
IMMUNE MECHANISMS ASSOCIATED WITH THE REJECTION OF FETAL MURINE PROISLET ALLOGRAFTS AND PIG PROISLET...
9
0041-1337/99/6707-963/0 TRANSPLANTATION Vol. 67, 963–971, No. 7, April 15, 1999 Copyright © 1999 by Lippincott Williams & Wilkins, Inc. Printed in U.S.A. IMMUNE MECHANISMS ASSOCIATED WITH THE REJECTION OF FETAL MURINE PROISLET ALLOGRAFTS AND PIG PROISLET XENOGRAFTS: COMPARISON OF INTRAGRAFT CYTOKINE mRNA PROFILES 1 CHARMAINE J. SIMEONOVIC, 2 MICHELLE J. TOWNSEND, 2 CAROLYN F. MORRIS, 2 ANDREW J. HAPEL, 4 MING-CHIU FUNG, 6 DAVID A. MANN, 3 IAN G. YOUNG 3 AND J. DENNIS WILSON 5,7 Division of Molecular Medicine, Division of Biochemistry and Molecular Biology, Division of Immunology and Cell Biology, The John Curtin School of Medical Research, The Australian National University, G.P.O. Box 334, Canberra, A.C.T., 2601, Australia; Department of Endocrinology, The Canberra Hospital, P.O. Box 11, Woden, A.C.T., 2606, Australia; and the Department of Biology, The Chinese University of Hong Kong, Shatin, N.T., Hong Kong Background. Previous in vivo depletion studies of CD4 and CD8 T cells indicated that different rejection mech- anisms operate for proislet allografts and xenografts. The cellular and molecular mechanisms of acute proislet allograft and xenograft rejection have therefore been characterized and directly compared. Methods. The intragraft cytokine mRNA profile in re- jecting BALB/c (H-2 d ) proislet allografts was analyzed in control, CD4 T cell-depleted, and CD8 T cell-depleted CBA/H (H-2 k ) recipient mice using semi-quantitative re- verse transcriptase-assisted polymerase chain reaction (RT-PCR). The cytokine profiles for proislet allografts and pig proislet xenografts at 3–10 days posttransplant were directly compared and correlated with graft histo- pathology. Results. Allograft rejection was protracted (2–3 weeks), characterized by infiltrating CD8 T cells and CD4 T cells (no eosinophils) and was associated with a Th1-type CD4 T cell response (IL-2, IFN-g, and IL-3 mRNA) and a CD8 T cell-dependent spectrum of cyto- kine gene expression (IL-2, IFN-g, IL-3, and IL-10 mRNA). Xenograft rejection was rapid (6 – 8 days), in- volved predominantly CD4 T cells and eosinophils, and in contrast to allografts, exhibited intragraft mRNA ex- pression for the Th2 cytokines IL-4 and IL-5. Conclusions. Proislet allograft and xenograft rejection differ in the tempo of destruction, phenotype of the cel- lular response and intragraft profile of cytokine mRNA. The recruitment of eosinophils only to the site of xeno- rejection correlates with IL-4 and IL-5 mRNA expres- sion. These findings suggest that different anti-rejection strategies may need to be developed to optimally target the allograft and the xenograft response. For the most effective anti-graft rejection strategy to be developed, the mechanism of transplant rejection needs to be carefully analyzed and clearly defined. It is now evident from rodent studies that the T cell response to neovascularised “cellular” transplants is determined by the type of tissue transplanted and the recipient strain or species. Studies of pancreatic islet tissue transplantation have shown that the relative contribution of CD4 and CD8 T cells to the rejection of murine pancreatic islet allografts depends on the selection of the donor/recipient inbred mouse strains (1–3). In addition, we have previously reported different roles for CD4 and CD8 T cells in the rejection of adult islet allografts and fetal proislet allografts transplanted between the same histoin- compatible mouse strains (3, 4). Although anti-CD8 mAb therapy was sufficient to prevent the rejection of BALB/c (H-2 d ) adult islet allografts in streptozotocin-induced dia- betic CBA/H (H-2 k ) mice, the same treatment was only par- tially effective in prolonging the survival of fetal proislet (islet precursor) allografts in the same mouse strain combi- nation; optimal proislet allograft survival and function re- quired combined anti-CD4 and anti-CD8 mAb therapy (3). In contrast to the allograft response, the rejection of fetal pig proislet xenografts in the same recipient CBA/H strain is exquisitely a CD4 T cell-dependent process (5–7); anti-CD8 mAb treatment is not effective in preventing proislet xeno- graft rejection (5). A role for CD8 T cells in the rejection of adult islet allografts but not xenografts has also been re- ported from studies utilizing b 2 -microglobulin-deficient (CD8 T cell deficient) recipient mice (8). Analysis of the responses of T cell subpopulations to pancreatic islet tissue allografts and xenografts have therefore identified differences in xeno- and allo-rejection mechansims. The molecular mechanism of graft rejection has been inves- tigated by identifying cytokine mRNA profiles at the graft site. According to the Th1/Th2 paradigm, Th1 cells produce IL-2, IFN-g and lymphotoxin whereas Th2 cells produce primarily IL-4, IL-5, IL-6, and IL-10 (9). In addition IL-3, granulocyte, macrophage colony stimulating factor (GM-CSF), and TNF are produced by both Th subpopulations. The rejection of DBA/2 (H-2 d ) pancreatic islets in B6AF1 (H-2 b/k,d ) recipient mice, i.e., 1 This work was supported in part by grants from the Juvenile Diabetes Foundation International, The National Health and Med- ical Research Council (Australia), and the Canberra Hospital Private Practice Fund (Australia). 2 Division of Molecular Medicine, The John Curtin School of Med- ical Research. 3 Division of Biochemistry and Molecular Biology, The John Cur- tin School of Medical Research. 4 Division of Immunology and Cell Biology, The John Curtin School of Medical Research. 5 Department of Endocrinology, The Canberra Hospital. 6 Department of Biology, The Chinese University of Hong Kong, 7 Address correspondence to: C. J. Simeonovic, PhD, Division of Molecular Medicine, The John Curtin School of Medical Research, The Australian National University, G.P.O. Box 334, Canberra, A.C.T., 2601, Australia. 963
-
Upload
hongkonguniversity -
Category
Documents
-
view
0 -
download
0
Transcript of IMMUNE MECHANISMS ASSOCIATED WITH THE REJECTION OF FETAL MURINE PROISLET ALLOGRAFTS AND PIG PROISLET...

0041-1337/99/6707-963/0TRANSPLANTATION Vol. 67, 963–971, No. 7, April 15, 1999Copyright © 1999 by Lippincott Williams & Wilkins, Inc. Printed in U.S.A.
IMMUNE MECHANISMS ASSOCIATED WITH THE REJECTION OFFETAL MURINE PROISLET ALLOGRAFTS AND PIG PROISLET
XENOGRAFTS: COMPARISON OF INTRAGRAFT CYTOKINE mRNAPROFILES1
CHARMAINE J. SIMEONOVIC,2 MICHELLE J. TOWNSEND,2 CAROLYN F. MORRIS,2 ANDREW J. HAPEL,4
MING-CHIU FUNG,6 DAVID A. MANN,3 IAN G. YOUNG3 AND J. DENNIS WILSON5,7
Division of Molecular Medicine, Division of Biochemistry and Molecular Biology,Division of Immunology and Cell Biology, The John Curtin School of Medical Research,
The Australian National University, G.P.O. Box 334, Canberra, A.C.T., 2601, Australia; Department of Endocrinology,The Canberra Hospital, P.O. Box 11, Woden, A.C.T., 2606, Australia; and the Department of Biology,
The Chinese University of Hong Kong, Shatin, N.T., Hong Kong
Background. Previous in vivo depletion studies of CD4and CD8 T cells indicated that different rejection mech-anisms operate for proislet allografts and xenografts.The cellular and molecular mechanisms of acute proisletallograft and xenograft rejection have therefore beencharacterized and directly compared.
Methods. The intragraft cytokine mRNA profile in re-jecting BALB/c (H-2d) proislet allografts was analyzed incontrol, CD4 T cell-depleted, and CD8 T cell-depletedCBA/H (H-2k) recipient mice using semi-quantitative re-verse transcriptase-assisted polymerase chain reaction(RT-PCR). The cytokine profiles for proislet allograftsand pig proislet xenografts at 3–10 days posttransplantwere directly compared and correlated with graft histo-pathology.
Results. Allograft rejection was protracted (2–3weeks), characterized by infiltrating CD8 T cells andCD4 T cells (no eosinophils) and was associated with aTh1-type CD4 T cell response (IL-2, IFN-g, and IL-3mRNA) and a CD8 T cell-dependent spectrum of cyto-kine gene expression (IL-2, IFN-g, IL-3, and IL-10mRNA). Xenograft rejection was rapid (6–8 days), in-volved predominantly CD4 T cells and eosinophils, andin contrast to allografts, exhibited intragraft mRNA ex-pression for the Th2 cytokines IL-4 and IL-5.
Conclusions. Proislet allograft and xenograft rejectiondiffer in the tempo of destruction, phenotype of the cel-lular response and intragraft profile of cytokine mRNA.The recruitment of eosinophils only to the site of xeno-rejection correlates with IL-4 and IL-5 mRNA expres-sion. These findings suggest that different anti-rejection
strategies may need to be developed to optimally targetthe allograft and the xenograft response.
For the most effective anti-graft rejection strategy to bedeveloped, the mechanism of transplant rejection needs to becarefully analyzed and clearly defined. It is now evident fromrodent studies that the T cell response to neovascularised“cellular” transplants is determined by the type of tissuetransplanted and the recipient strain or species. Studies ofpancreatic islet tissue transplantation have shown that therelative contribution of CD4 and CD8 T cells to the rejectionof murine pancreatic islet allografts depends on the selectionof the donor/recipient inbred mouse strains (1–3). In addition,we have previously reported different roles for CD4 and CD8T cells in the rejection of adult islet allografts and fetalproislet allografts transplanted between the same histoin-compatible mouse strains (3, 4). Although anti-CD8 mAbtherapy was sufficient to prevent the rejection of BALB/c(H-2d) adult islet allografts in streptozotocin-induced dia-betic CBA/H (H-2k) mice, the same treatment was only par-tially effective in prolonging the survival of fetal proislet(islet precursor) allografts in the same mouse strain combi-nation; optimal proislet allograft survival and function re-quired combined anti-CD4 and anti-CD8 mAb therapy (3). Incontrast to the allograft response, the rejection of fetal pigproislet xenografts in the same recipient CBA/H strain isexquisitely a CD4 T cell-dependent process (5–7); anti-CD8mAb treatment is not effective in preventing proislet xeno-graft rejection (5). A role for CD8 T cells in the rejection ofadult islet allografts but not xenografts has also been re-ported from studies utilizing b2-microglobulin-deficient (CD8T cell deficient) recipient mice (8). Analysis of the responsesof T cell subpopulations to pancreatic islet tissue allograftsand xenografts have therefore identified differences in xeno-and allo-rejection mechansims.
The molecular mechanism of graft rejection has been inves-tigated by identifying cytokine mRNA profiles at the graft site.According to the Th1/Th2 paradigm, Th1 cells produce IL-2,IFN-g and lymphotoxin whereas Th2 cells produce primarilyIL-4, IL-5, IL-6, and IL-10 (9). In addition IL-3, granulocyte,macrophage colony stimulating factor (GM-CSF), and TNF areproduced by both Th subpopulations. The rejection of DBA/2(H-2d) pancreatic islets in B6AF1 (H-2b/k,d) recipient mice, i.e.,
1 This work was supported in part by grants from the JuvenileDiabetes Foundation International, The National Health and Med-ical Research Council (Australia), and the Canberra Hospital PrivatePractice Fund (Australia).
2 Division of Molecular Medicine, The John Curtin School of Med-ical Research.
3 Division of Biochemistry and Molecular Biology, The John Cur-tin School of Medical Research.
4 Division of Immunology and Cell Biology, The John CurtinSchool of Medical Research.
5 Department of Endocrinology, The Canberra Hospital.6 Department of Biology, The Chinese University of Hong Kong,7 Address correspondence to: C. J. Simeonovic, PhD, Division of
Molecular Medicine, The John Curtin School of Medical Research,The Australian National University, G.P.O. Box 334, Canberra,A.C.T., 2601, Australia.
963

H-2K 1 I 2 mismatched allogeneic strains, has been corre-lated with enhanced expression of IL-2 and IFN-g transcriptsat day 8 and days 8–12 posttransplant, respectively; en-hanced IL-4 mRNA expression was not detected at any time-point. These findings suggested that the rejection of adultislet allografts resembles a Th1-type response generated, atleast in part, by CD8 T cells (10). In contrast, our studies ofthe acute rejection of fetal pig proislet xenografts in CBA/Hmice showed a clear correlation between rejection and CD4 Tcell-dependent intragraft expression of IL-3, IL-4, and IL-5mRNA (7); this cytokine profile is consistent with a Th2-typeresponse. To clearly identify differences in the mechanisms ofallograft and xenograft rejection, we have now examined thecytokine mRNA profile in acutely rejecting BALB/c (H-2d)mouse proislet allografts in completely MHC-mismatchedCBA/H (H-2k) recipient mice and directly compared the cy-tokine mRNA expression in pig proislet xenografts trans-planted to the same recipient mouse strain. Our study dem-onstrates that unlike xenograft rejection, the allograftrejection response does not involve a Th2-type reaction; thepresence/absence of strong intragraft expression of Th2 cyto-kine mRNAs (IL-4, IL-5) correlates with the presence/absence of eosinophil infiltration.
MATERIALS AND METHODS
Animals. CBA/H male mice (H-2k; 7.5–13 wk old), were providedby the Animal Services Division, The Australian National University(Canberra, Australia) and were used as transplant recipients.BALB/c and CBA/H fetuses at 17 days of gestation were obtained byhysterotomy of time-mated pregnant BALB/c and CBA/H femalemice, respectively (11, 12). Swiss outbred nude mice obtained fromthe Animal Services Division, The Australian National Universitywere used for the generation of ascites preparations of monoclonalantibodies. Fetal piglets of the SLAdd haplotype (68–75 days ofgestation) were obtained via Caesarian section of pregnant SLAdd
gilts/sows belonging to the inbred pig colony maintained at theAnimal Services Division, The Australian National University, Can-berra. This colony was derived from the inbred SLAdd herd (13)developed by Professor David H. Sachs (Boston, MA).
Preparation and transplantation of fetal proislets. Proislets (isletprecursors) were prepared from BALB/c (H-2d) or CBA/H (H-2k) fetalpancreases (17 days of gestation) by collagenase digestion followedby culture of the digested tissue in 10% CO2, 90% air for 4 days (11,12). Fetal pig proislets were prepared from fetal pig pancreas bycollagenase digestion followed by organ culture of the digested tissuefor 5–6 days in 10% CO2, 90% air (7, 14). To facilitate transplanta-tion proislets were embedded in a plasma clot prepared from recip-ient mouse strain plasma (1/20 fetal pig pancreas equivalent ofproislets or two fetal mouse pancreas equivalents of proislets/clot);six plasma clots containing mouse proislets or one or two plasmaclot(s) containing pig proislets were transplanted beneath the kidneycapsule of recipient mice anesthetised with avertin (7, 14). The dayof transplantation was designated day 0.
Preparation and administration of mAb. GK1.5 (anti-CD4) (15)and 49–11.1 (anti-CD8) (16) hybridoma cells were grown in culturefor the preparation of ascites in Swiss nude mice pretreated withpristane (Sigma Chemical Co., St. Louis, MO) (7). The Ig content ofpooled GK1.5 or 49–11.1 ascites preparations was determined by asandwich enzyme linked immunosorbent assay (ELISA) (7). GK1.5monoclonal antibody (1.0 mg Ig/dose) or 49–11.1 monoclonal anti-body (1.1–1.7 mg Ig/dose) was administered i.p. to transplant recip-ient mice on days 21 (day before transplantation), 1, 3, 7, and 10.CD4 or CD8 T cell depletion in peripheral lymph nodes of treatedmice was monitored by two-color flow cytometry using H129.19 (anti-CD4) (17) and 53.6.7 (anti-CD8) (18) rat anti-mouse monoclonal
antibodies to identify CD4 and CD8 T cells, respectively (7, 14).Monoclonal antibody-treated mice received oral antibiotics (oxytet-racycline hydrochloride, Terramycin 50, Pfizer Agricare Pty Ltd,West Ryde, NSW, Australia) in their drinking water (125 mg terra-mycin/300 ml); the antibiotic supplement was replaced every 3–4days.
Harvest of transplants and tissues for RNA extraction and immu-nohistochemistry. Transplant recipient mice at day 3–21 posttrans-plant were killed by cervical dislocation. The graft-bearing kidneywas promptly removed and 80–100% of the transplant was strippedfrom the kidney parenchyma and frozen in liquid nitrogen for sub-sequent RNA extraction (7). The remainder of such transplants orentire additional transplants were frozen in situ in liquid freon(preequilibrated with liquid nitrogen) for histology and immunohis-tochemistry. BALB/c and CBA/H mouse proislets, pig proislets, andkidney tissue harvested from CBA/H mice were also frozen in liquidnitrogen as control tissues for RNA extraction and subsequent anal-ysis of cytokine mRNA by reverse transcriptase-assisted polymerasechain reaction (RT-PCR*).
Histology and immunohistochemistry. Freon-frozen graft tissuewas sectioned (6-m thickness) and stained for presence of infiltratingCD4 and CD8 T cells using rat anti-mouse monoclonal antibodiesH129.19 and 53.6.7, respectively, and avidin-biotin-peroxidase im-munostaining methods incorporating a second step incubation withbiotinylated sheep anti-rat Ig (Cat. RPN.1002, Amersham, LittleChalfont, UK) followed by the peroxidase substrate 0.05% 3,39-dia-mino-benzidine tetrahydrochloride (Cat. D-5637, Sigma)/0.07% imi-dazole (pH 7.4)/0.02% H2O2 in phosphate-buffered saline (14). Alter-natively T cell subpopulations were identified using peroxidase-conjugated sheep anti-rat Ig (Cat. NA.932, Amersham; 1/50 inphosphate-buffered saline) (19). Freon-frozen sections of mouse thy-mus were used as positive controls for the immunostaining. Eosino-phils were identified in control sections incubated with phosphate-buffered saline due to their high level of endogenous peroxidaseactivity (7, 14, 20). Immunostained sections were counterstainedwith Mayer’s hematoxylin. Additional frozen sections were stainedwith hematoxylin and eosin to assess graft integrity/rejection.
RNA extraction and reverse transcription. Total graft/tissue RNAwas extracted using the guanidine isothiocyanate method (21) aspreviously described (7), and stored at 270°C. RNA (1–5 mg) wasreverse transcribed in 20–100 ml volume, respectively, using 100 UMaloney murine leukemia virus reverse transcriptase (Gibco BRL,Life Technologies, Inc., Gaithersburg, MD)/mg RNA, 40U RNasin(Promega, Madison, WI)/mg RNA, 0.5 mM (each) dNTP mix (Phar-macia, Piscataway, NJ), 0.1 mg oligo dT12–18 (Pharmacia)/mg RNA, 10mM dithiothreitol (supplied with enzyme) and reverse transcriptasebuffer (supplied with enzyme) for 1 hr at 37°C (7). Grafts to bedirectly compared were reverse transcribed concurrently using thesame reverse transcriptase mix; cDNA samples were then stored at220°C.
PCR analysis. Semiquantitative PCR was performed using aDNA thermal cycler (Gene Machine, Innovonics Pty Ltd, Melbourne,Vic, Australia). cDNA (20 ml) was amplified in a 50 ml reactionvolume containing 0.2 mM (each) deoxynucleotide triphosphates,1–5 mM MgCl2, 0.4–1 mM primers, 1 U Taq (Promega), 0.25U SU-PER TAQ (P.H. Stehelin & CIE AG, Basel, Switzerland) or 2 UThermalase rec-Tbr DNA polymerase (Amresco, Solon, OH) andreaction buffer (supplied with enzyme). The reaction mix was over-laid with mineral oil (Sigma); 10-ml aliquots of the reaction mix weresampled at five cycle intervals from 20 to 45 cycles. Standard reac-tion parameters were 96°C (1 min), 45–72°C (1 min), 70–72°C (1min). Primers for b-actin were used as controls to indicate that therewere similar levels of amplifiable cDNA in tissue samples. Annealingtemperatures and nucleotide sequences of the primer pairs were as
* Abbreviations: RT-PCR, reverse transcriptase-assisted polymer-ase chain reaction; IL, interleukin; INF, interferon; APC, antigen-presenting cells.
TRANSPLANTATION964 Vol. 67, No. 7

follows: IL-2 (72°C), TTGATGGACCTACAGGAGCTCCTGAGCA andAGAGAGCCTTATGTGTTGTAAGCAGGAGG ; IFN-g (56–62°C), TCCT-GCAGAGCCAGATTATCTC and GTGCTGGCAGAATTATTCTTATTG;IL-3 (72°C), ATAGGGAAGCTCCCAGAACCTGAACTC and AGACCCCT-GGCAGCGCAGAGTCATTC; IL-4 (45–62°C), TGACGCACAGAGCTAT-TGATGG and ATGATGCTCTTTAGGCTTTCCAG; IL-5 (52–54°C), AA-GATGCTTCTGCACTTGA and ACACCAAGGAACTCTTGCA (22);IL-10 (56°C), TAGAGCTGCGGACTGCCTTCA and TCATGGCCTT-GTAGACACCTTG; b-actin (55°C) ATGGATGACGATATCGCT andATGAGGTAGTCTGTCAGGT.
Con A (2 mg/ml) and phorbol myristate acetate (5 ng/ml)-activatedmouse spleen cells served as a positive control for interleukin (IL) -2,interferon-g, IL-4, IL-10 and b-actin mRNA (7); WEHI-3B cells (23),and D10.G4.1 cells (24) were used as positive controls for IL-3 andIL-5 transcripts, respectively. PCR products were size-fractionatedby electrophoresis through 1.5–2.0% agarose gels; the gels werestained in ethidium bromide solution (2 mg/ml) for 10–20 min andphotographed under UV illumination. The sizes of the PCR productswere verified against a marker of known standards (1 kb DNAladder, Gibco, BRL, Life Technologies Inc., Gaithersburg, MD).
RESULTS
Histological and Immunohistochemical Analyses of ProisletGraft Rejection in CBA/H Recipient Mice
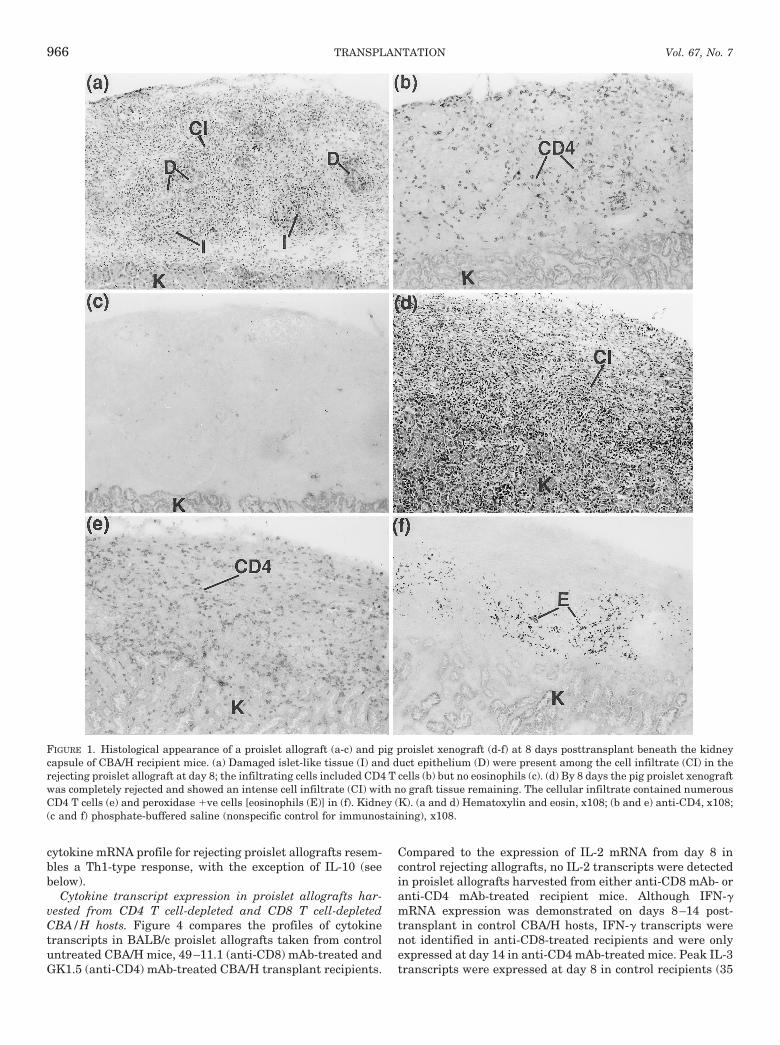
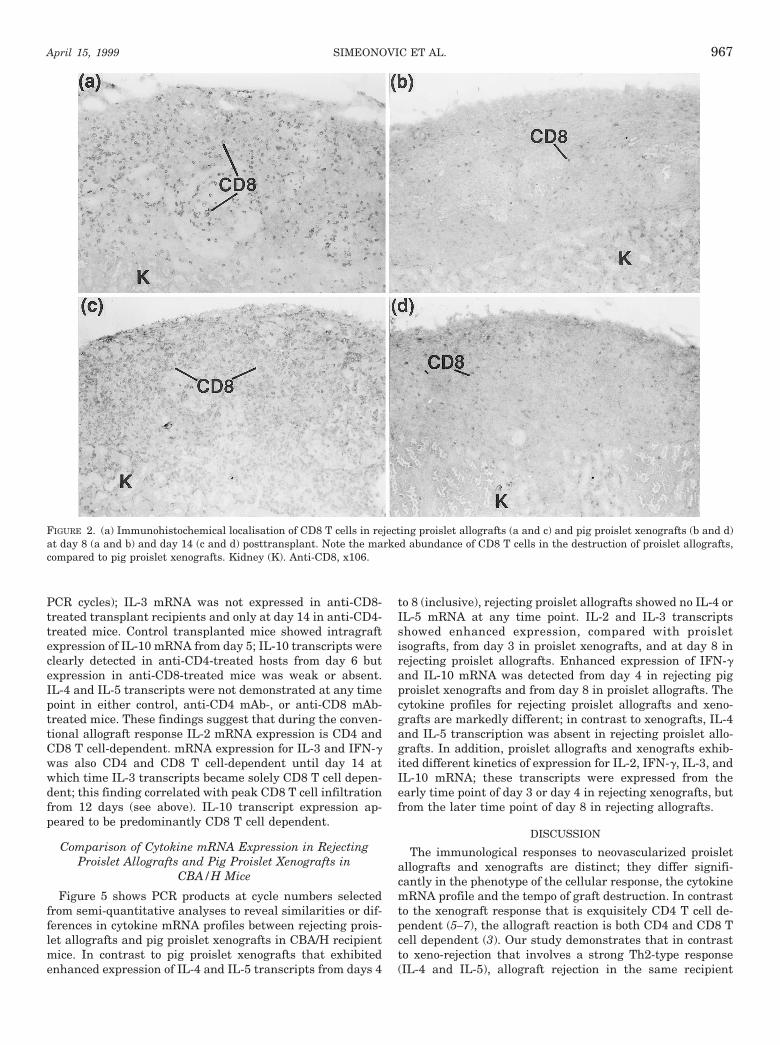
BALB/c proislet allografts. At 3–4 days posttransplantBALB/c proislet allografts in CBA/H recipient mice showedpresence of intact developing islet tissue and duct epithelium(from which precursor islet tissue differentiates) with sparselyscattered CD4 and CD8 T cells. By 6 days posttransplant theproislet allografts showed evidence of graft damage with morepronounced infiltration by CD4 and CD8 T cells (Table 1);intense cell infiltration was observed from 8 days (Fig. 1a) andmaximal CD8 T cell infiltration from 12 days (Fig. 2c). Dam-aged islet tissue and duct epithelium were identified in graftsharvested at 6–14 days posttransplant, suggesting a chronicprocess of graft destruction. Eosinophils were not detected atany time point during proislet allograft rejection (Table 1, Fig.1c). In contrast to proislet allografts, isografts of CBA/H prois-lets showed excellent development of islet tissue and persis-
tence of duct epithelium by 2 weeks posttransplant (Table 1).The rejection response to proislet allografts in CBA/H miceconsisted of CD4 T cells, CD8 T cells but no eosinophils (Figs. 1and 2); graft destruction was well advanced by 2 weeks andcomplete by 3 weeks posttransplantation (data not presented).
Comparison of the histopathology of proislet allograft andxenograft rejection. Pig proislet xenografts at 3–4 days aftertransplantation contained histological evidence of intact de-veloping pig islet tissue and duct epithelium (Table 1). At 5days, intact islet-like tissue and damaged graft tissue wereidentified at the graft site and by 6 days posttransplant thexenografts were either extensively or completely destroyed(Table 1). There was no evidence of graft tissue remaining by8 days (Fig. 1d). Compared with proislet allografts thatshowed on-going graft damage at 8 days posttransplant,proislet xenografts were completely destroyed, i.e., the tempoof the allograft response was markedly slower. The xenograftinfiltrate consisted mainly of CD4 T cells, eosinophils (Fig. 1,e and f) and some CD8 T cells (Fig. 2, b and d). Compared tothe xenograft reaction, CD8 T cell infiltration was more pro-nounced and eosinophils did not participate in the rejectionof proislet allografts. CD4 T cells were prominent in thecellular infiltrates of both rejecting allografts and xenografts.
Expression of Intragraft Cytokine Transcripts DuringRejection of BALB/c Proislet Allografts in CBA/H
Recipient Mice
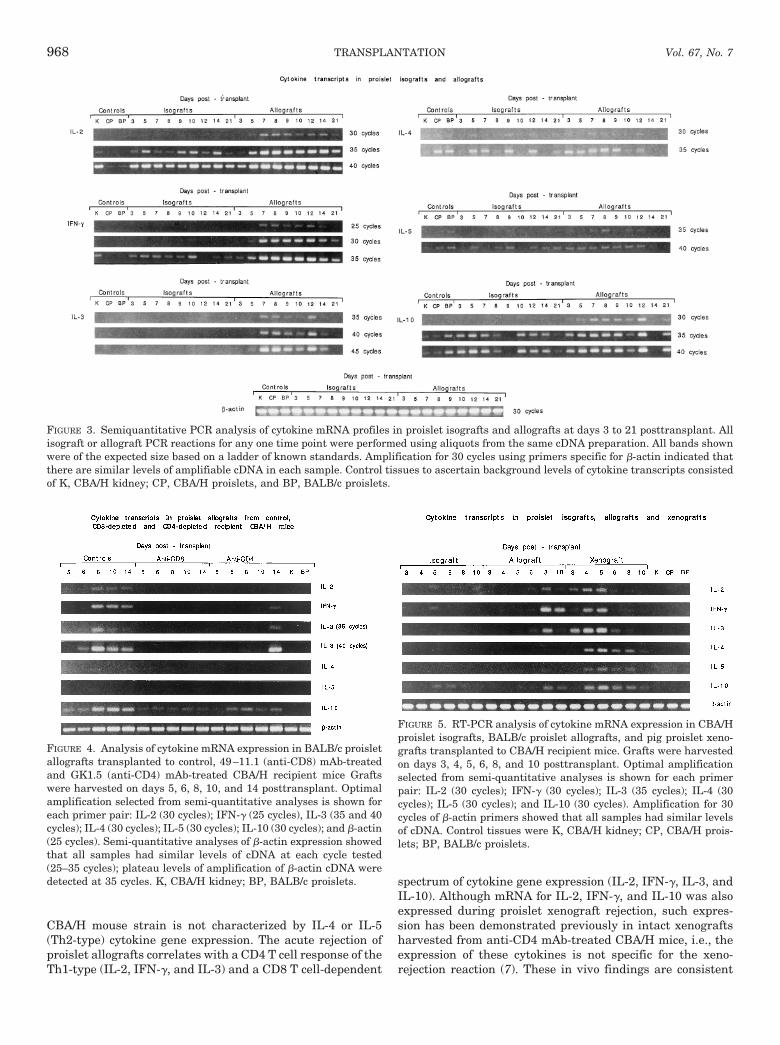
Characterization of the cytokine profile for allograft rejec-tion. Figure 3 shows semi-quantitative PCR analyses of cy-tokine mRNA expression in proislet allografts and isograftsfrom day 3 to 21 posttransplant. Compared with proisletisografts, proislet allografts showed enhanced transcript ex-pression from day 7 for IL-2, IFN-g, IL-3, and IL-10; mRNAexpression persisted to day 14 for IL-2 and IL-3, and up today 21 for IFN-g and IL-10. No enhanced IL-4 transcriptexpression was detected in proislet allografts at any timepoint; IL-5 mRNA was weakly expressed only at day 8. The
TABLE 1. Histological and immunohistochemical assessment of proislet allografts and pig proislet xenografts in CBA/H recipient mice
Transplant Day post transplant(n5no. of grafts)
Number of grafts with histological/immunohistochemical score
Total cellinfiltratea
CD4 T cellinfiltratea
CD8 T cellinfiltratea
Eosinophilinfiltrateb
Proislet/ductintegrityc
0 1 2 3 0 1 2 3 0 1 2 3 0 1 2 3 0 1 2 3
Isograftday 3 – day 4 (n52) 1 1 1 1 2 2 2day 5 – day 9 (n55) 5 5 3 2 5 5day 10 – day 14 (n53) 3 3 2 1 3 3
Allograftday 3 – day 4 (n52) 2 2 2 2 1 1day 5 – day 9 (n55) 1 2 2 1 4 1 4 5 3 2day 10 – day 14 (n53) 3 2 1 1 2 3 3
Xenograftday 3 – day 4 (n53) 1 1 1 1 2 2 1 2 1 1 2day 5 – day 8 (n55) 5 1 4 4 1 2 3 1 3 1day 10 – day 14 (n54) 1 3 4 2 2 4 4
a Score for total cell, CD4 and CD8 T cell infiltrate: 0, none or few infiltrating cells; 1, scattered infiltrate or focal accumulation; 2, moderateinfiltrate; 3, intense cell infiltrate.
b Score for eosinophil infiltrate: 0, none or few peroxidase 1ve cells; 1, sparsely scattered infiltrate; 2, mild infiltrate; 3, marked or intenseinfiltrate.
c Score for proislet/duct integrity: 0, only tissue remnants or no evidence of islet/duct tissue; 1, much damaged islet/duct tissue present; 2,intact islet/duct tissue1damaged tissue; 3, much intact islet/duct tissue.
SIMEONOVIC ET AL.April 15, 1999 965
cytokine mRNA profile for rejecting proislet allografts resem-bles a Th1-type response, with the exception of IL-10 (seebelow).
Cytokine transcript expression in proislet allografts har-vested from CD4 T cell-depleted and CD8 T cell-depletedCBA/H hosts. Figure 4 compares the profiles of cytokinetranscripts in BALB/c proislet allografts taken from controluntreated CBA/H mice, 49–11.1 (anti-CD8) mAb-treated andGK1.5 (anti-CD4) mAb-treated CBA/H transplant recipients.
Compared to the expression of IL-2 mRNA from day 8 incontrol rejecting allografts, no IL-2 transcripts were detectedin proislet allografts harvested from either anti-CD8 mAb- oranti-CD4 mAb-treated recipient mice. Although IFN-gmRNA expression was demonstrated on days 8–14 post-transplant in control CBA/H hosts, IFN-g transcripts werenot identified in anti-CD8-treated recipients and were onlyexpressed at day 14 in anti-CD4 mAb-treated mice. Peak IL-3transcripts were expressed at day 8 in control recipients (35
FIGURE 1. Histological appearance of a proislet allograft (a-c) and pig proislet xenograft (d-f) at 8 days posttransplant beneath the kidneycapsule of CBA/H recipient mice. (a) Damaged islet-like tissue (I) and duct epithelium (D) were present among the cell infiltrate (CI) in therejecting proislet allograft at day 8; the infiltrating cells included CD4 T cells (b) but no eosinophils (c). (d) By 8 days the pig proislet xenograftwas completely rejected and showed an intense cell infiltrate (CI) with no graft tissue remaining. The cellular infiltrate contained numerousCD4 T cells (e) and peroxidase 1ve cells [eosinophils (E)] in (f). Kidney (K). (a and d) Hematoxylin and eosin, x108; (b and e) anti-CD4, x108;(c and f) phosphate-buffered saline (nonspecific control for immunostaining), x108.
TRANSPLANTATION966 Vol. 67, No. 7
PCR cycles); IL-3 mRNA was not expressed in anti-CD8-treated transplant recipients and only at day 14 in anti-CD4-treated mice. Control transplanted mice showed intragraftexpression of IL-10 mRNA from day 5; IL-10 transcripts wereclearly detected in anti-CD4-treated hosts from day 6 butexpression in anti-CD8-treated mice was weak or absent.IL-4 and IL-5 transcripts were not demonstrated at any timepoint in either control, anti-CD4 mAb-, or anti-CD8 mAb-treated mice. These findings suggest that during the conven-tional allograft response IL-2 mRNA expression is CD4 andCD8 T cell-dependent. mRNA expression for IL-3 and IFN-gwas also CD4 and CD8 T cell-dependent until day 14 atwhich time IL-3 transcripts became solely CD8 T cell depen-dent; this finding correlated with peak CD8 T cell infiltrationfrom 12 days (see above). IL-10 transcript expression ap-peared to be predominantly CD8 T cell dependent.
Comparison of Cytokine mRNA Expression in RejectingProislet Allografts and Pig Proislet Xenografts in
CBA/H Mice
Figure 5 shows PCR products at cycle numbers selectedfrom semi-quantitative analyses to reveal similarities or dif-ferences in cytokine mRNA profiles between rejecting prois-let allografts and pig proislet xenografts in CBA/H recipientmice. In contrast to pig proislet xenografts that exhibitedenhanced expression of IL-4 and IL-5 transcripts from days 4
to 8 (inclusive), rejecting proislet allografts showed no IL-4 orIL-5 mRNA at any time point. IL-2 and IL-3 transcriptsshowed enhanced expression, compared with proisletisografts, from day 3 in proislet xenografts, and at day 8 inrejecting proislet allografts. Enhanced expression of IFN-gand IL-10 mRNA was detected from day 4 in rejecting pigproislet xenografts and from day 8 in proislet allografts. Thecytokine profiles for rejecting proislet allografts and xeno-grafts are markedly different; in contrast to xenografts, IL-4and IL-5 transcription was absent in rejecting proislet allo-grafts. In addition, proislet allografts and xenografts exhib-ited different kinetics of expression for IL-2, IFN-g, IL-3, andIL-10 mRNA; these transcripts were expressed from theearly time point of day 3 or day 4 in rejecting xenografts, butfrom the later time point of day 8 in rejecting allografts.
DISCUSSION
The immunological responses to neovascularized proisletallografts and xenografts are distinct; they differ signifi-cantly in the phenotype of the cellular response, the cytokinemRNA profile and the tempo of graft destruction. In contrastto the xenograft response that is exquisitely CD4 T cell de-pendent (5–7), the allograft reaction is both CD4 and CD8 Tcell dependent (3). Our study demonstrates that in contrastto xeno-rejection that involves a strong Th2-type response(IL-4 and IL-5), allograft rejection in the same recipient
FIGURE 2. (a) Immunohistochemical localisation of CD8 T cells in rejecting proislet allografts (a and c) and pig proislet xenografts (b and d)at day 8 (a and b) and day 14 (c and d) posttransplant. Note the marked abundance of CD8 T cells in the destruction of proislet allografts,compared to pig proislet xenografts. Kidney (K). Anti-CD8, x106.
SIMEONOVIC ET AL.April 15, 1999 967
CBA/H mouse strain is not characterized by IL-4 or IL-5(Th2-type) cytokine gene expression. The acute rejection ofproislet allografts correlates with a CD4 T cell response of theTh1-type (IL-2, IFN-g, and IL-3) and a CD8 T cell-dependent
spectrum of cytokine gene expression (IL-2, IFN-g, IL-3, andIL-10). Although mRNA for IL-2, IFN-g, and IL-10 was alsoexpressed during proislet xenograft rejection, such expres-sion has been demonstrated previously in intact xenograftsharvested from anti-CD4 mAb-treated CBA/H mice, i.e., theexpression of these cytokines is not specific for the xeno-rejection reaction (7). These in vivo findings are consistent
FIGURE 3. Semiquantitative PCR analysis of cytokine mRNA profiles in proislet isografts and allografts at days 3 to 21 posttransplant. Allisograft or allograft PCR reactions for any one time point were performed using aliquots from the same cDNA preparation. All bands shownwere of the expected size based on a ladder of known standards. Amplification for 30 cycles using primers specific for b-actin indicated thatthere are similar levels of amplifiable cDNA in each sample. Control tissues to ascertain background levels of cytokine transcripts consistedof K, CBA/H kidney; CP, CBA/H proislets, and BP, BALB/c proislets.
FIGURE 4. Analysis of cytokine mRNA expression in BALB/c proisletallografts transplanted to control, 49–11.1 (anti-CD8) mAb-treatedand GK1.5 (anti-CD4) mAb-treated CBA/H recipient mice Graftswere harvested on days 5, 6, 8, 10, and 14 posttransplant. Optimalamplification selected from semi-quantitative analyses is shown foreach primer pair: IL-2 (30 cycles); IFN-g (25 cycles), IL-3 (35 and 40cycles); IL-4 (30 cycles); IL-5 (30 cycles); IL-10 (30 cycles); and b-actin(25 cycles). Semi-quantitative analyses of b-actin expression showedthat all samples had similar levels of cDNA at each cycle tested(25–35 cycles); plateau levels of amplification of b-actin cDNA weredetected at 35 cycles. K, CBA/H kidney; BP, BALB/c proislets.
FIGURE 5. RT-PCR analysis of cytokine mRNA expression in CBA/Hproislet isografts, BALB/c proislet allografts, and pig proislet xeno-grafts transplanted to CBA/H recipient mice. Grafts were harvestedon days 3, 4, 5, 6, 8, and 10 posttransplant. Optimal amplificationselected from semi-quantitative analyses is shown for each primerpair: IL-2 (30 cycles); IFN-g (30 cycles); IL-3 (35 cycles); IL-4 (30cycles); IL-5 (30 cycles); and IL-10 (30 cycles). Amplification for 30cycles of b-actin primers showed that all samples had similar levelsof cDNA. Control tissues were K, CBA/H kidney; CP, CBA/H prois-lets; BP, BALB/c proislets.
TRANSPLANTATION968 Vol. 67, No. 7

with in vitro studies of mixed lymphocyte reaction reactivitythat revealed polarized Th cytokine mRNA expression afterallo- or xeno-antigen activation; murine alloreactivity corre-lated with a Th1-type cytokine profile (IL-2, IFN-g) and themouse anti-rat proliferative response generated a Th2-typeprofile (25).
Proislet xenografts are more rapidly destroyed than prois-let allografts, i.e., 1 vs. 2 to 3 weeks, respectively; neverthe-less proislet xeno-rejection does not reflect a hyperacute an-tibody-mediated mechanism as has been demonstrated forimmediately vascularized organ xenografts (26). Proisletxenografts are cellular transplants that become neovascular-ized by recipient-type vascular endothelium. As a result, thetarget for hyperacute rejection, donor antigens displayed ondonor-type vascular endothelium, is therefore absent. Theacute destruction of fetal pig proislet and islet-like cell clus-ter xenografts in mMT -/- (antibody-deficient) recipient mice(27, 28) supports this premise and argues against a majorrole for humoral immunity in the conventional process ofacute proislet xenograft rejection. The apparent differencesin proislet allograft and xenograft rejection mechanisms cantherefore not be explained by implicating a major role forxenoantibody; instead both rejection mechanisms involve cel-lular immune responses. It is unlikely that the slower tempofor allograft rejection, compared to the xenograft rejectionprocess, can be attributed to a lower precursor frequency foralloreactive T cells; other groups have demonstrated thatalloreactive T cells can exhibit a 6- to 20-fold higher precur-sor frequency than xenoreactive T cells (29, 30). Our studyshows that whereas CD4 T cells and eosinophils dominatethe cellular response to proislet xenografts, CD8 T cells andCD4 T cells (but not eosinophils) represent major partici-pants in the allograft reaction in CBA/H recipient mice. Wehave previously reported that the recruitment of esoinophilsto the site of proislet xenograft rejection is regulated by IL-4,IL-5, and IL-3 (20). We therefore conclude that the absence ofeosinophil recruitment to the proislet allograft rejection re-action is most likely due to absence of intragraft expression ofIL-4 and IL-5 mRNA. Expression of IL-3 transcripts duringthe allograft response appears to be insufficient stimulus foreosinophil mobilization or does not result in IL-3 proteinproduction. The absence of an eosinophil response to reject-ing proislet allografts is in contrast to eosinophil accumula-tion in rejecting, vascularized human renal and liverallografts (31, 32); the latter correlates with intragraft ex-pression of the Th2 cytokine, IL-5 (33). The recruitment ofeosinophils during graft rejection therefore acts as a usefulmarker for the intragraft expression of Th2 cytokines (e.g.,IL-5) and provides indirect evidence of Th2 cytokine proteinproduction. Although eosinophil infiltration correlates withpig proislet xenograft rejection, we have demonstrated thatxenograft rejection proceeds unimpeded in IL-4 gene knock-out and IL-5 gene knockout recipient mice, in the absence ofan eosinophil response (20). Eosinophils therefore are notcritical mediators of pig proislet xenograft rejection; it ispossible, however, that after binding to anti-pig IgG depos-ited within the graft eosinophils may function as an acessoryeffector of proislet tissue destruction via release of toxic prod-ucts such as major basic protein, eosinophil peroxidase,and/or reactive oxygen intermediates (34–36). Whether theprincipal destructive function of CD8 T cells during allograftrejection is achieved via secretion of pro-inflammatory cyto-
kine(s) (37) or via cytotoxicity (38) is unclear. The recentreport of adult islet allograft rejection in the absence of bothperforin/granzyme and Fas/FasL cytolytic pathways of de-struction (39) suggests that one or more cytokine(s) or in-flammatory mediator(s) may be responsible for mediatinggraft injury. The rejection of adult islet allografts in IL-2gene knockout mice has clearly indicated that IL-2 is notnecessary for allograft damage (40). IL-10 that is predomi-nantly regulated by CD8 T cells in our proislet allograftmodel and that has been shown to exhibit pro-inflammatoryproperties (41) and to enhance CD8 T cell proliferation/cytotoxicity (42) may therefore represent an important me-diator of proislet allograft rejection.
The involvement of both CD4 and CD8 T cells in therejection of proislet allografts is consistent with the knownmechanism of activation for alloreactive T cells. Alloantigens(class I and class II MHC) expressed on donor antigen-pre-senting cells (APCs) (passively carried within the islet tissue)are immunogenic by virtue of the co-expression of murineco-stimulatory ligand B7 (particularly B7–2) (43) and thuscan directly stimulate alloreactive CD8 T cells and CD4 Tcells, i.e., recipient APCs are not required in the allotrans-plant model (44). The individual contribution of these T cellsubpopulations to an allo-rejection reaction will then dependon the expression of donor class I and class II MHC alloan-tigens on graft parenchymal cells; the participation of CD4 Tcells in the rejection of proislet allografts is attributable torecognition of class II MHC alloantigens on associated ductepithelium (3). In contrast, the distinct role for only CD4 Tcells in the rejection of pancreatic islet tissue xenografts hasbeen correlated with the need for xenoantigens to be pro-cessed by recipient-type APCs and presented in the context ofrecipient class II MHC, i.e., thereby generating a class IIMHC-restricted CD4 T cell response (45, 46). Donor andrecipient APCs therefore appear to play a critical role inregulating the involvement of different T cell subpopulationsin the rejection of proislet allografts and xenografts. In vitro(47–49) and in vivo (50, 51) studies have also provided evi-dence supporting a contributing role for APCs in determiningthe Th1 and Th2 cytokine responses secreted by antigen-reactive T cells. It is possible that the preferential inductionof Th1 and Th2 type responses to proislet allografts andxenografts, respectively, may be due to a bias in the func-tional capacity of different APC populations to present en-dogenous MHC alloantigens and exogenous MHC xenoanti-gens and/or to signal activation of Th-specific cytokinetranscription factors in the antigen-responsive T cell (52). Itis noteworthy that dendritic cells are not only potent stimu-lators of alloreactive T cells in vitro (53) and in vivo (54) butalso potent producers of IL-12 and hence promote Th1 cyto-kine responses (55, 56) ; in addition, mature dendritic cellsare much less efficient at processing exogenous protein anti-gen(s) than macrophages (56, 57). These properties lend sup-port to the speculation that donor dendritic cells may be thepreferred APC for stimulating allo-rejection (via presenta-tion of endogenous major histocompatibility complex pep-tide); macrophages however are more capable of efficientlyprocessing unrefined proteins such as exogenous graft-derived xenoantigens. In the context of transplantation, therole of different APC populations in alloantigen versus xeno-antigen processing/presentation and in influencing distinct
SIMEONOVIC ET AL.April 15, 1999 969

intragraft cytokine profiles will require further investigationand clarification.
Analyses of the contribution of individual T cell-dependentcytokines to the rejection of proislet xenografts (20) and al-lografts8 have failed to identify a single cytokine that regu-lates either of these rejection reactions. Although both rejec-tion processes are T cell dependent, evidence argues againstdirect T cell-mediated cytotoxicity as the mechanism of de-struction of allogeneic tissue (39); in addition the dependenceof proislet xenograft rejection on CD4 T cells further suggeststhat cytokines or inflammatory mediators are required. It ispossible that tissue injury is achieved via a cascade of cyto-kines or via the cooperative involvement of cytokines andinflammatory mediators. Our study nevertheless suggeststhat different anti-rejection strategies may need to be devel-oped to optimally target the allograft and xenograft re-sponses.
The authors thank Debra Brown and Katja McKenzie for experttechnical assistance and Dr Simon Bain for the large animal surgery.We also thank Julie Macklin, Stuart Butterworth and Marc Fenningfor the photography.
REFERENCES
1. Hao L, Wang Y, Gill RG, Lafferty KJ. Role of L3T41 T cell inallograft rejection. J Immunol 1987; 139: 4022.
2. Yamamato H, Monden M, Kawai M, et al. The role of CD81 andCD41 cells in islet allograft rejection. Transplantation 1990;50: 120.
3. Simeonovic CJ, Brown DJ, Townsend MJ, Wilson JD. Differ-ences in the contribution of CD41 T cells to proislet and isletallograft rejection correlate with constitutive class II MHCalloantigen expression. Cell Transplant 1996; 5: 525.
4. Warren HS, Simeonovic CJ, Dixon JE, Pembrey, RG, LaffertyKJ. Sensitized Lyt-21 T cells trigger rejection of grafts express-ing class I major histocompatibility complex alloantigens.Transplant Proc 1986; 18: 310.
5. Wilson JD, Simeonovic CJ, Ting JHL, Ceredig R. Role of CD41
T-lymphocytes in rejection by mice of fetal pig proislet xeno-grafts. Diabetes 1989; 38(suppl 1): 217.
6. Simeonovic CJ, Wilson JD, Ceredig R. Antibody-induced rejec-tion of pig proislet xenografts in CD41 T cell-depleted diabeticmice. Transplantation 1990; 50: 657.
7. Morris CF, Simeonovic CJ, Fung M-C, Wilson JD, Hapel AJ. In-tragraft expression of cytokine transcripts during pig proisletxenograft rejection and tolerance in mice. J Immunol 1995;154: 2470.
8. Desai NM, Bassiri H, Kim J, et al. Islet allograft, islet xenograft,and skin allograft survival in CD81 T lymphocyte-deficientmice. Transplantation 1993; 55: 718.
9. Mosmann TR, Schumacher JH, Street NF, et al. 1991. Diversityof cytokine synthesis and function of mouse CD41 T cells.Immunol Rev 1991; 123: 209.
10. O’Connell PJ, Pacheco-Silva A, Nickerson PW, et al. Unmodifiedpancreatic islet allograft rejection results in the preferentialexpression of certain T cell activation transcripts. J.Immunol1993; 150: 1093.
11. Simeonovic CJ, Lafferty KJ. Immunogenicity of mouse fetal pan-creas and proislets. A comparison. Transplantation 1988; 45:824.
12. Simeonovic CJ, Brown DJ, Teittinen KUS, Wilson JD. Prepara-tion and transplantation of fetal proislets. In: Ricordi C, ed.
Pancreatic islet cell transplantation. Austin: RG Landes Co.,1992: 238.
13. Kirkman RL, Colvin RB, Flye MW, et al. Transplantation inminiature swine VI.: factors influencing survival of renal allo-grafts. Transplantation 1979; 28: 18.
14. Simeonovic CJ, Ceredig R, Wilson JD. Effect of GK1.5 monoclo-nal antibody dosage on survival of pig proislet xenografts inCD41 T cell-depleted mice. Transplantation 1990; 49: 849.
15. Dialynas DP, Quan ZS, Wall KA, et al. Characterisation of themurine T cell surface molecule designated L3T4, identified bymonoclonal antibody GK1.5: similarity of L3T4 to the humanLeu-3/T4 molecule. J Immunol 1983; 131: 2445.
16. Hogarth PM, Edwards J, McKenzie IFC, Goding JW, Liew FY.Monoclonal antibodies to the murine Ly-2.1 cell surface anti-gen. Immunology 1982; 46: 135.
17. Pierres A, Naquet P, Van Agthoven A, et al. A rat anti-mouse T4monoclonal antibody (H129.19) inhibits the proliferation of Ia-reactive T cell clones and delineates two phenotypically dis-tinct (T41, Lyt-2,3- and T4-, Lyt-2,31) subsets among anti-Iacytolytic T cell clones. J Immunol 1984; 132: 2775.
18. Ledbetter JA, Herzenberg LA. Xenogeneic monoclonal antibod-ies to mouse lymphoid differentiation antigens. Immunol Rev1979; 47: 63.
19. Warren HS, Hodder MJ, Allan W, Hume DA, Simeonovic CJ. In-duction of class II major histocompatibility antigens on thy-roid, adult pancreatic islet and fetal proislet allografts. Trans-plantation 1992; 53: 834 .
20. Simeonovic CJ, Townsend MJ, Wilson JD, et al. Eosinophils arenot required for the rejection of neovascularised fetal pig prois-let xenografts in mice. J Immunol 1997; 158: 2490.
21. Chirgwin JM, Przybyla AE, MacDonald RJ, Rutter WJ. Isolationof biologically active ribonucleic acid from sources enriched inribonuclease. Biochemistry 1979; 18: 5294 .
22. Montgomery RA, Dallman MJ. Analysis of cytokine expressionduring fetal thymic ontogeny using the polymerase chain re-action. J Immunol 1991; 147: 554.
23. Ralph P, Moore MAS, Nilsson K. Lysozyme synthesis by estab-lished human and murine histiocytic lymphoma cell lines.J Exp.Med 1976; 143: 1528.
24. Kaye J, Porchelli S, Tite J, Jones B, Janeway CA Jr. Both amonoclonal antibody and antisera specific for determinantsunique to individual cloned helper T cell lines can substitutefor antigen and antigen-presenting cells in the activation of Tcells. J Exp Med 1983; 158: 836.
25. Wren SM, Wang SC, Thai NL, et al. Evidence for early Th2 T cellpredominance in xenoreacivity. Transplantation 1993; 56: 905.
26. Auchincloss H Jr. Xenogeneic transplantation. Transplantation1988; 46: 1.
27. Benda B, Karlsson-Parra A, Ridderstad A, Korsgren O. Xeno-graft rejection of porcine islet-like cell clusters in immunoglob-ulin- or Fc-receptor g-deficient mice. Transplantation 1996; 62:1207.
28. Simeonovic CJ, McKenzie KUS, Wilson JD, Zarb JC, HodgkinPD. Role of anti-donor antibody in the rejection of pig proisletxenografts in mice. Xenotransplantation 1998; 5: 18.
29. Benfield MR, Witson JC, Alter BJ, Bach FH. Murine APC acti-vation in the xenogeneic MLC. Scand J Immunol 1993; 38: 130.
30. Cunningham AC, Butler TJ, Kirby JA. Demonstration of directxenorecognition of porcine cells by human cytotoxic T lympho-cytes. Immunology 1994; 81: 268.
31. Weir MR, Hall-Craggs M, Shen SY, et al. The prognostic value ofthe eosinophil in acute renal allograft rejection. Transplanta-tion 1986; 41: 709.
32. Foster PF, Sankary HN, Hart M, Ashmann M, Williams JW.Blood and graft eosinophilia as predictors of rejection in hu-man liver transplantation. Transplantation 1989; 47: 72.
33. Martinez OM, Ascher NL, Ferrell L, et al. Evidence for a non-classical pathway of graft rejection involving interleukin 5 and
8 Simeonovic CJ, Townsend MJ, Karupiah G, et al. Analysis of theTh1/Th2 paradigm in transplantation: Interferon-g-deficiency con-verts Th1-type proislet allograft rejection to a Th2-type xenograft-like response. Cell Transplant, in press.
TRANSPLANTATION970 Vol. 67, No. 7

eosinophils. Transplantation 1993; 55: 909.34. Corrigan CJ, Kay AB. T cells and eosinophils in the pathogenesis
of asthma. Immunol Today 1992; 13: 501.35. Spry CJF, Kay AB, Gleich G. J Eosinophils. Immunol Today
1992; 13: 384.36. Lopez AF, Sanderson CJ, Gamble JR, et al. Recombinant human
interleukin 5 is a selective activator of human eosinophil func-tion. J Exp Med 1988; 167: 219.
37. Sad S, Marcotte R, Mosmann TR. Cytokine differentiation ofprecursor mouse CD81 T cells into cytotoxic CD81 T cellssecreting Th1 or Th2 cytokines. Immunity 1995; 2: 271.
38. Hao L, Wang Y, Gill RG, et al. Role of lymphokine in isletallograft rejection. Transplantation 1990; 49: 609.
39. Ahmed KR, Guo TB, Gaal KK. Islet rejection in perforin-defi-cient mice. The role of perforin and Fas. Transplantation 1997;63: 951.
40. Steiger J, Nickerson PW, Steurer, W, Moscovitch-Lopatin M,Strom TB. IL-2 knockout recipient mice reject islet cell allo-grafts. J Immunol1995; 155: 489.
41. Wogensen L, Xiaojian H, Sarvetnick N. Leukocyte extravasationinto the pancreatic tissue in transgenic mice expressing inter-leukin 10 in the islets of Langerhans. J Exp Med 1993; 178:175.
42. Chen W- F, Zlotnick A. IL-10: a novel cytotoxic T cell differenti-ation factor. J Immunol 1991; 147: 528.
43. Lenschow DJ, Zeng Y, Hathcock KS, et al. Inhibition of trans-plant rejection following treatment with anti-B7–2 and anti-B7–1 antibodies. Transplantation 1995; 60: 1171.
44. Wolf LA, Coulombe M, Gill RG. Donor antigen-presenting cell-independent rejection of islet xenografts. Transplantation1995; 60: 1164.
45. Moses RD, Pierson RNI, Winn HJ, Auchincloss HJ. Xenogeneicproliferation and lymphokine production are dependent onCD41 T helper cells and self antigen-presenting cells in themouse. J Exp Med 1990; 172: 567.
46. Yoshizawa K, Yano A. Mouse T lymphocyte proliferative re-sponses specific for human MHC products in mouse anti-hu-man xenogeneic MLR. J Immunol 1984; 132: 2820.
47. Chang T-L, Shea CM, Urioste S, Thompson RC, Boom WH,
Abbas AK. Heterogeneity of helper/inducer T lymphocytes. III.Responses of IL-2- and IL-4- producing (Th1 and Th2) clones toantigens presented by different accessory cells. J Immunol1990; 145: 2803.
48. Gajewski TF, Pinnas M, Wong T, Fitch FW. Murine Th1 and Th2clones proliferate optimally in response to distinct antigen-presenting cell populations. J Immunol 1991; 146: 1750.
49. Schmitz J, Assenmacher M, Radbruch A. Regulation of T helpercell cytokine expression: functional dichotomy of antigen-pre-senting cells. Eur J Immunol 1993; 23: 191.
50. Doherty TM, Coffman RL. Leishmania antigens presented byGM-CSF-derived macrophages protect susceptible miceagainst challenge with Leishmania major. J Immunol 1993;150: 5476.
51. Sacks DL, Scott PA, Asofsky R, Sher AF. Cutaneous leishman-iasis in anti-IgM- treated mice: enhanced resistance due tofunctional depletion of a B cell-dependent T cell involved in thesuppressor pathway. J Immunol 1984; 132: 2072.
52. Lederer JA, Perez VL, DesRoches L, Kim SM, Abbas AK, Lich-tman AH. Cytokine transcriptional events during helper T cellsubset differentiation. J Exp Med 1996; 184: 397.
53. Steinman RM, Gutchinov B, Witmer MD, Nussenzweig MC.Dendritic cells are the principal stimulators of the primarymixed leukocyte reaction in mice. J Exp Med 1983; 157: 613.
54. Setum CM, Hegre OD, Serie JR, Moore WV. The potency ofsplenic dendritic cells as alloantigen presenters in vivo. Trans-plantation 1990; 49: 1175.
55. Heufler C, Koch F, Stanzl U, et al. Interleukin- 12 is produced bydendritic cells and mediates T helper 1 development as well asinterferon-g production by T helper cells. Eur J Immunol 1996;26: 659.
56. Winzler C, Rovere P, Rescigno M, et al. Maturation stages ofmouse dendritic cells in growth factor-dependent long-termcultures. J Exp Med 1997; 185: 317.
57. Austyn JM. Lymphoid dendritic cells. Immunology 1987; 62:161.
Received 16 December 1997.Accepted 21 October 1998.
SIMEONOVIC ET AL.April 15, 1999 971




















