
The Jacob2 Lectin of the Entamoeba histolytica Cyst Wall Binds Chitin and Is Polymorphic
Upload
independentCategory
view
2download
0
ORIGINAL PAPER
E. Orozco · R. Gharaibeh · A. M. RiveronD. M. Delgadillo · M. Mercado · T. SanchezE. Gomez Conde · M. A. Vargas · R. Lopez-Revilla
A novel cytoplasmic structure containing DNA networksin Entamoeba histolytica trophozoites
Received: 7 August 1996 / Accepted: 7 October 1996
Abstract We report here the presence of cytoplasmicDNA arranged in networks in the trophozoites of thehuman parasite Entamoeba histolytica. CytoplasmicDNA was detected in live trophozoites in a structurethat we called EhkO, using the fluorescent dye acridineorange, and by in situ hybridization to trophozoites witha rDNA probe. The EhkO was found in the axenicallygrown clones A, L6 (strain HM1:IMSS) and MAVax(strain MAV) and in the polyxenically grown cloneMAVpx (strain MAV). Bacteria present in MAVpx didnot cross hybridize with the DNA probe neither in insitu hybridization or in Southern blot experiments.Autoradiography of metabolically [3H]thymidine-la-beled trophozoites showed the presence of EhkO, and anEhkO-enriched fraction, purified from a nuclei-free ex-tract and examined by light microscopy, exhibited[3H]thymidine incorporation into this structure. DNAwas purified from the EhkO and enriched nuclear frac-tions and analyzed by transmission electron microscopy.The EhkO fraction contained DNA networks resem-bling those of trypanosome kDNA, whereas nuclearDNA was present mainly as linear molecules and some
circles. Our findings imply that E. histolytica may betaxonomically more closely related to the Trypanoso-matidae than previously suspected.
Key words DNA networks · Entamoebahistolytica · DNA organization
Introduction
Entamoeba histolytica, the microaerophilic protozoanspecies responsible for human amebiasis, belongs to thephylum Sarcomastigophora, like the aerobic Try-panosomatidae. A mitochondrion with kinetoplastDNA (kDNA) formed by maxicircles and minicirclescatenated into a network characterizes Trypanosomat-idae (Simpson and Silva 1971; Borst 1991; Cozzarelli1993; Ferguson et al. 1994) and places them in the OrderKinetoplastidae, formed by organisms which are knownto contain kDNA networks in their mitochondria (Borst1991). Some 25 years ago Albach et al. (1969) suggestedthe presence of cytoplasmic DNA in E. histolytica.However, Albach et al. (1980) later on explained thesefindings as being due to bacterial contamination.Therefore, it is currently accepted that E. histolyticalacks extranuclear DNA and mitochondria (MartınezPalomo 1986; Clark and Roger 1995). Phylogeneticstudies of the chaperonin 60 protein (cpn60) place E.histolytica between Trypanosoma cruzi and Rickettsiatsutsugamushi, suggesting that it may form the firstbranch of amitochondrial organisms (Clark and Roger1995). In contrast, studies of the rDNA small subunitsequence locate E. histolytica on a phylogenetic branchthat emerged after the appearance of organisms withtypical eukaryotic organelles (Cavalier-Smith 1993;Hasegawa et al. 1993). Ubiquinone (CoQ) has beenidentified (Ellis et al. 1994) and several housekeepingmitochondrial genes have been cloned from this parasite(Clark and Roger 1995; Ellis et al. 1994); their presencehas been explained as deriving from the nuclear remnantof a mitochondrial primordium, which was secondarily
Mol Gen Genet (1997) 254:250 – 257 Springer-Verlag 1997
Communicated by W. Goebel
E. Orozco · D. M. Delgadillo · E. Gomez CondeM. A. Vargas · R. Lopez-RevillaProgram of Molecular Biomedicine,CINVESTAV-IPN, A.P. 14-470,Mexico 07300 D.F.
R. Lopez-RevillaDepartment of Cell Biology,CINVESTAV-IPN A.P. 14-740,Mexico 07300 D.F.
A. M. RiveronNational Center for Scientific Research (CNIC)P.O. Box 6990, Cubanacan, Playa,Havana, Cuba
E. Orozco (&) · R. Gharaibeh · M. Mercado · T. SanchezDepartment of Experimental Pathology, CINVESTAV-IPN,A.P. 14-470, Mexico 07300 D.F.

lost (Clark and Roger 1995). However, the presence ofcytoplasmic DNA as a remnant of this mitochondrialprimordium has not been totally excluded. We havecarried out a number of experiments to elucidate someaspects of the unique DNA organization in this parasite.Our recent results demonstrate the presence of extra-nuclear DNA, and rosette-like structures were observedin total DNA (Baez-Camargo et al., in press). In thispaper we report the identification, isolation and initialcharacterization of a novel E. histolytica cytoplasmicstructure, designated the EhkO, that contains DNAnetworks (EhkDNA), similar in some respects to thekDNA of Trypanosomatidae (Simpson and Silva 1971;Borst 1991; Cozzarelli 1991; Ferguson et al. 1994;Robinson and Gull 1994; Perez-Morga and England1993). These findings place E. histolytica probably muchcloser to the Trypanosomatidae than expected; addi-tionally, they open the possibility that the EhkO corre-sponds to an organelle, the functions of which arecurrently under study.
Materials and methods
Culture of E. histolytica trophozoites
Trophozoites of clones A, L6 (strain HM1:IMSS) (Orozco et al.1983) and MAVax (Vargas and Orozco 1993) were axenicallycultured in TY1-S-33 medium as described before (Diamond et al.1978), while clone MAVpx was cultured in Robinson medium inthe presence of its original intestinal flora (Vargas and Orozco1993). All trophozoites were harvested in logarithmic phase fromrich culture media.
Acridine orange staining of liveand paraformaldehyde-fixed trophozoites
Trophozoites in logarithmic phase were chilled on ice, centrifugedfor 5 min at 1,200 rpm and resuspended in PBS at 106 trophozoitesper ml. Aliquots of 100 ll were spotted on coverslips that were placedin 50 ml conical tubes with TYS-1–33 medium, and horizontallyincubated for 30 min at 37° C. Then, the coverslips were taken outwith forceps, rinsed twice with phosphate-buffered saline PBS pH 6,prewarmed at 37° C, and incubated for 10 min at 37° C in a humidchamber with 0.1% acridine orange prepared in PBS pH 6.
Trophozoites were washed twice with PBS at 37° C and thecoverslips were placed on slides with a drop of PBS prewarmed at37° C and immediately observed with a fluorescence microscope(Olympus). In parallel experiments, trophozoites were fixed on theslides with 4% paraformaldehyde before incubation with the ac-ridine orange dye. As a control, the trophozoites on the coverslipswere treated with 50 U DNase for 1–6 h at 37° C before adding theacridine orange.
In situ hybridization of E. histolytica trophozoiteswith the ribosomal DNA EhVR1 probe
Trophozoites of the axenically growing clones A, L6 and MAVax,and of the clone MAVpx growing with intestinal flora, were fixedwith 4% paraformaldehyde, placed on slides at 37° C for 1 h andincubated for 3 h with 50 U RNase/ml at 37° C. A 500 bp DNAprobe derived from the EhVR1 fragment (Orozco et al. 1993) of the5′ end of the 16S rDNA gene was labeled by random priming(Ausubel et al. 1993) using [35S]dATP. For control experiments, thesingle-stranded sense DNA probe was transcribed in vitro in the
presence of [35S]dUTP (Ausubel et al. 1993) from the Bluescriptphagemid in which it was cloned. Hybridization was carried out inthe presence of 50% formamide at 42° C and washing steps weredone at 65° C in 0.1 × SSC (1 × SSC is 0.15 M NaCl, 15 mM sodiumcitrate, pH 7.0) (Ausubel et al. 1993). Slides were covered with anemulsion film (Kodak) and developed after 5–7 days of exposure inthe dark, stained with Giemsa and observed with the immersionobjective of an optical microscope at 100 × (Olympus). As a controlfor the absence of bacteria in the axenic cultures we plated one ml ofthe trophozoite cultures in Luria and Terrific broth media (Ausubelet al. 1993). We also separated high molecular weight DNA bytransverse alternating field electro-phoresis (TAFE) (Baez-Camargoet al., in press) using 3 × 106 trophozoite of clones A, MAVax,MAVmx (grown in TY1-S-33 medium with Fusobacterium simbio-sum) and MAVpx, and approximately 108 bacteria from the intes-tinal flora that was used to culture the MAVpx clone (Vargas andOrozco 1993). Then, DNA was transferred to Nylon membranesand hybridized with the 500 bp probe for the16S rDNA gene asdescribed by Baez-Camargo et al. (in press).
In vivo [3H]thymidine labeling and isolationof the structures that incorporated [3H]thymidine
Trophozoites were grown for 14 h in TYI-S-33 medium supple-mented with 10 lCi [3H]thymidine/ml, washed with PBS pH 6.8and resuspended in a buffer containing 0.3 M sucrose, 1 mMEDTA and 10 mM TRIS-HCl, pH 8, before being mechanicallydisrupted in a Dounce homogenizer. The homogenate was centri-fuged three times at 2,500 rpm (Beckman TJ-C) at 4° C for 10 minto pellet the nuclei; the postnuclear supernatant was retained. Theintegrity of the nuclei was checked by phase-contrast microscopyand by autoradiography of the labeled nuclei. The postnuclearsupernatant, containing the EhkOs, was centrifuged again for15 min at 8 500 rpm (Beckman JA-20 rotor) and 4° C, and thepellet was resuspended in 4% paraformaldehyde, spread on slides,covered with autoradiographic film emulsion (Kodak), exposed for5 days in the dark, developed, stained with Giemsa, and observedwith the immersion objective of an optical microscope at 100 x.
Transmission electron microscopy (TEM) of DNA from EhkOand nuclear fractions prepared from clone A trophozoites
To perform TEM analysis, the EhkO and enriched nuclear frac-tions were treated for 1 h with 5 mg/ml proteinase K at 37° C,extracted once with phenol, and incubated for 1 h with 50 lg/mlRNase before extracting again with phenol:chloroform and pre-cipitating the DNA with ethanol. A hyperphase solution with 100ng of EhkO or nuclear DNA was prepared in a buffer containing200 mM TRIS-HCl (pH 7.5), 20 mM EDTA, 50% (v/v) formamideand 0.01 mg/ml cytochrome c. The solution was spread on a hy-pophase formed by 17% formamide, 100 mM TRIS-HCl (pH 8.0)and 1 mM EDTA, and samples were picked up on collodion-coated grids, and stained with 0.5 mM uranyl acetate. The gridswere rotary shadowed with platinum/palladium (80:20) at a 6 °angle (Davis et al. 1971). Circular pBR322 was used as a control forDNA integrity and for the absence of artifactual structures in thesamples. Samples were observed through a JEOL Temscan electronmicroscope.
Results
Localization of cytoplasmic DNA in acridineorange-stained live and paraformaldehyde-fixedtrophozoites
Most of the live and paraformaldehyde-fixed tropho-zoites treated with acridine orange showed green fluo-rescence in the central part of the nuclei, corresponding
251

to DNA (Fig. 1A–C), in agreement with the Feulgenstain experiments carried out by Pan and Geiman (1955)and by Albach et al. (1980), who found [3H]thymidine-labeled DNA in the endosome. Green fluorescence in thenuclei was surrounded by red or orange fluorescencecorresponding to RNA, in agreement to the results ofthe same authors. RNA was also very abundant in thecytoplasm (Fig. 1A–C). In 90% of the live and motile, aswell as the fixed trophozoites, green fluorescence wasalso detected in a cytoplasmic structure of a polymor-phic shape. The 50% of trophozoites with cytoplasmicfluorescence exhibited a green spot approximately 0.5–2.5 lm in diameter (Fig. 1A, B), brighter and smallerthan the nuclei; we called this structure the EhkO. Dif-ferences in fluorescence intensity between the nucleusand the EhKO may be due to differences in DNA con-tent or structure. The other 50% of the trophozoitespresented one or more fluorescent spots that weresmaller than 0.5 lm in diameter. Paraformaldehyde-fixed trophozoites displayed fluorescent patterns similarto those seen in live trophozoites (Fig. 1C). Tropho-zoites that had been treated for 6 h with DNase did notshow green fluorescence (Fig. 1D).
Labeling of EhkO by in situ hybridizationof the trophozoites of clone A with a 500 bp probefrom the E. histolytica 16S rDNA gene
In Fig. 2 we present details of the results obtained withtrophozoites of clone A by in situ hybridization with a500 bp fragment from the 5′ end of the 16S rDNA gene.In agreement with our previous results (Baez-Camargoet al., in press) the in situ hybridization experimentscarried out here showed the DNA to be distributed be-tween the nucleus and the cytoplasm (Fig. 2A–D, F). Aswas found in the acridine orange assays (Fig. 1), in pa-raformaldehyde-fixed trophozoites the ribosomal probedetected a faint signal in the nucleus and a strong signalin the cytoplasm (Fig. 2A–D). In most trophozoites thecytoplasmic hybridization was seen as a 0.5–2.5 lm long,compact V-shaped structure (Fig. 2A) and in 10–20% ofthe positive trophozoites we found a duplicate structurewhich may correspond to a dividing EhkO (Fig. 2B).
To prove that the hybridization signal indeed comesfrom DNA, we carried out several controls. (i) Thetrophozoites of clone A were also hybridized with a 1-kbfragment of the EhVR1 variable region located upstream
Fig. 1A–D Acridine orangestaining of trophozoites. A, B.Live trophozoites. C Para-formaldehyde-fixedtrophozoites. D Trophozoitesfixed with 2% paraformalde-hyde and treated with 50 UDNase for 6 h. A and D showwhole trophozoites, while B andC show the nucleus and EhkOarea. N, nucleus Bars, 5 lm
252

of the 5′ end of the 16S gene (Orozco et al. 1993). Hy-bridization with this fragment yielded the same results aswere obtained with the 5′ end of the 16S gene (Fig. 2D).(ii) The sensitivity of the EhkO signal to DNase wastested by incubating the trophozoites with 50 U of theenzyme for different times. Untreated trophozoitesshowed the 0.5–2.5 lm EhkO (Fig. 2A, B), after 3 hincubation they showed more diffuse labeling of theEhkO structure (Fig. 2C, D), and this disappearedcompletely after 6 h incubation (Fig. 2E). (iii) The tro-phozoites were hybridized with the single-stranded sensetranscript. No hybridization signal was detected inDNase treated trophozoites (Fig. 2E), while the un-treated trophozoites display the EhkO (Fig. 2F),strongly suggesting that the material in this structure isDNA. Results from these experiments suggest that theDNase acts first by relaxing the DNA and then digestingit. This assumption is supported by analogy with Try-panosomatidae, which have small histone-like proteinsthat protect kDNA from nuclease attack (Tittawellaet al. 1993). (iv) The trophozoites were grown in thepresence of [3H]uridine and submitted to RNase diges-tion for the same time as used for cells on slides. The
untreated trophozoites (Fig. 2G) showed labeled RNAdistributed throughout the cytoplasm, whereas theRNase treated cells showed no radioactive label(Fig. 2H). To ensure that no labeled RNA remained theslides were overexposed for three weeks. No RNA wasdetected after the RNase treatment, corroborating thatthe ribosomal probe hybridized specifically with DNA inthe RNase treated trophozoites used in our experiments.(v) The trophozoites were in situ hybridized with theBluescript phagemid in which the probes were cloned.The phagemid did not give any radioactive signal, rulingout the possibility of unspecific hybridization (Fig. 2I).
Hybridization of trophozoites from the axenicclones L6 and MAVax and the polyxenic cloneMAVpx with ribosomal DNA probes
In order to determine if EhkO was present in other E.histolytica clones and strains, we carried out the in situhybridization experiments with axenically grown tro-phozoites of clones MAVax and L6, and with thepolyxenically grown MAVpx trophozoites. EhkO was
Fig. 2A–I In situ hybridization of trophozoites with DNA probescontained in the EhVR1 fragment of the 16S rDNA gene.Paraformaldehyde-fixed trophozoites (4%) were treated with 60 URNase for 3 h and hybridized in situ with a denatured 500 bp double-stranded probe containing the 5′ end of the 16S rDNA gene (A–C) orwith the single-stranded sense transcript (E, F) or with a 1000 bpprobe located upstream of the 5′ end of the 16S rDNA gene (D). Insome experiments the trophozoites were incubated with 50 U DNaseat 37° C for 3 h incubated (C, D, F), or 6 h (E). Trophozoites were
also grown in the presence of 10 lCi/ml [3H]uridine for 24 h (G, H)and untreated (G) or treated (H) with 60 U RNase for 3 h.Trophozoites hybridized with the random primer labeled Bluescriptphagemid in which the probes were cloned are shown in I. Slides wereexposed for five days in the dark (A–F, I) or for three weeks in thedark (G, H) before developing and observing in the light microscope.Panels and H, I were photographed at 40 × and panelsA–G at 100 × magnification. N, nucleus. Bars: A–E = 6 lm, H, I =10 lm
253

detected in all clones tested (Fig. 3A–C). Shape and sizeof the EhkO in axenically growing clones L6 and MA-Vax presented similar characteristics to those exhibitedby the EhkO detected in clone A. Although hybridiza-tion signals in the polyxenically growing MAVpx tro-phozoites were larger (Fig. 3C), no cross hybridizationwith bacterial DNA was observed. Albach et al. (1980)reported that the cytoplasmic signal obtained in meta-bolically [3H]thymidine labeled trophozoites was due tobacterial contamination; however, none of our axenic-ally growing cultures showed any evidence of bacterialgrowth when plated in rich media. To confirm the ab-sence of cross hybridization of the ribosomal probeswith bacteria we hybridized TAFE fractionated totalDNA of clones A, MAVpx, MAVmx and MAVax, andDNA from intestinal bacteria (Fig. 3D), with a 3.5 kbribosomal probe containing the16S rDNA gene of E.histolytica and the variable region at the 5′ end of thisgene. Figure 3E shows that under the stringency condi-tions used in these experiments E. histolytica rDNA didnot cross-hybridize with bacterial DNA, in agreement
with our published results (Orozco et al. 1993). TheDNA of clone MAVpx gave a fainter hybridizationsignal than the DNA of the other clones; we believe thatthis is because the amount of E. histolytica DNA wasoverestimated due to the presence of bacteria growntogether with the trophozoites. These experiments con-firmed that the hybridization signals associated with theEhkO in the in situ hybridization experiments are notdue to bacterial contamination and that the EhkO ispresent in the cytoplasm of all four clones tested.
In vivo [3H]thymidine labeling of trophozoitesand isolation of an EhkO-enriched cell fraction
About 90% of trophozoites grown in the presence of[3H]thymidine showed autoradiographically detectablecytoplasmic label and half of them contained a labeledEhkO larger than 0.5 lm in diameter, and of variableshape (Fig. 4A). We also observed differences in labelingintensity between the nucleus and the EhKO, probablydue to different levels of thymidine incorporation duringthe cell cycle. We isolated an EhkO-enriched fractionfrom nuclei-free lysates of trophozoites that had beengrown for 14 h in the presence of [3H]thymidine. Theseexperiments were performed under the assumption thatthe DNA in EhkO would replicate once during the cellcycle, which lasts for 14–18 h in E. histolytica tropho-zoites (Orozco et al. 1988). EhkOs isolated by the pro-cedure used here increased in size, probably due to DNAexpansion, but retained their varied shapes. One hun-dred isolated EhkOs were analyzed and classified ac-cording to their shape and size; the one shown in Fig. 4Bis 5.1 and 6.2 lm long in its horizontal (minor) andvertical (major) axes, respectively; 20% of the structuresobserved had similar shapes ranging from 4.2 to 6.2 lmand 5 to 6.8 lm in their minor and major axes, respec-tively. The structure in Fig. 4C measures 3.2 × 6.1 lm inits highly labeled portion; 60% of the structures presentin the enriched fraction had a similar appearance. TheEhkO in Fig. 4D measures 3.5 × 12 lm and appears tobe disrupted; around 5% of the isolated EhkO had thisappearance and showed labeled DNA forming smallspots. Representative ‘‘replicating’’ EhkOs are presentedin Fig. 4E and F. The ‘‘boomerang’’ structure in Fig. 4E(see also Fig. 2A) displays symmetrical wings extendingabout 12.3 lm from end to end. Two nearly symmetrical,6.8 lm long EhkOs are shown in Fig. 4F; they may justhave finished dividing. About 15% of the structures an-alyzed seemed to be dividing. In 10% of the isolatedEhkOs only part of the DNA was labeled (Fig. 4B–D),suggesting that [3H]thymidine incorporation varies dur-ing the cell cycle. Nuclei pelleted by centrifugation of thedisrupted labeled trophozoites at 2 500 rpm appeared tobe intact; they did not expand as isolated EhkOs did,and ranged in length from 3 to 6 lm. In Fig. 4N we showtwo representative nuclei, the one on the left has anelongated shape typical of dividing nuclei (Solıs andBarrios 1991), whereas the nucleus at the right is a typ-
Fig. 3A–E Trophozoites of clones L6 (strain HM1:IMSS), MAVaxand of the MAVpx (strain MAV) hybridized in situ with the 500 bpprobe from the 16S rDNA gene. In situ hybridization of the axenicallygrown clones L6 (A) and MAV ax (B) and polyxenically grown cloneMAVpx (C) was done as in Fig. 2. In C bacteria (b) ingested by, andoutside of, the trophozoites are shown. D Total DNA of the clones A(lane 1), MAVpx (lane 2), MAVmx (lane 3), MAVax (lane 4) andbacteria (lane 5) (bacterial flora including: Escherichia coli, Clostridiumsymbiosum, Pseudomonas aeruginosa and Streptococcus fecalis) frac-tinated by tranverse alternating field electrophoresis, stained withethidium bromide (D). TAFE was carried out in two steps, at 280 mAfor 48 h with 3-min pulses, and then at 400 mA for 24 h with 8-minpulses DNA from TAFE gels was transferred to nylon membranesand hybridized (E) with the complete EhVR1 fragment including the16S rDNA and the variable region. Final washings were carried out at50° C in 0.1 × SSC and 0.1% SDS. M, Saccharomyces cerevisiaemolecular weight markers
254

ical interphase nucleus (Orozco et al. 1988). These resultsconfirm that E. histolytica has two different types ofDNA-containing structures: the nucleus and the EhkO.
Transmission electron microscopy (TEM)of the DNA contained in enriched EhkOand nuclear fractions
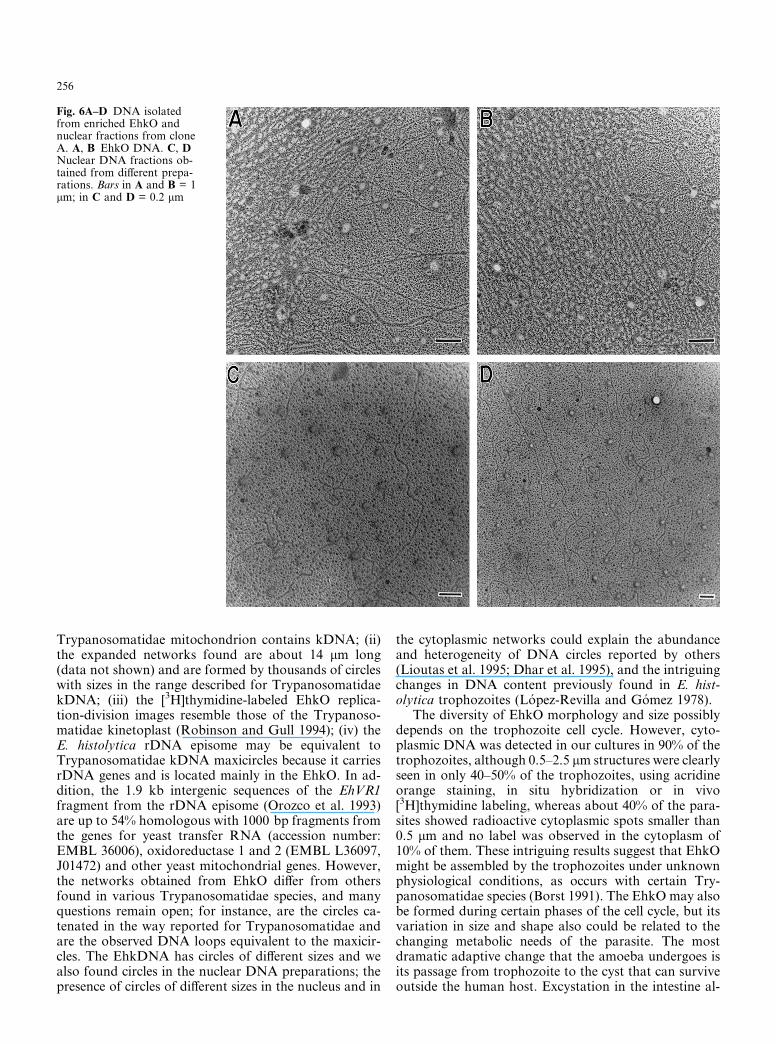
DNA was isolated from the EhkO and nuclear-enrichedfractions and observed by TEM. Intact and fragmentedDNA networks (EhkDNA) were observed in the EhkOfraction. A complete elongated network, approximately14.5 × 4 lm in size, composed of thousands of 0.4–0.8 kbminicircles with some DNA loops (maxicircles?) aroundthem, is shown in Fig. 5. Preparations incubated withDNase neither present any network nor DNA strands.Determination of the exact size and sequence of mini-circles and maxicircles depends on their future isolationand characterization, which is presently being carriedout in our laboratory. A magnification of a part of thenetwork is also shown in Fig. 5. We also observed par-tially lysed EhKOs with networks of DNA attached tothem (data not shown). In Fig. 6 we illustrate the dif-ferences between the EhkO and nuclear DNA structure.While the DNA from EhkO is mainly composed of freecircles, heterogeneous in size, and by networks of con-nected circles with loops in the extremes (Fig. 6A, B),the nuclear DNA did not exhibit network structures(Fig. 6C, D), but linear strands of different sizes andsome circles.
Discussion
The results presented here show for the first time thepresence of cytoplasmic DNA networks in E. histolyticatrophozoites; the networks resemble the kDNA of Try-
panosomatidae, and are contained in a structure calledthe EhkO by us. The following characteristics of theDNA-containing EhkO structure and of the EhkDNAsuggest an unexpectedly closer phylogenetic relationbetween E. histolytica and Trypanosomatidae: (i) theEhkO contains DNA networks (EhkDNA), just as the
Fig. 4A–F In vivo [3H]thymi-dine labeling of trophozoitesand isolated EhkO fractions.The trophozoites were grown inthe presence of 10 lCi [3H]thy-imidine for 14 h, washed twicewith PBS, spread on slides,covered with an emulsion filmand developed after 5 days ofexposure in the dark (A). Eh-kOs were isolated from a nu-cleus-free lysates, spread onslides, covered with emulsionand developed after 5 days(B–F). Nuclei were isolatedfrom the same trophozoites (N).Arrows in D point at small spotsof radiolabeled DNA.Bars = 10 lm
Fig. 5 Electron microscopy of isolated an EhkDNA network. Acomplete DNA network found in DNA purified from nuclei depletedlysate of E. histolytica clone A is shown. The inset is a magnified viewof the area in the square
255
Trypanosomatidae mitochondrion contains kDNA; (ii)the expanded networks found are about 14 lm long(data not shown) and are formed by thousands of circleswith sizes in the range described for TrypanosomatidaekDNA; (iii) the [3H]thymidine-labeled EhkO replica-tion-division images resemble those of the Trypanoso-matidae kinetoplast (Robinson and Gull 1994); (iv) theE. histolytica rDNA episome may be equivalent toTrypanosomatidae kDNA maxicircles because it carriesrDNA genes and is located mainly in the EhkO. In ad-dition, the 1.9 kb intergenic sequences of the EhVR1fragment from the rDNA episome (Orozco et al. 1993)are up to 54% homologous with 1000 bp fragments fromthe genes for yeast transfer RNA (accession number:EMBL 36006), oxidoreductase 1 and 2 (EMBL L36097,J01472) and other yeast mitochondrial genes. However,the networks obtained from EhkO differ from othersfound in various Trypanosomatidae species, and manyquestions remain open; for instance, are the circles ca-tenated in the way reported for Trypanosomatidae andare the observed DNA loops equivalent to the maxicir-cles. The EhkDNA has circles of different sizes and wealso found circles in the nuclear DNA preparations; thepresence of circles of different sizes in the nucleus and in
the cytoplasmic networks could explain the abundanceand heterogeneity of DNA circles reported by others(Lioutas et al. 1995; Dhar et al. 1995), and the intriguingchanges in DNA content previously found in E. hist-olytica trophozoites (Lopez-Revilla and Gomez 1978).
The diversity of EhkO morphology and size possiblydepends on the trophozoite cell cycle. However, cyto-plasmic DNA was detected in our cultures in 90% of thetrophozoites, although 0.5–2.5 lm structures were clearlyseen in only 40–50% of the trophozoites, using acridineorange staining, in situ hybridization or in vivo[3H]thymidine labeling, whereas about 40% of the para-sites showed radioactive cytoplasmic spots smaller than0.5 lm and no label was observed in the cytoplasm of10% of them. These intriguing results suggest that EhkOmight be assembled by the trophozoites under unknownphysiological conditions, as occurs with certain Try-panosomatidae species (Borst 1991). The EhkO may alsobe formed during certain phases of the cell cycle, but itsvariation in size and shape also could be related to thechanging metabolic needs of the parasite. The mostdramatic adaptive change that the amoeba undergoes isits passage from trophozoite to the cyst that can surviveoutside the human host. Excystation in the intestine al-
Fig. 6A–D DNA isolatedfrom enriched EhkO andnuclear fractions from cloneA. A, B EhkO DNA. C, DNuclear DNA fractions ob-tained from different prepa-rations. Bars in A and B = 1lm; in C and D = 0.2 lm
256

lows the parasite to invade the host tissues, where it mustadapt to various oxygen concentrations, which differ inthe intestine, liver and feces, the three main habitats of E.histolytica during its life cycle. The Krebs cycle and therespiratory chain have not been found to operate in E.histolytica trophozoites, which are known to have affinityfor oxygen (Weinbach and Diamond 1974), and to pos-sess CoQ (Ellis et al. 1994). In T. brucei bloodstreamforms, ATP is not synthesized by the Krebs cycle and therespiratory chain is absent in the mitochondrion (Borst1991). It is known that T. brucei lives for long periodswithout functional mitochondria and represses mi-tochondrial biogenesis surviving by glycolysis while inthe bloodstream of the mammalian host (Borst 1991).The kinetoplast provides metabolic energy to the Try-panosomatidae (Simpson 1987), whose flagellae are lo-cated right next to the kDNA. E. histolytica trophozoiteshave no flagellae but move actively by forming pseudo-podia; their relationship with the EhkO is unknown.
Acknowledgements We thank Pedro Chavez and the CINVES-TAV’s Electron Microscopy Unit and Department of Photographyfor excellent technical work. E. Orozco is an International Fellowof the Howard Hughes Medical Institute (USA). This work wasalso supported by CONACYT (Mexico), and INSERM (France).
References
Albach RA, Shaffer JG, Watson RH (1969) Autoradiographicstudies of 3H-thymidine uptake in Entamoeba histolytica in theCLG medium. J Protozool 13:349–355
Albach RA, Booden T, Boonlayagoor P, Dowing SW (1980)Concepts of function of peripheral non-chromatin and endo-some in Entamoeba histolytica. Arch Inv Med 11:63–73
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG,Smith JA, Struhl K (1993). Current protocols in molecular bi-ology. Wiley and Sons, New York
Baez-Camargo M, Riveron AM, Delgadillo DM, Flores E, San-chez T, Orozco E Entamoeba histolytica: gene linkage groupsand relevant features of its karyotype. Mol Gen Genet, in press
Borst P (1991) Why kinetoplast DNA networks? Trends Genet7:139–141
Cavalier-Smith T (1993) Kingdom Protozoa and its 18 phyla.Microbiol Rev 57:953–994
Cozzarelli NR (1993) The structure and function of DNA super-coiling and catenanes. The Harvey Lectures 1991–1992. Series87. Wiley-Liss, New York, pp 35–55
Clark GC, Roger AJ (1995) Direct evidence for secondary loss ofmitochondria in Entamoeba histolytica. Proc Natl Acad SciUSA 92, 6518–6521
Davis RW, Simon M, Davidson N (1971) Electron microscopeheteroduplex methods for mapping regions of base sequencehomology in nucleic acids. Methods Enzymol 21D:413–428
Diamond LS, Harlow DR, Cunick CC (1978) A new medium forthe axenic cultivation of Entamoeba histolytica and other Ent-amoeba. Trans R Soc Trop Med Hyg 72:431–432
Dhar SK, Choundhury NR, Bhattacharya A, Bhattacharya S(1995) A multitude of circular DNAs exist in the nucleus ofEntamoeba histolytica. Mol Biochem Parasitol 70:203–206
Ellis JE, Setchel DRK, Kaneshiro ES (1994) Detection of ubiqui-none in parasitic and free-living protozoa, including speciesdevoid of mitochondria. Mol Biochem Parasitol 65:213–224
Ferguson ML, Torri AF, Perez-Morga D, Ward DC, Englund PT(1994) Kinetoplast DNA replication: mechanistic differencesbetween Trypanosoma brucei and Crithidia fasciculata. J CellBiol 126:631–639
Hasegawa M, Hashimoto T, Adachi J, Iwabe N, Miyata T (1993)Early branchings in the evolution of eukaryotes: ancient di-vergence of Entamoeba that lacks mitochondria revealed byprotein sequence data. J Mol Evol 36:380–388
Lioutas C, Schmetz C, Tannich E (1995) Identification of variouscircular DNA molecules in Entamoeba histolytica. Exp Parasitol80, 349–352
Lopez-Revilla R, Gomez R (1978) Entamoeba histolytica, E. inva-dens, and E. moshkowkii: fluctuations of DNA content ofaxenic trophozoites. Exp Parasitol 44:243–248
Martınez-Palomo A (1986) Amebiasis Editorial Medica Panamer-icana, Mexico City pp 29
Orozco E, Guarneros G, Martınez-Palomo A, Sanchez T (1983)Entamoeba histolytica: Phagocytosis as a virulence factor J ExpMed 158:1511–1521
Orozco E, Solıs F, Domınguez J, Chavez B, Hernandez F (1988)Entamoeba histolytica: cell cycle and nuclear division. Exp Pa-rasitol 67:85–95
Orozco E, Lazard D, Sanchez T, Sanchez MA, Hernandez R,Felix-Silva E (1993) A variable DNA region of Entamoebahistolytica is expressed in several transcripts which differ ingenetically related clones. Mol Gen Genet 241:271–279
Pan CT, Geiman QM (1955) Comparative studies of intestinalamebae. Distribution and cyclic changes of the nucleic acids inEntamoeba histolytica and Entamoeba coli. Amer J Hyg 62:66–70
Perez-Morga D, Englund PT (1993) The attachment of minicircles tokinetoplast DNA networks during replication. Cell 74:703–711
Robinson DR, Gull K (1994) The configuration of DNA replica-tion sites within the Trypanosoma brucei kinetoplast. J Cell Biol126:641–648
Simpson L, Silva AD (1971) Isolation and characterization ofkinetoplast DNA from Leishmania tarentolae. J Mol Biol56:443–473
Simpson L (1987) The mitochondrial genome of kinetoplastidprotozoa: genomic organization, transcription, replication andevolution. Annu Rev Microbiol 41:363–382
Solıs FJ, Barrios R (1991) Entamoeba histolytica microtubulemovement during mitosis. Exp Parasitol 73:276–284
Tittawella I, Carlsson L, Thornell LE (1993) Two proteins involvedin kinetoplast compaction. FEBS Lett 333:5–9
Vargas MA, Orozco E (1993) Entamoeba histolytica: changes in thezymodeme of cloned non-pathogenic trophozoites culturedunder different conditions. Parasitol Res 19:353–356
Weinbach EC, Diamond LS (1974) Entamoeba histolytica I. Aer-obic metabolism. Exp Parasitol 35:232–243
257
Copyright © 2022 FDOKUMEN




















