
Differentiation of Entamoeba histolytica: A possible role for enolase

Experimental Parasitology 115 (2007) 192–199
www.elsevier.com/locate/yexpr
A phagocytosis mutant of Entamoeba histolytica is less virulent due to deWcient proteinase expression and release
Ken K. Hirata a, Xuchu Que a, Samuel G. Melendez-Lopez a, Anjan Debnath b, Simona Myers a,1, D. Scott Herdman a, Esther Orozco c, Alok Bhattacharya d,
James H. McKerrow b, Sharon L. Reed a,¤
a Departments of Pathology and Medicine, University of California, San Diego, CA 92103-8416, USAb Sandler Center for Basic Research in Parasitic Diseases, University of California, San Francisco, CA 94158, USA
c Departamento de Patologia Experimental, Centro de Investigacion y de Estudios Avanzados del, I.P.N., Mexico City, Mexico 07000, Mexicod School of Life Sciences, Jawaharlal Nehru University, New Delhi 10067, India
Received 21 April 2006; received in revised form 24 June 2006; accepted 2 August 2006Available online 20 September 2006
Abstract
Cysteine proteinases are key virulence factors of Entamoeba histolytica that are released during the process of invasion. We used achemical mutant of E. histolytica strain HM-1:IMSS, clone L6, which is deWcient in virulence, phagocytosis, and cysteine proteinase activ-ity to help deWne the mechanisms of cysteine proteinase release. All cysteine proteinase genes of wild type HM-1 were present in the L6mutant genome, but three of the major expressed proteinases, ehcp1, ehcp2, and ehcp5 were both transcribed, translated, and released atlower levels in L6. We hypothesized that a central protein such as the calcium binding protein 1, EhCaBP1, which is required for bothphagocytosis and exocytosis might be deWcient in this mutant. We found that both mRNA and proteinase levels of EhCaBP1 weredecreased in L6. These Wndings provide an important link between phagocytosis, passive release of multiple cysteine proteinases, andattenuated virulence of this E. histolytica mutant.© 2006 Elsevier Inc. All rights reserved.
Index Descriptors and Abbreviations: Entamoeba histolytica; Amebae; Parasite; Phagocytosis; Cysteine proteinases
1. Introduction
Cysteine proteinases are key virulence factors of Ent-amoeba histolytica. Studies using native and recombinantproteinases have shown that they play a role in invasionthrough the degradation of extracellular matrix and epithe-lial cell basement membrane (Keene et al., 1986). Cysteineproteinases also have important interactions with the hostimmune system, including activation of complement (Reedet al., 1986) and degradation of the anaphylatoxins C3a andC5a (Reed et al., 1995), IgG (Tran et al., 1998), IgA (Kelsalland Ravdin, 1993), and pro-IL-18 (Que et al., 2003). The
* Corresponding author. Fax: +1 619 543 6614.1 Present address: Department of Obstetrics and Gynecology, University
of California, Davis, Sacramento, CA, USA.
0014-4894/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.exppara.2006.08.004
E. histolytica genome project revealed that »40 genesencode cysteine proteinases (Loftus et al., 2005), but only aminority were expressed in cultured trophozoites (Bruch-haus et al., 2003). A recent study of the transcriptome of E.histolytica trophozoites during intestinal colonizationfound that the main up-regulated cysteine proteinases wereEhCP4 and EhCP6 (Gilchrist et al., 2006), proteinases thatare not expressed in culture (Bruchhaus et al., 2003).
The study of the localization and activity of individualproteinases has been aided by the puriWcation of native andactive, recombinant enzymes (Keene et al., 1986; Jacobset al., 1998; Li et al., 1995; Que et al., 2002). At least threeproteinases, EhCP2 (ACP2), EhCP5, and CP112 localize tothe plasma membrane (Jacobs et al., 1998; Que et al., 2002;Garcia-Rivera et al., 1999), and EhCP3 (ACP1) is cytoplas-mic (Que et al., 2002). Studies of the role of cysteine

K.K. Hirata et al. / Experimental Parasitology 115 (2007) 192–199 193
proteinases in the pathogenesis of other parasites have beengreatly aided by gene knock-outs (reviewed in Rosenthal,1999; Sajid and McKerrow, 2002), but homologous recom-bination has yet to be achieved in E. histolytica. In vivostudies using cysteine proteinase inhibitors (Stanley et al.,1995) and antisense constructs (Ankri et al., 1999) showedthat loss of cysteine protease activity by amebae lead toreduced liver abscess formation in SCID mice and ham-sters. However, cysteine proteinase inhibitors used to dateare active against all of the amebic cathepsins tested (Queet al., 2002), and the EhCP5-antisense construct likelyaVects multiple genes as well (Ankri et al., 1999). The onlystable proteinase deWcient strain of E. histolytica, L6, wasinitially isolated as a phagocytosis deWcient clone (Orozcoet al., 1983), and the basis for its decreased virulence has notbeen completely understood. We now show that thismutant also has decreased passive release of multiple cys-teine proteinases, which is due to lower levels of calciumbinding protein 1 (EhCaBP1), a central protein controllingphagocytosis and exocytosis.
2. Materials and methods
2.1. Entamoeba strains
Entamoeba histolytica strain HM1:IMSS and the cloneL6 (Orozco et al., 1983), were grown axenically in TYI-S-33medium and subcultured every 48–72 h. Monoxenic Ent-amoeba dispar, SAW 1734, was grown in YI supplementedwith Crithidia.
2.2. Cysteine proteinase activity
Cysteine proteinase activity was measured in superna-tants and lysates of trophozoites (5£106/ml) in PBS-Cys++(PBS supplemented with 20 mM cysteine, 0.15 mM CaCl2,and 0.5 mM MgCl2) for 2 h at 37 °C as previously described(Reed et al., 1993). The liberation of the Xuorescent leavinggroup, 7-amino-4-methyl coumarin (AMC) from the pep-tide substrate, benzyloxycarbonyl-Arg-Arg-AMC (EnzymeSystems Products, Livermore, CA) was determined and cal-culated as the initial velocity of cleavage as nM/ml/min(Reed et al., 1993).
2.3. PCR ampliWcation of cysteine proteinase genes
Genomic DNA was puriWed from E. histolytica, L-6 andE. dispar trophozoite nuclei using the Wizard DNA puriW-cation kit (Promega). Oligonucleotide primers were speciW-cally designed to amplify each of the 40 cysteine proteinasegenes (Bruchhaus et al., 2003). Products were visualized byethidium bromide staining of agarose gels.
2.4. Kinetic RT-PCR
Total RNA was extracted from trophozoites using theRNeasy Mini Kit (Qiagen) and cDNA synthesized with
oligo (dT) (Stratagene, La Jolla, CA). PCRs were per-formed in triplicate using SYBR Green PCR CoreReagents (Perkin-Elmer). The PCR product was continu-ally monitored in a Perkin-Elmer Gene Amp 5700Sequence Detection system with Quantitation software,which analyzes the ampliWcation curves derived from theincorporation of SYBR Green into the double strandedDNA product. The relative amount of product generatedwas measured by the threshold cycle (CT), when the level ofspeciWc PCR product increases exponentially and crossesthe threshold of a passive reference dye in each sample.Individual CT values were based on three separate mea-surements. Each experimental mRNA level was normalizedto an equivalent expression level of glyceraldehyde-3-phos-phate dehydrogenase (GenBank Accession No. M89790).The speciWcity of the PCR ampliWcation was veriWed bymelting curves of the Wnal amplicons.
2.5. Microarray hybridizations
Total RNA was isolated from HM-1 and L6 trophozo-ites using TRIZOL (Invitrogen). cDNA synthesis, micro-array hybridization, and microarray scanning wereperformed as described (Debnath et al., 2004). BrieXy,cDNA was synthesized from total RNA extracted fromHM-1:IMSS or L6, coupled to CyScribe Cy3 or Cy5(Amersham), and hybridized to the shotgun DNA micro-array constructed by use of 6144 random inserts from anE. histolytica genomic DNA library (Debnath et al., 2004).Four hybridizations were performed with HM-1 trophozo-ite RNA labeled with Cy3 and L6 trophozoite RNAlabeled with Cy5 (EhCS1p1-095, EhCS1p1-096, EhCS1p1-102, and EhCS1p1-103); one hybridization with HM-1trophozoite RNA labeled with Cy5 and L6 trophozoiteRNA labeled with Cy3 (EhCS1p1-099). Scanning and dataanalysis were performed as previously described (Debnathet al., 2004). Only spots in which almost more than 70% ofthe pixels had a signal of at least twice the standard devia-tion (SD) of the local background were considered in sub-sequent data analysis. Clones whose expression decreasedat least 4-fold in L6 relative to HM-1 were selected for fur-ther sequencing analysis.
2.6. Cysteine proteinase ELISA
To develop an ELISA to quantify the amount of pro-teinases released into media, EhCP1, EhCP2, EhCP3, andEhCP5 were all expressed as recombinant proteins as previ-ously described (Que et al., 2002). Monoclonal antibody toEhCP3 (ACP1) and polyclonal anti-peptide antibody toEhCP2 (ACP2) were made as previously described (Queet al., 2002). Polyclonal antibodies were raised againstrecombinant EhCP1 (expressed in pRSETA, Invitrogen)and EhCP5 (in pBAD/Thio-TOPO, Invitrogen) and puri-Wed by nickel chromatography. Rabbits were injected sub-cutaneously three times with 100 �g of each recombinantproteinase with Titermax (Sigma), and the IgG fraction was

194 K.K. Hirata et al. / Experimental Parasitology 115 (2007) 192–199
puriWed using Protein A chromatography (Pharmacia). Totest for speciWcity of the antibodies, 1.0 �g of each recombi-nant antigen (rEhCP1, 2, 3, and 5) was electrophoresedon 12% gels, transferred to nitrocellulose, reacted with a1–5 �g/ml dilution of each antibody, and detected with goatanti-mouse or anti-rabbit IgG alkaline phosphatase andBCIP/NBT (5-bromo, 4-chloro, 3-indolylphosphate/nitro-blue tetrazolium, Zymed, San Francisco, CA).
To quantify the amount of released proteinases, stan-dard curves were generated with 6.5–50 ng of each recombi-nant protein. Released proteinases were collected as above(2.2), and a 1:100 dilution of the media (50 �l) was used tocoat 96 well plates in duplicate overnight at 4 °C andblocked with PBS-3% bovine serum albumin. Plates wereextensively washed with PBS-0.05% Tween between eachstep and sequentially incubated with an »1.0 �g/ml dilutionof the polyclonal or monoclonal antibodies, followed bygoat anti-mouse or anti-rabbit IgG horseradish peroxidaseconjugates. Plates were developed with peroxidasesubstrate SureBlue, KPL, Gaithesburg, Md), 100 �l Stopsolution was added, and the absorbance read at 450 nm.
2.7. Phagocytosis assay
Phagocytosis was measured by incubating trophozoites(1£ 104) in TY-I-S33 without serum with Xuorescent NileRed Sulfate polystyrene latex beads (2£ 107, InterfacialDynamics, Portland, Ore.) for 30 min at 37 °C. The extracel-lular beads were removed by washing, trophozoites Wxedfor 10 min in 4% formaldehyde at 4 °C, and the number ofphagocytosed beads determined in aliquots of trophozoitesby Xuorescent microscopy.
2.8. EhCaBP1 immunoblots and Xuorescence microscopy
The levels of EhCaBP1 in HM-1 and L6 were comparedin lysates by immunoblots. Freshly harvested Entamoebacells were resuspended at 106/ml in lysis buVer (50 mMTris, 2 mM EDTA, pH 7.4, 0.1% NP-40 with inhibitors(Complete Mini, Roche) plus additional E64 to a Wnal con-centration of 100 �M). The cell lysates were vortexed andfrozen at ¡70 °C. After two freeze thaw cycles and a 15 ssonication, the lysates were centrifuged at 10,000g for 5mins at 4 °C, and the cleared supernatants were assayed forprotein concentration. Equal amounts of the lysates(400 ng) were electrophoresed (4–20% gradient gels;Novex) and transferred to PVDF membranes. Followingblocking with 5% milk/TBS/0.05% Tween 20, the blotswere probed with rabbit anti-EhCaBP1 serum (Sahooet al., 2004) (1:2000 dilution) or monoclonal antibodies tothe peroxiredoxin (FP-10,11, and 19, 0.1 �g/ml Wnal con-centration, Choi et al., 2005), followed by Zymax™ goatanti-rabbit or goat-anti mouse IgG HRP conjugate. Theblots were developed with SuperSignal ChemiluminescentSubstrate (Pierce). After imaging the developed blots, banddensities were determined using Quantity One software(Bio-Rad).
Trophozoites were allowed to attach to chamber slides(Lab Tek, Nunc), phagocytose human erythrocytes for10 min, and Wxed for immunoXuoresence labeling as previ-ously described (Sahoo et al., 2004) with 3.7% warmedparaformaldehyde and permeabilized with 0.1% TritonX-100/PBS. EhCaBP1 polyclonal antibody was used at a1:50 dilution, anti-rabbit Alexa 488 (Molecular Probes) at1:500 and phalloidin (Sigma) at 1:500. Imaging was per-formed on a Nikon Optiphot Xuorescent microscope.
3. Results
3.1. L6 has less total cysteine proteinase activity than E. histolytica or E. dispar
To deWne the defect in L6, we Wrst compared the amountof cysteine proteinase activity in lysates and released inmedia by E. histolytica, L6, and E. dispar. E. histolyticalysates had signiWcantly more cysteine proteinase activitythan L6 or E. dispar (p < 0.001 by Student’s paired t test,Fig. 1). E. histolytica (HM-1:IMSS) also released signiW-cantly more proteinase into the media than E. dispar or L6(p < 0.001 by Student’s paired t test, Fig. 1).
3.2. All cysteine proteinase genes are present in L6
To determine if chemical mutagenesis in L6 resulted infull or partial deletions of cysteine proteinase genes, weampliWed products containing the translated regions of allforty cysteine proteinases. All genes were detected in the L6genome (data not shown).
Fig. 1. L6 has less intracellular and released cysteine proteinase activity.The total cysteine proteinase activity in lysates and released into media byE. histolytica, E. dispar, and L6 was measured by the initial velocity of thecleavage of a Xuorescent peptide substrate (Z-Arg-Arg-AMC). Valuesrepresent the means § SE of 17 determinations.
K.K. Hirata et al. / Experimental Parasitology 115 (2007) 192–199 195
3.3. Cysteine proteinases genes are transcribed at lower levels in L6
We next evaluated the transcription levels of the Wvemajor expressed cysteine proteinase genes using real-time(RT) PCR. We found that all Wve genes were transcribed inL6, but ehcp1, ehcp2, and ehcp5 were all transcribed at 4- to8-fold lower levels than E. histolytica (Table 1). The levelsof EhCP3 and EhCP112 were unchanged.
3.4. Transcriptional diVerences between HM-1 and L6
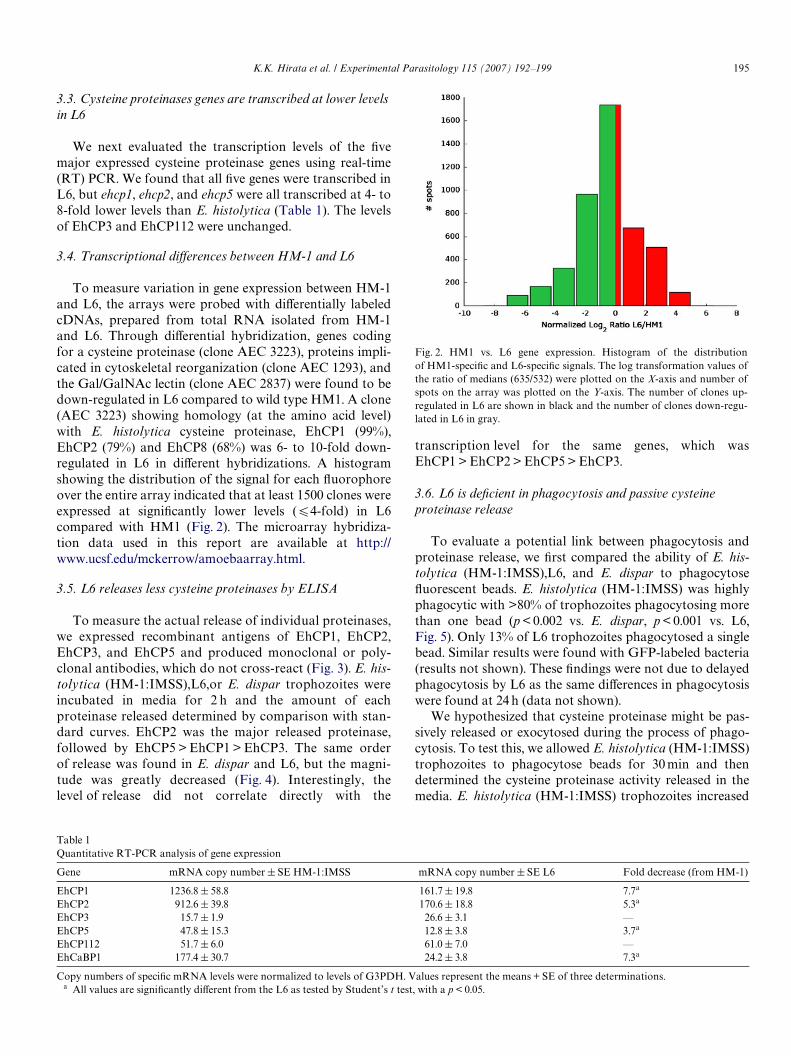
To measure variation in gene expression between HM-1and L6, the arrays were probed with diVerentially labeledcDNAs, prepared from total RNA isolated from HM-1and L6. Through diVerential hybridization, genes codingfor a cysteine proteinase (clone AEC 3223), proteins impli-cated in cytoskeletal reorganization (clone AEC 1293), andthe Gal/GalNAc lectin (clone AEC 2837) were found to bedown-regulated in L6 compared to wild type HM1. A clone(AEC 3223) showing homology (at the amino acid level)with E. histolytica cysteine proteinase, EhCP1 (99%),EhCP2 (79%) and EhCP8 (68%) was 6- to 10-fold down-regulated in L6 in diVerent hybridizations. A histogramshowing the distribution of the signal for each Xuorophoreover the entire array indicated that at least 1500 clones wereexpressed at signiWcantly lower levels (64-fold) in L6compared with HM1 (Fig. 2). The microarray hybridiza-tion data used in this report are available at http://www.ucsf.edu/mckerrow/amoebaarray.html.
3.5. L6 releases less cysteine proteinases by ELISA
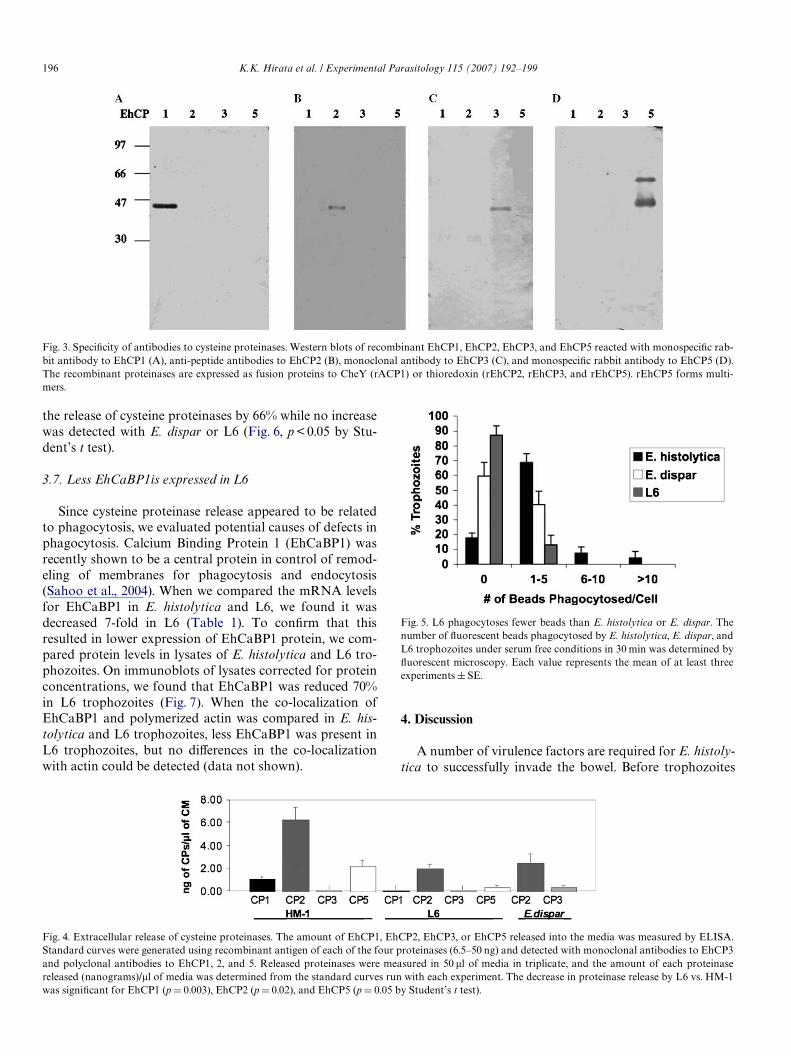
To measure the actual release of individual proteinases,we expressed recombinant antigens of EhCP1, EhCP2,EhCP3, and EhCP5 and produced monoclonal or poly-clonal antibodies, which do not cross-react (Fig. 3). E. his-tolytica (HM-1:IMSS),L6,or E. dispar trophozoites wereincubated in media for 2 h and the amount of eachproteinase released determined by comparison with stan-dard curves. EhCP2 was the major released proteinase,followed by EhCP5 > EhCP1 > EhCP3. The same orderof release was found in E. dispar and L6, but the magni-tude was greatly decreased (Fig. 4). Interestingly, thelevel of release did not correlate directly with the
transcription level for the same genes, which wasEhCP1 > EhCP2 > EhCP5 > EhCP3.
3.6. L6 is deWcient in phagocytosis and passive cysteine proteinase release
To evaluate a potential link between phagocytosis andproteinase release, we Wrst compared the ability of E. his-tolytica (HM-1:IMSS),L6, and E. dispar to phagocytoseXuorescent beads. E. histolytica (HM-1:IMSS) was highlyphagocytic with >80% of trophozoites phagocytosing morethan one bead (p < 0.002 vs. E. dispar, p < 0.001 vs. L6,Fig. 5). Only 13% of L6 trophozoites phagocytosed a singlebead. Similar results were found with GFP-labeled bacteria(results not shown). These Wndings were not due to delayedphagocytosis by L6 as the same diVerences in phagocytosiswere found at 24 h (data not shown).
We hypothesized that cysteine proteinase might be pas-sively released or exocytosed during the process of phago-cytosis. To test this, we allowed E. histolytica (HM-1:IMSS)trophozoites to phagocytose beads for 30 min and thendetermined the cysteine proteinase activity released in themedia. E. histolytica (HM-1:IMSS) trophozoites increased
Fig. 2. HM1 vs. L6 gene expression. Histogram of the distributionof HM1-speciWc and L6-speciWc signals. The log transformation values ofthe ratio of medians (635/532) were plotted on the X-axis and number ofspots on the array was plotted on the Y-axis. The number of clones up-regulated in L6 are shown in black and the number of clones down-regu-lated in L6 in gray.
Table 1Quantitative RT-PCR analysis of gene expression
Copy numbers of speciWc mRNA levels were normalized to levels of G3PDH. Values represent the means + SE of three determinations.a All values are signiWcantly diVerent from the L6 as tested by Student’s t test, with a p < 0.05.
Gene mRNA copy number § SE HM-1:IMSS mRNA copy number § SE L6 Fold decrease (from HM-1)
EhCP1 1236.8 § 58.8 161.7§ 19.8 7.7a
EhCP2 912.6 § 39.8 170.6§ 18.8 5.3a
EhCP3 15.7 § 1.9 26.6 § 3.1 —EhCP5 47.8 § 15.3 12.8 § 3.8 3.7a
EhCP112 51.7 § 6.0 61.0 § 7.0 —EhCaBP1 177.4 § 30.7 24.2 § 3.8 7.3a
196 K.K. Hirata et al. / Experimental Parasitology 115 (2007) 192–199
the release of cysteine proteinases by 66% while no increasewas detected with E. dispar or L6 (Fig. 6, p < 0.05 by Stu-dent’s t test).
3.7. Less EhCaBP1is expressed in L6
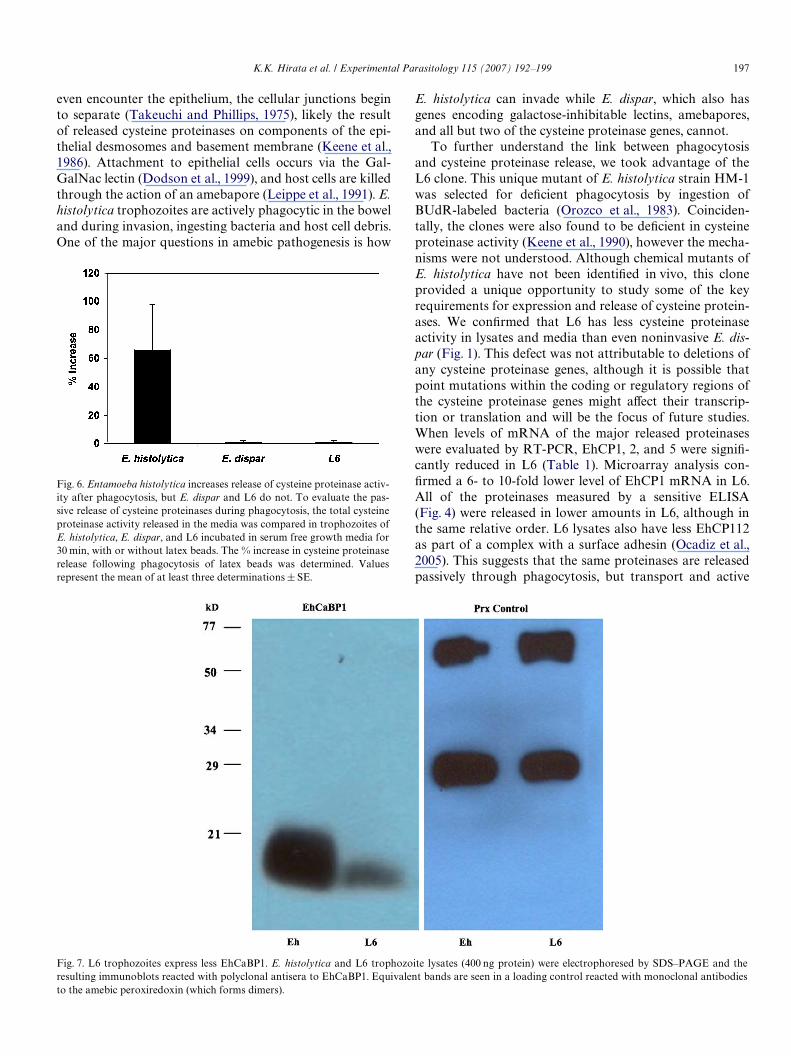
Since cysteine proteinase release appeared to be relatedto phagocytosis, we evaluated potential causes of defects inphagocytosis. Calcium Binding Protein 1 (EhCaBP1) wasrecently shown to be a central protein in control of remod-eling of membranes for phagocytosis and endocytosis(Sahoo et al., 2004). When we compared the mRNA levelsfor EhCaBP1 in E. histolytica and L6, we found it wasdecreased 7-fold in L6 (Table 1). To conWrm that thisresulted in lower expression of EhCaBP1 protein, we com-pared protein levels in lysates of E. histolytica and L6 tro-phozoites. On immunoblots of lysates corrected for proteinconcentrations, we found that EhCaBP1 was reduced 70%in L6 trophozoites (Fig. 7). When the co-localization ofEhCaBP1 and polymerized actin was compared in E. his-tolytica and L6 trophozoites, less EhCaBP1 was present inL6 trophozoites, but no diVerences in the co-localizationwith actin could be detected (data not shown).
4. Discussion
A number of virulence factors are required for E. histoly-tica to successfully invade the bowel. Before trophozoites
Fig. 5. L6 phagocytoses fewer beads than E. histolytica or E. dispar. Thenumber of Xuorescent beads phagocytosed by E. histolytica, E. dispar, andL6 trophozoites under serum free conditions in 30 min was determined byXuorescent microscopy. Each value represents the mean of at least threeexperiments § SE.
Fig. 4. Extracellular release of cysteine proteinases. The amount of EhCP1, EhCP2, EhCP3, or EhCP5 released into the media was measured by ELISA.Standard curves were generated using recombinant antigen of each of the four proteinases (6.5–50 ng) and detected with monoclonal antibodies to EhCP3and polyclonal antibodies to EhCP1, 2, and 5. Released proteinases were measured in 50 �l of media in triplicate, and the amount of each proteinasereleased (nanograms)/�l of media was determined from the standard curves run with each experiment. The decrease in proteinase release by L6 vs. HM-1
was signiWcant for EhCP1 (pD 0.003), EhCP2 (p D 0.02), and EhCP5 (p D 0.05 by Student’s t test).Fig. 3. SpeciWcity of antibodies to cysteine proteinases. Western blots of recombinant EhCP1, EhCP2, EhCP3, and EhCP5 reacted with monospeciWc rab-bit antibody to EhCP1 (A), anti-peptide antibodies to EhCP2 (B), monoclonal antibody to EhCP3 (C), and monospeciWc rabbit antibody to EhCP5 (D).The recombinant proteinases are expressed as fusion proteins to CheY (rACP1) or thioredoxin (rEhCP2, rEhCP3, and rEhCP5). rEhCP5 forms multi-mers.
K.K. Hirata et al. / Experimental Parasitology 115 (2007) 192–199 197
even encounter the epithelium, the cellular junctions beginto separate (Takeuchi and Phillips, 1975), likely the resultof released cysteine proteinases on components of the epi-thelial desmosomes and basement membrane (Keene et al.,1986). Attachment to epithelial cells occurs via the Gal-GalNac lectin (Dodson et al., 1999), and host cells are killedthrough the action of an amebapore (Leippe et al., 1991). E.histolytica trophozoites are actively phagocytic in the boweland during invasion, ingesting bacteria and host cell debris.One of the major questions in amebic pathogenesis is how
Fig. 6. Entamoeba histolytica increases release of cysteine proteinase activ-ity after phagocytosis, but E. dispar and L6 do not. To evaluate the pas-sive release of cysteine proteinases during phagocytosis, the total cysteineproteinase activity released in the media was compared in trophozoites ofE. histolytica, E. dispar, and L6 incubated in serum free growth media for30 min, with or without latex beads. The % increase in cysteine proteinaserelease following phagocytosis of latex beads was determined. Valuesrepresent the mean of at least three determinations § SE.
E. histolytica can invade while E. dispar, which also hasgenes encoding galactose-inhibitable lectins, amebapores,and all but two of the cysteine proteinase genes, cannot.
To further understand the link between phagocytosisand cysteine proteinase release, we took advantage of theL6 clone. This unique mutant of E. histolytica strain HM-1was selected for deWcient phagocytosis by ingestion ofBUdR-labeled bacteria (Orozco et al., 1983). Coinciden-tally, the clones were also found to be deWcient in cysteineproteinase activity (Keene et al., 1990), however the mecha-nisms were not understood. Although chemical mutants ofE. histolytica have not been identiWed in vivo, this cloneprovided a unique opportunity to study some of the keyrequirements for expression and release of cysteine protein-ases. We conWrmed that L6 has less cysteine proteinaseactivity in lysates and media than even noninvasive E. dis-par (Fig. 1). This defect was not attributable to deletions ofany cysteine proteinase genes, although it is possible thatpoint mutations within the coding or regulatory regions ofthe cysteine proteinase genes might aVect their transcrip-tion or translation and will be the focus of future studies.When levels of mRNA of the major released proteinaseswere evaluated by RT-PCR, EhCP1, 2, and 5 were signiW-cantly reduced in L6 (Table 1). Microarray analysis con-Wrmed a 6- to 10-fold lower level of EhCP1 mRNA in L6.All of the proteinases measured by a sensitive ELISA(Fig. 4) were released in lower amounts in L6, although inthe same relative order. L6 lysates also have less EhCP112as part of a complex with a surface adhesin (Ocadiz et al.,2005). This suggests that the same proteinases are releasedpassively through phagocytosis, but transport and active
Fig. 7. L6 trophozoites express less EhCaBP1. E. histolytica and L6 trophozoite lysates (400 ng protein) were electrophoresed by SDS–PAGE and theresulting immunoblots reacted with polyclonal antisera to EhCaBP1. Equivalent bands are seen in a loading control reacted with monoclonal antibodies
to the amebic peroxiredoxin (which forms dimers).
198 K.K. Hirata et al. / Experimental Parasitology 115 (2007) 192–199
release of the cysteine proteinases may diVer, an area we areactively investigating.
L6 trophozoites had signiWcantly reduced phagocytosis,even less than noninvasive E. dispar (Fig. 5). Cysteine pro-teinase activity might be co-regulated in some fashion withthe process of phagocytosis, either involving passive releaseor exocytosis (see below) (Fig. 6). There was no increasedrelease of cysteine proteinases into the media with thepoorly phagocytic E. dispar or L6, while E. histolyticareleased 66% more cysteine proteinase activity.
Phagocytosis is an active process in E. histolytica,involving actin polymerization controlled by a number ofproteins. Overexpression of the mutant p21racA-V12, whichforces GTPase to be Wxed in the active state, caused accu-mulation of actin under the plasma membrane and mark-edly decreased phagocytosis (Ghosh and Samuelson,1997). The Rab proteins are also important regulatorsof vesicular transport, including cysteine proteinases(Temesvari et al., 1999; Saito-Nakano et al., 2004; Nak-ada-Tsukui et al., 2005). RabB was overexpressed in L6,but the complex remained in cytoplasmic vesicles and wasineYciently translocated to phagocytic vesicles (Guzman-Medrano et al., 2005). Both EhRab5 and Rab7 have beenlinked with the transport of cysteine proteinases via aprephagosomal vacuole, but the pathways controllingspeciWc cysteine proteinases remain to be determined(Saito-Nakano et al., 2004; Nakada-Tsukui et al., 2005;Okada et al., 2005).
Because both phagocytosis and the passive release ofmultiple cysteine proteinases were disrupted in this clone,we hypothesized that a central control protein wasinvolved. A potential candidate is EhCaBP1, which co-localizes with polymerized F-actin and inhibition by anti-sense expression impaired both processes (Sahoo et al.,2004).We found that less EhCaBP1 mRNA expression(Table 1) and less EhCaBP1 protein were detected in L6(Fig. 7). Calcium Xuxes have recently been linked withmembrane repair and lysosomal exocytosis, manifest asgenetic defects in beige-J mice and the human recessivedisorder, Chediak-Higashi syndrome (Huynh et al.,2004). The disruption of calcium homeostasis mediatedby EhCaBP1 in the L6 mutant lends further support tothis link in a lower eukaryote, leading to the defectivephagocytosis, proteinase release, and virulence in thismutant.
Acknowledgments
This work was presented in part at the XIV Seminar onAmebiasis, Mexico City, November 27–30, 2000 (AbstractS237). The work was supported by grants from the UnitedStates Public Health Service Grant RO1AI49531 (S.R.),DK-35108 (S.R.), and AI35707 (J.M.), and the Sandler Cen-ter for Basic Research in Parasitic Diseases. S.G.M.L was arecipient of a PROMEP-UABC Scholarship (UABC-123,Clave P/PROMEP: UABC-2000-12-01; Ministry of PublicEducation, México.
References
Ankri, S., Stolarsky, T., Bracha, R., Padilla-Vaca, F., Mirelman, D., 1999.Antisense inhibition of expression of cysteine proteinases aVects Ent-amoeba histolytica- induced formation of liver abscess in hamsters.Infection and Immunity 67, 421–422.
Bruchhaus, I., Loftus, B.J., Hall, N., Tannich, E., 2003. The intestinal pro-tozoan parasite Entamoeba histolytica contains 20 cysteine proteasegenes, of which only a small subset is expressed during in vitro cultiva-tion. Eukaryotic Cell 2, 501–509.
Choi, M.H., Sajed, D., Poole, L., Hirata, K., Herdman, S., Torian, B.E.,Reed, S.L., 2005. An unusual surface peroxiredoxin protects invasiveEntamoeba histolytica from oxidant attack. Molecular and Biochemi-cal Parasitology 143, 80–89.
Debnath, A., Das, P., Sajid, M., McKerrow, J.H., 2004. IdentiWcation ofgenomic responses to collagen binding by trophozoites of Entamoebahistolytica. Journal of Infectious Diseases 190, 448–457.
Dodson, J.M., Lenkowski, P., Eubanks, A.C., Jackson, G.H., Napodano, J.,Lyerly, D.M., Lockhard, L.A., Mann, B.J., Petri, W.A., 1999. Infectionand immunity mediated by the carbohydrate recognition domain ofthe Entamoeba histolytica Gal/GalNac lectin. Journal of InfectiousDiseases 179, 460–466.
Garcia-Rivera, G., Rodriguez, M.A., Ocadiz, R., Martinez-Lopez, M.C.,Arroyo, R., Gonzalez-Robles, A., Orozco, E., 1999. Entamoeba histoly-tica: a novel cysteine protease and an adhesin form the 112 kDa surfaceprotein. Molecular Microbiology 33, 556–568.
Ghosh, S.K., Samuelson, J., 1997. Involvement of p21racA, phosphoinosi-tide 3-kinase, and vacuolar ATPase in phagocytosis of bacteria anderythrocytes by Entamoeba histolytica: suggestive evidence for coinci-dental evolution of amebic invasiveness. Infection and Immunity 65,4243–4249.
Gilchrist, C.A., Houpt, E., Trapaidze, N., Fei, Z., Crasta, O., Asgharpour,A., Evans, C., Martino-Catt, S., Baba, D.J., Stroup, S., Hamano, S.,Ehrenkaufer, G., Okada, M., Singh, U., Nozaki, T., Mann, B.J., PetriJr., W.A., 2006. Impact of intestinal colonization and invasion on theEntamoeba histolytica transcriptome. Molecular and Biochemical Par-asitology (Epub).
Guzman-Medrano, R., Castillo-Juarez, B.A., Garcia-Perez, R.M., Salas-Cases, A., Orozco, E., Rodriguez, M.A., 2005. Entamoeba histolytica:Alterations in EhRabB protein in a phagocytosis deWcient mutant cor-relate with the Entamoeba dispar RabB sequence. Experimental Parasi-tology 110, 259–264.
Huynh, C.W., Diane, M., Kaplan, J., Andrews, N.W., 2004. Defective lyso-somal exocytosis and plasma membrane repair in Chediak-Higashi/beige cells. Proceedings of the National Academy of Sciences of theUnited States of America 101, 16795–16800.
Jacobs, T., Bruchhaus, I., Dandekar, T., Tannich, E., Leippe, M., 1998. Iso-lation and molecular characterization of a surface-bound proteinase ofEntamoeba histolytica. Molecular Microbiology 27, 269–276.
Keene, W.E., Hidalgo, M.E., Orozco, E., McKerrow, J.H., 1990. Entamoebahistolytica: correlation of the cytopathic eVect of virulent trophozoiteswith secretion of a cysteine proteinase. Experimental Parasitology 71,199–206.
Keene, W.E., Pettit, M.G., Allen, S., McKerrow, J.H., 1986. The major neu-tral proteinase of Entamoeba histolytica. Journal of Experimental Med-icine 163, 536–549.
Kelsall, B.L., Ravdin, J.I., 1993. Degradation of human immunoglobulin Aby Entamoeba histolytica. Journal of Infectious Diseases 168, 1319–1322.
Leippe, M., Sebastian, E., Schoenberger, O.L., Horstmann, R.D., Muller-Eberhard, H.J., 1991. Pore-forming peptide of pathogenic Entamoebahistolytica. Proceedings of the National Academy of Sciences of theUnited States of America 88, 7659–7663.
Li, E., Yang, W.-G., Zhang, T., Stanley, S.L., 1995. Interaction of lamininwith Entamoeba histolytica cysteine proteinases and its eVect on amebicpathogenesis. Infection and Immunity 63, 4150–4153.
Loftus, B., Anderson, I., Davies, R., Alsmark, U.C.M., Samuelson, J., Ame-deo, P., Roncaglia, P., Berriman, M., Hirt, R.P., Mann, B.J., Nozaki, T.,

K.K. Hirata et al. / Experimental Parasitology 115 (2007) 192–199 199
Suh, B., Pop, M., Duchene, M., Ackers, J., Tannich, E., Leippe, M.,Hofer, M., Bruchhaus, Iris., Willhoeft, U., Bhattacharya, A., Chilling-worth, T., Churcher, C., Hance, Z., Harris, B., Harris, D., Jagels, K.,Moule, S., Mungall, K., Ormond, D., Squares, R., Whitehead, S., Quail,M.A., Rabbinowitsch, E., Norbertczak, H., Price, C., Wang, Z., Guillén,N., Gilchrist, C., Stroup, S.E., Bhattacharya, S., Lohia, A., Foster, P.G.,Sicheritz-Ponten, T., Weber, C., Singh, U., Mukherjee, C., El-Sayed,N.M., Petri Jr., W.A., Clark, C.G., Embley, T.M., Barrell, B., Fraser,C.M., Hall, N., 2005. The genome of the protist parasite Entamoebahistolytica. Nature 433, 865–868.
Nakada-Tsukui, K., Saito-Nakano, Y., Ali, V., Nozaki, T., 2005. A retro-merlike complex is a novel Rab7 eVector that is involved in thetransport of the virulence factor cysteine protease in the enteric proto-zoan parasite Entamoeba histolytica. Molecular Biology of the Cell 16,5294–5303.
Ocadiz, R., Orozco, E., Carrillo, E., Quintas, L.I., Ortega-Lopez, J., Garcia-Perez, R.M., Sanchez, T., Castillo-Juarez, B.A., Garcia-Rivera, G.,Rodriguez, M.A., 2005. EhCP112 is an Entamoeba histolytica secretedcysteine protease that may be involved in the parasite-virulence. Cellu-lar Microbiology 7, 221–232.
Okada, M., Huston, C.D., Mann, B.J., Petri, W.A., Kita, K., Nozaki,T., 2005. Proteomic analysis of phagocytosis in the enteric protozoanparasite Entamoeba histolytica. Eukaryotic Cell 4, 827–831.
Orozco, E., Guarneros, G., Martinez-Palomo, A., Sanchez, T., 1983.Entamoeba histolytica phagocytosis as a virulence factor. Journal ofExperimental Medicine 158, 1511–1521.
Que, X., Brinen, L.S., Perkins, P., Herdman, D.S., Hirata, K., Torian, B.E.,Rubin, H.M., Reed, S.L., 2002. Cysteine proteinases from distinct cellu-lar compartments are recruited to phagocytic vesicles by Entamoebahistolytica. Molecular Biochemistry and Parasitology 119, 23–32.
Que, X., Kim, S.-H., Sajid, M., Eckmann, L., Dinarello, C.A., McKerrow,J.H., Reed, S.L., 2003. A surface amebic cysteine proteinase inactivatesInterleukin-18. Infection and Immunity 71, 1274–1280.
Reed, S.L., Bouvier, J., Pollack, A.S., Engel, C., Brown, M., Hirata, K., Que,X., Eakin, A., Hagblom, P., Gillin, F.D., McKerrow, J.H., 1993. Cloningof a virulence factor of Entamoeba histolytica: pathogenic strains
possess a unique cysteine proteinase gene. Journal of Clinical Investi-gation 91, 1532–1540.
Reed, S.L., Curd, J.G., Gigli, I., Gillin, F.D., Braude, A.I., 1986. Activationof complement by pathogenic and nonpathogenic Entamoeba histoly-tica. Journal of Immunology 136, 2265–2270.
Reed, S.L., Ember, J.A., Herdman, D.S., DiScipio, R.G., Hugli, T.E., Gigli,I., 1995. The extracellular neutral cysteine proteinase of Entamoeba his-tolytica degrades anaphylatoxins C3a and C5a. Journal of Immunol-ogy 155, 266–274.
Rosenthal, P.J., 1999. Proteases of protozoan parasites. Advances inParasitology 43, 106–139.
Sahoo, N., Labruyere, E., Bhattacharya, S., Sen, P., Guillen, N., Bhattach-arya, A., 2004. Calcium binding protein 1 of the protozoan parasiteEntamoeba histolytica interacts with actin and is involved in cytoskele-ton dynamics. Journal of Cell Science 117, 3625–3634.
Saito-Nakano, Y., Yasuda, T., Nakada-Tsukui, K., Lieppe, M., Nozaki, T.,2004. Rab5-associated vacuoles play a unique role in phagocytosis ofthe enteric protozoan parasite Entamoeba histolytica. Journal ofBiological Chemistry 279, 49497–49507.
Sajid, M., McKerrow, J.H., 2002. Cysteine proteases of parasitic organ-isms. Molecular Biochemistry and Parasitology 120, 1–21.
Stanley, S.L., Zhang, T., Rubin, D., Li, E., 1995. Role of the Entamoeba his-tolytica cysteine proteinase in amebic liver abscess formation in severecombined immunodeWcient mice. Infection and Immunity 63, 1587–1589.
Takeuchi, A., Phillips, B.P., 1975. Electron microscope studies of experi-mental Entamoeba histolytica infections in the guinea pig. I. Penetra-tion of the intestinal epithelium by trophozoites. American Journal ofTropical Medicine and Hygiene 24, 34–48.
Temesvari, L.A., Harris, E.N., Stanley Jr., S.L., Cardelli, J.A., 1999. Earlyand late endosomal compartments of Entamoeba histolytica areenriched in cysteine proteases, acid phosphatase and several Ras-related Rab GTPases. Molecular and Biochemical Parasitology 103,225–241.
Tran, V.Q., Herdman, D.S., Torian, B.E., Reed, S.L., 1998. The neutral cys-teine proteinase of Entamoeba histolytica degrades IgG and preventsits binding. Journal of Infectious Diseases 177, 508–511.
Copyright © 2022 FDOKUMEN




















