
Inhibition of Obliterative Airway Disease Development in Murine Tracheal Allografts by Matrix...
13
American Journal of Transplantation 2005; 5: 671–683 Blackwell Munksgaard Copyright C Blackwell Munksgaard 2005 doi: 10.1111/j.1600-6143.2005.00751.x Inhibition of Obliterative Airway Disease Development in Murine Tracheal Allografts by Matrix Metalloproteinase-9 Deficiency F´ elix G. Fern ´ andez a , Lacey G. Campbell a , Wei Liu a , J. Michael Shipley b,c , Shigeyoshi Itohara d , G. Alexander Patterson a , Robert M. Senior b,c , T. Mohanakumar a,e and Andr ´ es Jaramillo a, ∗ Departments of a Surgery, b Internal Medicine, c Cell Biology & Physiology and e Pathology & Immunology, Washington University School of Medicine, Saint Louis, MO 63110 and d Laboratory for Behavioral Genetics, Brain Science Institute, Riken, Saitama 351-0198, Japan ∗ Corresponding author: Andr ´ es Jaramillo, [email protected]. This study was designed to define the roles of matrix metalloproteinase (MMP)-2 and MMP-9 in obliterative airway disease (OAD) in heterotopic murine tracheal allografts, considered a suitable animal model for chronic lung allograft rejection. BALB/c tracheal allo- grafts were transplanted into MMP-2-deficient (−/−) and MMP-9−/− mice. Also, wild-type recipients were treated with doxycycline, a nonspecific MMP inhibitor. After 10, 20 and 30 days, allografts were analyzed for OAD development, intragraft levels of MMP-2 and MMP-9 and the frequency and cytokine/chemokine production profile of alloreactive T cells. Allografts transplanted into wild-type mice developed OAD lesions within 30 days. These allografts revealed sig- nificant upregulation of both MMP-2 and MMP-9. Allografts transplanted into MMP-9−/− and doxycycline-treated recipients did not develop OAD. In contrast, allografts transplanted into MMP-2−/− mice developed OAD lesions with normal kinetics. Interest- ingly, MMP-9−/− recipients showed an enhanced T cell alloreactivity associated with an abnormal profile of cytokine/chemokine production. The enhanced T cell alloreactivity in MMP-9−/− mice was mediated by enhanced dendritic cell stimulatory capacity as well as enhanced T cell responsive capacity. These results suggest that MMP-9 plays an important role in the pathogenesis of OAD and may represent a target for the therapeutic intervention of chronic lung allograft rejection. Key words: Bronchiolitis obliterans syndrome, cy- tokines, MMP-2, MMP-9, obliterative airway disease, T cells Received 28 April 2004; revised 28 October 2004; ac- cepted for publication 1 November 2004 Introduction Lung transplantation is recognized as a viable treatment option for a variety of end-stage pulmonary diseases. Long- term survival of lung transplant patients is limited by the development of bronchiolitis obliterans syndrome (BOS), which is considered to be the manifestation of chronic lung allograft rejection (1,2). The histopathologic correlate of BOS, obliterative bronchiolitis, is characterized histolog- ically by extensive fibrosis and obliteration of the small air- ways that results in a progressive decline of the pulmonary function (3). A growing body of evidence suggests that BOS is mediated by the coordinated infiltration and effec- tor functions of alloreactive T cells (4,5). In this regard, it has become clear that novel immune mediator molecules such as matrix metalloproteinases (MMP), in particular MMP-2 and MMP-9, may also play an important role in the patho- genesis of BOS (6–9). Currently, conventional immunosup- pressive protocols are the only available treatment for BOS. However, it is clear that these therapeutic protocols are not sufficient to prevent the progression of the disease. There- fore, there is a critical need for novel therapeutic protocols for the prevention and treatment of BOS. Interfering with the infiltration of activated T cells into the allograft, as may be achieved with MMP inhibition, has been shown to be an effective approach for preventing the rejection process in experimental models (10). MMPs constitute a multigene family of over 25 secreted and cell surface enzymes capable of proteolytic activ- ity against many components of the extracellular matrix (11,12). Knowledge about the role of MMPs in the immune response is rapidly expanding, yet it still is not understood how these enzymes are involved in the regulation of the im- mune response during homeostasis and disease (6,11,12). In this regard, CD4+ T cells produce both MMP-2 and MMP-9 while CD8+ T cells predominantly produce MMP- 9 upon antigenic stimulation (13–17). Additionally, T cells induce production of MMP-2 and MMP-9 by macrophages, endothelial cells, epithelial cells, vascular smooth muscle cells and fibroblasts (18–24). It has been shown that MMP- 9 plays a crucial role in the development of autoimmune en- cephalomyelitis, experimental bullous pemphigoid, exper- imental myasthenia gravis and experimental arthritis (25– 28). In this regard, MMP-9 plays an important role in the transmigration of T cells and dendritic cells (DC) through 671
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Inhibition of Obliterative Airway Disease Development in Murine Tracheal Allografts by Matrix...

American Journal of Transplantation 2005; 5: 671–683Blackwell Munksgaard
Copyright C© Blackwell Munksgaard 2005
doi: 10.1111/j.1600-6143.2005.00751.x
Inhibition of Obliterative Airway Disease Developmentin Murine Tracheal Allografts by MatrixMetalloproteinase-9 Deficiency
Felix G. Fernandeza, Lacey G. Campbella, Wei
Liua, J. Michael Shipleyb,c, Shigeyoshi Itoharad,
G. Alexander Pattersona, Robert M. Seniorb,c,
T. Mohanakumara,e and Andres Jaramilloa,∗
Departments of aSurgery, bInternal Medicine, cCellBiology & Physiology and ePathology & Immunology,Washington University School of Medicine, Saint Louis,MO 63110 and dLaboratory for Behavioral Genetics, BrainScience Institute, Riken, Saitama 351-0198, Japan∗Corresponding author: Andres Jaramillo,[email protected].
This study was designed to define the roles of matrixmetalloproteinase (MMP)-2 and MMP-9 in obliterativeairway disease (OAD) in heterotopic murine trachealallografts, considered a suitable animal model forchronic lung allograft rejection. BALB/c tracheal allo-grafts were transplanted into MMP-2-deficient (−/−)and MMP-9−/− mice. Also, wild-type recipients weretreated with doxycycline, a nonspecific MMP inhibitor.After 10, 20 and 30 days, allografts were analyzed forOAD development, intragraft levels of MMP-2 andMMP-9 and the frequency and cytokine/chemokineproduction profile of alloreactive T cells. Allograftstransplanted into wild-type mice developed OADlesions within 30 days. These allografts revealed sig-nificant upregulation of both MMP-2 and MMP-9.Allografts transplanted into MMP-9−/− anddoxycycline-treated recipients did not develop OAD. Incontrast, allografts transplanted into MMP-2−/− micedeveloped OAD lesions with normal kinetics. Interest-ingly, MMP-9−/− recipients showed an enhanced Tcell alloreactivity associated with an abnormal profileof cytokine/chemokine production. The enhanced Tcell alloreactivity in MMP-9−/− mice was mediatedby enhanced dendritic cell stimulatory capacity as wellas enhanced T cell responsive capacity. These resultssuggest that MMP-9 plays an important role in thepathogenesis of OAD and may represent a target forthe therapeutic intervention of chronic lung allograftrejection.
Key words: Bronchiolitis obliterans syndrome, cy-tokines, MMP-2, MMP-9, obliterative airway disease,T cells
Received 28 April 2004; revised 28 October 2004; ac-cepted for publication 1 November 2004
Introduction
Lung transplantation is recognized as a viable treatmentoption for a variety of end-stage pulmonary diseases. Long-term survival of lung transplant patients is limited by thedevelopment of bronchiolitis obliterans syndrome (BOS),which is considered to be the manifestation of chroniclung allograft rejection (1,2). The histopathologic correlateof BOS, obliterative bronchiolitis, is characterized histolog-ically by extensive fibrosis and obliteration of the small air-ways that results in a progressive decline of the pulmonaryfunction (3). A growing body of evidence suggests thatBOS is mediated by the coordinated infiltration and effec-tor functions of alloreactive T cells (4,5). In this regard, it hasbecome clear that novel immune mediator molecules suchas matrix metalloproteinases (MMP), in particular MMP-2and MMP-9, may also play an important role in the patho-genesis of BOS (6–9). Currently, conventional immunosup-pressive protocols are the only available treatment for BOS.However, it is clear that these therapeutic protocols are notsufficient to prevent the progression of the disease. There-fore, there is a critical need for novel therapeutic protocolsfor the prevention and treatment of BOS. Interfering withthe infiltration of activated T cells into the allograft, as maybe achieved with MMP inhibition, has been shown to bean effective approach for preventing the rejection processin experimental models (10).
MMPs constitute a multigene family of over 25 secretedand cell surface enzymes capable of proteolytic activ-ity against many components of the extracellular matrix(11,12). Knowledge about the role of MMPs in the immuneresponse is rapidly expanding, yet it still is not understoodhow these enzymes are involved in the regulation of the im-mune response during homeostasis and disease (6,11,12).In this regard, CD4+ T cells produce both MMP-2 andMMP-9 while CD8+ T cells predominantly produce MMP-9 upon antigenic stimulation (13–17). Additionally, T cellsinduce production of MMP-2 and MMP-9 by macrophages,endothelial cells, epithelial cells, vascular smooth musclecells and fibroblasts (18–24). It has been shown that MMP-9 plays a crucial role in the development of autoimmune en-cephalomyelitis, experimental bullous pemphigoid, exper-imental myasthenia gravis and experimental arthritis (25–28). In this regard, MMP-9 plays an important role in thetransmigration of T cells and dendritic cells (DC) through
671

Fernandez et al.
endothelial and epithelial tight junctions (21,29). In addi-tion, MMP-9 has been shown to be involved in the modu-lation of the activity of several cytokines and chemokines(6,12,30).
Hertz et al. (31) developed a murine heterotopic trachealtransplantation model to study the pathogenesis of BOS.These tracheal allografts developed obliterative airway dis-ease (OAD) within 30 days, which is characterized by aprogression of mononuclear cellular infiltration, epithelialdamage and subepithelial and luminal fibrosis. Since OADreproduces the histopathology of BOS, this is currentlyconsidered the best animal model for the study of theimmunopathogenesis of BOS. Thus, the objective of thisstudy was to characterize the role of MMP-2 and MMP-9 inthe immunopathogenesis of OAD toward the potential useof these enzymes as therapeutic targets for the preventionof BOS after lung transplantation.
Materials and Methods
Animals
MMP-2-deficient (−/−) and MMP-9−/− mice were originally developedby homologous recombination on C57BL/6 (H-2b) and 129SvEv (H-2b) ge-netic backgrounds, respectively (32,33). Wild-type C57BL/6 (MMP-2+/+)and BALB/c (H-2d) mice were obtained from The Jackson Laboratories (BarHarbor, ME). Wild-type 129/SvEv (MMP-9+/+) mice were obtained fromTaconic Farms (Germantown, NY). All mice were housed in a pathogen-freefacility and all experimental procedures were performed in adult males 8–12 weeks of age. All animal procedures were performed according to theguidelines of the animal care committee of Washington University Schoolof Medicine.
Heterotopic tracheal transplantation
Recipient mice were transplanted as previously described (34,35). Briefly,donor mice were anesthetized and exsanguinated. The tracheas were ex-posed via an anterior midline incision, excised and flushed with 2 mL ofice-cold Euro-Collins solution (Mediatech, Herndon, VA). Then, the trachealgrafts were then placed in a subcutaneous pocket on the back of recipientmice anesthetized with intraperitoneal ketamine (50 mg/kg) and xylazine(10 mg/kg). Then, the wound was closed with staples. The tracheal graftswere harvested at 10, 20 and 30 days for histopathologic analysis, cytokineanalysis substrate zymography and RT-PCR.
Inhibition of MMP enzymatic activity in vivo
Tracheal allograft recipients were injected s.c. with doxycycline(20 mg/kg/day, Sigma, St. Louis, MO), a potent inhibitor of MMP activity(36), diluted in PBS starting at day 0 until tracheal grafts were harvested.Control mice were given carrier alone for the duration of the study. Allograftrecipients were sacrificed at the time of rejection.
Histological analysis
Tracheal grafts were fixed in 10% paraformaldehyde, embedded in paraf-fin, sectioned at 5 micron thickness and stained with H&E and Masson’strichrome. The specimens were examined in blinded fashion for evidenceof OAD lesions. Grafts that displayed epithelial abnormalities, mononuclearcellular infiltration and fibroproliferative obliteration of the airway lumenwere considered positive for OAD development. Grafts were considered
normal if there were no epithelial abnormalities, mononuclear cellular infil-tration and fibroproliferation.
Immunohistochemical analysis
Allografts were frozen, embedded in TBS Tissue Freezing Medium (Trian-gle Biomedical Sciences, Durham, NC), and sectioned at 5 lm thickness.The sections were treated with 3% H2O2 for 10 min to quench endoge-nous peroxidase activity and blocked for 30 min with PBS supplementedwith 0.5% blocking reagent (TSA Biotin System, PerkinElmer, Boston, MA)and 2% normal goat serum (Jackson ImmunoResearch Laboratories, WestGrove, PA). Then, the sections were incubated for 2 h with purified rat anti-mouse CD4, CD8, CD19 and CD11b mAbs (5.0 lg/mL, BD Pharmingen, SanDiego, CA). Then, the sections were incubated for 1 h with biotin-conjugatedF(ab′)2 goat anti-rat IgG (1:500, Jackson ImmunoResearch Laboratories).Then, the sections were incubated for 30 min with HRP-conjugated strep-tavidin (BD Pharmingen). The presence of graft-infiltrating CD4+ and CD8+T cells, CD19+ B cells and CD11b+ macrophages was detected with theDAB Substrate Kit (BD Pharmingen), counter-stained with hematoxylin andexamined using a light microscope. The number of graft-infiltrating cellswas determined as the mean of CD4+ cells, CD8+ cells, CD19+ cells andCD11b+ cells present in four separate fields of view evaluated in each graft(magnification: ×400).
Substrate zymography
Tracheal grafts were harvested, homogenized in PBS containing 0.1% BRIJ-35 (Sigma) and a protease inhibitor cocktail (Roche Diagnostics, Mannheim,Germany), and centrifuged at 15,000 rpm for 10 min at 4
◦C. The protein con-
centration of the supernatants was determined using the BCA protein as-say (Pierce, Rockford, IL), and 15 lg of supernatant proteins were resolvedby nonreducing 10% SDS-PAGE through Novex Tris-Glycine gels contain-ing 0.1% gelatin (Invitrogen, Carlsbad, CA). The gels were then washedfor 30 min in Novex zymogram renaturing buffer (Invitrogen) to remove theSDS, and then incubated in Novex zymogram developing buffer (Invitrogen)for 24 h at 37
◦C. Areas of protease activity were visualized after staining
the gels with 0.5% Coomassie blue R-250.
RT-PCR
Tracheal grafts were harvested, homogenized with TRIzol reagent (LifeTechnologies, Gaithersburg, MD), and total RNA was isolated accordingto the manufacturer’s instructions. cDNA was synthesized by extensionof primers using Powerscript reverse transcriptase (Clontech, Palo Alto,CA) according to manufacturer’s instructions. PCR was then performedusing Taq DNA polymerase and buffer mix (Invitrogen) in the presenceof oligonucleotide primers for MMP-2, MMP-9 and GAPDH (control).MMP-2 primer sequences were: 5′-CT T TGCAGGAGACAAGT TCTGG-3′ and 5′-T TAAGGTGGTGCAGGTATCTGG-3′ (701-bp product); MMP-9primer sequences were: 5′-TGGCAGAGGCATACT TGTAC-3′ and 5′-AACTGGATGACAATGTCCGC-3′ (375-bp product); GAPDH primersequences were: 5′-CCATGT T TGTGATGGGTGTGAACC-3′ and 5′-TGTGAGGGAGATGCTCAGTGT TGG-3′ (712-bp product). The MMP-9primer sequences were designed to amplify a specific sequence withinthe region of the gene disrupted in the MMP-9−/− mice. Thus, no MMP-9mRNA is detected in mice carrying this targeted mutation. The PCR cyclingconditions for MMP-2 were as follows: initial denaturation at 94
◦C for
5 min, followed by 40 cycles of 94◦C for 20 s, 51
◦C for 30 s and 72
◦C
for 1 min, and a final extension step at 72◦C for 10 min. The PCR cycling
conditions for MMP-9 were as follows: initial denaturation at 94◦C for
5 min, followed by 40 cycles of 94◦C for 20 s, 55
◦C for 30 s and 72
◦C
for 1 min, and a final extension step at 72◦C for 10 min. The PCR cycling
conditions for GAPDH were as follows: initial denaturation at 94◦C for
1 min, followed by 30 cycles of 94◦C for 20 s, 58
◦C for 1 min and 72
◦C for
1 min, and a final extension set at 72◦C for 10 min. The amplified cDNA
672 American Journal of Transplantation 2005; 5: 671–683

MMPs in Obliterative Airway Disease
products were electrophoresed on 2% agarose gels, stained with ethidiumbromide and visualized using an UV trans-illuminator.
Cytokine/chemokine production assay
To compare the cytokine/chemokine production ex vivo, spleen cells wereharvested from MMP-9+/+ and MMP-9−/− tracheal allograft recipients at20 days after transplantation. The cells were then treated with erythro-cyte lysis buffer (0.1 mM EDTA, 155 mM ammonium chloride, 10 mMpotassium bicarbonate) for 3 min, washed (3×), and resuspended in RPMI-1640 medium (Gibco, Grand Island, NY) supplemented with 10% FBS(Hyclone, Logan, UT), 2 mM L-glutamine, 100 lM nonessential amino acids,25 mM HEPES buffer, 1 mM sodium pyruvate, 100 U/mL penicillin, 100lg/mL streptomycin and 50 lM 2-ME. The cells were plated in 96-well flat-bottom plates in triplicate wells at a concentration of 3 × 105/well. Spleencells from BALB/c (allograft donor) and 129/SvEv (syngeneic control) micewere prepared as described above, irradiated at 3000 rad, and plated in thewells containing responder cells at a concentration of 6 × 105/well. After72 h of incubation, the culture supernatants were collected and analyzed bymeans the TranSignal cytokine/chemokine Ab array (Panomics, RedwoodCity, CA) following the manufacturer’s instructions. To determine the cy-tokine/chemokine production in situ, BALB/c allografts were harvested fromMMP-9+/+ and MMP-9−/− tracheal allograft recipients at 20 days aftertransplantation. The grafts were then treated with lysis buffer (Panomics),diluted to a concentration of 75 lg of protein/mL and analyzed by meansof the TranSignal cytokine/chemokine Ab array (Panomics) following themanufacturer’s instructions. The cytokine blots were analyzed using theQuantity One software (BioRad, Hercules, CA). Results are expressed asdensitometry units.
ELISPOT assay
Priming of alloantigen-specific T cells from tracheal allograft recipients wastested by enumerating IFN-c -producing T cells using an ELISPOT assay.Briefly, MultiScreen 96-well filtration plates (Millipore, Bedford, MA) werecoated for 2 h at 37◦C with a capture IFN-c -specific mAb (5.0 lg/mL, BDPharmingen) in 0.05 M carbonate-bicarbonate buffer (pH: 9.6). Then, theplates were blocked with 1% BSA for 1 h at 37◦C and washed with PBS(3×). Spleen cells from MMP-9+/+ and MMP-9−/− recipients were har-vested at 20 and 30 days after transplantation. Then, the cells were pre-pared as described above and plated at a concentration of 3 × 105/wellin quadruplicate wells in the mAb-coated plates. Spleen cell from BALB/c(allograft donor) and 129/SvEv (syngeneic control) mice were prepared asdescribed above, irradiated at 3000 rad, and plated in the wells containingresponder cells at a concentration of 6 × 105/well. After 48 h of incuba-tion, the plates were washed with PBS (3×) and PBS supplemented with0.5% Tween-20 (3×). Then, a biotinylated IFN-c -specific mAb (2.0 lg/mL,BD Pharmingen) diluted in PBS/BSA/Tween-20 was added to the wells.After 2 h at room temperature, the plates were washed with PBS/Tween-20 (3×) and HRP-labeled streptavidin (1:100, BD Pharmingen) diluted inPBS/BSA/Tween-20 was added to the wells. After 2 h at room temperature,the plates were washed with PBS/Tween-20 (3×) and PBS (3×). Then, AECsubstrate reagent (BD Pharmingen) was added to the wells for 2–5 min.The plates were washed with tap water to stop the reaction and air-dried.Spots were analyzed in an ImmunoSpot Series I analyzer (Cellular Technol-ogy, Cleveland, OH) that was designed to detect spots with pre-determinedcriteria for spot size, shape and colorimetric density.
DC preparation
Mature DCs were purified as previously described (37). Briefly, spleenswere injected with 100 units/mL of collagenase D (Boehringer Mannheim,Indianapolis, IN), teased and passed through a cell strainer with a poresize of 40 lm. The remaining tissue fragments were then treated with400 units/mL of collagenase D for 45–60 min at 37
◦C. After this incubation,
the spleen fragments were passed trough a cell strainer with a pore size of
40 lm and pooled with the single cell suspension collected before. Then,the cells were centrifuged, incubated with anti-mouse CD11c-coated mag-netic beads (Miltenyi Biotech, Auburn, CA), and positively selected witha MACS separation column following the manufacturer’s instructions. Iso-lated DCs were then allowed to undergo maturation by cultivation in culturemedium for 48 h. Purified DCs were >98% CD11c+ as determined by flowcytometric analysis (data not shown).
T cell preparation
Spleens were teased and passed through a cell strainer with a pore size of 40lm. Then, the T cells were negatively selected using the pan-T cell isolationkit (Miltenyi Biotech) following the manufacturer’s instructions. Purified Tcells were >98% CD3+ as determined by flow cytometric analysis (datanot shown).
APC stimulatory capacity assay
To determine the direct stimulatory capacity of MMP-9+/+ versus MMP-9−/− alloantigen-presenting cells, T cells from BALB/c mice were platedin 96-well flat-bottom plates at a concentration of 3 × 105/well in tripli-cate wells, and used as responder cells. Then, varying numbers (1.5–24 ×103/well) of DCs from naıve MMP-9+/+ and MMP-9−/− mice were irra-diated at 3000 rad and used as stimulators. After 72 h of incubation, theremaining cultures were pulsed with 1 lCi/well [3H]-TdR (ICN, Costa Mesa,CA) for 24 h and the incorporation of [3H]-TdR into DNA was determinedby liquid scintillation counting. BALB/c T cells cultured in the presence ofsyngeneic BALB/c DCs were used as negative controls. The mean countsper minute (cpm) in the negative control cultures was subtracted from themean cpm in the experimental cultures to give a final result as net cpm.
In turn, to determine the direct alloresponsive capacity of MMP-9+/+ versusMMP-9−/− T cells, varying numbers (1.5–24 × 103/well) of irradiated DCsfrom BALB/c mice were cultured in the presence of T cells from naive MMP-9+/+ and MMP-9−/− mice. Then, the proliferative response of alloreactiveT cells was determined as described above. MMP-9+/+ and MMP-9−/− Tcells cultured in the presence of syngeneic 129/SvEv were used as negativecontrols. The mean cpm in the negative control cultures was subtractedfrom the mean cpm in the experimental cultures to give a final result as netcpm.
Statistical analysis
Differences in alloreactive T cell frequencies in tracheal allograft recipi-ents and the levels of cellular infiltration into the allografts were assessedby means of the 2-tailed Student’s t-test. Differences in T cell responsesagainst allogeneic DCs in naıve mice were assessed by means of the 2-wayANOVA test. Differences in the frequency of OAD lesions in tracheal graftswere assessed by means of the Fisher exact test. All statistical analyseswere performed using the Prism 4.0 Software (GraphPad, San Diego, CA).The a was set at p < 0.05 in all analyses.
Results
Increased MMP-2 and MMP-9 expression during
OAD development
Previous studies have shown that BALB/c allografts displaycomplete development of OAD at 30 days after transplan-tation (31,34,35). As shown in our previous studies (35),BALB/c and C57BL/6 tracheal grafts were transplanted intoC57BL/6 recipients and harvested at 10, 20 and 30 daysafter transplantation. As shown in Figure 1 and Table 1,C57BL/6 isografts did not show any histopathologic signsof OAD development at any time point (Figure 1A–C). Incontrast, BALB/c allografts showed epithelial metaplasia
American Journal of Transplantation 2005; 5: 671–683 673

Fernandez et al.
Figure 1: Inhibition of OAD development by treatment with doxycycline. C57BL/6 isografts (A–C) and BALB/c allografts (D–F)were transplanted into C57BL/6 mice. In parallel experiments, BALB/c allografts were transplanted into doxycycline-treated C57BL/6recipients (G–I). The grafts were harvested at 10, 20 and 30 days after transplantation. Then, histological analysis was performed with H&E(A,B,D,E,G,H) and Masson’s trichrome (C,F,I) stains. Results shown are representative of five different experiments. Original magnification:×100, inserts: ×400.
and severe mononuclear cellular infiltration at day 10 af-ter transplantation (Figure 1D), severe mononuclear cellularinfiltration, loss of epithelium and partial lumen occlusion(25%) in two of eight specimens at day 20 after transplan-tation (Figure 1E), and moderate mononuclear infiltrationas well as complete luminal occlusion in all specimens atday 30 after transplantation (Figure 1F).
To determine whether MMP-2 and/or MMP-9 play a rolein the immunopathogenesis of OAD, we determined thelevels of MMP-2 and MMP-9 mRNA in BALB/c allograftsand C57BL/6 isografts harvested at 10, 20 and 30 daysafter transplantation.
As shown in Figure 2, C57BL/6 isografts revealed small de-tectable levels of MMP-2 mRNA transcription after 10 daysand no MMP-9 transcription at any time point. In con-trast, BALB/c allografts revealed a significant upregulationof MMP-2 and MMP-9 mRNA transcription after 10 and 20
days but not at 30 days after transplantation. To determinethe expression of MMP-2 and MMP-9 at the protein level,we measured the enzymatic activity of MMP-2 and MMP-9 in BALB/c allografts and C57BL/6 isografts harvested atthe same time points after transplantation. As shown inFigure 3, C57BL/6 isografts displayed very low levels ofpro-MMP-2 or pro-MMP-9 enzymatic activity (gelatinolyticbands at approximately 105 kDa and 64 kDa) and no activeforms of MMP-2 or MMP-9 (gelatinolytic bands at approx-imately 94 kDa and 60 kDa). In contrast, rejected BALB/callografts displayed two prominent gelatinolytic bands cor-responding to the pro-MMP-9 and pro-MMP-2 zymogensafter 10 and 20 days and two less prominent gelatinolyticbands corresponding to the active MMP-9 and MMP-2forms zymogens after 20 days. Only high levels of the pro-and active forms of MMP-2 were detected after 30 days.The gelatinolytic activities were blocked by the addition ofEDTA, which inhibit MMP activity, but not by the serineprotease inhibitor PMSF (data not shown).
674 American Journal of Transplantation 2005; 5: 671–683

MMPs in Obliterative Airway Disease
Table 1: Development of OAD in BALB/c tracheal allografts
OAD lesions (percentage of grafts affected)
Time after Epithelial Mononuclear cellular LumenDonor Recipient Tx (days) Treatment n damage infiltration occlusiona
C57BL/6 C57BL/6 10 – 8 0 0 0C57BL/6 C57BL/6 20 – 8 0 0 0C57BL/6 C57BL/6 30 – 8 0 0 0
BALB/c C57BL/6 10 – 8 100b 100d 0BALB/c C57BL/6 20 – 8 100c 100d 8BALB/c C57BL/6 30 – 8 100c 100e 100
BALB/c C57BL/6 10 Doxycycline 5 0 0 0BALB/c C57BL/6 20 Doxycycline 5 0 20f 0BALB/c C57BL/6 30 Doxycycline 5 40b 80f 0
BALB/c MMP2−/− (C57BL/6) 20 – 8 87c 87d 4BALB/c MMP2−/− (C57BL/6) 30 – 8 87c 87e 92
129/SvEv 129/SvEv 10 – 8 0 0 0129/SvEv 129/SvEv 20 – 8 0 0 0129/SvEv 129/SvEv 30 – 8 0 0 0
BALB/c 129/SvEv 10 – 8 100b 100d 0BALB/c 129/SvEv 20 – 8 100b 100d 0BALB/c 129/SvEv 30 – 8 100c 100c 100
BALB/c MMP9−/− (129/SvEv) 20 – 8 25b 25f 0BALB/c MMP9−/− (129/SvEv) 30 – 8 50b 50f 8aLumen occlusion: results represent the mean percentage of total and partial levels of occlusion.Epithelial damage: bmetaplasia, cabsence.Mononuclear cellular infiltration: dsevere, emoderate and fmild.
Figure 2: Increased MMP-2 and MMP-9
mRNA expression during OAD develop-
ment. C57BL/6 isografts and BALB/c al-lografts were transplanted into C57BL/6mice. In parallel experiments, BALB/c allo-grafts were transplanted into doxycycline-treated C57BL/6 recipients. The graftswere harvested at 10, 20 and 30 days aftertransplantation. Then, the MMP-2, MMP-9 and GAPDH (control) mRNA levels weremeasured by RT-PCR. Results shown arerepresentative of three different experi-ments.
Inhibition of OAD development by abrogation of
MMP enzymatic activity
To further determine whether MMPs play a role in thepathogenesis of OAD, BALB/c allografts were transplantedinto C57BL/6 recipients systemically treated with doxycy-cline (36). As shown in Figure 1 and Table 1, inhibitionof MMP activity significantly delayed the development ofOAD. Doxycycline prevented the epithelial damage and the
mononuclear cellular infiltration observed in these BALB/callografts at days 10 and 20 after transplantation (Figure1G and H, respectively). At day 30 after transplantation,these allografts revealed mild mononuclear cellular infiltra-tion, mild epithelial metaplasia and no luminal occlusion inany of the five specimens studied (Figure 1I). The inhibi-tion of OAD development in the presence of doxycyclinecorrelated with a complete inhibition of MMP-9 mRNA
American Journal of Transplantation 2005; 5: 671–683 675

Fernandez et al.
Figure 3: Increased MMP-2 and MMP-9 enzymatic activity during OAD development. C57BL/6 isografts and BALB/c allografts weretransplanted into C57BL/6 mice. In parallel experiments, BALB/c allografts were transplanted into doxycycline-treated C57BL/6 recipients.The grafts were harvested at 10, 20 and 30 days after transplantation. Then, the MMP-2 and MMP-9 enzymatic activity was measuredby zymographic analysis. Results shown are representative of three different experiments.
transcription and enzymatic activity at days 10 and 20 af-ter transplantation (Figures 2 and 3, respectively). Interest-ingly, only a partial inhibition of MMP-2 mRNA transcriptionand enzymatic activity was observed at days 10 and 20 af-ter transplantation (Figures 2 and 3, respectively). It is im-portant to note that the MMP-2 and MMP-9 mRNA levelsobserved at day 30 after transplantation from doxycycline-treated recipients correlates with the mononuclear cellularinfiltration observed in these allografts at this time point.These data indicate that MMP-2 and/or MMP-9 are in-volved in the process of mononuclear cellular infiltration,and ultimately, OAD development.
Inhibition of OAD development in MMP-9−/− but not
in MMP-2−/− mice
To determine the role of MMP-2 in the pathogenesis ofOAD, BALB/c allografts were transplanted into MMP-2+/+and MMP-2−/− mice. As shown in Figure 4 and Table 1,75% of the BALB/c allografts transplanted into MMP-2−/− mice displayed severe mononuclear cellular infiltra-tion and loss of epithelium as well as partial luminal oc-clusion (25%) in one of eight specimens at day 20 aftertransplantation (Figure 4A). In addition, 87% of the BALB/callografts transplanted into MMP-2−/− mice displayedmoderate mononuclear cellular infiltration and completedestruction of the epithelium at 30 days after transplan-tation (Figure 4B). Seven of eight specimens displayed to-tal luminal occlusion (100%) and one of eight specimensshowed partial luminal occlusion (50%) at day 30 aftertransplantation (Figure 4B). As mentioned above, BALB/callografts transplanted into MMP-2+/+ mice displayed se-vere mononuclear cellular infiltration and epithelial meta-
plasia at day 20 after transplantation (Figure 4C), and mod-erate mononuclear cellular infiltration, loss of epitheliumand luminal obliteration by fibrosis at day 30 after trans-plantation (Figure 4D). Thus, the kinetics of OAD develop-ment in allografts transplanted into MMP-2−/− recipientswas comparable to the one observed in MMP-2+/+ mice.
To determine the role of MMP-9 in the pathogenesis ofOAD, BALB/c allografts were transplanted into MMP-9+/+and MMP-9−/− mice. As shown in Figure 4 and Table 1,only 25% of the BALB/c allografts transplanted into MMP-9−/− mice showed mild mononuclear cellular infiltrationand epithelial metaplasia at day 20 after transplantation(Figure 4E). In addition, only 50% of the BALB/c allo-grafts transplanted into MMP-9−/− mice displayed mildmononuclear cellular infiltration and epithelial metaplasiaat day 30 after transplantation (Figure 4F). Moreover, onlyone of eight specimens displayed partial luminal occlusion(50%) at day 30 after transplantation. In contrast, BALB/callografts transplanted into MMP-9+/+ mice displayed se-vere mononuclear cellular infiltration and epithelial meta-plasia at day 20 after transplantation (Figure 4G), and se-vere mononuclear cellular infiltration, loss of epitheliumand complete luminal occlusion (100%) in all specimensat day 30 after transplantation (Figure 4H). 129/SvEv iso-grafts transplanted into 129/SvEv mice did not show anyhistopathologic signs of OAD development at 10, 20 and 30days after transplantation (Table 1, histology not shown).
Additional morphometric analysis was performed in theseallografts to determine the levels cellular infiltration forT cells (CD4+ and CD8+), B cells (CD19+) andmacrophages (CD11b+) at 20 days after transplantation.
676 American Journal of Transplantation 2005; 5: 671–683
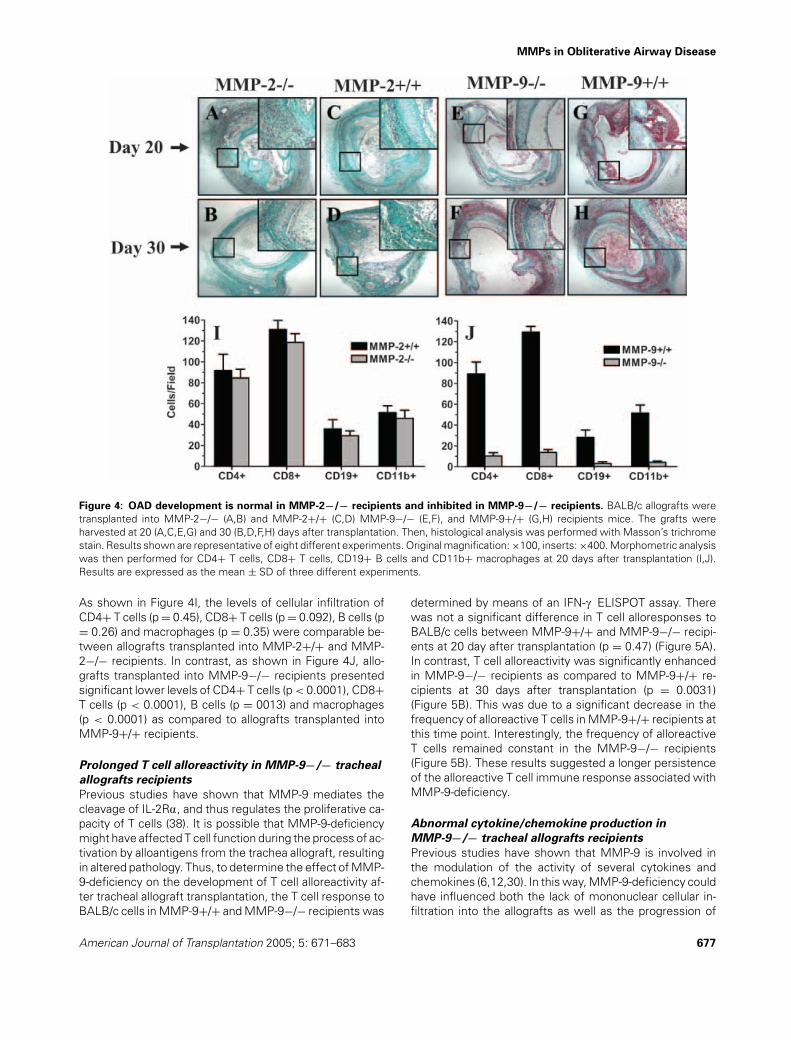
MMPs in Obliterative Airway Disease
Figure 4: OAD development is normal in MMP-2−/− recipients and inhibited in MMP-9−/− recipients. BALB/c allografts weretransplanted into MMP-2−/− (A,B) and MMP-2+/+ (C,D) MMP-9−/− (E,F), and MMP-9+/+ (G,H) recipients mice. The grafts wereharvested at 20 (A,C,E,G) and 30 (B,D,F,H) days after transplantation. Then, histological analysis was performed with Masson’s trichromestain. Results shown are representative of eight different experiments. Original magnification: ×100, inserts: ×400. Morphometric analysiswas then performed for CD4+ T cells, CD8+ T cells, CD19+ B cells and CD11b+ macrophages at 20 days after transplantation (I,J).Results are expressed as the mean ± SD of three different experiments.
As shown in Figure 4I, the levels of cellular infiltration ofCD4+ T cells (p = 0.45), CD8+ T cells (p = 0.092), B cells (p= 0.26) and macrophages (p = 0.35) were comparable be-tween allografts transplanted into MMP-2+/+ and MMP-2−/− recipients. In contrast, as shown in Figure 4J, allo-grafts transplanted into MMP-9−/− recipients presentedsignificant lower levels of CD4+ T cells (p < 0.0001), CD8+T cells (p < 0.0001), B cells (p = 0013) and macrophages(p < 0.0001) as compared to allografts transplanted intoMMP-9+/+ recipients.
Prolonged T cell alloreactivity in MMP-9−/− tracheal
allografts recipients
Previous studies have shown that MMP-9 mediates thecleavage of IL-2Ra, and thus regulates the proliferative ca-pacity of T cells (38). It is possible that MMP-9-deficiencymight have affected T cell function during the process of ac-tivation by alloantigens from the trachea allograft, resultingin altered pathology. Thus, to determine the effect of MMP-9-deficiency on the development of T cell alloreactivity af-ter tracheal allograft transplantation, the T cell response toBALB/c cells in MMP-9+/+ and MMP-9−/− recipients was
determined by means of an IFN-c ELISPOT assay. Therewas not a significant difference in T cell alloresponses toBALB/c cells between MMP-9+/+ and MMP-9−/− recipi-ents at 20 day after transplantation (p = 0.47) (Figure 5A).In contrast, T cell alloreactivity was significantly enhancedin MMP-9−/− recipients as compared to MMP-9+/+ re-cipients at 30 days after transplantation (p = 0.0031)(Figure 5B). This was due to a significant decrease in thefrequency of alloreactive T cells in MMP-9+/+ recipients atthis time point. Interestingly, the frequency of alloreactiveT cells remained constant in the MMP-9−/− recipients(Figure 5B). These results suggested a longer persistenceof the alloreactive T cell immune response associated withMMP-9-deficiency.
Abnormal cytokine/chemokine production in
MMP-9−/− tracheal allografts recipients
Previous studies have shown that MMP-9 is involved inthe modulation of the activity of several cytokines andchemokines (6,12,30). In this way, MMP-9-deficiency couldhave influenced both the lack of mononuclear cellular in-filtration into the allografts as well as the progression of
American Journal of Transplantation 2005; 5: 671–683 677

Fernandez et al.
MMP-9-/- MMP-9+/+0
150
300
450
600
750
900
BALB/c (Donor)
129/SvEv (Self)
Time: 20 days
Sp
ots
/1x1
06 S
ple
en c
ells
MMP-9-/- MMP-9+/+0
150
300
450
600
750
900
BALB/c (Donor)129/SvEv (Self)
Time: 30 days
Sp
ots
/1x1
06 S
ple
en c
ells
A
B
Figure 5: Prolonged T cell alloreactivity in MMP-9−/− tra-
cheal allografts recipients. BALB/c allografts were transplantedinto MMP-9+/+ and MMP-9−/− mice. Then, spleen cells wereharvested at 20 (A) and 30 (B) days after transplantation. Sub-sequently, spleen cells from the MMP-9+/+ and MMP-9−/− re-cipients were cultured in the presence of irradiated BALB/c and129/SvEv (syngeneic control) cells. After 48 h, the frequency of al-loreactive T cells was determined by means of an IFN-c ELISPOTassay. The results are expressed as the mean ± SD of quadru-plicate cultures and are representative of three different experi-ments. T cell response MMP-9+/+ versus MMP-9−/− at 20 and30 days: p = 0.47 and 0.0031, respectively.
the alloreactive immune response by affecting the func-tion of both cytokines and chemokines. To elucidate themechanisms responsible for the lack of mononuclear cel-lular infiltration in the presence of a persistent alloreactiv-ity in MMP-9−/− recipients, we performed a semiquan-titative analysis of cytokine and chemokine levels in thesupernatants from the above cultures. As shown inFigure 6A, spleen cells from MMP-9+/+ recipients showedsignificantly higher levels of IL-4, IL-9, IL-17, MCP-5 andGM-CSF as compared to spleen cells from MMP-9−/− re-cipients. In contrast, spleen cells from MMP-9−/− recipi-ents showed significantly higher levels of IL-2, IL-3, IL-12,IL-12p70 and sTNFR1 as compared to spleen cells fromMMP-9+/+ recipients. No differences were observed inthe levels of IL-10, IFN-c and RANTES between MMP-9+/+ and MMP-9−/− spleen cells. No detectable produc-tion of IL-5, IL-6, IL-13, TNF-a and MCP-1 was observedin the cultures (data not shown). In addition, no cytokineproduction was observed by spleen cells from MMP-9+/+and MMP-9−/− recipients cultured in the presence of syn-geneic 129/SvEv cells (data not shown).
To determine whether graft-infiltrating cells displayed asimilar cytokine production profile as the one observedin MMP-9+/+ and MMP-9−/− spleen cells activated exvivo, we performed an analysis of the in situ cytokine
IL-2
IL-3
IL-4
IL-9
IL-1
0
IL-1
2
IL-1
2p70
IL-1
7 γIF
N-
sTN
FR
1
MC
P-5
RA
NT
ES
GM
-CS
F
0
500
1000MMP-9+/+MMP-9-/-
3500
5000
10500
12000
Den
sito
met
y u
nit
s
IL-2
IL-3
IL-4
IL-9
IL-1
0
IL-1
2
IL-1
2p70
IL-1
7 γIF
N-
sTN
FR
1
MC
P-5
RA
NT
ES
GM
-CS
F
0
1000
2000
3000
4000
5000
6000
MMP-9+/+MMP-9-/-
Den
sito
met
ry u
nit
s
A
B
Figure 6: Abnormal cytokine/chemokine production in MMP-
9−/− tracheal allografts recipients. BALB/c allografts weretransplanted into MMP-9+/+ and MMP-9−/− mice. Grafts andspleen cells were harvested at 20 days after transplantation.(A) The MMP-9+/+ and MMP-9−/− spleen cells were culturedin the presence of irradiated BALB/c cells. After 72 h, the cul-ture supernatants were then analyzed by means of an anti-cytokine/chemokine Ab array. (B) Concomitantly, harvested graftswere lysed and analyzed by means an anti-cytokine/chemokine Abarray. Results are expressed as the mean ± SD of duplicate blotsand are representative of three different experiments.
and chemokine production profile in allografts harvestedat 20 days after transplantation. As shown in Figure 6B,the in situ cytokine and chemokine production profile inallografts harvested from MMP-9+/+ and MMP-9−/− re-cipients is remarkably similar to the one displayed by thecorresponding spleen cells activated ex vivo. Allografts har-vested from MMP-9+/+ recipients also showed signifi-cantly higher levels of IL-4, IL-9, MCP-5 and GM-CSF ascompared to allografts harvested from MMP-9−/− recip-ients. In contrast, allografts harvested from MMP-9−/−recipients also showed significantly higher levels of IL-2,IL-3, IL-12, IL-12p70 and TNFR1 as compared to allo-grafts harvested from MMP-9+/+ recipients. No differ-ences were observed in the levels of IL-10, IL-17, IFN-c andRANTES between allografts harvested from MMP-9+/+and MMP-9−/− recipients.
Overall, these results indicate that T cell alloreactivityin MMP-9−/− recipients presents a Th1 predominancewith lower production of IL-4 and a higher productionof cytokines that promote T cell proliferation (IL-2), Th1
678 American Journal of Transplantation 2005; 5: 671–683

MMPs in Obliterative Airway Disease
maturation (IL-12) and inflammation (IL-3, IL-17 and GM-CSF) that may lead to a persistence of the peripheral re-sponse as compared to MMP-9+/+ mice. These resultsalso suggest that MMP-9−/− T cells may have an impairedability to infiltrate the tracheal allografts due to their inabilityto produce sufficient levels of MCP-5 leading to a delayeddevelopment of OAD even in the presence of persistentalloreactivity.
Enhanced innate T cell alloreactivity
in MMP-9−/− mice
Previous studies have shown that MMP-9 is involved inthe process of maturation of Langerhans cells (39). Also,MMP-9 has been shown to be involved in the modula-tion of the activity of several cytokines and chemokines(6,12,30). To determine whether enhanced T cell allore-activity as well as the abnormal production of cytokinesand chemokines in MMP-9−/− recipients was due to a dif-ference in antigen-presentation by DCs or a difference inT cell responsiveness against alloantigens, we first deter-
1500
3000
6000
1200
0
2400
00
40000
80000
120000
160000
200000
MMP-9-/- T CellsMMP-9+/+ T Cells
A
BALB/c DCs/Well
[3 H]-
Td
R u
pta
ke (C
PM
)
1500
3000
6000
1200
0
2400
00
25000
50000
75000
100000
125000
MMP-9-/- DCsMMP-9+/+ DCs
B
129/SvEv DCs/Well
[3 H]-
TdR
upt
ake
(CP
M)
Figure 7: Enhanced innate T cell alloreactivity and DC al-
lostimulatory capacity in MMP-9−/− mice. T cells and DCsfrom MMP-9+/+, MMP-9−/− and BALB/c naıve mice were puri-fied using Ab-coated magnetic beads. (A) T cells from MMP-9+/+and MMP-9−/− mice were cultured in the presence of BALB/cDCs. After 72 h, the incorporation of [3H]-thymidine into DNA wasdetermined by liquid scintillation counting. T cell response: MMP-9+/+ versus MMP-9−/−: p = 0.028. (B) In turn, T cells fromBALB/c mice were cultured in the presence of MMP-9+/+ andMMP-9−/− DCs. After 72 h, the incorporation of [3H]-TdR intoDNA was determined by liquid scintillation counting. DC stimula-tion: MMP-9+/+ versus MMP-9−/−: p < 0.0001. The results areexpressed as the mean ± SD of triplicate cultures and are repre-sentative of three different experiments.
mined the ability of BALB/c DCs to stimulate the prolifer-ation of naıve MMP-9+/+ and MMP-9−/− T cells. Inter-estingly, as shown in Figure 7A, MMP-9−/− T cells dis-played a significantly higher proliferative response againstallogeneic BALB/c DCs as compared to MMP-9+/+ T cells(p = 0.028). It should be noted, however, that the pro-liferative responses of MMP-9−/− T cells to syngeneic129/SvEv DCs were approximately two-fold higher thanthe proliferative responses of MMP-9+/+ T cells to syn-geneic 129/SvEv DCs (data not shown). For this reason,the mean cpm in these negative controls was subtractedfrom the mean cpm in the experimental cultures to presentthe results as net cpm.
Enhanced innate DC allostimulatory capacity in
MMP-9−/− mice
Subsequently, the ability of MMP-9+/+ and MMP-9−/−DCs to stimulate the proliferation of T cells from naıveBALB/c mice was determined. Interestingly, as shown inFigure 7B, DCs from MMP-9−/− mice also showed a sig-nificantly higher capacity to stimulate allogeneic BALB/cT cell proliferation as compared to DCs from MMP-9+/+mice (p < 0.0001). These results indicate that the en-hanced T cell alloreactivity observed in MMP-9−/− recipi-ents is mediated through an enhanced stimulatory capacityof their APCs in addition to an inherently enhanced T cellresponsive capacity.
Production of MMP-9 by parenchymal cells of the
allograft during OAD development
Several studies have shown that activated T cells and in-flammatory cytokines can induce the production of MMPsin several parenchymal cell types such as vascular smoothmuscle cells, fibroblasts, epithelial cells and endothelialcells (18–24). Therefore, the possibility existed that primedalloreactive T cells could have induced the production ofMMP-9 by parenchymal cells of the allograft. To deter-mine whether donor-derived MMP-9 had a contributionto the process of OAD development in the MMP-9−/−recipients, we determined the MMP-9 mRNA and enzy-matic activity levels in BALB/c allografts harvested fromMMP-9+/+ and MMP-9−/− recipients at 20 and 30 daysafter transplantation. As shown in Figure 8A, allograftsharvested from MMP-9−/− recipients revealed the pres-ence of MMP-9 mRNA at 20 days after transplantation.No detectable levels of MMP-9 mRNA were detected inthe allografts harvested from MMP-9−/− recipients at 30days after transplantation. The levels of MMP-9 mRNA inallografts harvested from the MMP-9+/+ recipients was1.7-fold higher than the one observed in the MMP-9−/−recipients at 20 days after transplantation as measuredby densitometric analysis (Figure 8A). Rejected allograftsharvested from MMP-9−/− recipients also showed de-tectable levels of MMP-9 enzymatic activity (Figure 8B,gelatinolytic band at approximately 105 kDa correspondingto pro-MMP-9). No detectable levels of MMP-9 enzymaticactivity were detected in the allografts harvested from
American Journal of Transplantation 2005; 5: 671–683 679
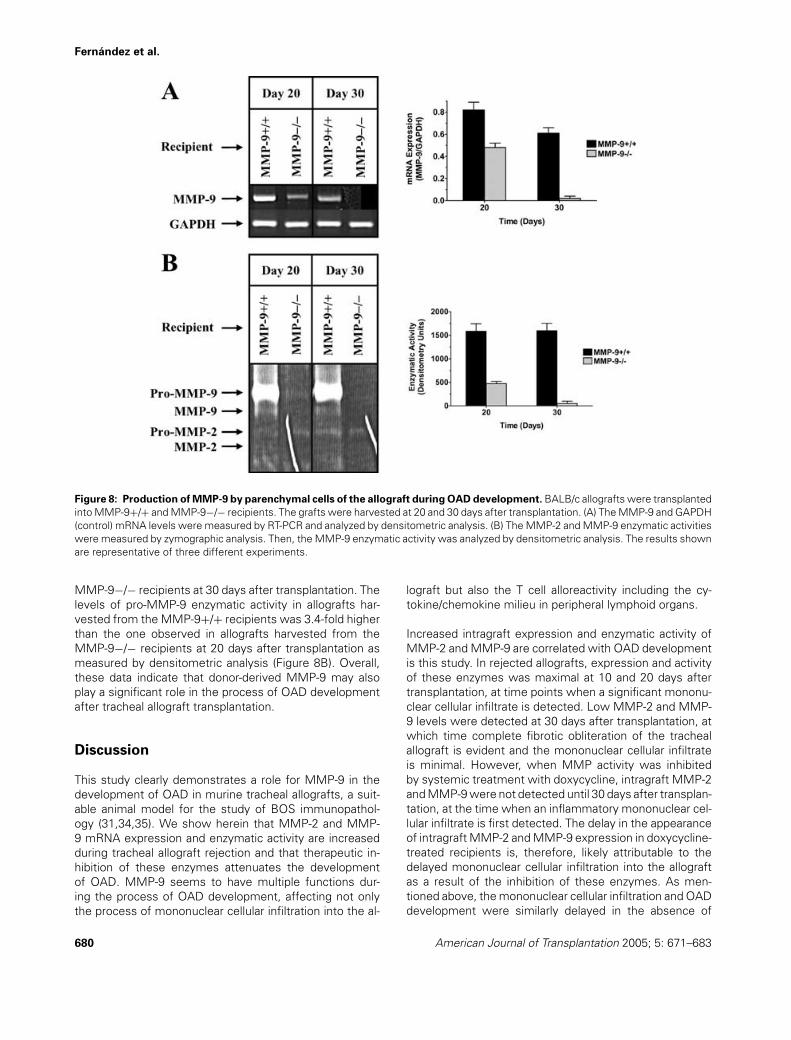
Fernandez et al.
Figure 8: Production of MMP-9 by parenchymal cells of the allograft during OAD development. BALB/c allografts were transplantedinto MMP-9+/+ and MMP-9−/− recipients. The grafts were harvested at 20 and 30 days after transplantation. (A) The MMP-9 and GAPDH(control) mRNA levels were measured by RT-PCR and analyzed by densitometric analysis. (B) The MMP-2 and MMP-9 enzymatic activitieswere measured by zymographic analysis. Then, the MMP-9 enzymatic activity was analyzed by densitometric analysis. The results shownare representative of three different experiments.
MMP-9−/− recipients at 30 days after transplantation. Thelevels of pro-MMP-9 enzymatic activity in allografts har-vested from the MMP-9+/+ recipients was 3.4-fold higherthan the one observed in allografts harvested from theMMP-9−/− recipients at 20 days after transplantation asmeasured by densitometric analysis (Figure 8B). Overall,these data indicate that donor-derived MMP-9 may alsoplay a significant role in the process of OAD developmentafter tracheal allograft transplantation.
Discussion
This study clearly demonstrates a role for MMP-9 in thedevelopment of OAD in murine tracheal allografts, a suit-able animal model for the study of BOS immunopathol-ogy (31,34,35). We show herein that MMP-2 and MMP-9 mRNA expression and enzymatic activity are increasedduring tracheal allograft rejection and that therapeutic in-hibition of these enzymes attenuates the developmentof OAD. MMP-9 seems to have multiple functions dur-ing the process of OAD development, affecting not onlythe process of mononuclear cellular infiltration into the al-
lograft but also the T cell alloreactivity including the cy-tokine/chemokine milieu in peripheral lymphoid organs.
Increased intragraft expression and enzymatic activity ofMMP-2 and MMP-9 are correlated with OAD developmentis this study. In rejected allografts, expression and activityof these enzymes was maximal at 10 and 20 days aftertransplantation, at time points when a significant mononu-clear cellular infiltrate is detected. Low MMP-2 and MMP-9 levels were detected at 30 days after transplantation, atwhich time complete fibrotic obliteration of the trachealallograft is evident and the mononuclear cellular infiltrateis minimal. However, when MMP activity was inhibitedby systemic treatment with doxycycline, intragraft MMP-2and MMP-9 were not detected until 30 days after transplan-tation, at the time when an inflammatory mononuclear cel-lular infiltrate is first detected. The delay in the appearanceof intragraft MMP-2 and MMP-9 expression in doxycycline-treated recipients is, therefore, likely attributable to thedelayed mononuclear cellular infiltration into the allograftas a result of the inhibition of these enzymes. As men-tioned above, the mononuclear cellular infiltration and OADdevelopment were similarly delayed in the absence of
680 American Journal of Transplantation 2005; 5: 671–683

MMPs in Obliterative Airway Disease
MMP-9, but not MMP-2, indicating an important role forMMP-9 in the immunopathogenesis of OAD. The reasonthat mononuclear cellular infiltration does eventually oc-cur remains to be elucidated. This may be explained bythe overlapping functions of MMP family members or bya MMP-independent mechanism. In this regard, a limita-tion of this study is the lack of long-term follow-up at latertime points of MMP-9−/− as well as doxycycline-treatedtracheal allograft recipients to determine the kinetics ofOAD development in these conditions.
Related studies have demonstrated that MMP-9 is involvedin the proteolytic degradation of several active chemokinessuch as growth-related oncogene-a, connective tissue ac-tivating peptide-III and platelet factor-4 (6,12,30). Con-versely, MMP-9 is involved in the activation of TGF-b, TNF-a, IL-1b and IL-8 (6,12,30). Our study corroborates thesefindings as the cytokine profiles produced by spleen cellsex vivo and graft-infiltrating cells in vivo from MMP-9−/−recipients were altered in comparison to MMP-9+/+ recip-ients. The production IL-4, IL-9, IL-17, MCP-5 and GM-CSFproduction was decreased in MMP-9−/− recipients dur-ing OAD development. In contrast, significant increasesin IL-2, IL-3, IL-12, IL-12p70 and sTNFR1 were observedin MMP-9−/− recipients during OAD development. Ourresults on T cell proliferative alloresponses in vitro are inagreement with the results of McMillan et al. (39) show-ing that MMP-9-deficiency results in an enhanced allergen-induced airway inflammation and T cell reactivity to theactivating antigen.
Our results strongly indicate that MMP-9 plays several im-portant roles in the modulation of cytokine and chemokinefunction and, hence, of leukocyte activity during the pro-cess of OAD development. These data indicate that MMP-9 may play a dual role in the process of OAD developmentby modulating mononuclear cellular infiltration into the allo-graft as well as the activation and expansion of alloreactiveT cells. Our results are in agreement with the results ofCataldo et al. (40) that show that MMP-9-deficiency im-pairs lymphocytic inflammation and peribronchial mononu-clear cellular infiltration during allergen-induced airway in-flammation. Additionally, MMP-9-deficiency significantlyaffected the production of MCP-5 upon allogeneic activa-tion. These results suggest that MMP-9 may be involved inthe chemotactic process that drives mononuclear cellularinfiltration into tracheal allografts through the modulationof MCP-5 production. This may explain the lack of mononu-clear cellular infiltration into the allografts in the presenceof a significantly enhanced T cell alloreactivity after trachealtransplantation in MMP-9−/− recipients.
To further determine the role of MMP-9-deficiency in theprocess of T cell activation during the development of OAD,we examined the allostimulatory capacity of APCs and thealloresponsive capacity of T cell from naıve MMP-9+/+ andMMP-9−/− mice. Interestingly, we determined that MMP-9−/− DCs have an enhanced stimulatory capacity as com-
pared to MMP-9+/+ DCs. Surprisingly, we also demon-strated that MMP-9−/− T cells display an enhanced in-nate alloresponsive capacity as compared to MMP-9+/+T cells. These results indicate that MMP-9-deficiency af-fects not only T cell activation and expansion but also al-loantigen presentation by DCs. Our results are in directcontrast with previous studies showing that MMP-9−/−Langerhans cells display normal stimulatory capacity to al-loreactive BALB/c T cells (41). These marked differences inthe stimulatory capacity may be explained by the differentorigin of these DCs.
Allografts harvested from MMP-9−/− recipients revealedthe presence of detectable levels of MMP-9 production.These results indicate that, upon activation, parenchymalcells in the allograft have a significant contribution to theprocess of cellular influx into allograft by means of MMP-9production. Thus, to better define the effect of MMP-9-deficiency on the pathogenesis of OAD, current studies inour laboratory are focused on the introduction of the MMP-9−/− gene into the BALB/c background to use these miceas donors in our tracheal transplantation model.
In conclusion, we show herein that MMP-9 plays adual role during the development of OAD, regulating themononuclear cellular influx into the allograft while modu-lating the activation and expansion of alloreactive T cells.Thus, MMP-9 may represent a novel therapeutic targetfor the treatment and/or prevention of BOS after lungtransplantation.
Acknowledgments
The authors would like to thank Dr. Jeff Atkinson for critical review ofthe manuscript. This work was supported by a Basic Scientist FacultyGrant from the American Society of Transplantation (A.J.), an award fromthe American Heart Association (A.J.), grants HL66452 (T.M.), HL29594(R.M.S.) and HL47328 (R.M.S.) from the National Institutes of Health, theAlan A. and Edith L. Wolff Charitable Trust (R.M.S.) and a Research Fel-lowship from the International Society for Heart and Lung Transplantation(F.G.F.).
References
1. Sundaresan RS, Trulock EP, Mohanakumar T, Cooper JD, Pat-terson GA. Prevalence and outcome of bronchiolitis obliteranssyndrome after lung transplantation. Ann Thorac Surg 1995; 60:1341–1347.
2. Smith MA, Sundaresan RS, Mohanakumar T et al. Effect of devel-opment of antibodies to HLA and CMV mismatch on lung trans-plant survival and development of bronchiolitis obliterans syn-drome. Am J Thorac Surg 1998; 116: 812–820.
3. Zheng L, Ward C, Snell GI et al. Scar collagen deposition in theairways of allografts of lung transplant recipients. Am J RespirCrit Care Med 1997; 155: 2072–2077.
4. Reznik SI, Jaramillo A, SivaSai KSR et al. Indirect allorecognitionof mismatched donor HLA class II peptides in lung transplantrecipients with bronchiolitis obliterans syndrome. Am J Transplant2001; 1: 228–235.
American Journal of Transplantation 2005; 5: 671–683 681

Fernandez et al.
5. Lu KC, Jaramillo A, Mendeloff EN et al. Concomitant allorecog-nition of mismatched donor HLA class I- and II-derived peptidesin pediatric lung transplant recipients with bronchiolitis obliteranssyndrome. J Heart Lung Transplant 2003; 22: 35–43.
6. Goetzl EJ, Banda MJ, Leppert D. Matrix metalloproteinases inimmunity. J Immunol 1996; 156: 1–4.
7. Trello CA, Williams DA, Keller CA, Crim C, Webster RO, Ohar JA.Increased gelatinolytic activity in bronchoalveolar lavage fluid instable lung transplant recipients. Am J Respir Crit Care Med 1997;156: 1978–1986.
8. Beeh KM, Beier J, Kornmann O, Micke P, Buhl R. Sputum levelsof metalloproteinase-9 and tissue inhibitor of metalloproteinase-1and their ratio correlate with airway obstruction in lung transplantrecipients: relation to tumor necrosis factor-a and interleukin-10.J Heart Lung Transplant 2001; 20: 1144–1151.
9. Yano M, Omoto Y, Yamakawa Y et al. Increased matrixmetallopreteinase-9 activity and mRNA expression in lungischemia-reperfusion injury. J Heart Lung Transplant 2001; 20:679–686.
10. Hammer MH, Zhai Y, Katori M et al. Homing of in vitro-generateddonor antigen-reactive CD4+ T lymphocytes to renal allografts isa4b1 but not aLb2 integrin dependent. J Immunol 2001; 166:596–601.
11. Nagase H, Woessner JF Jr. Matrix metalloproteinases. J BiolChem 1999; 274: 21491–21494.
12. Sternlicht MD, Werb Z. How matrix metalloproteinases regulatecell behaviour. Annu Rev Cell Dev Biol 2001; 17: 463–516.
13. Esparza J, Vilardell C, Calco J et al. Fibronectin upregulatesgelatinase B (MMP-9) and induces coordinated expression ofgelatinase A (MMP-2) and its activator MT1-MMP (MMP-14) byhuman T lymphocyte cell lines. A process repressed throughRAS/MAP kinase signaling pathways. Blood 1999; 94: 2754–2766.
14. Kambara C, Nakamura T, Furuya T et al. Vascular cell ad-hesion molecule-1-mediated matrix metalloproteinase-2 induc-tion in peripheral blood T cells is up-regulated in patients withHTLV-I-associated myelopathy. J Neuroimmunol 1999; 99: 242–247.
15. Graesser D, Mahooti S, Haas T, Davis S, Clark RB, Madri JA. Theinterrelationship of (alpha)4 integrin and matrix metalloproteinase-2 in the pathogenesis of experimental autoimmune en-cephalomyelitis. Lab Invest 1998; 78: 1445–1458.
16. Graesser D, Mahooti S, Madri JA. Distince roles for matrixmetalloproteinase-2 and a4 integrin in autoimmune T cell extrava-sation and residency in brain parenchyma during experimental au-toimmune excephalomyelitis. J Neuroimmunol 2000; 109: 121–131.
17. Yakubenko VP, Lobb RR, Plow EF, Ugarova TP. Differential in-duction of gelatinase B (MMP-9) and gelatinase A (MMP-2) inT lymphocytes upon a4b1-mediated adhesion to VCAM-1 andthe CS-1 peptide of fibronectin. Exp Cell Res 2000; 260: 73–84.
18. Lacraz S, Isler P, Vey E, Welgus HG, Dayer JM. Direct contact be-tween T lymphocytes and monocytes is a major pathway for in-duction of metalloproteinase expression. J Biol Chem 1994; 269:22027–22033.
19. Miltenburg AM, Lacraz S, Welgus HG, Dayer JM. Immobilizedanti-CD3 antibody activates T cell clones to induce the produc-tion of interstitial collagenase, but not tissue inhibitor of metal-loproteinases, in monocyte THP-1 cells and dermal fibroblasts. JImmunol 1995; 154: 2655–2667.
20. Schonbeck U, Mach F, Sukhova GK et al. Regulation of matrixmetalloproteinase expression in human vascular smooth muscle
cells by T lymphocytes: a role for CD40 signaling in plaque rupture.Circ Res 1997; 81: 448–454.
21. Burger D, Rezzonico R, Li JM et al. Imbalance between inter-stitial collagenase and tissue inhibitor of metalloproteinases 1 insynoviocytes and fibroblasts upon direct contact with stimulatedT lymphocytes: involvement of membrane-associated cytokines.Arthritis Rheum 1998; 41: 1748–1759.
22. Mach F, Schonbeck U, Fabunmi RP et al. T lymphocytes induceendothelial cell matrix metalloproteinase expression by a CD40L-dependent mechanism: implications for tubule formation. Am JPathol 1999; 154: 229–238.
23. van der Velden VH, Naber BA, van der Spoel P, HoogstedenHC, Versnel MA. Cytokines and flucocorticoids modulate hu-man bronchial epithelial cell peptidases. Cytokine 1998; 10: 55–65.
24. Johansson N, Westermarck J, Leppa S et al. Collagenase-3(matrix metalloproteinase-13) gene expression by HaCaT ker-atinocytes is enhanced by tumor necrosis factor-alpha and trans-forming growth factor-beta. Cell Growth Differ 1997; 8: 243–250.
25. Dubois B, Masure S, Hurtenbach U et al. Resistance of younggelatinase B-deficient mice to experimental autoimmune ex-cephalomyelitis and necrotizing tail lesions. J Clin Invest 1999;104: 1507–1515.
26. Liu Z, Shipley JM, Vu TH et al. Gelatinase-B-deficient mice areresistant to experimental bullous pemphigoid. J Exp Med 1998;188: 475–482.
27. Faber-Elmann A, Grabovsky V, Dayan M, Sela M, Alon R, MozesE. An altered peptide ligand inhibits the activities of matrixmetalloproteinase-9 and phospholipase C, and inhibits T cell in-teractions with VCAM-1 induced in vivo by a myasthenogenic Tcell epitope. FASEB J 2001; 15: 187–194.
28. Itoh T, Matsuda H, Tanioka M, Kuwabara K, Itohara S,Suzuki R. The role of matrix metalloproteinase-2 and matrixmetalloproteinase-9 in antibody-induced arthritis. J Immunol2002; 169: 2643–2647.
29. Ichiyasu H, McCormack JM, McCarthy KM, Dombkowski D, Pref-fer F, Schneeberger EE. MMP-9-deficient dendritic cells have im-paired migration through tracheal epithelial tight junctions. Am JRespir Cell Mol Biol 2004; 30: 761–770.
30. Opdenakker G, Van den Steen PE, Dubois B et al. Gelatinase Bfunctions as regulator and effector in leukocyte biology. J LeukocBiol 2001; 69: 851–859.
31. Hertz MI, Jessurun J, King MB, Murray JJ. Reproduction of theobliterative bronchiolitis lesion after heterotopic transplantationof mouse airways. Am J Pathol 1993; 142: 1945–1951.
32. Itoh T, Ikeda T, Gomi H, Nakao S, Suzuki T, Itohara S. Unalteredsecretion of beta-amyloid precursor protein in gelatinase A (ma-trix metalloproteinase-2)-deficient mice. J Biol Chem 1997; 272:22389–22392.
33. Vu TH, Shipley JM, Bergers G et al. MMP-9/gelatinase B is a keyregulator of growth plate angiogenesis and apoptosis of hyper-trophic chondryocytes. Cell 1998; 90: 411–422.
34. Aris RM, Walsh S, Chalermskulrat W, Hathwar V, Neuringer IP.Growth factor upregulation during obliterative bronchiolitis in themouse model. Am J Respir Crit Care Med 2002; 166: 417–422.
35. Higuchi T, Jaramillo A, Kaleem Z, Patterson GA, MohanakumarT. Different kinetics of obliterative airway disease developmentinduced by CD4+ and CD8+ T cells in murine heterotopic trachealallografts. Transplantation 2002; 74: 646–651.
36. Roach DM, Fitridge RA, Laws PE, Millard SH, Varelias A, CowledPA. Up-regulation of MMP-2 and MMP-9 leads to degradationof type IV collagen during skeletal muscle reperfusion injury;
682 American Journal of Transplantation 2005; 5: 671–683

MMPs in Obliterative Airway Disease
protection by the MMP inhibitor, doxycycline. Eur J Vasc En-dovasc Surg 2002; 23: 260–269.
37. Fukao T, Frucht DM, Yap G, Gadlins M, O’Shea JJ, Koyasu S.Inducible expression of stat4 in dendritic cells and macrophagesand its critical role in innate and adaptive immune responses. JImmunol 2001; 166: 4446–4455.
38. Sheu BC, Hsu SM, Ho HN, Lien HC, Huang SC, Lin RH. A novelrole of metalloproteinase in cancer-mediated immunosuppres-sion. Cancer Res 2001; 61: 237–242.
39. McMillan SJ, Kearley J, Campbell JD et al. Matrix
metalloproteinase-9 deficiency results in enhanced allergen-induced airway inflammation. J Immunol 2004; 172: 2586–2594.
40. Cataldo DD, Tournoy KG, Vermaelen K et al. Ma-trix metalloproteinase-9 deficiency impairs cellular infiltration andbronchial hyperresponsiveness during allergen-induced airwayinflammation. Am J Pathol 2002; 161: 491–498.
41. Ratzinger G, Stoitzner P, Ebner S et al. Matrix metalloproteinases-9 and -2 are necessary for the migration of Langerhans cells anddermal dendritic cells from human and murine skin. J Immunol2002; 168: 4361–4371.
American Journal of Transplantation 2005; 5: 671–683 683




















