
Evidence for a “Wattle and Daub” Model of the Cyst Wall of Entamoeba

Parasitol Res (1993) 79:153-159
Parasitnlngy Research �9 Springer-Verlag 1993
Expression of sequences related to c-myc in Entamoeba
Margarita Leyva Leyva 1, Patricio Gariglio 2, 3, Luz Mafia Rangel 3, Jes6s Vald~s 2, Patricia Ayala 2, Esther Orozeo 1
1 Department of Experimental Pathology, CINVESTAV-IPN, A.P. 14-740, M~xico, 07000 D.F. M6xico 2 Department of Genetics and Molecular Biology, CINVESTAV-IPN, A.P. 14-740, M6xieo, 07000 D.F. Mexico
CISEI, INSP, Av. Universidad No. 115, Cuernavaca, Mor. M~xico
Accepted: 25 November 1992
Abstract. The evolutionarily conserved proto-oncogene c-rnyc is involved in both proliferation and differentia- tion processes of higher eukaryotic cells. We report here the identification and characterization of sequences ho- mologous to c-myc in different Entamoeba species using a fragment of the mammalian c-myc gene as a probe. This probe hybridized with fragments of 3.5 and 3.4 ki- lobases (kb) in E. histolytica HindIII of EcoRI digested DNA. In E. invadens it recognized fragments of 3.1 and 2.8 kb, and in Laredo strain (reported as E. moshkovskii by Clark and Diamond in 1991) the probe hybridized with fragments of 17 kb. The c-myc probe identified transcripts of 3.3 and 1.5 kb in E. histolytica, transcripts of 1.8 and 1.3 kb in Laredo strain, and transcripts of 3.7, 1.8, 1.5 and 1.1 kb in E. invadens. Antibodies against a highly conserved region of the c-myc protein recog- nized in E. histolytica polypeptides of 35, 40, and 60 kDa. The expression of the 60 kDa polypeptide was temperature-inducible in Laredo strain. In E. invadens a 110 kDa strong band was detected by the antibodies.
Surprisingly, E. invadens myc-like sequences and pro- teins showed greater homology to mammalian c-myc gene and proteins. Expression of proteins antigenically related to c-mye varied according to the cell cycle phase of E. histotytica. These proteins peaked during D, G1, and S phases and declined during G2.
The Entamoeba genus comprises two pathogenic species: E. histolytica and E. invadens, the protozoan parasites responsible for human and reptilian amebiasis, respec- tively. E. histolytica probably infects more than 10% of the world's population and kills approximately 60000 people each year (Walsh 1986). Several reports have cor- related virulence of E. histolytica strains and biological properties of the trophozoites. These include: (1) ability to invade mammalian tissues; (2) agglutination by con-
Correspondence to: E. Orozco
canavalin A; (3) ability to form colonies in semisolid agar; and (4) negative surface charge (Trissl et al. 1977; Gillin and Diamond 1978). These properties resemble some characteristics of transformed cells (Topp et al. 1980).
On the other hand, proto-oncogene products exert a controlling influence on cellular proliferation and dif- ferentiation (Heldin and Westermark 1984; Weinberg 1985). The differential expression of proto-oncogenes during embryonic development (Muller et al. 1982), their homology to both growth factors (Doolittle et al. 1983; Waterfield etal. 1983) and growth factor receptors (Downward et al. 1984; Green and Chambon 1986; Sap et al. 1986; Weinberger et al. 1986), and their evolution- ary conservation (Bishop 1983; Varmus 1984) support this concept. The participation of proto-oncogenes in normal cellular metabolism has also been suggested (Denhardt et al. 1986). The identification of sequences homologous to vertebrate proto-oncogenes in inverte- brates (Shilo and Weinberg 1981) indicates that these genes present both a high degree of conservation and a wide distribution among divergent species.
The human c-myc gene is involved in growth regula- tion and in human cancer (Varmus 1984; Kelly and Sie- benlist 1986). Its expression might be controlled both at the transcriptional (Leder et al. 1983) and post-tran- scriptional levels (Gariglio et al. 1987a, b). The human c-rnyc protein is predicted to be 439 amino acids long, with a molecular weight of 49000 (Battey et al. 1983; Watson et al. 1983; Watt et al. 1983), but specific anti- sera precipitate proteins of 62 67 kDa (Hann and Eisen- man 1984; Ramsay et al. 1984; Hann et al. 1988). Molec- ular mechanisms of growth and invasiveness in Enta- moeba species could be related to the expression of prod- ucts related evolutionarily to mammalian oncogenes. The study of these molecules in lower eukaryotic cells such as Entamoeba will contribute to the understanding of cellular growth and differentiation phenomena in un- icellular parasites and their possible participation in vir- ulence. In this work we report the identification and partial characterization of sequences homologous to c-

154
myc and their corresponding proteins in three different Entamoeba species .
Materials and methods
Cultures
Trophozoites of E. histolytica, E. invadens and Laredo strain (E. moshkovskii) were grown and harvested as described previously (Diamond et al. 1978). E. histolytica was grown at 37 ~ C, E. inva- dens at room temperature, and Laredo strain at both 37 ~ C and room temperature. HeLa cells were grown in modified minimal essential medium supplemented with 10% fetal calf serum in a 10% CO2 humidified atmosphere.
Isolation and hybridization o f D N A and R N A
For the detection of sequences homologous to c-myc in Entamoeba, I kilobase (kb) of complementary DNA (cDNA) clone containing the second and third exons of the human c-myc gene (Marcu et al. 1983) was used as probe (Ryc-7.4). The probe (purified mye frag- ment without plasmid vector) was labeled with 32P-ATP by the random oligonucleotide primer reaction using a commercial kit (Pharmacia, USA) (Feinberg and Vogelstein 1983). For Southern hybridizations (Southern 1975) 10 gg amebic DNA, isolated as de- scribed (Hernfindez et al. 1986), was HindIII or EcoRI digested and electrophoresed on 0.8% agarose gels for 16 h at 20 V, trans- ferred to Nytran membranes (Schleicher & Schuell, USA), hybrid- ized for 20 h with the radiolabeled probe and washed several times at 42 ~ C in 5 x standard saline citrate (SSC), 1% sodium dodecyl sulfate (SDS). For Northern experiments, 10 gg total amebic RNA, isolated as described previously (Leyva-Leyva et al. 1986), was elec- trophoresed through a 1.2% agarose gel containing 1% formalde- hyde (Merck, USA) in 3-(N-morpholino) propanesulfonic acid (MOPS) (Sigma, St. Louis, Mo., USA) buffer (Ausubel et al. 1987). RNA was transferred to Nytran membranes, hybridized for 20 h with the radiolabeled probe and washed as described for DNA. HeLa D N A was used in all experiments as a control. Kodak films with DuPont Cronex Lightning-Plus intensifying screens were ex- posed to the membranes for different times at - 7 0 ~ C.
Cell cycle
For synchronization experiments, 5 • 102 trophozoites ml 1 in amebic medium were incubated for 25 h at 37 ~ C in 10 ml of TYI-S- 33 medium containing 200 txg ml- i colchicine. Afterwards, tropho- zoites were washed and incubated in 10 ml fresh medium (Orozco et al. 1988). Aliquots were taken at different times after colchicine washing: I h for division phase (D), 5 h for gap-1 phase (G1), 9 h for synthesis phase (S) and 12 h for gap-2 phase (G2) (Orozco et aL 1988). Synchronization of the cultures was monitored by counting the cell numbers at different times.
Immunofluor escence
Formaldehyde-fixed trophozoites in different cell cycle phases were permeabilized by incubation for 5 rain in acetone at - 20 ~ C. Then, they were incubated for 1 h at 37 ~ C with the mAb anti-c-myc, washed three times with PBS, and incubated for 1 h at 37 ~ C with a fluoresceinated second antibody. Trophozoites were washed again and fuorescence was monitored by microscopy (Polybar).
Results
Entamoeba genome has transcriptionally active c-myc-l ike D N A sequences
T h e f i rs t s tep in this w o r k was to d e t e r m i n e w h e t h e r h o m o l o g o u s s equences to the m a m m a l i a n c-myc gene were p r e s e n t in Entamoeba D N A . HindII I or EcoRI di-
i .
Hind31 Eco RI
Q; �9 O "~ Q; - "I- W _J uJ I W --J W
Protein extraetion, gel electrophoresis and immunoblott ing
Trophozoites were washed with phosphate buffered saline (PBS) and lysed in the presence of 100 mM p-hydroxy-mercuribenzoate (PHMB) (Sigma) in 100 mM TRIS-HC1 pH 8.0. The mixture was vortexed at 4 ~ C, diluted with Laemmli buffer (Laemmli 1970) and boiled for 5 rain in a water bath. Protein concentration was deter- mined by the Lowry technique (Lowry et al. 1951), and aliquots containing t00 gg protein were subjected to 10% SDS-polyacryl- amide gel electrophoresis (PAGE). Proteins were transferred to nitrocellulose (NC) (Towbin et al. 1979) and filters were incubated for 3 h at room temperature with the monoclona[ antibodies (mAb) against a highly conserved region of the c-myc protein (amino acids 43 to 55; Microbiological Associates, Bethesda Laboratories, Md., USA). Binding of specific antibodies (IgGs) was detected by incubation of NC filters with 12SI-labeled protein G (1 x ] 0 6 epm) for 2 h at room temperature. NC filters were washed and dried, and Kodak X-Omat XAR-ray films with an intensifying screen were exposed to the filters. To determine whether proteolytic degradation due to E. histolytica proteases was a factor in these experiments, trophozoites or protein extracts from E. invadens were mixed with an increasing number of trophozoites, or amount of proteins, from E. histolytica. Cell mixtures were incubated at 4 ~ C for 15 min, and proteins were coextracted and analyzed as men- tioned above.
3 3 |
Fig. 1. Southern blot analysis of c-myc gene in Entamoeba. DNA from E. histolytica (E.h.), Laredo strain (Lar), E. invadens (E.i.) and HeLa cells was Hind III or EcoRI digested, separated in 0.8% agarose gel, transferred to Nytran membranes and hybridized with the Ryc-7.4 probe as described in Materials and methods. Kodak films were exposed 10 days to the membranes. HeLa lanes were exposed for only 2 days
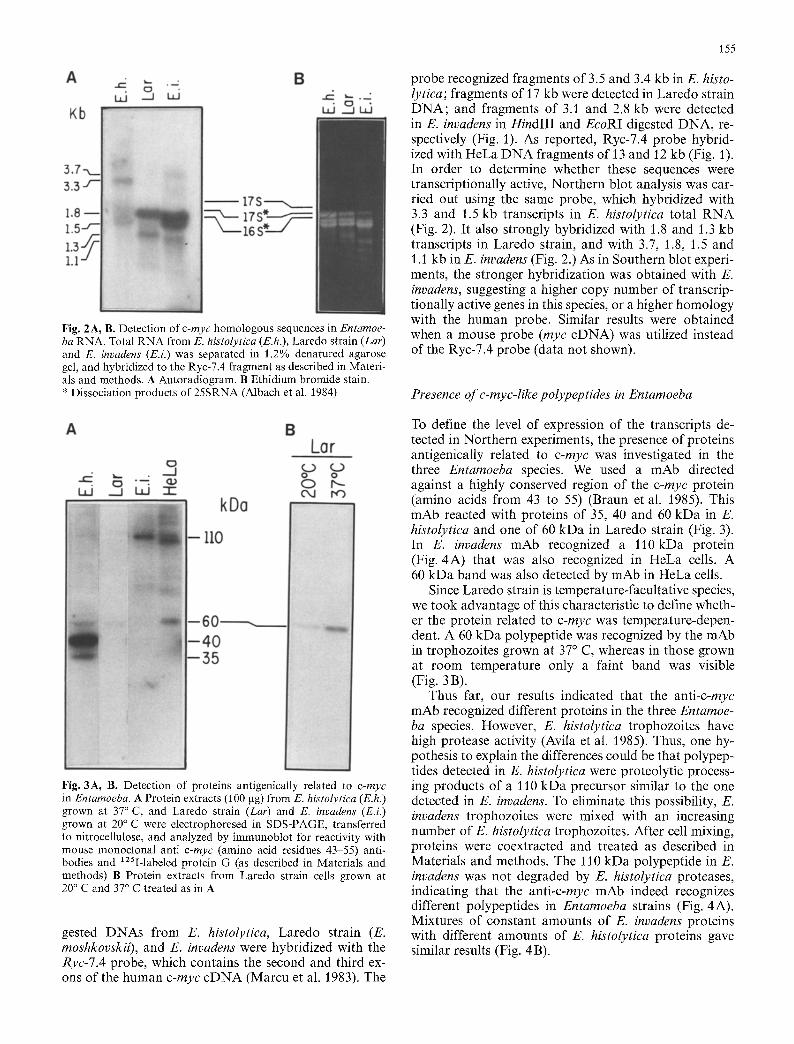
155
A .= ~ . - B ,,; w _d,-. �9
Fig. 2 A, B. Detection of c-myc homologous sequences in Entamoe- ba RNA. Total RNA from E. histolytica (E.h.), Laredo strain (Lar) and E. invadens (E.i.) was separated in 1.2% denatured agarose gel, and hybridized to the Ryc-7.4 fragment as described in Materi- als and methods. A Autoradiogram. B Ethidium bromide stain. * Dissociation products of 25SRNA (Albach et al. 1984)
probe recognized fragments of 3.5 and 3.4 kb in E. histo- lytica; fragments of 17 kb were detected in Laredo strain DNA; and fragments of 3.1 and 2.8 kb were detected in E. invadens in HindIII and EcoRI digested DNA, re- spectively (Fig. 1). As reported, Ryc-7.4 probe hybrid- ized with HeLa DNA fragments of 13 and 12 kb (Fig. 1). In order to determine whether these sequences were transcriptionally active, Northern blot analysis was car- ried out using the same probe, which hybridized with 3.3 and 1.5 kb transcripts in E. histolytica total RNA (Fig. 2). It also strongly hybridized with 1.8 and 1.3 kb transcripts in Laredo strain, and with 3.7, 1.8, 1.5 and 1.1 kb in E. invadens (Fig. 2.) As in Southern blot experi- ments, the stronger hybridization was obtained with E. invadens, suggesting a higher copy number of transcrip- tionally active genes in this species, or a higher homology with the human probe. Similar results were obtained when a mouse probe (myc cDNA) was utilized instead of the Ryc-7.4 probe (data not shown).
Presence of c-myc-like polypeptides in Entamoeba
A B Lar
o o o "-~ ~ 0
uJ _A ~ -r od
Fig. 3A, B. Detection of proteins antigenically related to c-myc in Entamoeba. A Protein extracts (100 btg) from E. histolytica (E.h.) grown at 37 ~ C, and Laredo strain (Lar) and E. invadens (E.i.) grown at 20~ were electrophoresed in SDS-PAGE, transferred to nitrocellulose, and analyzed by immunoblot for reactivity with mouse monoclonal anti c-myc (amino acid residues 43-55) anti- bodies and 125I-labeled protein G (as described in Materials and methods) B Protein extracts from Laredo strain cells grown at 20 ~ C and 37 ~ C treated as in A
gested DNAs from E. h&tolytica, Laredo strain (E. moshkovskii), and E. invadens were hybridized with the Ryc-7.4 probe, which contains the second and third ex- ons of the human c-myc cDNA (Marcu et al. 1983). The
To define the level of expression of the transcripts de- tected in Northern experiments, the presence of proteins antigenically related to c-myc was investigated in the three Entamoeba species. We used a mAb directed against a highly conserved region of the c-myc protein (amino acids from 43 to 55) (Braun et al. 1985). This mAb reacted with proteins of 35, 40 and 60 kDa in E. histolytica and one of 60 kDa in Laredo strain (Fig. 3). In E. invadens mAb recognized a 110kDa protein (Fig. 4A) that was also recognized in HeLa cells. A 60 kDa band was also detected by mAb in HeLa cells.
Since Laredo strain is temperature-facultative species, we took advantage of this characteristic to define wheth- er the protein related to c-myc was temperature-depen- dent. A 60 kDa polypeptide was recognized by the mAb in trophozoites grown at 37 ~ C, whereas in those grown at room temperature only a faint band was visible (Fig. 3 B).
Thus far, our results indicated that the anti-c-myc mAb recognized different proteins in the three Entamoe- ba species. However, E. histolytica trophozoites have high protease activity (Avila et al. 1985). Thus, one hy- pothesis to explain the differences could be that polypep- tides detected in E. histolytica were proteolytic process- ing products of a 110 kDa precursor similar to the one detected in E. invadens. To eliminate this possibility, E. invadens trophozoites were mixed with an increasing number of E. histolytica trophozoites. After cell mixing, proteins were coextracted and treated as described in Materials and methods. The 110 kDa polypeptide in E. invadens was not degraded by E. histolytica proteases, indicating that the anti-c-myc mAb indeed recognizes different polypeptides in Entamoeba strains (Fig. 4A). Mixtures of constant amounts of E. invadens proteins with different amounts of E. histolytica proteins gave similar results (Fig. 4 B).

156
A l 2 3 4 5 6 7
B 1 2 5 4
1 n
)
Fig. 4A, B. Interaction between protein extracts obtained from E. invadens and E. histolytica trophozoites. A Immunoblot using the mAb anti c-myc (amino acid residues 43-55) of Entamoeba proteins was performed with cell mixtures containing different numbers of E. invadens: E. histolytica cells: lane 1, 1 x 106:0; lane 2, 1 x 106 : 1 X 103 ; lane 3, I x 106 : 1 x 104; lane 4, 9 x 10 s : 1 x 105 ; lane
Expression of c-myc-like proteins through the cell cycle of E. histolytiea
We determined the cellular distribution and accumula- tion through the cell cycle of proteins related to c-rnyc. Trophozoites were synchronized by colchicine and c- rnyc-like peptides were localized by immunofluorescence through the cell cycle phases of E. histolytica. An intense signal was detected in a high percentage of cells in S and G1 phases. Signal was also detected in D phase, but only a few fluorescent cells were seen during G2 phase (Fig. 5). Since no more than 90% of the tropho- zoites were synchronous, these positive cells may not actually be in G2 phase. This is supported by the fact that some of the fluorescent cells in G2 showed two nuclei, indicating that trophozoites were dividing (specif- ically, in late telophase) (Orozco et al. 1988).
Discussion
We report here the identification and partial character- ization of genes and proteins related to c-myc in E. histo- lytica and E. invadens, two pathogenic species, and in Laredo, a non-pathogenic strain corresponding to the E. moshkovskii species (Clark and Diamond 1991). In Trypanosoma brucei the presence of three DNA frag-
5, 5 x 105:5x 105; lane 6, 0:1 x 106, lane 7, HeLa cells as positive control (1 x 106). B Same as in A, except that a different ratio of proteins (E. invadens :E. histolytiea) was used: lane 1, HeLa cells as positive control (30gg); lane 2, 100gg:0gg; lane 3, 50 ~tg: 50 gg; lane 4, 0:100 gg. The immunoblots were carried out as described in Fig. 3
ments hybridizing with a v-myc probe has been reported (Davis et al. 1989).
The Ryc-7.4 probe, a cDNA fragment containing only human c-myc sequences (without plasmid vector sequences), hybridized with DNA from the three Enta- moeba species studied here. The stronger hybridization was obtained with E. invadens, suggesting a higher gene copy number or a higher homology with the human c-myc. Northern experiments also showed a stronger hy- bridization with the transcripts in E. invadens, suggesting again a higher homology of the transcripts with the Ryc- 7.4 probe, or a higher amount of these RNA species. The molecular weight of Entamoeba transcripts was vari- able in the three species, although similar patterns were evident in Laredo strain and E. invadens. Transcripts of 1.5 and 3.3 kb were found in E. histolytica, the latter similar in length to the single transcript reported for T. brucei (Davis et al. 1989). We used low stringency conditions since, except for T. brucei, the c-myc gene has been identified in lower eukaryotes where it has been looked for extensively. The data in this paper, while suggestive, do not prove the existence of myc-like se- quences in Entamoeba and we are currently trying to clone and sequence these genes.
Northern and Western analysis suggested that c-myc- like sequences are both transcriptionally and translation- ally active in Entamoeba. The latter is indicated by the

Fig. 5. c-myc like protein expression during the cell cycle of E. histolytica. Trophozoites were synchronized by incubation for 25 h in 200 gg ml 1 colchicine. After washing, trophozoites were incu- bated at 37~ in fresh amebic medium and aliquots were taken at t h (M), 5 h (G1), 9 h (S) and 12 h (G2). Trophozoites were permeabilized by 5 min incubation in acetone, and immunofluores- cence was carried out using the monoclonal antibody against c-mye and a fluoresceinated second antibody. After the cultures had been synchronized the trophozoites were counted every 4 h. Permeabili- zation of the plasma and nuclear membranes was monitored using an anti-RNA polymerase I1 antibody (kindly provided by Dr. M. Dahrnus, Davis, Calif., USA) (c~POL). HeLa cells (HeLa) were also analyzed in parallel, n, Nucleus. Right-hand panels cell stage are photomicrographs
recognition of amebic proteins by antibodies generated against the highly conserved 43 55 amino acid region of the c-myc mammalian protein (Braun et al. ]986).
Proteins antigenically related to c-myc could be members of the myc family or could be nuclear proteins
157
presenting the highly conserved domain to which the antibodies are directed. Bands obtained with the mAb differed in intensity in the three Entamoeba species, but HeLa cells and E. invadens gave similar patterns. The 1 ] 0 kDa band was very strong in HeLa and in E. inva- dens, but it was very faint in E. histolytica and in Laredo strain. The 60 kDa band appeared in HeLa cells and in E. histolytica, but it was weak in E. invadens and in Laredo. This protein was temperature-induced in Lar- edo strain. In E. histolytica a strong 40 kDa band was present, but it was not detected in the other species.
Experiments using both protein and cell mixtures of E. histolytica and E. invadens excluding the possibility that the lower molecular weight proteins were degrading products due to the E. histolytica proteases. Thus, differ- ential gene expression of proteins related to c-myc in the tree Entamoeba species could account for these re- sults. This assumption is supported by the different tran- scripts detected in the three species, suggesting either that we are dealing with different genes or different con- trol mechanisms at both transcriptional and translation- al levels.
Transcripts of 3.3 and 1.5 kb were detected in E. histolytica that may encode for proteins of about 110 and 50 kDa, respectively. In Laredo strain the probe detected 1.8 and J.3 kb transcripts that may encode for proteins of 60 and 43 kDa, respectively. In E. invadens, four transcripts of 3.7, 1.8, ] .5, and 1.1 kb were detected; the predicted weights for proteins would be 120, 60, 50 and 37 kDa, respectively. A reasonable correlation was found between the molecular weights of the transcripts recognized by Ryc-7.4 and the proteins detected by anti- bodies. These results suggest the presence of evolutionar- ily conserved c-myc-like sequences. Several clones and strains o fE . histolytica have given similar results to those reported in this paper (data not shown). In T. brucei proteins of 40, 50 and 63 kDa were detected by polyclon- al antibodies against c-myc (Davis et al. 1989). A high molecular weight protein was surprisingly found in E. invadens that probably shared the same epitope with a 110 kDa myc-like protein found in lymphoblast, activat- ed lymphocytes and HeLa cells (Miranda et al. 1991).
On the other hand, previous reports have shown Lar- edo to differ from E. histolytica in zymodeme (Sargeaunt etal . 1980), nuclear division (Gicquaud 1979; Orozco et al. 1988), and optimal growth temperature (Martinez- Palomo 1982). Recently Clark and Diamond (1991) re- ported that Laredo belongs to E. moshkovskii species. Results obtained here confirm that Laredo belongs to a different species than E. histolytica.
In mammalian cells, expression of c-myc occurs at a constant level throughout the cell cycle (Hann et al. 1985; Thompson et al. 1985), although a transient in- crease in expression is observed upon treatment with mitogens (Kelly et al. 1983). We found expression of pro- teins antigenically related to c-myc in the nucleus of tro- phozoites of E. histotytica at a constant level through the cell cycle, except perhaps in G2. Subcellular localiza- tion is further evidence that Entamoeba has proteins re- lated to c-myc, and probably they carry out similar func- tions to those reported for mammalian cells. However,

158
fur ther s tudies are necessary to d e m o n s t r a t e the def ini te funct ions o f p ro te ins re la ted to c - m y c in paras i tes . Func- t iona l mo lecu l a r s tudies in paras i tes , on the role o f the c - m y c produc t s , w o u l d e luc ida te whe the r or no t they are impl ica ted in D N A rep l ica t ion (Studzinski et al. 1986; G u l l & f e z e t a l . 1987; Iguch i -Ar iga et al. 1987), t r ansc r ip t ion (Kings ton et al. 1984; K a d d u r a h - D a o u k e t a l . 1987) and R N A process ing (Spec tor et al. 1987), as has been found in m a m m a l i a n s .
Acknowledgements. The research of Dr. Orozco was supported in part by an International Research Scholars award from the How- ard Hughes Medical Institute. This work was also partially sup- ported by CoNaCyT (M6xico), The MacArthur Foundation (USA) and Givaudan de Maxico S. A. The authors gratefully acknowledge Drs. Adolfo Martlnez-Palomo, Debra Peattie and Pedro Huertas for critical reading of this manuscript. We also thank Biol. Pedro Chfivez for his help with the illustrations and Alicia Romero for secretarial assistance.
References
Alhach RA, Prachayasittikul V, Heebner GM (1984) Isolation and characterization of RNA of Entamoeba histolytica. Mol Bio- chem Parasitol 12: 261-272
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K (eds) (1987) Current protocols in molecular biology, vol 1. Greene Publishing Associates and Wiley-lnlersei- once, Chapter 4
Avila EE, S/mchez-Garza M, Calder6n J 0985) Entamoeba histoly- tica and E. invadens: sulfhydryl-dependent proteolytic activity. J Protozool 32:163-166
Battey J, Moulding C, Taub R, Murphy W, Stewart T, Potter H, Lenoir G, Leder P (1983) The human c-myc oncogene : struc- tural consequences of translocation into the IgH locus in Bur- kitt lymphoma. Cell 34:779 787
Bishop JM (1983) Cellular oncogenes and retroviruses. Annu Roy Biochem 52: 301-354
Braun M J, Deininger PL, Casey JW (1985) Nucleotide sequence of a transduced myc gene from a defective feline leukemia pro- virus. J Virol 55:177-183
Clark CG, Diamond LS (1991) The Laredo strain and other 'Enta- moeba histolytica-like' amoebae are Entamoeba moshkovskii. Mol Biochem Parasitol 46:11-18
Davis CE, Colmerauer ME, Kim CH, Matthews B, Guiney DG (1989) myc-related proteins and DNA sequences in Trypanoso- ma brucei. Microb Pathog 7:45 53
Denhardt DT, Edwards DR, Parfett CL (1986) Gene expression during the mammalian cell cycle. Biochim Biophys Acta 865:83 125
Diamond LS~ Harlow DR, Cunnichk CC (1978) A new medium for the axenic cultivation of Entamoeba histolytica and other Entamoeba. Trans R Soc Trop Med Hyg 72:431-432
Doolittle RF, Hunkapiller MW, Hood LE, Devare SG, Robbins KC, Aaronson SA, Antoniades HN (1983) Simian sarcoma vi- rus oncogene, v-sis, is derived from the gene (or genes) encoding a platelet-derived growth factor, Science 221:275-277
Downward J, Yarden Y, Mayes E, Scrace G, Totty N, Stockwell P, Ullrich A, Schlessinger J, Waterfield MD (1984) Close simi- larity of epidermal growth factor receptor and v-erb-B onco- gene protein sequences. Nature 307 : 521-527
Feinberg AP, Vogelstein B (1983) A technique for radiolabeling DNA restriction endonuclease fragments to high specific activi- ty. Anal Biochem 137: 266~267
Gariglio P, OcAdiz R, Sauceda R (1987a) Human papillomavirus DNA sequences and c-myc oncogene alterations in uterine cer- vix carcinoma. Cancer cells, vol 5. Papillomavirus. Spring Har- bor Laboratory, New York, pp 343-348
Gariglio P, delAngel R, Herrera A, Bonilla M (1987b) Theoretical model for the post-transcriptional regulation of the human c- myc gene expression, involving double-stranded RNA process- ing. J Theor Biol 125:83-92
Gicquaud CR (1979) Etude de l'ultrastructure du noyau et de la mitose de Entamoeba histolytica. Bio[ Cell 35:305 312
Gillin FD, Diamond LS (1978) Clonal growth of Entamoeba histo- lytica and other species of Entamoeba in agar. J Protozool 25:539-543
Green S, Chambon P (1986) A superfamily of potentially oncogenic hormone receptors. Nature 324:615-617
Gutiarrez C, Guo ZS, Farrell-Towt J, Ju G, DePamphilis ML (i 987) c-myc protein and DNA replication: separation of c-myc antibodies from an inhibitor of DNA synthesis. Mol Cell Biol 7:4549~4589
Harm SR, Eisemnan RN (1984) Proteins encoded by the human c-myc oncogene: differential expression in neoplastic cells. Mol Cell Biol 4: 2486-2497
Hann ST, Thompson CB, Eisenman RN (1985) c-myc oncogene protein synthesis is independent of the cell cycle in human and avian cells. Nature 314:366-369
Harm SR, King MW, Bentley DL, Anderson CW, Eisenman RN (1988) A non-AUG translational initiation in c-myc exon 1 generates an N-terminally distinct protein whose synthesis is disrupted in Burkitt's lymphomas. Cell 52:185-195
Heldin CH, Westermark B (1984) Growth factors: mechanism of action and relation to oncogenes. Cell 37:9-20
Hern~ndez FC, Oc~tdiz JR, Orozco E (1986) AnAlisis de la organi- zaci6n del DNA de varias cepas y clonas de Enamoeba histolyti- ca. Arch Invest M6d 17 [Suppl 1 ] : 107-112
Iguchi-Ariga SM, ltani T, Kiji Y, Ariga H (1987) Possible function of the c-myc product : promotion of cellular DNA replication. EMBO J 6:2365-2371
Kaddurah-Daouk R, Green M J, Baldwin AS, Kingston RE (1987) Activation and repression of mammalian gene expression by the c-myc protein. Genes Dev 1:347-357
Kelly K, Siebenlist U (1986) The regulation and expression of c- myc in normal and malignant cells. Annu Rev Immunol 4:317- 338
Kelly K, Cochran BH, Stiles CD, Leder P (1983) Cell-specific regu- lation of the c-myc gene by lymphocyte mitogens and platelet- derived growth factor. Cell 35:603-610
Kingston RE, Baldwin AS, Sharp PA (1984) Regulation of heat shock protein 70 gene expression by c-myc. Nature 312:280-282
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685
Leder P, Battey J, Lenoir G, Moulding C, Murphy W, Potter H, Stewart T, Taub R (1983) Translocations among antibody genes in human concer. Science 222:765-771
Leyva-Leyva M, Hidalgo ME, Orozco E (1986) Aislamiento, carac- terizaci6n y traducci6n in vitro de RNA de donas de Entamoe- ba histolytica deficientes en adhesi6n. Arch Invest Mad 17 [Suppl. 1]: 155-159
Lowry OH, Rosebrough N J, Farr AL, Randall RJ (1951) Protein measurement with the Folin-phenol reagent. J Blot Chem 193:265-275
Marcu KB, Harris LJ, Stanton LW, Erickson J, Watt R, Croce CM (1983) Transcriptionally active c-myc oncogene is con- tained within NIARD, a DNA sequence associated with chro- mosome translocations in B-cell neoplasia. Proc Natl Acad Sci USA 80:519-523
Martinez-Palomo A (1982) The biology of Entamoeba histolytica. Research Studies Press, Wiley, Chichester
Miranda E, Valles Y, Hernfindez L, Rangel L, Castrej6n A, Collaza J, Gutiarrez M, Gonzfilez R, Gariglio P (1991) Estudio de la protelna myc y de proteinas antig~nicamente relacionadas a myc en leucemia linfoblfistica aguda. Rev Invest Clin 43:139- 145
Muller R, Slamon DJ, Tremblay JM, Cline MI, Verma IM (1982) Differential expression of cellular oncogenes during pre- and postnatal development of the mouse, Nature 299 : 640-644

159
Orozco E, Solis FJ, Dominguez J, ChS, vez B, Hernfindez F (1988) Entamoeba histolytica: Cell cycle and nuclear division. Exp Par- asitol 67: 85-95
Ramsay G, Evan G, Bishop JM (1984) The protein encoded by the human proto-oncogene c-myc. Proc Natl Acad Sci USA 81 : 7742-7746
Sap J, Mufioz A, Damm K, Goldberg Y, Gysdael J, Leutz A, Beug H, Vennstrom B (1986) The c-erb-A protein is a high- affinity receptor for thyroid hormone. Nature 324:635-640
Sargeaunt PG, Williams JE, Neal RA (1980) A comparative study of Entamoeba histolytica (NIH : 200, HKg, etc.), "E. histolytica- like" and other morphologically identical amoebae using isoen- zyme electrophoresis. Trans R Soc Trop Med Hyg 74:469-474
Shilo BZ, Weinberg RA (1981) DNA sequences homologous to vertebrate oncogenes are conserved in Drosophila melanogaster. Proc Nat Acad Sci USA 78 : 6789 6792
Southern EM (1975) Detection of specific sequences among DNA fragments separated by gel electrophoresis. J Mol Biol 98 : 503 517
Spector D, Watt R, Sullivan N (1987) The v-myc and c-myc pro- teins colocalize in situ with small nuclear ribonucleoprotein par- ticles. Oncogene 1 : 5-12
Studzinski GP, Brelvi ZS, Feldman SC, Watt RA (1986) Participa- tion of c-myc protein in DNA synthesis of human cells. Science 234: 467-470
Thompson CB, Challoner PB, Neiman PE, Groudine M (1985) Levels of c-rnyc oncogene mRNA are invariant throughout the cell cycle. Nature 314:363 366
Topp WC, Lane D, Pollack R (1980) Transformation by SV40 and polyoma virus. In: Tooze J (ed) Molecular biology of tumor viruses, 2nd edn, vol 4, part 2. DNA tumor viruses. Cold Spring Harbor Laboratory, New York, pp 205-296
Towbin H, Staehelin T, Gordon J (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Nat Acad Sci USA 76:4350-4354
TrissI D, Martinez-Palomo A, Arg~ieIio C, De la Torre M, de Ia Hoz R (1977) Surface properties related to concanavalin A- induced agglutination. A comparative study of several Enta- moeba strains. J Exp Med 145:65~655
Varmus HE (1984) The molecular genetics of cellular oncogenes. Anna Rev Genet 18 : 553-612
Walsh JA (1986) Problems in recognition and diagnosis of amebia- sis : estimation of the global magnitude of morbidity and mor- tality. Rev Infect Dis 8:228-238
Waterfield MD, Scrace T, Whittle N, Stroobant P, Johnsson A, Wasteson A, Westermark B, Heldin CH, Huang JS, Deuel TF (1983) Platetet derived growth factor is structurally related to the putative transforming protein p28-sis of simian sarcoma virus. Nature 304:35-39
Watson DK, Psallidopoulos MC, Samuel KP, Dalla-favera R, Pa- pas TS (1983) Nucleotide sequence analysis of human c-myc locus, chicken homologue, and myelocytomatosis virus MC29 transforming gene reveals a highly conserved gene product. Proc Natl Acad Sci USA 80:3642-3645
Watt R, Stanton LW, Marcu KB, Gallo RC, Croce CM, Rovera G (1983) Nucleotide sequence of cloned cDNA of human c-myc oncogene. Nature 303 : 725-728
Weinberg RA (1985) The action of oncogenes in the cytoplasm and nucleus. Science 230 : 770-776
Weinberger C, Thompson CC, Ong ES, Lebo R, Gruol D J, Evans RM (1986) The c-erb-A gene encodes a thyroid hormone recep- tor. Nature 324:641-646
Copyright © 2022 FDOKUMEN




















