al. / Physiology & Behavior 80 (2003) 49–5654
control demonstrated a significant decrease in response
latency across trial blocks (Friedman test: X(2) = 49.17,
P < .0001). Group ablation also exhibited a significance
decrease in latency scores (X(2) = 6.26, P < .05), whereas
group S-only showed and minor but significant decrease in
response latency in the last block of trials (X(2) = 10.225,
P < .01). The analyses of latency performance indicated
significant group differences at each trial block (Kruskal–
Wallis tests: X(2)z 73.66, Ps < .001). Pairwise comparisons
showed that groups control and ablation exhibited differences
in the second and third trial blocks (first: U = 14, P>.1;
second: U = 0, P < .001; third: U = 0, P < .001), their perfor-
mance was different that of S-only group in all trial blocks
(control vs. S-only: UsV 1, Ps < .01; ablation vs. S-only:
UsV 3, Ps < .01).
3.3. Discussion
The impairment of avoidance learning in telencephalon-
ablated goldfish puts them at similar level to that of
goldfish exposed to S-only training. Moreover, previous
experiments showed that telencephalon-ablated goldfish
can be conditioned in a food-reinforced color discrimina-
tion task [23–25]. Therefore, such an impairment cannot
Results from statistical analysis of the escape and responseless percentagebetween group control, ablation, and S-only.
Fig. 3. Results from the second experiment. Data are grouped in blocks of
10 trials training (one trial training per day). (A) Means of avoidance
response percentage F S.E.M. (B) Means of escape response percentage
F S.E.M. (C) Means of responseless percentage (absence of shuttle
response) F S.E.M. (D) Means of shuttle response latencies F S.E.M. All
variables were grouped in sets of 10 daily trials. S-only group did not
perform avoid and escape responses since unconditioned stimulus was not
present. However, its shuttle responses were classified as escape or
avoidance responses in relation to its latency for comparative analysis
(Kruskal–Wallis test: * *P < .01).
be attributed to an interference in the goldfish’s visual
capacities.
Ablation of the telencephalon impairs acquisition of the
spaced-trial shuttle avoidance response just as it does in the
massed trial situation [12]. This similarity is consistent with
the hypothesis that these two training protocols engage
similar telencephalic mechanisms.
4. General discussion
These experiments demonstrate robust, telencephalon-
dependent, spaced-trial avoidance learning in goldfish. To
the best of our knowledge, this is the first demonstration of
spaced-trial (i.e., one trial per day) avoidance learning in any
species. This result has implications for the fear = frustration
hypothesis [2] when integrated with the results of appetitive
spaced-trial experiments. In mammals, such spaced-trial
experiments with food reinforcers involving the surprising
omission of rewards, or the surprising reduction in reward
magnitude or quality, lead to emotional responses collective-
ly referred to as frustration [1,2]. Goldfish, as well as other
species of bony fish, amphibians, turtles, and pigeons, have
thus far provided no evidence that requires the assumption of
an emotional mediational state of frustration [19–21]. For
example, goldfish trained with large rewards acquire a
swimming alleyway response faster than goldfish trained
with small rewards; however, when shifted from the large to
the small reward magnitude, they show little or no change in

M. Portavella et al. / Physiology & Behavior 80 (2003) 49–56 55
behavior [26]. That is, they provide no evidence of succes-
sive negative contrast—a deterioration of behavior that
follows a downward shift in reward magnitude in rats [27].
Such frustration-related instrumental effects in rats and other
mammals can be reduced by treatment with benzodiazepine
anxiolytics [28,29], induce pituitary–adrenal activation
[30–32], and support escape conditioning [33]. These are
properties shared by frustration- and fear-inducing situations
that suggest some degree of overlap in underlying mecha-
nisms. The fact that spaced-trial avoidance learning is readily
obtained in a species—the goldfish—that shows no spaced-
trial contrast and related phenomena suggests a fundamental
asymmetry in the evolution of the neural systems involved in
the associative reinstatement of fear and frustration. The
potential dissociation between fear and frustration in the
goldfish could be approached in at least two directions. One
possibility is that there is a dissociation of the basic mech-
anisms subserving appetitive and aversive learning in gold-
fish. The other possibility is that the experiments with
goldfish involving reward omission manipulation were not
particularly sensitive to detect the effects of frustration on
instrumental behavior.
The present results are also important to understand the
deficit in avoidance learning induced by telencephalic
lesions. The spacing of the trials leaves little doubt that
goldfish can retrieve the appropriate information to perform
the shuttle response purely on the basis of the WS. By
contrast, in relatively mass conditions of training, there are
additional sources of control over the behavior, including
the recent presentation of shocks and the performance of the
shuttle response. Recent powerful events like these may
enhance retrieval and allow the information to carry-over
across trials. The present results suggest that telencephalic
lesions affect avoidance learning by impairing the retrieval
of an anticipatory fear response by the WS or by interfering
with the ability of an internal state of fear to induce an
avoidance response. In light of the success of the present
procedure, it would be interesting to determine if transfer of
control from a Pavlovian signal for shock to the shuttle
avoidance response would occur in ablated goldfish trained
under spaced conditions. A previous experiment showed
that lesioned animals shuttled in the presence on the
Pavlovian signal trained off-baseline just as much as normal
controls [8]. However, testing of the Pavlovian signal was
carried out after five avoidance conditioning trials involving
both shuttling responses and exposure to the electric shock.
The spaced-trial procedure used in the present experi-
ments can contribute to clarify the role of the telencephalon
in avoidance learning. The conditioned emotional response
in mammals is acquired in a few trials. The spaced-trial
avoidance learning procedure used in the present experi-
ments shows a similar effectiveness. For example, 10 trials
were enough to acquire the avoidance response in Experi-
ment 2. Additional experimental evidence shows that the
total or partial ablation of the telencephalon in goldfish
produces specific impairments in spatial learning
[23,25,34–36]. The specificity of these effects is similar
to what has been described in mammals with hippocampal
lesions [17,18,37]. These results suggest the presence of a
telencephalon-based spatial learning system in the goldfish.
Recent work provided evidence of the presence of two
memory systems in the teleost telencephalon based on
discrete pallial regions: dorsomedial (emotional system)
and dorsolateral (spatial system) [38,39]. The absence of
carry-over effects, the rapid acquisition of avoidance
responses, and the impairment exhibited by such avoidance
behavior in animals with telencephalic ablation provide
further support for the hypothesis that the fish telencepha-
lon contains an emotional system that is critical for fear
conditioning.
Acknowledgements
This research was supported by grants from the Spanish
D.G.E.S. (PB.96-1334) and Junta de Andalucıa. The
participation of M.R.P. in the project was supported by
grants from Junta de Andalucıa, Spain. We thank G.
Labrador and M. T. Gutierrez for technical assistance.
References
[1] Amsel A. Frustration theory. Cambridge (UK): Cambridge Univ
Press; 1992.
[2] Gray JA. The psychology of fear and stress. Cambridge (UK): Cam-
bridge Univ Press; 1987.
[3] Mowrer OH. On the dual nature of learning—a re-interpretation of
‘‘conditioning’’ and ‘‘problem solving’’. Harvard Educ Rev 1947;17:
102–48.
[4] Wagner AR. Frustrative non-reward: a variety of punishment? In:
Campbell BA, Church RM, editors. Punishment and aversive behav-
ior. New York: Appleton-Century-Crofts; 1969. p. 157–81.
[5] Brown RT, Wagner AR. Resistance to punishment and extinction
following training with shock or non-reinforcement. J Exp Psychol
1964;68:503–7.
[6] Papini MR, Dudley RT. Consequences of surprising reward omis-
sions. Rev Gen Psychol 1997;1:175–97.
[7] Gallon RL. Effects of pre-training with fear and escape conditioning
on shuttle-box avoidance acquisition by goldfish. Psychol Rep 1972;
31:919–24.
[8] Overmier JB, Starkman N. Transfer of control of avoidance behavior
in normal and telencephalon ablated goldfish (Carassius auratus).
Physiol Behav 1974;12:605–8.
[9] Zhuikov AY, Couvillon PA, Bitterman ME. Quantitative two-process
analysis of avoidance conditioning in goldfish. J Exp Psychol, Anim
Behav Processes 1994;20:32–43.
[10] Flood NB, Overmier JB, Savage GE. The teleost telencephalon and
learning: an interpretative review of data and hypotheses. Physiol
Behav 1976;16:783–98.
[11] Flood NB, Overmier JB. Learning in teleost fish: role of the telence-
phalon. In: Laming PR, editor. Brain mechanisms of behavior in lower
vertebrates. Cambridge: Cambridge Univ. Press; 1981. p. 259–79.
[12] Hainsworth FR, Overmier JB, Snowdon CT. Specific and permanent
deficits in instrumental avoidance learning following forebrain abla-
tion in the goldfish. J Comp Physiol Psychol 1967;63:111–6.
[13] Overmier JB, Hollis KL. Fish in the think tank: learning, memory and

M. Portavella et al. / Physiology & Behavior 80 (2003) 49–5656
integrated behavior. In: Kesner RP, Olton DS, editors. Neurobiology
of comparative cognition. Hillsdale (NJ): Erlbaum; 1990. p. 204–36.
[14] Overmier JB, Papini MR. Factors modulating the effects of teleost
telencephalon ablation on retention, relearning, and extinction of in-
strumental avoidance behavior. Behav Neurol 1986;100:190–9.
[15] Aggleton JP. The amygdala: neurobiological aspects of emotion,
memory, and mental dysfunction. New York: Wiley-Liss; 1992.
[16] Aggleton JP. The amygdala: a functional analysis. Oxford: Oxford
Univ. Press; 2001.
[17] Eichenbaum H, Otto T, Cohen NJ. The hippocampus. What does it
do? Behav Neural Biol 1992;57:2–36.
[18] O’Keefe J, Nadel L. The hippocampus as a cognitive map. Oxford:
Clarendon Press; 1978.
[19] Bitterman ME. The comparative analysis of learning. Science 1975
(May 16);188:699–709.
[20] Papini MR. Role of reinforcement in spaced-trial operant learning in
pigeons (Columba livia). J Comp Psychol 1997;111:175–97.
[21] Papini MR, Muzio RN, Segura ET. Instrumental learning in toads
(Bufo arenarum): reinforcer magnitude and medial pallium. Brain
Behav Evol 1995;46:61–71.
[22] Miller RR. Effects of inter-trial reinstatement of training stimuli on
complex maze learning in rats: evidence that ‘‘acquisition’’ curves
reflect more than acquisition. J Exp Psychol, Anim Behav Processes
1982;8:86–109.
[23] Lopez JC, Bingman VP, Rodrıguez F, Gomez Y, Salas C. Dissociation
of place and cue learning by telencephalic ablation in goldfish. Behav
Neurosci 2000;117:687–99.
[24] Ohnishi K. Telencephalic function implicated in food-reinforced col-
our discrimination learning in goldfish. Physiol Behav 1989;46:
707–12.
[25] Salas C, Broglio CR, Rodrıguez F, Lopez JC, Portavella M, Torres B.
Telencephalic ablation in goldfish impairs performance in a ‘‘spatial
constancy’’ problem but not in a cued one. Behav Brain Res 1996;79:
193–200.
[26] Lowes G, Bitterman ME. Reward and learning in the goldfish.
Science 1967 (July 28);157:455–7.
[27] Crespi LP. Quantitative variation of incentive and performance in the
white rat. Am J Psychol 1942;55:467–517.
[28] Feldon J, Gray JA. The partial reinforcement extinction effect after
treatment with chlordiazepoxide. Psychopharmacology 1981;73:
269–75.
[29] McNaughton N. Effects of anxyolitic drugs on the partial reinforce-
ment effects in runway and Skinner box. Q J Exp Psychol 1984;36B:
319–30.
[30] Coe CL, Stanton ME, Levine S. Adrenal responses to reinforcement
and extinction: role of expectancy versus instrumental responding.
Behav Neurosci 1983;97:654–7.
[31] Dantzer R, Arnone M, Mormede P. Effects of frustration on behaviour
and plasma corticosteroid levels in pigs. Physiol Behav 1980;24:1–4.
[32] Osborne B, Silverhart T, Markgraf C, Seggie J. Effects of fornix
transection and pituitary–adrenal modulation on extinction behavior.
Behav Neurosci 1987;101:504–12.
[33] Daly HB. Reinforcing properties of escape from frustration aroused in
various learning situations. Psychol Learn Motiv 1974;8:187–231.
[34] Lopez JC, Broglio CR, Rodrıguez F, Thinus-Blanc C, Salas C. Re-
versal learning deficit in a spatial task but not in a cued one after
telencephalic ablation in goldfish. Behav Brain Res 2000;109:91–8.
[35] Rodrıguez F, Lopez JC, Vargas JP, Gomez Y, Broglio C, Salas C.
Conservation of spatial memory function in the pallial forebrain of
reptiles and ray-finned fishes. J Neurosci 2002;22(7):2894–903.
[36] Salas C, Rodrıguez F, Vargas JP, Duran E, Torres B. Spatial learning
and memory deficits after telencephalic oblation in goldfish trained in
place and turn maze procedures. Behav Neurosci 1996;110:965–80.
[37] Olton DS, Meck WH, Church RM. Separation of hippocampal and
amygdaloid involvement ion temporal memory dysfunction. Brain
Res 1987;404 [180B188].
[38] Portavella M. Implication of different forebrain areas in two-way
active avoidance conditioning in goldfish (Carassius auratus). Un-
published PhD dissertation. Sevilla: University of Sevilla; 2000.
[39] Portavella M, Vargas JP, Torres B, Salas C. The effects of telence-
phalic pallial lesions on spatial, temporal, and emotional learning in
goldfish. Brain Res Bull 2002;57(3/4):397–9.




















