
Differentiation of Entamoeba histolytica: A possible role for enolase

mcpt0fe
Archives of Biochemistry and BiophysicsVol. 375, No. 2, March 15, pp. 304–314, 2000doi:10.1006/abbi.1999.1592, available online at http://www.idealibrary.com on
Ectonucleotide Diphosphohydrolase Activitiesin Entamoeba histolytica
Fernanda S. Barros,* Lucia F. De Menezes,† Ana A. S. Pinheiro,* Edward F. Silva,‡Angela H. C. S. Lopes,§ Wanderley De Souza,† and Jose R. Meyer-Fernandes*,1
*Departamento de Bioquımica Medica, Instituto de Ciencias Biomedicas, †Instituto de Biofısica Carlos Chagas Filho,§Instituto de Microbiologia Professor Paulo de Goes, Universidade Federal do Rio de Janeiro, CCS,Bloco H, Cidade Universitaria, Ilha do Fundao, 21541-590, Rio de Janeiro, RJ, Brazil; and‡Departamento de Parasitologia, Universidade Federal de Minas Gerais, MG, Brazil
Received July 21, 1999, and in revised form October 15, 1999
tc
ui
et
In this work, we describe the ability of living cells ofEntamoeba histolytica to hydrolyze extracellular ATP.In these intact parasites, whose viability was deter-mined by motility and by the eosin method, ATP hy-drolysis was low in the absence of any divalent metal(78 nmol Pi/h/105 cells). Interestingly, in the presenceof 5 mM MgCl2 an ecto-ATPase activity of 300 nmolPi/h/105 cells was observed. The addition of MgCl2 tothe extracellular medium increased the ecto-ATPaseactivity in a dose-dependent manner. At 5 mM ATP,half-maximal stimulation of ATP hydrolysis was ob-tained with 1.23 mM MgCl2. Both activities were linearwith cell density and with time for at least 1 h. Theecto-ATPase activity was also stimulated by MnCl2
and CaCl2 but not by SrCl2, ZnCl2, or FeCl3. In fact,FeCl3 inhibited both Mg21-dependent and Mg21-inde-pendent ecto-ATPase activities. The Mg21-indepen-dent ATPase activity was unaffected by pH in therange between 6.4 and 8.4, in which the cells wereviable. However, the Mg21-dependent ATPase activitywas enhanced concomitantly with the increase in pH.In order to discard the possibility that the ATP hydro-lysis observed was due to phosphatase or 5*-nucleoti-dase activities, several inhibitors for these enzymeswere tested. Sodium orthovanadate, sodium fluoride,levamizole, and ammonium molybdate had no effecton the ATPase activities. In the absence of Mg21 (basalactivity), the apparent Km for ATP42 was 0.053 6 0.008
M, whereas at saturating MgCl2 concentrations, theorresponding apparent Km for Mg-ATP22 for Mg21-de-endent ecto-ATPase activity (difference between to-al and basal ecto-ATPase activity) was 0.503 mM 6.062. Both ecto-ATPase activities were highly specificor ATP and were also able to hydrolyze ADP lessfficiently. To identify the observed hydrolytic activi-
1 To whom correspondence should be addressed. Fax: 155-21-270-8647. E-mail: [email protected].
304
ies as those of an ecto-ATPase, we used suramin, aompetitive antagonist of P2 purinoreceptors and an
inhibitor of some ecto-ATPases, as well as the imper-meant agent 4*-4*-diisothiocyanostylbenzene-2*-2*-dis-
lfonic acid. These two reagents inhibited the Mg21-ndependent and the Mg21-dependent ATPase activi-
ties to different extents, and the inhibition by bothagents was prevented by ATP. A comparison amongthe ecto-ATPase activities of three amoeba speciesshowed that the noninvasive E. histolytica and thefree-living E. moshkovskii were less efficient than thepathogenic E. histolytica in hydrolyzing ATP. As E.histolytica is known to have a galactose-specific lectinon its surface, which is related to the pathogenesis ofamebiasis, galactose was tested for an effect on ecto-ATPase activities. It stimulated the Mg21-dependentcto-ATPase but not the Mg21-independent ATPase ac-ivity. © 2000 Academic Press
Key Words: Entamoeba histolytica; ecto-ATPase;ecto-phosphatase.
Entamoeba histolytica is an enteric protozoan thatinfects 10% of the world’s human population. This par-asite causes 50 to 100 million cases of invasive colitis orliver abscess and up to 100,000 deaths per year (1, 2).Amebiasis is one of the great parasitic diseases of hu-mankind, which predominantly affects individuals oflower socioeconomic status, who live in developingcountries. During its life cycle, E. histolytica exists ineither trophozoite or cyst form. Infection is acquired byingestion of the cyst form, which is resistant to the acidpH of the stomach, and excystation occurs in the smallbowel, in which the cysts divide and differentiate intotrophozoites (1). The pathogenic, invasive E. histolyticaand the most prevalent, nonpathogenic E. dispar aretwo morphologically indistinguishable species that ex-
0003-9861/00 $35.00Copyright © 2000 by Academic Press
All rights of reproduction in any form reserved.

idOctac
hgpecreacn(hc
oC
aeztcpawE
s
np
305ECTO-ATPases IN Entamoeba histolytica
hibit several antigenic, genetic, and isoenzymatic dif-ferences (1, 3).
Cell–cell recognition and adherence are processescentral to many fundamental areas of biology. Onecommon theme for many pathogenic microorganisms isthe use of microbial lectins for colonization and inva-sion. It is not surprising that lectin-based protein–carbohydrate interactions are important in adherence,as the extracellular surfaces of eukaryotic cells are richin complex carbohydrates that are attached to glycop-roteins and glycolipids. Lectins make up a class ofproteins that bind carbohydrates specifically and non-covalently (4). Parasite recognition of glycoconjugatesplays an important role in the pathogenesis of amebi-asis. Killing of host cells by E. histolytica trophozoitesn vitro occurs only upon direct contact, which is me-iated by an amebic adhesin that recognizes N- and-linked oligosaccharides (5). This adhesin is specifi-
ally inhibited by millimolar concentrations of galac-ose (Gal)2 and N-acetyl-D-galactosamine (GalNAc)nd avoidance of membrane lysis by the complementomplex attack is also mediated by this lectin (6).
During the last two decades, considerable progressas been achieved in the study of ectonucleotidases ineneral (7) and ecto-ATPases in particular (8, 9). Thisrogress is related to the finding that, in contrast to thestablished view, nucleotides can be found in signifi-ant concentrations outside cells (9, 10). Nucleotideseleased to the extracellular medium may exert theirffects on other cells in the vicinity of the secretion sitend modulate biological processes by binding to specificell surface receptors (9). ATP may be one of the sig-aling molecules involved in cell-mediated cytotoxicity11). It has been demonstrated that extracellular ATPas profound effects on cellular functions (11, 12),ausing plasma membrane depolarization, Ca21 influx,
and cell death (11, 12). Fillipini et al. (12) have shownthat ATP can kill various types of cells, with the ex-ception of those that express a high level of ATP-break-down activity on their surface.
Ecto-ATPases are glycoproteins present in theplasma membrane and have their active sites facingthe external medium rather than the cytoplasm (8).The activities of these enzymes can be measured usingviable cells and can be modulated by divalent cations,such as Mg21 and Ca21 (13). A previous study on nucle-tidases of E. histolytica (14) described the activity of aa21-dependent nucleoside diphosphatase associated
with the particulate fraction. This enzyme was shownto be responsible for the hydrolysis of ADP, UDP, GDP,and thiamine pyrophosphate. Subsequently, McCauland Bird (15) showed that thiamine pyrophosphatase
2 Abbreviations used: Gal, galactose; GalNAc, N-acetyl-D-galac-tosamine; p-NPP, p-nitrophenylphosphate; DIDS, 49-49-diisothiocya-
ostylbenzene-29-29-disulfonic acid; NTPase, nucleoside triphos-hate hydrolase.
was associated with the cytoplasmic vacuole of thisparasite. Moreover, Serrano et al. (16) reported thepresence of a Mg21-dependent ATPase activity in theplasma membrane of this parasite. Recently, upon ad-dition of ATP to intact cells of E. histolytica, it wasdemonstrated that a Ca21–ATPase activity of this par-site is exposed to the medium and is inhibited byxtracellular addition of the impermeant reagent dia-otized sulfanilic acid (17). It has been proposed thathis E. histolytica ecto-ATPase could participate in ahain of nucleosidase activities on the surface of thearasite and that this pathway could provide the par-sites with a source of extracellular adenosine (17),hich is known to be part of the dietary requirement of. histolytica and is taken up by these cells (18).Here, we show the presence of Mg21-independent
and Mg21-dependent ecto-ATPase activities on the cellurface of living E. histolytica trophozoites. We char-
acterize the properties of these enzymes and demon-strate the effects of galactose. We also compare ATPhydrolysis by the invasive E. histolytica with that cat-alyzed by the noninvasive E. histolytica and the free-living E. moshkovskii.
MATERIAL AND METHODS
Reagents. All reagents were purchased from E. Merck (D-6100Darmstadt, Germany) or Sigma Chemical Co. (St. Louis, MO).[g32P]ATP was prepared as described by Glynn and Chappell (19).Distilled water was deionized using a MilliQ system of resins (Mil-lipore Corp., Bedford, MA) and was used in the preparation of allsolutions. Concentrations of free and complexed species at equilib-rium were calculated by using an iterative computer program thatwas modified (20) from that described by Fabiato and Fabiato (21).
Microorganisms. Invasive E. histolytica (strain HM1:IMSS),noninvasive E. histolytica (strain RPS) (22), and free-living E. mosh-kovskii (strain FIC) trophozoites were cultured axenically in TYI-S-33 medium (22, 23), supplemented with 10% adult bovine serumand 3% vitamin (Diamond Vitamin, Tween 80; JRH Biosciences) at37°C, and were harvested at the stationary phase of growth, 48 hafter inoculation. The parasites were then collected by centrifuga-tion, washed twice, and kept in 116.0 mM NaCl, 5.4 mM KCl, 5.5 mMD-glucose, 50.0 mM Hepes, pH 7.2. Cellular viability was assessed,before and after incubations, by motility and the eosin method (23).The viability was not affected by the conditions employed.
Ecto-ATPase activity measurements. Intact cells were incubatedfor 1 h at 30°C in 0.5 ml of a mixture containing, unless otherwisespecified, 116.0 mM NaCl, 5.4 mM KCl, 5.5 mM D-glucose, 50.0 mMHepes–Tris buffer, pH 7.2, 5.0 mM ATP, and 1.0 3 106 cells/ml, inthe presence or in the absence of 5.0 mM MgCl2. The ATPase activitywas determined by measuring the hydrolysis of [g-32P]ATP (104
Bq/nmol ATP) (24). The experiments were started by the addition ofliving cells and terminated by the addition of 1.0 ml of a cold mixturecontaining 0.2 g charcoal in 1.0 M HCl. The tubes were then centri-fuged at 2000 rpm for 10 min at 4°C. Aliquots (0.5 ml) of thesupernatant containing the released 32Pi were transferred to scintil-lation vials containing 9.0 ml of scintillation fluid (2.0 g PPO in 1 literof toluene). The ATPase activity was calculated by subtracting thenonspecific ATP hydrolysis measured in the absence of cells. TheATP hydrolysis was linear with time under the assay conditions usedand was proportional to the cell number. In the experiments in whichother nucleotides were used, the hydrolytic activity measured underthe same conditions described above was assayed spectrophotometri-
tscmo
ewe(4
cs
paast
nM
bhn
Tlsratcsp
lai
lh
mw
ipF
wi
m(bi5wMpn
306 BARROS ET AL.
cally by measuring the release of Pi from the nucleotides (25). Thevalues obtained for ATPase activities measured using both methods(colorimetric and radioactive) are exactly the same. In the experi-ments in which high concentrations of Ca21, Mn21, and Fe31 wereested, possible precipitates formed were checked as previously de-cribed (26). Under the conditions employed, in the reaction mediumontaining 50 mM Hepes, pH 7.2, 116 mM NaCl, 5.4 mM KCl, 5.5M D-glucose, and 5 mM ATP, no phosphate precipitates were
bserved in the presence of these cations.Phosphatase measurements. In addition to the measurements of
cto-ATPase activity, the ecto-p-nitrophenylphosphatase activityas determined in the same medium as that for ATP hydrolysis,xcept that ATP was replaced by 5.0 mM p-nitrophenylphosphatep-NPP). The reaction was determined spectrophotometrically at25.0 nm using an extinction coefficient of 14.3 3 103 M21 cm21.Statistical analysis. All experiments were performed in tripli-
ate, with similar results obtained in at least three separate celluspensions. Apparent Km and Vmax values were calculated using a
computerized nonlinear regression analysis of the data to theMichaelis–Menten equation (24). Statistical significance was deter-mined by Student’s t test. Significance was considered as P , 0.05.
RESULTS
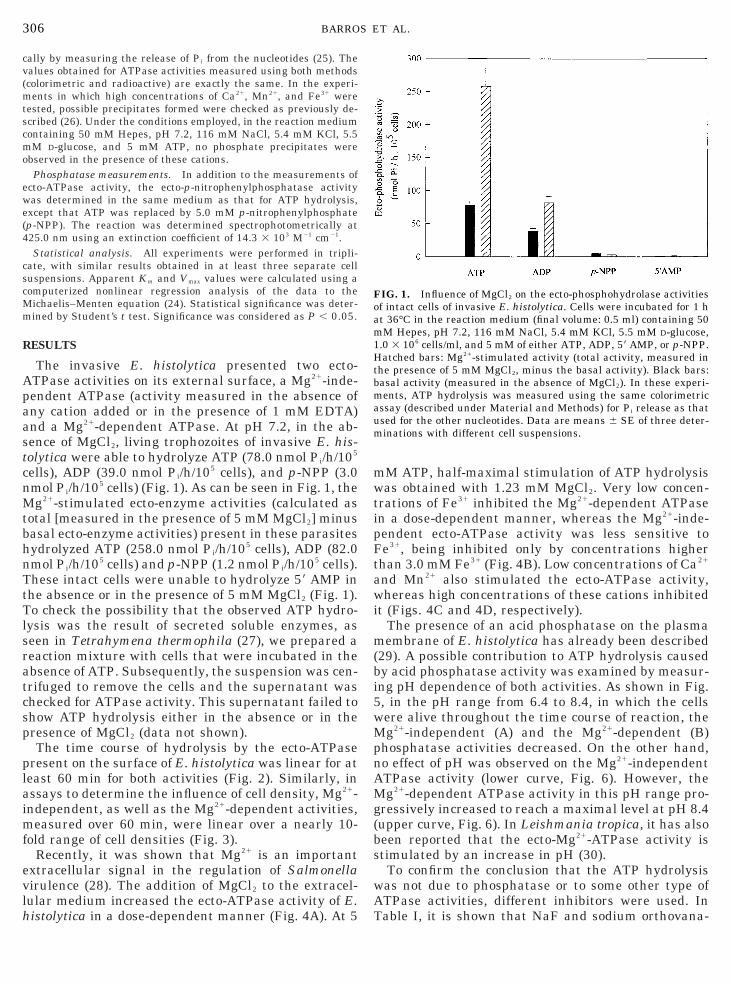
The invasive E. histolytica presented two ecto-ATPase activities on its external surface, a Mg21-inde-
endent ATPase (activity measured in the absence ofny cation added or in the presence of 1 mM EDTA)nd a Mg21-dependent ATPase. At pH 7.2, in the ab-ence of MgCl2, living trophozoites of invasive E. his-olytica were able to hydrolyze ATP (78.0 nmol Pi/h/105
cells), ADP (39.0 nmol Pi/h/105 cells), and p-NPP (3.0mol Pi/h/105 cells) (Fig. 1). As can be seen in Fig. 1, theg21-stimulated ecto-enzyme activities (calculated as
total [measured in the presence of 5 mM MgCl2] minusasal ecto-enzyme activities) present in these parasitesydrolyzed ATP (258.0 nmol Pi/h/105 cells), ADP (82.0mol Pi/h/105 cells) and p-NPP (1.2 nmol Pi/h/105 cells).
These intact cells were unable to hydrolyze 59 AMP inthe absence or in the presence of 5 mM MgCl2 (Fig. 1).
o check the possibility that the observed ATP hydro-ysis was the result of secreted soluble enzymes, aseen in Tetrahymena thermophila (27), we prepared aeaction mixture with cells that were incubated in thebsence of ATP. Subsequently, the suspension was cen-rifuged to remove the cells and the supernatant washecked for ATPase activity. This supernatant failed tohow ATP hydrolysis either in the absence or in theresence of MgCl2 (data not shown).The time course of hydrolysis by the ecto-ATPase
present on the surface of E. histolytica was linear for ateast 60 min for both activities (Fig. 2). Similarly, inssays to determine the influence of cell density, Mg21-ndependent, as well as the Mg21-dependent activities,
measured over 60 min, were linear over a nearly 10-fold range of cell densities (Fig. 3).
Recently, it was shown that Mg21 is an importantextracellular signal in the regulation of Salmonellavirulence (28). The addition of MgCl2 to the extracel-ular medium increased the ecto-ATPase activity of E.istolytica in a dose-dependent manner (Fig. 4A). At 5
M ATP, half-maximal stimulation of ATP hydrolysisas obtained with 1.23 mM MgCl2. Very low concen-
trations of Fe31 inhibited the Mg21-dependent ATPasen a dose-dependent manner, whereas the Mg21-inde-endent ecto-ATPase activity was less sensitive toe31, being inhibited only by concentrations higher
than 3.0 mM Fe31 (Fig. 4B). Low concentrations of Ca21
and Mn21 also stimulated the ecto-ATPase activity,hereas high concentrations of these cations inhibited
t (Figs. 4C and 4D, respectively).The presence of an acid phosphatase on the plasmaembrane of E. histolytica has already been described
29). A possible contribution to ATP hydrolysis causedy acid phosphatase activity was examined by measur-ng pH dependence of both activities. As shown in Fig., in the pH range from 6.4 to 8.4, in which the cellsere alive throughout the time course of reaction, theg21-independent (A) and the Mg21-dependent (B)
hosphatase activities decreased. On the other hand,o effect of pH was observed on the Mg21-independent
ATPase activity (lower curve, Fig. 6). However, theMg21-dependent ATPase activity in this pH range pro-gressively increased to reach a maximal level at pH 8.4(upper curve, Fig. 6). In Leishmania tropica, it has alsobeen reported that the ecto-Mg21-ATPase activity isstimulated by an increase in pH (30).
To confirm the conclusion that the ATP hydrolysiswas not due to phosphatase or to some other type ofATPase activities, different inhibitors were used. InTable I, it is shown that NaF and sodium orthovana-
FIG. 1. Influence of MgCl2 on the ecto-phosphohydrolase activitiesof intact cells of invasive E. histolytica. Cells were incubated for 1 hat 36°C in the reaction medium (final volume: 0.5 ml) containing 50mM Hepes, pH 7.2, 116 mM NaCl, 5.4 mM KCl, 5.5 mM D-glucose,1.0 3 106 cells/ml, and 5 mM of either ATP, ADP, 59 AMP, or p-NPP.Hatched bars: Mg21-stimulated activity (total activity, measured inthe presence of 5 mM MgCl2, minus the basal activity). Black bars:basal activity (measured in the absence of MgCl2). In these experi-ments, ATP hydrolysis was measured using the same colorimetricassay (described under Material and Methods) for Pi release as thatused for the other nucleotides. Data are means 6 SE of three deter-minations with different cell suspensions.

awtpaMnpoco
ahdho
etT
n
n
a
307ECTO-ATPases IN Entamoeba histolytica
date, two potent inhibitors of acid phosphatase activi-ties (31, 32), strongly inhibited the Mg21-independentnd the Mg21-dependent phosphatase activities,hereas no effect was observed on the ATPase activi-
ies. Levamizole, a specific inhibitor of alkaline phos-hatases (33, 34), also failed to inhibit the ATPasectivities and the phosphatase activities (Table I).oreover, p-NPP, a substrate for phosphatases, did
ot inhibit the ATPase activities. Another possible ex-lanation for the ATP hydrolysis might be a 59-nucle-tidase (35). However, as shown in Fig. 1, these intactells were unable to hydrolyze 59 AMP in the absencer in the presence of 5 mM MgCl2. Moreover, the lack of
response to ammonium molybdate (Table I), a potentinhibitor of 59-nucleotidase activity (35), indicates thatthis enzyme did not contribute to the ATP hydrolysis.It is also shown in Table I that the ATPase and phos-
FIG. 2. Time course of the ecto-ATPase activities of intact cells of inmedium (final volume: 0.5 ml) containing 50 mM Hepes, pH 7.2, 116Tris–ATP [g-32P]ATP [sp act 5 104 Bq/nmol ATP], without (A) or wdeterminations with different cell suspensions.
FIG. 3. Cell density dependence of the ecto-ATPase activity of invreaction medium (final volume: 0.5 ml) as that described in the legere means 6 SE of three determinations with different cell suspens
phatase activities were insensitive to oligomycin andsodium azide, two known inhibitors of mitochondrialMg-ATPase, to bafilomycin A1, a V-ATPase inhibitor,nd to ouabain, a Na/K-ATPase inhibitor (36). The ATPydrolysis described here may be due to an ATP-iphosphohydrolase activity, as it was inhibited byigh concentrations of ADP and AMP, but not by aden-sine (Table I).The Mg21-dependent and the Mg21-independent
cto-ATPase activities showed different kinetic pat-erns with respect to the ATP concentration (Fig. 7).he Mg21-independent activity had an apparent Km for
ATP42 of 0.053 6 0.008 mM and a Vmax of 87.6 6 9.2mol Pi/h/105 cells (Fig. 7A). For the Mg21-dependent
ecto-ATPase activity, the apparent Km for MgATP22
was 0.503 6 0.062 mM and the Vmax was 276.6 6 30.6mol Pi/h/105 cells (Fig. 7B). We also analyzed the
ive E. histolytica. Cells were incubated for 1 h at 36°C in the reactionNaCl, 5.4 mM KCl, 5.5 mM D-glucose, 1 3 106 cells/ml, and 5 mM(B) the addition of 5 mM MgCl2. Data are means 6 SE of three
ve E. histolytica. Cells were incubated for 1 h at 36°C in the sameof Fig. 2, without (A) or with (B) the addition of 5 mM MgCl2. Datas.
vasmMith
asindion

) thm .
5od
308 BARROS ET AL.
specificity of these ecto-ATPase activities for variousother nucleotides. In Table II, it is shown that ATP wasthe best substrate for these enzymes, although they
FIG. 4. Influence of different cation concentrations on the ecto-Aincubated for 1 h at 36°C in the same reaction medium (final volumincreasing concentrations of Mg21 (A), Fe31 [with (F) or without (E
eans 6 SE of three determinations with different cell suspensions
FIG. 5. Effect of pH on the p-nitrophenylphosphatase activities of inin the reaction medium (final volume: 0.5 ml) containing 116 mM Na0 mM Hepes–Tris buffer, adjusted to pH values between 6.4 and 8.f the experiments. The assays were performed without (A) or wieterminations with different cell suspensions.
were also able to hydrolyze ADP. Other nucleoside59-triphosphates (ITP, CTP, GTP, and UTP) producedvery low reaction rates.
ase activities of intact cells of invasive E. histolytica. Cells were.5 ml) as that described in the legend of Fig. 2, with the addition ofe addition of 5 mM MgCl2] (B), Ca21 (C), and Mn21 (D). Data are
ct cells of invasive E. histolytica. Cells were incubated for 1 h at 36°C5.4 mM KCl, 5.5 mM D-glucose, 5 mM p-NPP, 1.0 3 106 cells/ml, andn this pH range (6.4–8.4), the cells were viable thougout the course(B) the addition of 5 mM MgCl2. Data are means 6 SE of three
TPe: 0
taCl,4. Ith

AhasewAaasaAasiM
it
a
A
mh
aia
Adf
rm[b(ma
309ECTO-ATPases IN Entamoeba histolytica
The Mg21-dependent and the Mg21-independentTPase activities present on the external surface of E.istolytica were the same when cells were in log phases when they were in stationary phase (data nothown). Since we used intact cells for measuring thenzyme activities in all the experiments done in thisork, it likely that the described activities are ecto-TPases. To confirm this, we applied the criterion thatn authentic ectoenzyme should be inhibited by andded extracellular impermeant reagent (13, 30, 37)uch as 49-49-diisothiocyanostylbene-29-29-disulfoniccid (DIDS) (30, 37), and by an inhibitor of some ecto-TPases, such as suramin, which is also a competitiventagonist of P2-purinergic receptors (38). Figure 8hows that 1 mM DIDS inhibited 100% of the Mg21-ndependent ecto-ATPase activity (A) and 60% of the
g21-dependent activity (B) and that 1 mM suraminpromoted a 54% reduction of the E. histolytica Mg21-ndependent ecto-ATPase activity (A) and a 64% reduc-ion of the Mg21-dependent activity (B). The inhibition
promoted by either DIDS or suramin was suppressedwhen the cells were preincubated in the presence of 5mM ATP (Figs. 8A and 8B, insets).
The noninvasive E. histolytica and the free-living E.moshkovskii were compared to the pathogenic E. his-tolytica concerning the presence of Mg21-dependentnd Mg21-independent ecto-ATPase activities. As
shown in Fig. 9, all three species express the Mg21-independent (A) and the Mg21-dependent (B) ecto-
TPase activities, and pathogenic E. histolytica showed
FIG. 6. Effects of pH on the ecto-ATPase activities of intact cells ofinvasive E. histolytica. Cells were incubated for 1 h at 36°C in theeaction medium (final volume: 0.5 ml) containing 116 mM NaCl, 5.4M KCl, 5.5 mM D-glucose, 1 3 106 cells/ml, 5 mM Tris–ATP
g-32P]ATP [sp act 5 104 Bq/nmol ATP], and 50 mM Hepes–Trisuffer, adjusted to pH values between 6.4 and 8.4. In this pH range6.4–8.4) the cells were viable througout the course of the experi-ents. The assays were performed with (F) or without (E) the
ddition of 5 mM MgCl2. Data are means 6 SE of three determina-tions with different cell suspensions.
uch higher ATPase activities than the noninvasive E.istolytica and the free-living E. moshkovskii.It is well known that the pathogenesis of invasive
mebiasis is due to an amebic Gal- and GalNAc-inhib-table adherence lectin that exclusively mediates thedherence of E. histolytica to mammalian target cells
(6, 39, 40). The physiological role of the ecto-ATPases isstill unknown, but a possible involvement in cellularadhesion has been suggested (41–43). For these rea-sons, we examined the effect of D-galactose on the ecto-
TPase activities of this parasite (Fig. 10). The Mg21-ependent ecto-ATPase was stimulated more than two-old by 50 mM D-galactose (Fig. 10B), although the
Mg21-independent ecto-ATPase was not affected bythis carbohydrate (Fig. 10A). D-Glucose had no effect oneither of the hydrolytic activities tested.
DISCUSSION
In this paper, we report the characterization of Mg21-independent and Mg21-dependent ecto-ATPase activi-ties present on the external surface of E. histolytica(Fig. 1). Cellular viability was assessed, before andafter the reactions, by motility and by the eosin method(23). The viability was not affected by any of the con-ditions used in the assays. The external localization ofthe ATP-hydrolyzing site is supported by its sensitivityto the impermeant reagent DIDS and to suramin,which is a noncompetitive inhibitor of ecto-ATPasesand an antagonist of P2 purinoreceptors, which medi-ate the physiological functions of extracellular ATP(38, 44). Also, the use of a battery of inhibitors for otherATPases that have intracellular ATP binding sitesshowed no change in the ATPase activities, either inthe absence or in the presence of Mg21 (Table I). For theabove reasons, we interpret our data according to thecriteria for ectolocalization of the ATPase activitieshere described (8). These criteria include the use ofnonpenetrating substrates, which have access to theenzyme in whole, intact cells, the demonstration ofproduct formation extracellularly, and the demonstra-tion that the activity can be inhibited by exposure ofintact cells to impermeant reagents (13, 30, 45). Ourdata are consistent with observations showing thatecto-ATPases are widely distributed in nature, as theyhave been described in plants (46), several mammaliancell types (8, 47), insects (48), helminth endoparasites(49, 50), and protozoa, including T. thermophila (27),Leishmania tropica (30), Toxoplasma gondii (51), andE. histolytica (17), among others.
The ATPase activities shown in this work cannot beattributed to a mitochondrial ATPase, since this activ-ity was insensitive to oligomycin and sodium azide(Table I), two known F-type ATPase inhibitors (30). InLeishmania donovani, a H1-ATPase responsible forintracellular pH maintenance has been identified inthe plasma membrane (52). This H1-ATPase was de-

pdp
s
rtS
310 BARROS ET AL.
scribed as a P-type cation transport ATPase, highlysensitive to orthovanadate (53). Since the Mg21-inde-
endent and the Mg21-dependent ecto-ATPases hereescribed did not respond to vanadate (Table I), theossibility that these activities were due to a H1-
TA
Influence of Various Agents on ATPase and p-N
Additions
p-NPPase
Independent of MgCl2 Dependen
Control 100.0 6 9.8 100.0Levamizole (1.0 mM) 100.8 6 12.5 95.2Vanadate (1.0 mM) 5.7 6 0.7 24.1Molybdate (1.0 mM) 9.2 6 1.3 31.3Tartrate (10.0 mM) 90.1 6 10.8 98.1NaF (10.0 mM) 12.6 6 1.7 36.7Ouabain (1.0 mM) 95.8 6 7.4 98.7Azide (10.0 mM) 98.6 6 8.9 93.3Bafilomycin (1 mM) 91.4 6 10.6 93.0Oligomycin (2 mg/ml) 98.1 6 12.6 95.4Adenosine (10.0 mM) 112.6 6 10.8 103.2AMP (10.0 mM) 81.0 6 7.2 79.2ADP (10.0 mM) 62.8 6 7.1 41.5Pi (10.0 mM) 59.3 6 4.8 75.9p-NPP (10.0 mM) —
Note. ATPase and phosphatase activities were measured in the sta5 mM p-NPP. Activities are expressed as a percentage of that mMg21-independent ATPase (78.2 6 5.9 nmol Pi/h/105 cells), the Mindependent p-NPPase (2.8 6 0.2 nmol Pi/h/105 cells), and the Mg21-das 100%. The standard errors were calculated from the absolute acconverted to percentage of the control values. The unpaired t test statistical differences (P . 0.05) from the control.
FIG. 7. Dependence on ATP42 (A) and Mg-ATP22 (B) concentrationwere incubated at 36°C in the same reaction medium (final volume:ATP42 (A) or Mg-ATP22 (B) concentrations varying as shown on theegression using the Michaelis–Menten equation as described undero form the desired ATP42 (A) and Mg-ATP22 (B) concentrations wereE of three determinations with different cell suspensions.
ATPase present on the surface of the plasma membrane(30) was discarded. This ATP hydrolysis cannot be dueto a phosphatase activity present on the external sur-face of Entamoeba membrane because, as shown in Fig.5, the ecto-phosphatase activities present on the exter-
I
ophenylphosphatase Activities of E. histolytica
Relative activity (%)
ATPase
n MgCl2 Independent of MgCl2 Dependent on MgCl2
8.7 100.0 6 7.6 100.0 6 12.412.9 95.3 6 12.4 95.7 6 9.4
3.4 92.8 6 8.6 92.2 6 7.44.2 92.5 6 10.7 92.7 6 12.6
13.8 104.7 6 11.8 104.8 6 11.63.7 81.5 6 9.3 81.0 6 6.9
10.7 100.7 6 12.7 100.8 6 10.58.7 100.4 6 7.9 100.6 6 9.69.7 100.3 6 8.6 100.0 6 11.7
10.4 100.3 6 12.8 100.6 6 7.312.5 94.9 6 10.9 94.4 6 12.96.6 60.4 6 7.2 60.3 6 7.93.2 61.6 6 4.4 58.2 6 6.68.1 98.9 6 7.9 98.8 6 10.1
104.1 6 12.0 99.0 6 8.7
rd assay described under Material and Methods with 5 mM ATP orured under control conditions, i.e., without other additions. The
-dependent ATPase (258.6 6 31.9 nmol Pi/h/105 cells), the Mg21-ndent p-NPPase (1.3 6 0.1 nmol Pi/h/105 cells) activities were takenty values of three experiments with different cell suspensions andwed, in all measurements of ATPase activities, that there were no
ecto-ATPase activities of intact cells of invasive E. histolytica. Cellsml) as that described in the legend of Fig. 2, which corresponds toscissa. Curves represent the fit of experimental data by nonlinearterial and Methods. The total amounts of ATP and MgCl2 necessaryulated as described under Material and Methods. Data are means 6
BLE
itr
t o
66666666666666
—
ndaeasg21
epetiviho
s of0.5ab
Macalc

w
p(aAtwdM
l
ftstc
A
A
a
s
311ECTO-ATPases IN Entamoeba histolytica
nal surface of E. histolytica are acid phosphatases,hereas the optimum pH for the Mg21-dependent ecto-
ATPase lies in the alkaline range and the Mg21-inde-endent activity was not affected by variation of the pHFig. 6). In addition, potent inhibitors for phosphatasectivities were not capable of modifying the ecto-TPase activities (Table I). Similar results were ob-
ained for L. tropica Mg21-dependent ecto-ATPase,hich also exhibits higher activity in alkaline pH andoes not respond to phosphatase inhibitors (30). Theg21-independent and the Mg21-dependent ecto-
ATPase activities described here cannot be attributedto a 59-nucleotidase, since AMP hydrolysis is not cata-yzed by the intact cells (Fig. 1), and ammonium
TABLE II
Substrate Specificity of Ecto-ATPase Activities
Nucleotides
Relative activity
Mg21-independent Mg21-dependent
ATP 100.0 6 7.6 100.0 6 12.4CTP 1.3 6 0.1 4.2 6 0.3GTP 1.0 6 0.1 1.1 6 0.1ITP 9.3 6 0.6 3.2 6 0.4UTP 3.6 6 0.3 1.0 6 0.1ADP 50.6 6 5.7 32.1 6 3.6
Note. ATPase activities were measured in a standard assay de-scribed under Material and Methods with the nucleotides listed (5mM), without or with the addition of 5 mM MgCl2. Reactions were at30°C. The Mg21-independent ATPase (78.2 6 5.9 nmol Pi/h/105 cells)nd the Mg21-dependent ATPase (258.6 6 31.9 nmol Pi/h/105 cells)
activities were taken as 100%. The standard errors were calculatedfrom the absolute activity values of three experiments with differentcell suspensions and converted to percentage of the control values. Inthese experiments, ATP hydrolysis was measured using the samecolorimetric assay of Pi release from other nucleotides as that de-cribed under Material and Methods.
FIG. 8. Effects of DIDS and suramin on the ecto-ATPase activitiesat 36°C in the same reaction medium (final volume: 0.5 ml) as thatDIDS or suramin. After 1 h, the reaction was started by the additioncells were incubated in the same conditions described above in the p(B) the addition of 5 mM MgCl2. Data are means 6 SE of three det
molybdate does not inhibit the ecto-ATPase activities,although it does inhibit the phosphatase activities (Ta-ble I). These data are consistent with the previouslydescribed biochemical and ultrastructural localizationof an ATPase, an ADPase, and an acid phosphataseand exclusion of a 59-nucleotidase on the plasma mem-brane and on the inner surface of the cytoplasmic vacu-oles in E. histolytica (16, 54). Here, we further charac-terize the E. histolytica ecto-ATPase activities, show-ing that the addition of MgCl2 to the extracellularmedium increased the ecto-ATPase activity in a dose-dependent manner (Fig. 4A). Low concentrations ofCa21 and Mn21 also stimulated the ecto-ATPase activ-ity, whereas high concentrations of these cations inhib-ited this activity (Figs. 4C and 4D, respectively). Verylow concentrations of Fe31 (as low as 0.3 mM) were ableto inhibit the Mg21-dependent ATPase but not theMg21-independent ecto-ATPase activity (Fig. 4B). Dif-erences in the response to pH variation observed forhe two ecto-ATPases here described, as well as inensitivity to the inhibition promoted by Fe31, suggesthat these activities are due to different enzymes. Asan be also seen in Figs. 7A and 7B, the Mg21-depen-
dent ATPase activity was higher than the Mg21-inde-pendent ecto-ATPase activity, although with lower af-finity for the corresponding substrates Mg-ATP22 and
TP42, respectively.It has been shown that the nucleoside triphosphate
hydrolase (NTPase) purified from T. gondii is not asingle enzyme but a mixture of two isozymes, termedNTPase I and NTPase II, and that a primary differencebetween these isozymes is that NTPase II hydrolyzesnucleoside triphosphate and diphosphate substrates atalmost the same rate, whereas NTPase I hydrolyzesnucleoside diphosphate at a much slower rate (55).The Mg21-independent and the Mg21-dependent ecto-
TPases of E. histolytica characterized here demon-
ntact cells of invasive E. histolytica. Cells were preincubated for 1 hcribed in the legend of Fig. 2, with or without the addition of 1 mMmM Tris–ATP [g-32P]ATP [sp act 5 104 Bq/nmol ATP]. (Insets) The
ence of 5 mM ATP. The assays were performed without (A) or withinations with different cell suspensions.
of idesof 5res
erm

C(
1p
312 BARROS ET AL.
strate high specificity for ATP, being much less activetoward other nucleoside triphosphate substrates, suchas ITP, CTP, UTP, and GTP (Table II), but are alsoable to hydrolyze ADP (Fig. 1, Table II). The ADPhydrolysis was 32 and 50% of the ATP hydrolysis forthe Mg21-dependent and the Mg21-independent activi-ties, respectively (Table II). Furthermore, ADP inhib-ited the two ATPase activities by about 40% (Table I).These data are highly suggestive that the ATP hydro-lysis characterized here is due to an ATP-diphospho-hydrolase activity, as already described in other organ-isms (9, 56–59). In contrast, in L. tropica, a Mg21-dependent ecto-ATPase that was not considered anATP-diphosphohydrolase because of the lack of inhibi-tion of the ATP hydrolysis by high concentrations ofADP at nonsaturating concentrations of ATP has beencharacterized (30). In T. gondii, it has been shown thatdifferent strains exhibit NTPases from two differentgroups, one group with ATPase/ADPase ratios of 4.0 or5.0 and another group with ATPase/ADPase ratios of
FIG. 9. Ecto-ATPase activities of intact cells of invasive E. histolytiells were incubated for 1 h at 36°C in the same reaction medium (
A) or with (B) the addition of 5 mM MgCl2. Data are means 6 SE
FIG. 10. Effects of D-glucose and D-galactose on the ecto-ATPase ach at 36°C in the same reaction medium (final volume: 0.5 ml) as t
resence of 50 mM glucose (Glu) or 50 mM galactose (Gal) without (Adeterminations with different cell suspensions.
approximately 1.0 (51). This ratio is similar to thoseshown by the ecto-ATPases of aortic and lung endothe-lial cells, erythrocytes, and other blood cells (8).
The identity and function of ecto-ATPases have beenreviewed and the nomenclature “E-type ATPases” wasproposed for these enzymes (9), whose physiologicalrole is still unknown. However, several hypotheseshave been suggested, such as protection from cytolyticeffects of extracellular ATP (11, 12), regulation of ec-tokinase substrate concentration (9), involvement withcellular adhesion and signal transduction (40, 60, 61),and regulation of NO release in smooth muscle cells(62). Considering the fact that E. histolytica is respon-sible for complicated infections, whose ulceration of theintestinal mucosa allows the parasite to penetrate intothe host circulation, one could speculate that the pres-ence of an ecto-ATPase activity might reflect some formof evasion of the parasite from host defense mecha-nisms in the circulation (1, 8). A highly active ecto-ATP-diphosphohydrolase was localized on the external
A), noninvasive E. histolytica (B), and free-living E. moshkovskii (C).l volume: 0.5 ml) as that described in the legend of Fig. 2, without
hree determinations with different cell suspensions.
ties of intact cells of invasive E. histolytica. Cells were incubated fordescribed in the legend of Fig. 2, in the absence (Control) or in thewith (B) the addition of 5 mM MgCl2. Data are means 6 SE of three
ca (finaof t
tivihat) or

As
cr
e
1
313ECTO-ATPases IN Entamoeba histolytica
surface of the tegument of Schistosoma mansoni (49,50) and the authors suggested that this enzyme mightbe involved in an escape mechanism allowing the par-asite to split ATP or ADP released in its vicinity byactivated platelets or activated cytotoxic T lympho-cytes (49, 50).
The results presented in Fig. 9 demonstrate that thepathogenic amoeba E. histolytica has much higherecto-ATPase activities than the noninvasive amoeba E.histolytica or the free-living amoeba E. moshkovskii. Itis well known that coevolution of parasites and hosts isbased upon gain or loss of certain characteristics thatare acted on by natural selection, so that one organismbecomes adapted to live as a parasite of the other,which becomes its host (63). An example is the acqui-sition of cytoplasmic virus-like particles by somestrains of Endotrypanum spp., which has been attrib-uted to the apparent faster rate of evolution and to thegeographic distribution of these trypanosomatid para-sites, as compared to other strains of the same genus(64, 65). Differences in the structure of glycoconjugatesthat are exposed on the parasites’ exterior surfaces arecertainly among the most important acquisitions forintracellular parasitism (66–68). Since ecto-ATPasesare glycoproteins themselves (8, 9), it is possible thatthe differences observed here in the ecto-ATPase activ-ities of the free-living E. moshkovskii, the noninvasiveE. histolytica, and the pathogenic E. histolytica (Fig. 9)were significant in the evolution of these species, aprocess occurring over a long period of time, duringwhich they became so physiologically distinct that theyremain taxonomically separate (2, 3).
In T. gondii, it has been shown that there is nodifference between the ATPase activities of virulentand those of avirulent strains, although it was sug-gested that these activities must be essential for itsintracellular parasitism (51). If ecto-ATPases serve asadhesion molecules in Entamoeba, they could be con-sidered pathogenesis markers for these cells. Surfacemembrane interactions between E. histolytica and itshost are of critical importance, and understanding thebiochemical basis for parasite adherence and cytolyticactivities is crucial for vaccine development. Killing ofhost cells by E. histolytica trophozoites in vitro occursonly upon direct contact, which is mediated by an ame-bic adhesin, a lectin that recognizes N- and O-linkedoligosaccharides (6, 39). This lectin is specifically in-hibited by millimolar concentrations of galactose andN-acetyl-D-galactosamine (6). The essential role of theGal/GalNAc lectin in amebic adherence to target cellshas been demonstrated using carbohydrate inhibitorsand using target cells deficient in glycosylation. Inadherence assays in vitro, 50 mM Gal or GalNAc in-hibited about 90% of the binding of E. histolytica tro-phozoites to Chinese hamster ovary cells, whereasother sugars had no effect (69, 70). When we investi-gated the possible effect of D-galactose on both ecto-
TPases present in E. histolytica, 50 mM D-galactosetimulated the Mg21-dependent ecto-ATPase activity
by more than twofold but did not interfere with theMg21-independent ecto-ATPase activity (Fig. 10). Re-ently, it was shown that a 46-kDa lectin isolated fromoot extracts of the legume Doliclos biflorus is a Nod
factor binding protein as well as a nucleoside di- andtriphosphate hydrolase stimulated by carbohydrate li-gands (71). As it has been suggested that ecto-ATPasesmay be involved with cellular adhesion (41, 60, 61),we intend to investigate whether the amebic lectin(6, 39) may be associated with this Mg21-dependentcto-ATPase.
ACKNOWLEDGMENTS
We thank Dr. Martha M. Sorenson for critical reading of themanuscript. This work was partially supported by grants from theBrazilian agencies Conselho Nacional de Desenvolvimento Cientıficoe Tecnologico (CNPq), Financiadora de Estudos e Projetos (FINEP),Fundacao de Amparo a Pesquisa do Estado do Rio de Janeiro(FAPERJ), and Programa de Nucleos de Excelencia (PRONEX,Grant 0885).
REFERENCES
1. Ravdin, J. (1989) J. Infect. Dis. 159, 420–429.2. Aucott, J. N., and Ravdin, J. I. (1993) Infect. Dis. Clin. North Am.
7, 467–485.3. Jackson, T. F. H. G. (1998) Int. J. Parasitol. 28, 181–186.4. Sharon, N., and Lis, H. (1989) Science 246, 227–234.5. Guerrant, R. L, Brush, J., Ravdin, J. I., Sullivan, J. A., and
Mandell G. L. (1981) J. Infect. Dis. 143, 83–93.6. Ravdin, J. I., and Guerrant, R. L. (1981) J. Clin. Invest. 68,
1305–1313.7. Zimmermann, H. (1996) Drug Dev. Res. 39, 337–352.8. Plesner, L. (1995) Int. Rev. Cytol. 158, 141–214.9. Dombrowski, K., Ke, Y., Brewer, K. A., and Kapp, J. A. (1998)
Immunol. Rev. 161, 111–118.0. Gordon, J. L. (1986) Biochem. J. 233, 309–319.
11. Steinberg, T. H., and Di Virgilio, F. (1991) Curr. Opin. Immunol.3, 71–75.
12. Filippini, A., Taffs, R. E., Agui, T., and Sitkovsky, M. V. (1990)J. Biol. Chem. 265, 334–340.
13. De Pierre, J. W., and Karnovsky, M. L. (1973) J. Cell Biol. 56,275–303.
14. McLaughlin, J., Lindmark, D. G., and Muller, M. (1978) Bio-chem. Biophys. Res. Commun. 82, 913–920.
15. McCaul, T. F., and Bird, R. G. (1978) Int. J. Parasitol. 8, 501–506.
16. Serrano, R., Deas, J. E., and Swarren, L. G. (1977) Exp. Parasi-tol. 41, 370–384.
17. Bakker-Grunwald, T., and Parduhn, H. (1993) Mol. Biochem.Parasitol. 57, 167–170.
18. Boonlayagoor, P., Albach, R. A., Stern, M. L., and Booden, T.(1978) Exp. Parasitol. 45, 225–233.
19. Glynn, I. M., and Chappell, J. B. (1946) Biochem. J. 90, 147–149.20. Sorenson, M. M., Coelho, H. S. L., and Reuben, J. P. (1986) J.
Membrane Biol. 90, 219–230.21. Fabiato, A., and Fabiato, F. (1979) J. Physiol. (Paris) 75, 423–
429.

314 BARROS ET AL.
22. Gomes, M. A., Silva, E. F., Macedo, M. A., Vago, A. R. and Melo,M. N. (1997) Parasitology 114, 517–520.
23. Diamond, L. S., Harlow, D. R., and Cunnick, C. C. (1978) Trans.R. Soc. Trop. Med. Hyg. 72, 431.
24. Guilherme, A., Meyer-Fernandes, J. R., and Vieyra, A. (1991)Biochemistry 90, 147–149.
25. Lowry, H. O., and Lopez, M. (1946) J. Biol. Chem. 162, 421–428.26. Meyer-Fernandes, J. R., and Vieyra, A. (1988) Arch. Biochem.
Biophys. 266, 132–141.27. Smith, T. M., Kirley, T. L., and Hennessey, T. M. (1997) Arch.
Biochem. Biophys. 337, 351–359.28. Garcia-Vescovi, Soncini, F. C., and Groisman, E. A. (1996) Cell
84, 165–174.29. Anaya-Ruiz, M., Rosales-Encina, J. L., and Talamas-Rohama, P.
(1997) Arch. Med. Res. 28, 182–183.30. Meyer-Fernandes, J. R., Dutra, P. M. L., Rodrigues, C. O., Saad-
Nehme, J., and Lopes, A. H. C. S. (1997) Arch. Biochem. Biophys.341, 40–46.
31. Fernandes, E. C., Meyer-Fernandes, J. R., Silva-Neto, M. A. C.,and Vercesi, A. E. (1997) Z. Naturforsch. 52c, 351–358.
32. Dutra, P. M. L., Rodrigues, C. O., Jesus, J. B., Lopes, A. H. C. S.,Souto-Padron, T., and Meyer-Fernandes, J. R. (1998) Biochem.Biophys. Res. Commun. 253, 164–169.
33. Van Belle, H. (1972) Biochim. Biophys. Acta 289, 158–168.34. Van Belle, H. (1976) Clin. Chem. 22, 972–976.35. Gottlieb, M., and Dwyer, D. M. (1983) Mol. Biochem. Parasitol. 7,
303–317.36. Cunningham, H. B., Yazak, P. J., Domingo, R. C., Oades, K. V.,
Bohlen, H., Sabbadini, R. A., and Dahms, A. S. (1993) Arch.Biochem. Biophys. 303, 32–43.
37. Knowles, A. F. (1988) Arch. Biochem. Biophys. 263, 264–271.38. Hourani, S. M. O., and Chown, J. A. (1989) Gen. Pharmacol. 20,
413–416.39. Ravdin, J. I., Croft, B. Y., and Guerrant, R. L. (1980) J. Exp. Med.
152, 377–390.40. Salata, R. A., Pearson, R. P., Murphy, C. F., and Ravdin, J. I.
(1985) J. Clin. Invest. 76, 491–499.41. Stout, J. G., Strobel, R. S., and Kirley, T. L. (1995) J. Biol. Chem.
270, 11845–11850.42. Cheung, P. H., Luo, W., Qui, Y., Zhang, X., Earley, K., Milirons,
P., and Lin, S-H. (1993) J. Biol. Chem. 268, 24303–24310.43. Aurivillus, M., Hansen, O. C., Lazrek, M. B. S., Bock, E., and
Obrink, B. (1990) FEBS Lett. 264, 267–269.44. Chen, B. C., Lee, C-M, and Lin, W-W. (1996) Br. J. Pharmacol.
119, 1628–1634.45. Trams, E. G., and Lauter, C. J. (1974) Biochim. Biophys. Acta
345, 180–197.46. Traverso-Cori, A., Chaimovich, H., and Cori, O. (1965) Arch.
Biochem. Biophys. 109, 173–184.47. Komoszynski, M., and Wojtczak, A. (1996) Biochim. Biophys.
Acta 1310, 233–241.
48. Ribeiro, J. M. C., and Garcia, E. S. (1980) J. Insect Physiol. 26,303–307.
49. Vasconcelos, E. G., Nascimento, P. S., Meirelles, M. N. L., Ver-jovski-Almeida, S., and Ferreira, S. T. (1993) Mol. Biochem.Parasitol. 58, 205–214.
50. Vasconcelos, E. G., Ferreira, S. T., Carvalho, T. M. U., De Souza,W., Kettlun, A. M., Mancilla, M., Valenzuela, M. A., and Verjov-ski-Almeida, S. (1996) J. Biol. Chem. 271, 267–269.
51. Asai, T., and Suzuki, Y. (1990) FEMS Microbiol. Lett. 72, 89–92.52. Zilberstein, D., and Dwyer, D. M. (1985) Proc. Natl. Acad. Sci.
USA 82, 1716–1720.53. Zilberstein, D., and Dwyer, D. M. (1988) Biochem. J. 256, 13–21.54. Kobayashi, S., Takeushi, T., Asami, K., and Fujiwara, T. (1981)
Exp. Parasitol. 54, 202–212.55. Asai, T., Miura, S., Sibley, L. D., Okabayashi, H., and Takeuchi,
T. (1995) J. Biol. Chem. 270, 11391–11397.56. Sarkis, J. J. F., Guimaraes, J. A., and Ribeiro, J. M. C. (1986)
Biochem. J. 233, 885–890.57. Handa, M., and Guidotti, G. (1996) Biochem. Biophys. Res. Com-
mun. 218, 916–923.58. Wang, T-F., and Guidotti, G. (1996) J. Biol. Chem. 271, 9898–
9901.59. Zimmermann, H. (1999) Trends Pharmacol. Sci. 20, 231–236.60. Margolis, R. N., Schell, M. J., Taylor, and Hubbard, A. L. (1990)
Biochem. Biophys. Res. Commun. 166, 562–566.61. Dubyak, G. R., and El-Motassin, C. (1993) Am. J. Physiol. 34,
C577–C606.62. Yagi, K., Nishiro, I., Eguchi, M., Kitagawa, M., Miura, Y., and
Mizoguchi, T. (1994) Biochem. Biophys. Res. Commun. 203,1237–1243.
63. Ridley, M. (1993) in Evolution, pp. 267–301, Blackwell Sci.,Cambridge.
64. Lopes, A. H. C. S., Iovannisci, D., Petrillo-Peixoto, M. L., McMa-hon-Pratt, D., and Beverley, S. M. (1990) Mol. Biochem. Parasi-tol. 49, 151–162.
65. Fernandes, A. P., Nelson, K., and Beverley, S. M. (1993) Proc.Natl. Acad. Sci. USA 90, 11608–11612.
66. Mauel, J. (1996) Adv. Parasitol. 38, 1–51.67. Routier, F. H., Xavier da Silveira, E., Wait, R., Jones, C., Pre-
viato, J. O., and Mendonca-Previato, L. (1995) Mol. Biochem.Parasitol. 69, 81–92.
68. Xavier da Silveira, E., Jones, C., Wait, R., Previato, J. O., andMendonca-Previato, L. (1998) Biochem. J. 329, 665–673.
69. Saffer, L. D., and Petri, W. A., Jr. (1991) Exp. Parasitol. 72,106–108.
70. McCoy, J. J., Mann, B. J., and Petri, W. A., Jr. (1994) Infect.Immun. 62, 3045–3050.
71. Etzler, M. E., Kalsi, G., Ewing, N. N., Roberts, N. J., Day, R. B.,and Murphy, J. B. (1999) Proc. Natl. Acad. Sci. USA 96, 5856–5861.
Copyright © 2022 FDOKUMEN




















