
Liposomes enhance delivery and expression of an RGD-oligolysine gene transfer vector in human...
11
Gene Therapy (1998) 5, 1488–1498 1998 Stockton Press All rights reserved 0969-7128/98 $12.00 http://www.stockton-press.co.uk/gt Liposomes enhance delivery and expression of an RGD-oligolysine gene transfer vector in human tracheal cells M Colin 1 , RP Harbottle 2 , A Knight 2 , M Kornprobst 1 , RG Cooper 3 , AD Miller 3 , G Trugnan 4 , J Capeau 1 , C Coutelle 2 and MC Brahimi-Horn 1 1 Institut National de la Sante ´ et de la Recherche Me ´dicale U402, Faculte ´ de Me ´decine Saint-Antoine, Paris, France; 2 Molecular Genetics, Division of Biomedical Sciences, Imperial College, School of Medicine at St Mary’s, London; 3 Department of Chemistry, Imperial College, London, UK; and 4 Institut National de la Sante ´ et de la Recherche Me ´dicale CJF 96.07, Faculte ´ de Me ´decine Saint-Antoine, Paris, France Nonviral gene delivery systems consist predominantly of Expression was further enhanced 30-fold (P , 0.01) with lipoplexes or receptor-targeting and nontargeting poly- lipofectamine and the ratio of DNA/peptide/lipofectamine plexes. We examined integrin-mediated gene delivery was critical for specificity and expression. Fluorescence using an Arg-Gly-Asp/oligo-L-lysine ([K] 16 RGD) cyclic pep- and radioactive labelling of the complex showed that the tide and investigated its gene transfer efficiency when [K] 16 RGD peptide increased the endocytic uptake of DNA associated with a cationic liposome. We demonstrated that into cells. The cell association of both DNA and peptide human cystic fibrosis and noncystic fibrosis tracheal epi- increased even further with lipofectamine. Confocal thelial cells in culture express integrins that recognise the microscopy showed that the [K] 16 RGD peptide and the RGD integrin-binding motif. We found a 10-fold (P , 0.01) DNA internalised together within 30 min and localised to increased expression of a luciferase encoding plasmid in vesicles in the perinuclear region. These results show that these cells when complexing the plasmid to the [K] 16 RGD an integrin-binding ligand can deliver genetic material to peptide as compared with plasmid alone. This increase airway cells and that a cationic liposome can enhance the was specific to the [K] 16 RGD peptide since neither a efficacy of this nonviral vector system. [K] 16 RGE nor a [K] 16 peptide gave a comparable increase. Keywords: cystic fibrosis; gene therapy; integrin; liposome; oligo-L-lysine; RGD Introduction Gene therapy aims to introduce corrective genetic material into cells to relieve the symptoms of disease. A variety of diseases are being considered for application of gene transfer technology, in particular inherited dis- orders such as cystic fibrosis (for review see Refs 1 and 2). Nonviral systems for delivery of DNA, although less efficient than viral systems, provoke less host inflamma- tory and immune responses. They consist mainly of cat- ionic liposome–DNA complexes (lipoplexes) or receptor- targeting or nontargeting cationic polymer–DNA com- plexes (polyplexes). Cationic liposomes bind DNA by means of ionic interactions and facilitate DNA delivery by fusion with the plasma membrane, 3 and/or endo- cytosis. 4–7 Such vectors are easy to prepare and have lim- ited toxicity, 8 but achieve relatively low transfection lev- els. Receptor-targeting DNA delivery constructs consist mainly of a receptor-binding ligand, for example trans- ferrin or asialo-orosomucoid, conjugated to a DNA-bind- Correspondence: MC Brahimi-Horn, INSERM U402, Faculte ´ de Medicine Saint-Antoine, 27 rue Chaligny, 75571 Paris Cedex 12, France Received 9 April 1998; accepted 19 June 1998 ing moiety, usually high molecular weight oligo-l-lysine (for review see Refs 9 and 10). We have previously demonstrated the principal of inte- grin-mediated gene delivery using a bifunctional peptide. This molecule is comprised of an integrin receptor-tar- geting domain containing an arginine-glycine-aspartic acid tripeptide motif and a DNA binding moiety consisting of a stretch of 16 lysine residues, Arg-Gly- Asp/oligo-l-lysine ([K] 16 RGD). 11,12 We showed that this peptide condensed plasmid DNA (approximately 10 nm) rendering it resistant to nuclease degradation and that the level of reporter gene expression obtained in Caco- 2 cells under optimal conditions approached that of the commercial liposome lipofectamine. 12 Integrins are a class of related heterodimeric transmembrane surface receptors involved in cell–cell adhesion and in promotion of interactions between cells and components of the extracellular matrix (for review see Ref. 13). They are able to bind to a wide variety of ligands including extracellu- lar matrix glycoproteins such as fibronectin and vitronec- tin, while their intracellular domains interact with the cytoskeleton. Integrins are natural targets for receptor- mediated cell entry used by a variety of pathogenic micro-organisms, including bacteria 14 and viruses. 15,16 In particular, the adenovirus, a candidate for viral gene
Transcript of Liposomes enhance delivery and expression of an RGD-oligolysine gene transfer vector in human...

Gene Therapy (1998) 5, 1488–1498 1998 Stockton Press All rights reserved 0969-7128/98 $12.00
http://www.stockton-press.co.uk/gt
Liposomes enhance delivery and expression of anRGD-oligolysine gene transfer vector in humantracheal cells
M Colin1, RP Harbottle2, A Knight2, M Kornprobst1, RG Cooper3, AD Miller3, G Trugnan4,J Capeau1, C Coutelle2 and MC Brahimi-Horn1
1Institut National de la Sante et de la Recherche Medicale U402, Faculte de Medecine Saint-Antoine, Paris, France; 2MolecularGenetics, Division of Biomedical Sciences, Imperial College, School of Medicine at St Mary’s, London; 3Department of Chemistry,Imperial College, London, UK; and 4Institut National de la Sante et de la Recherche Medicale CJF 96.07, Faculte de MedecineSaint-Antoine, Paris, France
Nonviral gene delivery systems consist predominantly of Expression was further enhanced 30-fold (P , 0.01) withlipoplexes or receptor-targeting and nontargeting poly- lipofectamine and the ratio of DNA/peptide/lipofectamineplexes. We examined integrin-mediated gene delivery was critical for specificity and expression. Fluorescenceusing an Arg-Gly-Asp/oligo-L-lysine ([K]16RGD) cyclic pep- and radioactive labelling of the complex showed that thetide and investigated its gene transfer efficiency when [K]16RGD peptide increased the endocytic uptake of DNAassociated with a cationic liposome. We demonstrated that into cells. The cell association of both DNA and peptidehuman cystic fibrosis and noncystic fibrosis tracheal epi- increased even further with lipofectamine. Confocalthelial cells in culture express integrins that recognise the microscopy showed that the [K]16RGD peptide and theRGD integrin-binding motif. We found a 10-fold (P , 0.01) DNA internalised together within 30 min and localised toincreased expression of a luciferase encoding plasmid in vesicles in the perinuclear region. These results show thatthese cells when complexing the plasmid to the [K]16RGD an integrin-binding ligand can deliver genetic material topeptide as compared with plasmid alone. This increase airway cells and that a cationic liposome can enhance thewas specific to the [K]16RGD peptide since neither a efficacy of this nonviral vector system.[K]16RGE nor a [K]16 peptide gave a comparable increase.
Keywords: cystic fibrosis; gene therapy; integrin; liposome; oligo-L-lysine; RGD
IntroductionGene therapy aims to introduce corrective geneticmaterial into cells to relieve the symptoms of disease. Avariety of diseases are being considered for applicationof gene transfer technology, in particular inherited dis-orders such as cystic fibrosis (for review see Refs 1 and2). Nonviral systems for delivery of DNA, although lessefficient than viral systems, provoke less host inflamma-tory and immune responses. They consist mainly of cat-ionic liposome–DNA complexes (lipoplexes) or receptor-targeting or nontargeting cationic polymer–DNA com-plexes (polyplexes). Cationic liposomes bind DNA bymeans of ionic interactions and facilitate DNA deliveryby fusion with the plasma membrane,3 and/or endo-cytosis.4–7 Such vectors are easy to prepare and have lim-ited toxicity,8 but achieve relatively low transfection lev-els. Receptor-targeting DNA delivery constructs consistmainly of a receptor-binding ligand, for example trans-ferrin or asialo-orosomucoid, conjugated to a DNA-bind-
Correspondence: MC Brahimi-Horn, INSERM U402, Faculte de MedicineSaint-Antoine, 27 rue Chaligny, 75571 Paris Cedex 12, FranceReceived 9 April 1998; accepted 19 June 1998
ing moiety, usually high molecular weight oligo-l-lysine(for review see Refs 9 and 10).
We have previously demonstrated the principal of inte-grin-mediated gene delivery using a bifunctional peptide.This molecule is comprised of an integrin receptor-tar-geting domain containing an arginine-glycine-asparticacid tripeptide motif and a DNA binding moietyconsisting of a stretch of 16 lysine residues, Arg-Gly-Asp/oligo-l-lysine ([K]16RGD).11,12 We showed that thispeptide condensed plasmid DNA (approximately 10 nm)rendering it resistant to nuclease degradation and thatthe level of reporter gene expression obtained in Caco-2 cells under optimal conditions approached that of thecommercial liposome lipofectamine.12 Integrins are aclass of related heterodimeric transmembrane surfacereceptors involved in cell–cell adhesion and in promotionof interactions between cells and components of theextracellular matrix (for review see Ref. 13). They are ableto bind to a wide variety of ligands including extracellu-lar matrix glycoproteins such as fibronectin and vitronec-tin, while their intracellular domains interact with thecytoskeleton. Integrins are natural targets for receptor-mediated cell entry used by a variety of pathogenicmicro-organisms, including bacteria14 and viruses.15,16 Inparticular, the adenovirus, a candidate for viral gene

RGD-mediated gene internalisation and expressionM Colin et al
1489therapy, has been shown to require RGD-binding of inte-grins for internalisation.17
While both cationic liposome and receptor-mediatedmolecular conjugates suffer from relatively low transfec-tion efficiencies the association of the two can substan-tially increase transfection as demonstrated in lung endo-thelial cells when a poly-l-lysine anti-thrombomodulinantibody conjugate and DNA where associated with acationic liposome.18 An additional study also showedenhancement of asialoglycoprotein receptor-mediatedDNA transfer into human hepatoma HepG2 cells in thepresence of the lipid lipofectam.19 Hela cells transfectedwith the ligand transferrin also showed enhanced trans-fection with the liposome lipofectin.20 We recentlyreported first results demonstrating enhancement of inte-grin-mediated gene delivery by addition of a liposome tomelanoma, kidney and endothelial cell lines.21
In the present report, we extend our study on integrin-mediated gene delivery to non-cystic fibrosis and cysticfibrosis human tracheal cells and show that the combi-nation of the [K]16RGD peptide with the liposome lipofec-tamine increases the level of transgene expression about30-fold (P , 0.01) above that obtained with the peptidealone. Radioactive and fluorescence labelling studies toexamine the cellular delivery of the peptide and DNAcomponents in the presence and absence of lipofectaminedemonstrate that lipofectamine improves the efficiency ofuptake of the complexes with a certain degree of targetspecificity by which the RGD motif of the complex bindsto integrins on the cell surface of human tracheal epi-thelial cells. However, lipofectamine also increases non-specific DNA delivery. Lipofectamine does not seem tohave a major influence on endosomal location since con-focal microscopy shows rapid accumulation of the com-plexes in perinuclear vesicles both in its presence andabsence.
Results
Competitive inhibition of cell attachment to the integrin-binding domain of fibronectin by an RGD peptide andimmuno-detection of the b1-integrin subunitThe integrin targeting sequence RGDMFG used in our[K]16RGD ligand is known to have a high affinity forcellular integrin receptors. Cells normally attach andspread on cell culture wells coated with fibronectin (FN)or vitronectin (VN) through binding of their integrins tothe RGD-motif of FN and VN. We demonstrated pre-viously by competitive cell binding studies with the[K]16RGD peptide that the RGD domain in our constructmaintained its affinity for these integrins after the syn-thetic fusion with the highly positively charged oligo-l-lysine chain.12 These experiments were performed on theCaco-2 intestinal cell line which our group recentlyshowed to express the b1-integrin subunit.22 Since thepresent study uses tracheal epithelial cells with the long-term aim of correcting the cystic fibrosis defect in airwayepithelium, we verified that this cell line also demon-strates binding affinity for the RGD domain of the[K]16RGD peptide. The cells do not attach to wells coatedwith the control protein BSA, but attach to wells coatedwith FN; each in the absence of competitive ligands. Pre-incubation of the cell suspensions with [K]16RGDdecreases significantly the degree of attachment to FN
while preincubation with [K]16RGE does not modifyattachment. A similar pattern of results is obtained forthe Caco-2 cell line. In addition, VN gives similar resultsto those of FN. These results demonstrate that the RGDsequence of [K]16RGD competes with FN or VN forattachment of the epithelial 56FHTE8o− cell line andsuggests that these cells express integrins which recog-nise the RGD motif. In addition, we confirmed by fluor-escence flow cytometry that the tracheal cell line56FHTE8o− expresses the b1-integrin subunit. Controlcells incubated with the secondary FITC-labelled anti-body alone showed a low level of fluorescence while cellsincubated with a b1-anti-integrin antibody and then thesecondary antibody showed increased fluorescence. Simi-lar results for cell attachment and immunodetection werealso obtained with another non-cystic fibrosis epithelialtracheal cell line (NT-1) and with two other epithelial tra-cheal cystic fibrosis cell lines (CFT-1) and (CFT-2) (resultsare not shown).
Peptide/DNA complex mediated transgene expressionenhanced with lipofectamineOligo-l-lysine binds to DNA and retards its migrationduring electrophoresis in an agarose gel.12 By addition ofincreasing amounts of peptide to a constant amount ofplasmid DNA, we found that 0.48 nmol of either the[K]16RGD or [K]16RGE peptide (1.9 mg peptide/mg DNA)or of the oligo-l-lysine peptide, [K]16, are sufficient tofully retard DNA migration (not shown).
Transfection of tracheal cells with the [K]16RGD pep-tide induces a steep increase of luciferase gene expressionfrom its two-fold retardation value (0.96 nmol)(Figure 1a) to reach about 10-fold (P , 0.01) theexpression level of the [K]16RGE control peptide at three-fold retardation (1.44 nmol). [K]16RGE gave only very lowlevels of luciferase gene expression (Figure 1a) which cor-respond to those obtained with naked DNA and DNAcomplexed with [K]16 peptide respectively (not shown).The presence of lipofectamine (24 mg/mg DNA)(Figure 1b) enhances the luciferase activity steadily withincreasing amounts of peptide to attain a maximum atthree times the retardation amount of peptide (1.44 nmol)but with no obvious difference between [K]16RGD and[K]16RGE. Lipofectamine also increases the level ofexpression mediated by naked DNA and DNA com-plexed with the control peptide [K]16 (result not shown)by about 50-fold (P , 0.01). The [K]16RGD/DNA/lipo-some-mediated expression is about three orders of mag-nitude above the value observed with the optimal[K]16RGD/DNA ratio without liposome and is about six-fold (P , 0.01) higher than that of [K]16/DNA in the pres-ence of lipofectamine (result not shown). At this liposomeratio (24 mg/mg DNA) it appears that lipofectaminemasks the RGD sequence making it less accessible to cellsurface integrins and that transfection is mainly drivenby the nonspecific mechanism of liposome-mediatedcell entry.
Similar results were also obtained with the other cysticfibrosis and non-cystic fibrosis tracheal cell lines (resultsnot shown).
The obvious next experiment was therefore to try torestore the targeting specificity of the [K]16RGD peptideby reducing the lipid to complex ratio. At a ratio of 1:5:18of DNA/[K]16RGD peptide/lipofectamine, we obtainedoptimal activity whilst retaining about 50% (P , 0.05) of

RGD-mediated gene internalisation and expressionM Colin et al
1490
Figure 1 Peptide at three-fold the retardation amount (1.44 nmol) showsmaximal luciferase gene expression with or without lipofectamine. Lucifer-ase gene expression in tracheal cells with the indicated amount of peptidewas examined in the absence (a) and presence (b) of lipofectamine. Resultsare expressed as relative light units (RLU) per milligramme cell proteinper 60 s.
the targeting specificity (Figure 2). This result suggeststhat in the presence of lipofectamine, internalisation ofthe complex is occurring by a mechanism which is inde-pendent of integrins.
In conclusion, the [K]16RGD peptide not only increasesthe expression level compared with plasmid DNA by itsspecific targeting effect, it also enhances the lipofectamineactivity nonspecifically as does [K]16RGE.
The RGD peptide and lipofectamine increase uptake of[35S]-labelled plasmid DNA into cellsTo investigate the kinetics of binding and uptake of DNAcomplexed to peptide and/or lipofectamine, we labelledthe plasmid DNA with dATP-[35S]. After incubation ateither 4°C or 37°C with the labelled DNA, the cells wererinsed and detached with trypsin/EDTA. In the absence
Figure 2 The amount of lipofectamine influences the expression level andthe specificity of [K]16RGD. DNA complexed with either [K]16RGD or[K]16RGE and lipofectamine was added to semiconfluent tracheal epithelialcells in OptiMEM at the indicated DNA/peptide/lipofectamine ratios.After 4 h the cells were rinsed and a further 48 h later the luciferaseactivity was determined.
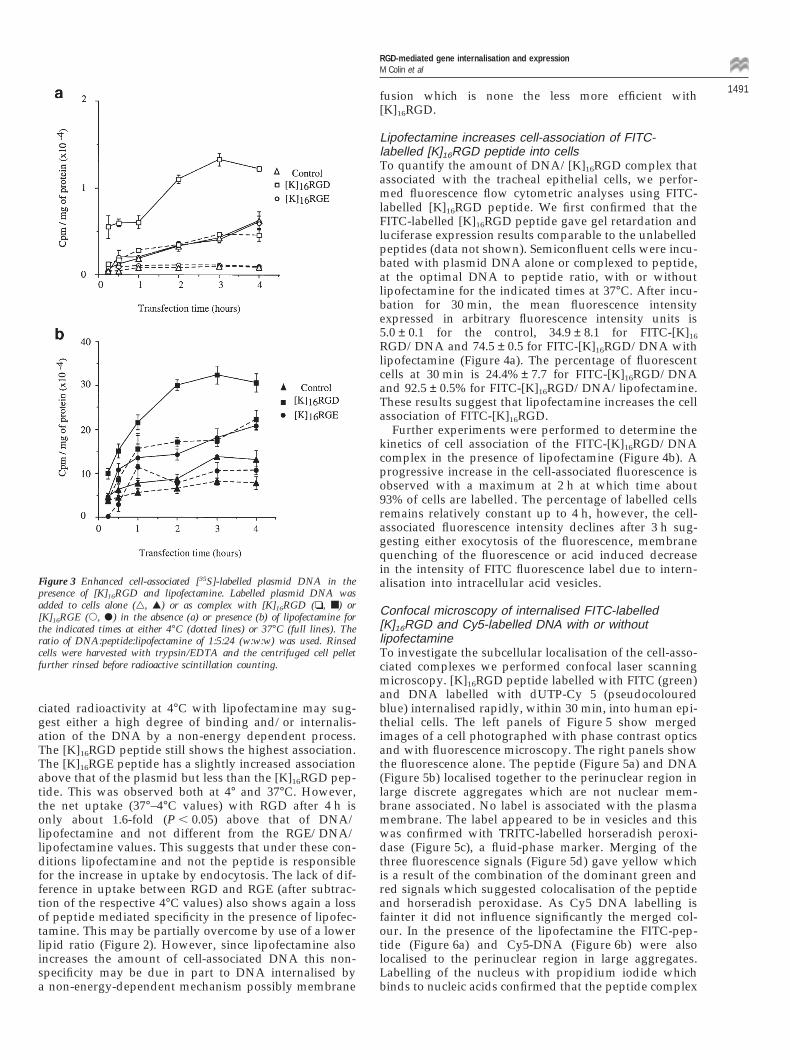
of lipofectamine cell-associated radioactivity at 37°Cwould include bound and internalised DNA. Becauseclassical endocytosis, an energy-dependent mechanism ofcell entry, is blocked at 4°C the cell-associated radio-activity would be limited to cell bound material or radio-active macromolecules that enter by a non-endocytic,possibly fusion, mechanism. Thus the difference betweenthe cell-associated radioactivity at 37°C and 4°C rep-resents uptake by endocytosis.
At 4°C only a constant low level of radioactivity is cell-associated with plasmid alone and plasmid complexed tothe [K]16RGE peptide whilst cell binding is increased withthe [K]16RGD peptide (Figure 3a). [K]16RGE would there-fore represent nonspecific electrostatic binding. Theincreased values for [K]16RGD/DNA suggest that theDNA is at least in part binding by means of integrinsthat recognise the RGD-binding domain. At 37°C the cell-associated [35S]-labelled plasmid DNA complexed to the[K]16RGD peptide increases with the transfection timeand reaches a plateau at around 2 to 3 h. The uptake (37°–4°C values) of labelled DNA was highest for the[K]16RGD/DNA complex while that for the [K]16
RGE/DNA complex is equivalent to plasmid alone(Figure 3a). Thus in the absence of lipofectamine the[K]16RGD/DNA complex enters the cells by means of areceptor-mediated endocytic pathway. A lower level ofendocytosis occurs for the [K]16RGE/DNA complex andis equivalent to that of DNA alone.
In the presence of lipofectamine the amount of cell-associated radioactivity increases rapidly with time andis higher for all combinations at both 4° and 37°C thanin the absence of lipofectamine (Figure 3b). After 3 to 4 hthe increase at 4°C is more than 40-fold (P , 0.02) for the[K]16RGD peptide. The relatively high level of cell-asso-
RGD-mediated gene internalisation and expressionM Colin et al
1491
Figure 3 Enhanced cell-associated [35S]-labelled plasmid DNA in thepresence of [K]16RGD and lipofectamine. Labelled plasmid DNA wasadded to cells alone (g, G) or as complex with [K]16RGD (❏, ■) or[K]16RGE (s, P) in the absence (a) or presence (b) of lipofectamine forthe indicated times at either 4°C (dotted lines) or 37°C (full lines). Theratio of DNA:peptide:lipofectamine of 1:5:24 (w:w:w) was used. Rinsedcells were harvested with trypsin/EDTA and the centrifuged cell pelletfurther rinsed before radioactive scintillation counting.
ciated radioactivity at 4°C with lipofectamine may sug-gest either a high degree of binding and/or internalis-ation of the DNA by a non-energy dependent process.The [K]16RGD peptide still shows the highest association.The [K]16RGE peptide has a slightly increased associationabove that of the plasmid but less than the [K]16RGD pep-tide. This was observed both at 4° and 37°C. However,the net uptake (37°–4°C values) with RGD after 4 h isonly about 1.6-fold (P , 0.05) above that of DNA/lipofectamine and not different from the RGE/DNA/lipofectamine values. This suggests that under these con-ditions lipofectamine and not the peptide is responsiblefor the increase in uptake by endocytosis. The lack of dif-ference in uptake between RGD and RGE (after subtrac-tion of the respective 4°C values) also shows again a lossof peptide mediated specificity in the presence of lipofec-tamine. This may be partially overcome by use of a lowerlipid ratio (Figure 2). However, since lipofectamine alsoincreases the amount of cell-associated DNA this non-specificity may be due in part to DNA internalised bya non-energy-dependent mechanism possibly membrane
fusion which is none the less more efficient with[K]16RGD.
Lipofectamine increases cell-association of FITC-labelled [K]16RGD peptide into cellsTo quantify the amount of DNA/[K]16RGD complex thatassociated with the tracheal epithelial cells, we perfor-med fluorescence flow cytometric analyses using FITC-labelled [K]16RGD peptide. We first confirmed that theFITC-labelled [K]16RGD peptide gave gel retardation andluciferase expression results comparable to the unlabelledpeptides (data not shown). Semiconfluent cells were incu-bated with plasmid DNA alone or complexed to peptide,at the optimal DNA to peptide ratio, with or withoutlipofectamine for the indicated times at 37°C. After incu-bation for 30 min, the mean fluorescence intensityexpressed in arbitrary fluorescence intensity units is5.0 ± 0.1 for the control, 34.9 ± 8.1 for FITC-[K]16
RGD/DNA and 74.5 ± 0.5 for FITC-[K]16RGD/DNA withlipofectamine (Figure 4a). The percentage of fluorescentcells at 30 min is 24.4% ± 7.7 for FITC-[K]16RGD/DNAand 92.5 ± 0.5% for FITC-[K]16RGD/DNA/lipofectamine.These results suggest that lipofectamine increases the cellassociation of FITC-[K]16RGD.
Further experiments were performed to determine thekinetics of cell association of the FITC-[K]16RGD/DNAcomplex in the presence of lipofectamine (Figure 4b). Aprogressive increase in the cell-associated fluorescence isobserved with a maximum at 2 h at which time about93% of cells are labelled. The percentage of labelled cellsremains relatively constant up to 4 h, however, the cell-associated fluorescence intensity declines after 3 h sug-gesting either exocytosis of the fluorescence, membranequenching of the fluorescence or acid induced decreasein the intensity of FITC fluorescence label due to intern-alisation into intracellular acid vesicles.
Confocal microscopy of internalised FITC-labelled[K]16RGD and Cy5-labelled DNA with or withoutlipofectamineTo investigate the subcellular localisation of the cell-asso-ciated complexes we performed confocal laser scanningmicroscopy. [K]16RGD peptide labelled with FITC (green)and DNA labelled with dUTP-Cy 5 (pseudocolouredblue) internalised rapidly, within 30 min, into human epi-thelial cells. The left panels of Figure 5 show mergedimages of a cell photographed with phase contrast opticsand with fluorescence microscopy. The right panels showthe fluorescence alone. The peptide (Figure 5a) and DNA(Figure 5b) localised together to the perinuclear region inlarge discrete aggregates which are not nuclear mem-brane associated. No label is associated with the plasmamembrane. The label appeared to be in vesicles and thiswas confirmed with TRITC-labelled horseradish peroxi-dase (Figure 5c), a fluid-phase marker. Merging of thethree fluorescence signals (Figure 5d) gave yellow whichis a result of the combination of the dominant green andred signals which suggested colocalisation of the peptideand horseradish peroxidase. As Cy5 DNA labelling isfainter it did not influence significantly the merged col-our. In the presence of the lipofectamine the FITC-pep-tide (Figure 6a) and Cy5-DNA (Figure 6b) were alsolocalised to the perinuclear region in large aggregates.Labelling of the nucleus with propidium iodide whichbinds to nucleic acids confirmed that the peptide complex

RGD-mediated gene internalisation and expressionM Colin et al
1492
Figure 4 Fluorescence flow cytometric analysis of the association of the FITC-labelled [K]16RGD peptide with tracheal epithelial cells. (a) Cells wereincubated for 30 min in DMEM/F-12 alone (control) or FITC-labelled [K]16RGD/DNA or FITC-labelled [K]16RGD/DNA/lipofectamine (L). (b) Kineticsof cell association of FITC-labelled [K]16RGD in the presence of lipofectamine. The ratio of DNA:peptide:lipofectamine of 1:5:18 (w:w:w) was used.
was associated with the perinuclear region (results notshown). The DNA label was more intense in the presenceof lipofectamine. This suggests that less DNA entered thecells in the absence of lipofectamine as indicated by theresults obtained with [35S]-labelled DNA (Figure 3). Inboth, the absence (Figure 5) and presence of lipofecta-mine (Figure 6) the merged images of the blue and greensignals show colocalisation (turquoise) (not shown). TheTRITC-horseradish peroxidase (Figure 6c) with complexcontaining lipofectamine also appeared to localise to thesame region as the peptide and DNA, as confirmed bythe merging of the three fluorescence labels (Figure 6d).Thus confocal microscopy demonstrates internalisation ofboth the peptide and the DNA in the absence and pres-ence of lipofectamine and localisation to vesicles, prob-
Figure 5 Confocal microscopy showing internalised fluorescent peptideand plasmid DNA in the absence of lipofectamine. Tracheal cells wereincubated with Cy5-labelled DNA/FITC-labelled [K]16RGD peptide com-plex at 37°C for 30 min. Rinsed cells were then incubated for 5 min withTRITC-horseradish peroxidase. The left panels show merged images of acentral section of the same cell photographed with phase contrast opticsand with fluorescence. The right panels show the fluorescence image alone.The FITC-labelled [K]16RGD peptide (a; green), the Cy5-labelled DNA (b;coded in blue), the TRITC-horseradish peroxidase (c; red) and the mergedfluorescence (d; green, blue and red) images are shown. The ratio of DNA:peptide was 1:5 and sections of 0.88 mm were obtained. The bar for theimages on the left panel represents 10 mm and on the right 5 mm.

RGD-mediated gene internalisation and expressionM Colin et al
1493a- a-
b- b-
c- c-
d-

RGD-mediated gene internalisation and expressionM Colin et al
1494 ably endosomes, in the perinuclear region of the cellafter 30 min.
DiscussionThe long-term aim of this work is to develop a vectorsystem that allows delivery of DNA sequences coding forthe normal cystic fibrosis transmembrane conductanceregulator gene, the gene which is defective in cysticfibrosis, to the airways in a cell-specific manner allowinghigh expression for gene therapy in vivo. Presently, bothviral and nonviral vectors for gene transfer are in devel-opment. Although viral vectors are considered to beefficient, their use may be limited by possible side-effects.Nonviral vectors are less toxic, easier to prepare and arenot limited in the size of the DNA they can deliver.Research efforts into nonviral vectors for gene therapyhave concentrated mainly on either lipoplex or on recep-tor-targeting polyplex vectors. We previously establishedthe principle of integrin-mediated gene transfer using anAsp-Gly-Arg/oligo-l-lysine ([K]16RGD) conjugate.11,12
We noted recently that the combination of these twotypes of nonviral vectors, a cationic lipid lipofectamineand an integrin-binding poly-l-lysine conjugateincreased substantially the expression level in human tra-cheal epithelial cells. Maximal expression was dependenton the amount of complex and on the ratio of DNA-to-peptide-to-lipofectamine.
An increase in delivery and expression in endothelialcells was observed when a poly-l-lysine antibody conju-gate and DNA were associated with a cationic lipo-some.18 Another study reported enhancement of asialo-glycoprotein receptor-mediated DNA transfer intohuman hepatoma HepG2 cells in the presence of the lipidlipofectam.19 Enhanced transferrin-mediated gene trans-fer in the presence of the cationic lipid lipofectin in Helacells has been reported.20 Using [35S]-labelled plasmidDNA the authors also showed enhanced intracellularDNA transfer by transferrin in the presence of lipofectin.
The enhanced effect may be due to increased uptakeof DNA, but also to protection of the DNA by the lipo-somes in the extracellular medium allowing the complexto interact with the cell surface. We found that both lipo-fectamine and peptide protects the plasmid DNA fromdegradation by DNase (not shown). Liposomes may alsoprotect the DNA by delaying its passage to lysosomesas suggested by a study which examined the subcellulardistribution of [35S]-labelled DNA in rat liver.23 Delayeddegradation may mean that there is a greater chance thatthe DNA passages to the nucleus.
The specificity of the integrin binding was demon-strated by an increase in luciferase activity with the[K]16RGD peptide over the [K]16RGE peptide in theabsence of lipofectamine. However, this specificity waslost in the presence of high ratios of lipofectamine andwas only partially restored at lower ratios. This suggeststhat the amount of lipid may modify the interaction ofthe [K]16RGD peptide with the cell surface integrinswhich we demonstrated by two independent methods tobe present on human epithelial tracheal cells. In fact thebinding affinity of vitronectin (VN) for its integrin recep-tor has been shown to be modulated by the amount andcomposition of liposomes.24 In addition, these resultssuggest that the complex is entering by a mechanism thatis not receptor-mediated. We also noted that the
[K]16RGE peptide gives higher expression than the [K]16
peptide in the presence of lipofectamine. This may beexplained by the resemblance of both the [K]16RGD and[K]16RGE peptides with a nuclear-localization signal(NLS), a cluster of positively charged [K]16 separated bya spacer from the positively charged arginine. The [K]16
peptide consists of only a single cluster of basic aminoacids and thus may not be as effective in nuclear trans-port. It may also be possible that [K]16RGE is more read-ily endocytosed by a nonspecific mechanism. The resultssuggest that the peptides play an intracellular role. Thesecharacteristics may facilitate active nuclear uptake ofcomplexes. Without lipofectamine neither [K]16RGE nor[K]16 enter the cell to a significant level, thus a nucleartargeting effect is absent and the two show the sameexpression level. These questions are being furtherinvestigated.
We also showed that lipofectamine increases the levelof cell-associated conjugate and DNA. This cell associ-ation represented predominantly endocytosed peptideRGD conjugate and DNA, as demonstrated by confocalmicroscopy, and suggested that lipofectamine facilitatesintracellular delivery of complex. The high percentage ofcells labelled with complex containing lipofectamine asdetected with fluorescence flow cytometry suggests thatcell entry in this system is not a limiting step to DNAdelivery. In addition, the confocal results showed coin-ternalisation of the peptide and DNA and localisation invesicles in the perinuclear region in both the presenceand absence of lipofectamine. RGD is a common motiffound in many extracellular matrix proteins that bind tointegrins including FN and VN. Internalisation of VN bymeans of the endosome–lysosome pathway has beenshown to be dependent on integrins25 and to be inhibitedby RGD peptides.26 Although endocytosis appears to bethe major entry route, as indicated by previous studies,4–7
this does not exclude the possibility that some of theuptake with lipofectamine is driven by an energy-inde-pendent process, probably membrane fusion, as sug-gested by the cell association of labelled DNA at 4°C andas proposed previously.3 Further studies into the endo-cytic mechanisms and intracellular trafficking of the vec-tor are proceeding.
Integrins are a family of ubiquitous proteins whichhave been shown to be present on the airways althoughthe FN and VN-binding integrins are either not or onlyexpressed at low levels in normal lung epithelia.27,28
However, their expression is increased during the woundrepair process in airway epithelia and in the injured air-ways of cystic fibrosis patients.28,29 Moreover, recombi-nant adenovirus-mediated gene transfer which is RGDdependent occurs preferentially in areas of injured epi-thelium.28 Thus the injured airway observed in cysticfibrosis may be susceptible to gene transfer with an inte-grin-mediated delivery system. In addition, cationic lipo-somes have been shown not to provoke adverse eventsin cystic fibrosis patients and to bring about a level ofgene transfer comparable to that reported for adeno-viral vectors.8
The results to date indicate that cationic liposomescoupled with integrin-mediated gene delivery enhancethe transfection efficiency of the RGD vector probably byimproving the efficiency of uptake of the complexesthrough a receptor-mediated mechanism and throughprotection of the DNA by lipofectamine and the peptide.

RGD-mediated gene internalisation and expressionM Colin et al
1495a- b-
c- d-
Figure 6 Confocal microscopy showing internalised fluorescent peptide and plasmid DNA in the presence of lipofectamine. Tracheal cells were incubatedwith Cy5-labelled DNA/FITC-labelled [K]16RGD peptide complex in the presence of lipofectamine at 37°C for 30 min. Rinsed cells were then incubatedfor 5 min with TRITC-horseradish peroxidase. The FITC-labelled [K]16RGD peptide (a; green), the Cy5-labelled DNA (b; coded in blue), the TRITC-horseradish peroxidase (c; red) and the merged fluorescence (d; green, blue and red) images are shown. The ratio of DNA:peptide:lipofectamine was1:5:18 and sections of 1.28 mm were obtained. The bar represents 2.5 mm and all images are of the same enlargement.
This nonviral vector system is a first step towards devel-opment of cell-specific targeting of the airway for genetherapy of cystic fibrosis.
Materials and methods
Peptide synthesis and characterisation of the [K]16RGDand [K]16RGE peptidesThe peptides N-[K]16GGCRGDMFGCA ([K]16RGD) andN-[K]16GGCRGEMFGCA ([K]16RGE) were synthesised onan ABI (Applied Biosystems, Perkin Elmer, Warrington,UK) solid-phase batch peptide synthesiser (model 431A)using standard fluorenylmethoxycarbonyl (FMOC)chemistry as previously described.12 Peptides were pur-ified by HPLC on a C18 preparative column and charac-terised by matrix-assisted laser desorption/ionisation-time of flight (MALDI-TOF) mass spectrometry andamino acid sequencing, as described previously.12 Thepurity of each peptide was in excess of 95%. Fluoresceinisothiocyanate (FITC)-labelled versions of [K]16RGD and[K]16RGE peptides were prepared by coupling FITC iso-mer I to solid phase resin-bound samples of both pep-tides. Labelled peptides were separated from unlabelledpeptides, after release from the resin, by HPLC asdescribed above. All peptides were cyclised before use byoxidation in HEPES buffered saline (HBS, 150 mm NaCl,10 mm Hepes, pH 7.3) and the formation of disulphidebonds monitored as previously described.12 The oligo-l-lysine [K]16 peptide was synthesised by Genosys Biotech-nologies (Cambridge, UK) and purified by HPLC on aVYDAC RP C-8 column to 98% purity.
Cell cultureA foetal human tracheal epithelial cell line 56FHTE8o−
obtained from a non-cystic fibrosis foetus was kindly pro-vided by D Gruenert.30 Another non-cystic fibrosishuman foetal tracheal cell line (NT-1) and two cysticfibrosis foetal tracheal cell lines CFT-1 and CFT-2 wereprepared and established in the same way in our labora-tory.31,32 Cells were grown in a 1:1 mixture of Dulbecco’sminimum essential medium (DMEM) and Ham’s F-12nutrient medium (DMEM/F-12) containing 50 U/mlpenicillin and 50 U/ml streptomycin (ATGC, Noisy leGrand, France) and supplemented with foetal calf serum(10% v/v) (Boehringer Mannheim, Meylan, France).Caco-2 cells were kindly provided by Dr A Zweibaum ofINSERM U178, France and cultured in DMEM (4.5 g/lglucose) supplemented with 10% foetal calf serum, 2 mml-glutamine and non-essential amino acids. Cells weremaintained at 37°C in 5% CO2 95% air and the mediumchanged every second day.
Cell attachment and inhibitionThe wells of non-tissue culture 96-well plates (TPP,Trafadingen, Switzerland) were coated either with fib-ronectin (FN), vitronectin (VN) (Sigma, Saint QuantinFallavier, France) both from human plasma (0.1 mg/mlin PBS) or bovine serum albumin (BSA) 3% overnight atroom temperature. The wells were then rinsed twice withPBS and blocked with 3% BSA/PBS for 1 h at 37°C. Cellsfrom 75-cm2 flasks were harvested in EDTA (2 mm) inPBS and centrifuged at 500 g for 10 min. The cell suspen-sion was diluted in DMEM/F-12 to 1 × 106 cells and

RGD-mediated gene internalisation and expressionM Colin et al
1496 mixed gently by rotation for 1 h at 4°C with 500 ml of0.1 mg/ml solutions of the integrin-binding peptide[K]16RGD (which blocks binding of fibronectin andvitronectin to integrins) or the non-binding control [K]16-RGE peptide or BSA. Fifty microlitres of cell suspension(5 × 104 cells) was added to each well preincubated witheither FN, VN or BSA and incubated at 37°C for 1.5 h.The wells were washed gently with PBS and the attachedcells fixed by the addition of 50 ml methanol for 10 min.The methanol was removed and the remaining cells werestained with 0.1% (w/v) crystal violet in distilled waterfor 10 min at room temperature. The excess crystal violetwas removed by washing with PBS and the plates air-dried. The stained cells were solubilised by the additionof 50 ml of 1% SDS to each well. The absorbance at 570 nmof each well was then measured using a multiwell platespectrometer. The protein concentration of parallel cul-tures of cells were assayed for protein by the Bradford33
method after lysis in 1 n NaOH.
DNA preparation and labellingAn American firefly (Phoptinus pyralis) luciferase DNAplasmid (5256 bp) under the control of an SV40 promoterand enhancer (Promega, Charbonnieres, France) wasused as a reporter gene. Plasmid DNA was purified usingQiagen columns and the endotoxin-free Maxi kit afteralkaline lysis of cells pelleted from overnight cultures ofEscherichia coli DH5a.
The plasmid was labelled by a modified nick-trans-lation technique, which uses either no or very low levelsof DNase and thereby preserves intact the plasmid DNAas previously described.34 The DNA was labelled witheither dUTP-fluorescein (Boehringer Mannheim), dUTP-Cy5 (Amersham, Les Ulis, France) or dATP-[35S](Amersham). One microgramme of DNA was labelled at16°C for 1 h in the presence of 5 ml of nick-translationbuffer (10 ×: 500 mm Tris-HCl, pH 7.8, 100 mg/ml nucle-ase-free BSA, 100 mm b-mercaptoethanol, 50 mm MgCl2),5 ml of enzyme mix (10 ×: 50 mm Tris-HCl, pH 7.5, 5 mmmagnesium acetate, 1 mm PMSF, 50% glycerol (v/v),100 mg/ml nuclease-free BSA, 0.25 U/ml DNA poly-merase I, 5 × 10−4 U/ml DNase I), 5 ml of dXTP mix (5 ×:0.5 mm dATP, 0.5 mm dCTP, 0.5 mm dGTP, 0.34 mmdTTP) and 1.6 ml of dUTP-fluorescein or dUTP-Cy5 in a50 ml reaction volume. The reaction was stopped by theaddition of 3 mm EDTA and by incubation at 65°C for10 min. Unincorporated nucleotides were removed byspinning through a Sephadex G50 column. LabelledDNA was ethanol precipitated and resuspended in 50 mlof OptiMEM medium (Gibco BRL, Cergy Pontoise,France).
For labelling with dATP-[35S], 1 mg DNA was incu-bated at 16°C, 1 h, in the presence of 20 ml dXTP mix(0.3 mm dCTP, dGTP, dTTP), 20 ml of enzyme mix con-taining 5 × 10−5 U/ml DNase, and 3.3 ml of dATP [35S](specific activity .1000 Ci/mmol). The labelled DNAwas recovered as described above.
Complex formationThe [K]16RGD or [K]16RGE (FITC labelled or nonlabelled)were mixed with the plasmid DNA pGL3 at a ratio of5:1 (w:w), respectively and incubated for 15 min at roomtemperature in HEPES buffered saline. Peptide at thisratio condenses the DNA as shown by electronmicroscopy and renders it resistant to DNase.12 Lipofecta-
mine (Gibco BRL) (2 mg/ml) a 3:1 (w:w) mixture of apolycationic lipid (DOSPA) and a neutral lipid (DOPE)was then added (final ratio DNA/peptide/lipofectamine;1:5:24 (w:w:w) unless otherwise indicated) and the mix-ture incubated a further 15 min. DNA retardation wasdetermined as previously described.12
Luciferase assayCells were seeded on 12-well cell culture plates at adensity of 4 × 105 cells per well and incubated at 37°C forabout 12 h until 50% confluency. The components ofthe vector complex (DNA/peptide/Lipofectamine,0.08 mg:0.4 mg:1.9 mg, per well) alone or together wereadded to the cells in a final volume of 200 ml of Opti-MEM. Cells were then incubated at 37°C for 4 h afterwhich the components of the complex were removed.The cells were then washed in PBS, the normal sup-plemented media added and incubated for a further 48 hafter the transfection period. Forty-eight hours was foundto give optimal activity. Luciferase activity was assayedafter washing cells twice with 1 ml of PBS and harvestingin 400 ml of lysis buffer (25 mm Tris/H3PO4, pH 7.8, 1%Triton X-100, 15% glycerol, 1 mm EDTA, 10 mm MgCl2,1 mm DTT) for 15 min at 4°C. The cell lysate was thentransferred to 1.5 ml Eppendorf tubes and centrifuged for10 min at 21 700 g to pellet cellular debris. Luciferaseactivity of cell lysate was then determined on an aliquot(100 ml) using a Berthold (Evry, France) luminometer(Lumat LB9507) with a 60 s integration period after auto-matic injection of 100 ml of a d-luciferin solution (lysisbuffer without DTT, with 100 mm ATP, 43 mg/ml d-luciferin). The protein concentration of cell lysates wasdetermined using the Bradford method.33
Cell association of [35S]-labelled DNACells at approximately 50% confluency (4 × 105 cells perwell) in 12-well plates were incubated with either [35S]-labelled plasmid DNA, [35S]-labelled DNA/[K]16RGD or[35S]-labelled DNA/[K]16RGE complex (0.02 mg DNA perwell) with or without lipofectamine for different times at37°C. The cells were rinsed with PBS, harvested with250 ml trypsin/EDTA per well for 10 min at 37°C or for1 h at 4°C before being blocked with 250 ml of 1% BSAin PBS and recovered by centrifugation. The cell pellet(10 min, 100 g) was rinsed once in PBS by centrifugationbefore lysis in 1 n NaOH. The protein concentration wasdetermined by the method of Bradford33 and the radioac-tivity measured by scintillation counting with Ecolite(ICN, Orsay, France) in an LKB Rackbeta 1209 counter.
Fluorescence flow cytometryFluorescence flow cytometric analysis was performed ona Cytofluorograph IIs (Ortho Diagnostic Systems, West-wood, MA, USA). For b1 integrin immunodetection cellswere detached with 5 mm EDTA in PBS, counted andrinsed three times with 3% BSA in PBS. 5 × 105 cells werethen incubated with a mouse monoclonal anti-humanCD29 (b1 integrin) antibody (0.1 mg/ml, Cliniscience,Birmingham, AL, USA) at a 1/50 dilution for 30 min at4°C. Cells were then washed three times with 3% BSA inPBS and then incubated with a fluorescein isothiocyanate(FITC)-conjugated goat anti-mouse IgG (2 mg/ml; Biosys,Compiegne, France) at a 1/20 dilution for 30 min at 4°C.After three rinses the cells were resuspended in DMEMwithout phenol red and analysed. Negative controls con-sisted of cells incubated with only the anti-IgG antibody.

RGD-mediated gene internalisation and expressionM Colin et al
1497For detection of cell-associated FITC-labelled peptidecells (2 × 106) were transfected with FITC-labelled[K]16RGD/DNA complex (2.5 mg/0.5 mg DNA), with orwithout lipofectamine (6 mg), for 0.25, 0.5, 1.0, 2.0, 3.0 or4.0 h at 37°C. Cells were detached with 0.05% trypsin,0.54 mm EDTA, centrifuged and resuspended beforeanalysis in DMEM at 106 cells per ml. The percentage offluorescent cells was determined after substraction of theautofluorescence of the control cells.
Confocal laser scanning microscopyCells (1.5 × 104 cells per well) were plated on Labtek 8-well plates (Gibco BRL) and the following day fluor-escein-labelled [K]16RGD/Cy5-labelled DNA complex, atthe above indicated optimal ratio, (0.3 mg DNA per well)was added together with or without lipofectamine inOptiMEM for 30 min; the same batch of labelled DNAand peptide were used. Tetramethylrhodamine-5-isothi-ocyanate (TRITC) labelled horseradish peroxidase(150 mg/ml; Sigma), a fluid phase marker which entersintracellular vesicles was then added to the cells rinsedtwice in PBS and incubated for 5 min in PBS containing5% BSA. The cells were washed twice in PBS and thefluorescence was examined with a Leica TCS microscopeequipped with a DMR inverted microscope and a 63/1.4objective. A krypton-argon mixed-gas laser was used togenerate three bands: 488 nm for FITC, 568 nm for TRITCand 657 nm for Cy5. A band pass filter was used torecover FITC fluorescence, a LP 590 filter was used forTRITC and a LP 650 filter was used for Cy5. Images wereprocessed using a Power Mac 7500/100 equipped withPhotoshop and printed on a Kodak XLS 8600 P.S. printer.
Statistical analysisThe results are representative of three experiments whereeach point was performed in triplicate and presented asthe mean ± s.e.m. Statistical analysis was performedusing the nonparametric rank test of Wilcoxon.Probability values ,0.05 were considered statisticallysignificant.
AcknowledgementsThis work was supported by grants from the InstitutNational de la Sante et de la Recherche Medicale(INSERM) and from the Association Francaise de Luttecontre la Mucoviscidose to MC, MCB-H and CC. CC,RPH and AK are supported by the MRC and the MullerBequest. We would like to thank Dr DG Gruenert of theGene Therapy Core Center, UCSF, USA for his kind giftof the foetal human tracheal epithelial cell line(56FHTE8o−) and Yves Chretien for help with treatmentof confocal images and statistical analysis.
References1 Curiel DT, Pilewski JM, Albelda SM. Gene therapy approaches
for inherited and acquired lung diseases. Am J Respir Cell MolBiol 1996; 14: 1–18.
2 Coutelle C. Gene therapy for cystic fibrosis – strategies, prob-lems and perspectives In: Strauss M, Baranger JA (eds). Conceptsin Gene Therapy. Walter de Gruyter: Berlin, 1997, pp 313–343.
3 Felgner PL et al. Lipofection: a highly efficient, lipid-mediatedDNA-transfection procedure. Proc Natl Acad Sci USA 1987; 84:7413–7417.
4 Zhou X, Huang L. DNA transfection mediated by cationic lipo-somes containing lipopolylysine: characterization and mech-anism of action. Biochim Biophys Acta 1994; 1189: 195–203.
5 Wrobel I, Collins D. Fusion of cationic liposomes with mam-malian cells occurs after endocytosis. Biochim Biophys Acta 1995;1235: 296–304.
6 Zabner J et al. Cellular and molecular barriers to gene transferby a cationic lipid. J Biol Chem 1995; 270: 18997–19007.
7 Matsui H, Johnson LG, Randell SH, Boucher RC. Loss of bindingand entry of liposome–DNA complexes decreases transfectionefficiency in differentiated airway epithelial cells. J Biol Chem1997; 272: 1117–1126.
8 Porteous DJ et al. Evidence for safety and efficacy of DOTAPcationic liposome mediated CFTR gene transfer to the nasal epi-thelium of patients with cystic fibrosis. Gene Therapy 1997; 4:210–218.
9 Cotten M, Wagner E. Non-viral approaches to gene therapy.Curr Opin Biotechnol 1993; 4: 705–710.
10 Lasic DD, Templeton NS. Liposomes in gene therapy. Adv DrugDel Rev 1996; 20: 221–266.
11 Hart SL et al. Gene delivery and expression mediated by an inte-grin-binding peptide. Gene Therapy 1995; 2: 552–554.
12 Harbottle RP et al. An Arg-Gly-Asp/oligolysine peptide: aprototype construct for integrin mediated gene delivery. HumGene Ther 1998; 9: 1037–1047.
13 Rouslahti E. RGD and other recognition sequences for integrins.Annu Rev Cell Dev Biol 1996; 12: 697–715.
14 Isberg RR, Tran Van Nhieu G. The mechanism of phagocyticuptake promoted by invasin–integrin interaction. Trends CellBiol 1995; 5: 120–124.
15 Logan D et al. Structure of a major immunogenic site on foot-and-mouth disease virus. Nature 1993; 362: 566–568.
16 Coulson BS, Londrigan SL, Lee DJ. Rotavirus contains integrinligand sequences and a disintegrin-like domain that are impli-cated in virus entry into cells. Proc Natl Acad Sci USA 1997; 94:5389–5394.
17 Wickham TJ, Filardo EJ, Cheresh DA, Nemerow GR. Integrinavb5 selectively promotes adenovirus mediated cell membranepermeabilization. J Cell Biol 1994; 127: 257–264.
18 Trubetskoy VS, Torchilin VP, Kennel S, Huang L. Cationic lipo-somes enhance targeted delivery and expression of exogenousDNA mediated by N-terminal modified poly(l-lysine)-antibodyconjugate in mouse lung endothelial cells. Biochim Biophys Acta1992; 1131: 311–313.
19 Remy J-S et al. Targeted gene transfer into hepatoma cells withlipopolyamine-condensed DNA particles presenting galactoseligands: a stage toward artificial viruses. Proc Natl Acad Sci USA1995; 92: 1744–1748.
20 Cheng P-W. Receptor ligand-facilitated gene transfer: enhance-ment of liposome-mediated gene transfer and expression bytransferrin. Hum Gene Ther 1996; 7: 275–282.
21 Hart SL et al. Lipid-mediated enhancement of transfection by anonviral integrin-targeting vector. Hum Gene Ther 1998; 9:575–585.
22 Levy P et al. Enterocytic differentiation of the human Caco-2 cellline correlates with alterations of integrin signaling. J Cell Physiol(in press).
23 Wattiaux R et al. Cationic lipids delay the transfer of plasmidDNA to lysosomes. Biochem Biophys Res Comm 1996; 227: 448–454.
24 Conforti G et al. Modulation of vitronectin receptor binding bymembrane lipid composition. J Biol Chem 1990; 265: 4011–4019.
25 Pijuan-Thompson V, Gladson CL. Ligation of integrin a5b1 isrequired for internalization of vitronectin by integrin avb3. JBiol Chem 1997; 272: 2736–2743.
26 Panetti TS, McKeown-Longo PJ. The avb5 integrin receptorregulates receptor-mediated endocytosis of vitronectin. J BiolChem 1993; 268: 11492–11495.
27 Palmer EL et al. Sequence and tissue distribution of the integrina9 subunit, a novel partner of b1 that is widely distributed inepithelia and muscle. J Cell Biol 1993; 123: 1289–1297.

RGD-mediated gene internalisation and expressionM Colin et al
1498 28 Pilewski JM, Latoche JD, Arcasoy SM, Albelda SM. Expressionof integrin cell adhesion receptors during human airway epi-thelial repair in vivo. Am J Physiol 1997; 273: L256–L263.
29 Herand A-L et al. Fibronectin and its a5b1-integrin receptor areinvolved in the wound repair process of airway epithelium. AmJ Physiol 1996; 271: L726–L733.
30 Gruenert DC et al. Characterization of human tracheal epithelialcells transformed by an origin-defective simian virus 40. ProcNatl Acad Sci USA 1988; 85: 5951–5955.
31 Lemnaouar M et al. Oncogene-mediated propagation of trachealepithelial cells from two cystic fibrosis fetuses with different
mutations. Characterization of CFT-1 and CFT-2 cells in culture.Eur J Clin Invest 1993; 23: 151–160.
32 Mergey M et al. CFTR gene transfer corrects defective glycocon-jugate secretion in human CF epithelial tracheal cells. Am J Phy-siol 1995; 269: L855–L864.
33 Bradford MA. A rapid and sensitive method for the quantifi-cation of microgram quantities of protein utilizing protein-dyebinding. Anal Biochem 1976; 72: 248–254.
34 Wheeler VC, Coutelle C. Nondegradative in vitro labeling ofplasmid DNA. Anal Biochem 1995; 225: 374–376.




















