
Inhibitory effect of phorbol ester on carbachol-induced signal transduction in cultured canine...
10
Original Paper J BiomedSci 1995;2:283-292 a Cellular and Molecular Pharmacology Laboratory,Departmentof Pharmacology, and b Departmentof Internal Medicine, Chang GungCollege of Medicineand Technology, Tao-Yuan,Taiwan, Republicof China • o, •o, ,• • • leel leoo ,,oe eeeei *e,eoe~eeooel**,,o tl KeyWords Phorbol ester Carbachol Protein kinase C Inositol phosphates Calcium ion Muscarinic receptor Tracheal smooth muscle cell Inhibitory Effect of Phorbol Ester on Carbachoi-lnduced Signal Transduction in Cultured Canine Tracheal Smooth Muscle Cells ~leo, • ~ualoo I• oollo I o*eee~ooooe~omt***~met t*oomoal o*~loeo* • *oell*omlooa***el,m~oeeleQ**oee,g**eee~ Abstract Regulation of the increases in inositol 1,4,5-trisphosphate (IP3) production and intracellular Ca 2+concentration ([Ca2+]i) by activation of protein kinase C (PKC) was investigated in cultured canine tracheal smooth muscle cells (TSMCs). Stimulation of TSMCs by carbachol led to IP3 formation and caused an initial transient peak of [Ca2+]i followed by a sustained elevation in a concentration-dependent manner. Pretreatment of TSMCs with phorbol 12- myristate 13-acetate (PMA, 1 gM) for 30 rain blocked the carbachol-induced IP3 formation and Ca 2+ mobilization. Following preincubation, carbachol- induced Ca 2+mobilization recovered within 24 h. The concentrations of PMA that gave half-maximal inhibition of carbachol-induced IP3 formation and increase in [Ca2+]i were 7 and 4 nM, respectively. Prior treatment of TSMCs with staurosporine (1 gM), a PKC inhibitor, inhibited the ability of PMA to attenuate carbachol-induced responses. Inactive phorbol ester, 4a-phorbol 12,13-didecanoate at 1 gM, did not inhibit these responses to carbachol. The 3 Kd and Bmax of the muscarinic receptor for [ H]N-methylscopolamine binding were not significantly changed by PMA treatment. PMA also decreased PKC activity in the cytosol of TSMCs, while increasing it transiently in the mem- branes within 30 min. Thereafter, the membrane-associated PKC activity decreased and persisted for at least 24 h of PMA treatment. Taken together, these results suggest that activation of PKC may inhibit phosphoinositide hydrolysis and consequently attenuate the [Ca2+]i increase or inhibit both responses independently. The inhibition by PMA of carbachol-induced re- sponses was inversely correlated with membranous PKC activity. ioe,oeiQeee,,eaee,o~we The contractile response of tracheal smooth muscle (TSM) to muscarinic agonists is mediated through musca- rinic receptors (mAChRs) [2]. Stimulation of mAChRs results in the hydrolysis of phosphoinositide (PI) through activation of phospholipase C (PLC), thus generating two intracellular messengers, inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG) [30, 32]. Studies to date suggest a major role for IP3 in the agonist-triggered rise in [Ca2+]i that accompanies activation in many cell types. Thus, in terms of second-messenger generation, contrac- Received: July22,1994 Accepted: November 3, 1994 Chuen Mao Yang, PhD Department of Pharmacology Chang Gtmg College of Medicine and Technology 259 Wen-Hwa I Road Kwei-San, Tao-Yuan, Taiwan (ROC) © 1995 National Science Council, ROC; S. Karger AG, Basel 1021-7770/95/0023-0283 $8.00/0
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Inhibitory effect of phorbol ester on carbachol-induced signal transduction in cultured canine...

Original Paper
J Biomed Sci 1995;2:283-292
a Cellular and Molecular Pharmacology Laboratory, Department of Pharmacology, and
b Department of Internal Medicine, Chang Gung College of Medicine and Technology, Tao-Yuan, Taiwan, Republic of China
• o , • o , , • • • l e e l l e o o , , o e e e e e i * e , e o e ~ e e o o e l * * , , o t l
KeyWords Phorbol ester Carbachol Protein kinase C Inositol phosphates Calcium ion Muscarinic receptor Tracheal smooth muscle cell
Inhibitory Effect of Phorbol Ester on Carbachoi-lnduced Signal Transduction in Cultured Canine Tracheal Smooth Muscle Cells
~leo, • ~ualoo I• ool lo I o*eee~ooooe~omt***~met t*oomoal o*~loeo* • *oell*omlooa***el,m~oeeleQ**oee,g**eee~
Abstract Regulation of the increases in inositol 1,4,5-trisphosphate (IP3) production and intracellular Ca 2+ concentration ([Ca2+]i) by activation of protein kinase C (PKC) was investigated in cultured canine tracheal smooth muscle cells (TSMCs). Stimulation of TSMCs by carbachol led to IP3 formation and caused an initial transient peak of [Ca2+]i followed by a sustained elevation in a concentration-dependent manner. Pretreatment of TSMCs with phorbol 12- myristate 13-acetate (PMA, 1 gM) for 30 rain blocked the carbachol-induced IP3 formation and Ca 2+ mobilization. Following preincubation, carbachol- induced Ca 2+ mobilization recovered within 24 h. The concentrations of PMA that gave half-maximal inhibition of carbachol-induced IP3 formation and increase in [Ca2+]i were 7 and 4 nM, respectively. Prior treatment of TSMCs with staurosporine (1 gM), a PKC inhibitor, inhibited the ability of PMA to attenuate carbachol-induced responses. Inactive phorbol ester, 4a-phorbol 12,13-didecanoate at 1 gM, did not inhibit these responses to carbachol. The
3 Kd and Bmax of the muscarinic receptor for [ H]N-methylscopolamine binding were not significantly changed by PMA treatment. PMA also decreased PKC activity in the cytosol of TSMCs, while increasing it transiently in the mem- branes within 30 min. Thereafter, the membrane-associated PKC activity decreased and persisted for at least 24 h of PMA treatment. Taken together, these results suggest that activation of PKC may inhibit phosphoinositide hydrolysis and consequently attenuate the [Ca2+]i increase or inhibit both responses independently. The inhibition by PMA of carbachol-induced re- sponses was inversely correlated with membranous PKC activity. ioe,oeiQeee, ,eaee,o~we
The contractile response of tracheal smooth muscle (TSM) to muscarinic agonists is mediated through musca- rinic receptors (mAChRs) [2]. Stimulation of mAChRs results in the hydrolysis of phosphoinositide (PI) through activation of phospholipase C (PLC), thus generating two
intracellular messengers, inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG) [30, 32]. Studies to date suggest a major role for IP3 in the agonist-triggered rise in [Ca2+]i that accompanies activation in many cell types. Thus, in terms of second-messenger generation, contrac-
Received: July22,1994 Accepted: November 3, 1994
Chuen Mao Yang, PhD Department of Pharmacology Chang Gtmg College of Medicine and Technology 259 Wen-Hwa I Road Kwei-San, Tao-Yuan, Taiwan (ROC)
© 1995 National Science Council, ROC; S. Karger AG, Basel 1021-7770/95/0023-0283 $8.00/0

t ion of TSM may be mediated by IP3-induced Ca 2+ mobi- lization from intracellular stores.
Activation of PI hydrolysis by mAChRs also leads to the formation of DAG, which activates an important reg- ulatory enzyme protein kinase C (PKC) [19]. The PI-Ca 2+ signaling system has been reported to be negatively regu- lated by PKC activation, by showing a decrease in PI hydrolysis in several cell types [22, 23, 25], intracellular Ca 2+ mobilization [16, 27], or both responses [21 ]. These reports suggest that there are many candidates for targets of PKC in the regulation of PI-Ca 2+ signaling in these cells. Phorbol esters, which mimic the effect of DA G and cause permanent activation of PKC [7], are useful re- agents for examining the role of D A G in the regulation of PI hydrolysis and Ca 2+ mobilization related to smooth muscle function. Furthermore, phorbol ester has been shown to inhibit histamine-induced IP3 formation and calcium mobilization in cultured TSM cells (TSMCs) [ 13, 18]. Therefore, a question arises as to whether there is an inhibitory or stimulatory effect o f phorbol 12-myristate 13-acetate (PMA) on muscarinic responses in canine TSMCs.
The precise molecular and biochemical mechanisms of the relation between phorbol esters and muscarinic desen- sitization of physiological responses in TSM are not clear. The purpose o f the present study was to investigate the role of the DAG-PKC pathway in the modulat ion of phar- macological properties o f mAChRs, PI breakdown, change in [Ca2+]i, and the involvement of PKC in the mechanism of desensitization of mAChRs induced by phorbol esters.
Materials and Methods
Materials Dulbecco's modified Eagle's medium (DMEM)/Ham's nutrient
mixture F-12 (F-12) medium and fetal bovine serum (FBS) were pur- chased from J.R. Scientific (Woodland, Calif., USA). Insulin and insulin-like growth factor (IGF)-I were from Boehringer Mannheim (Germany). Myo-[2-3H]inositol (18 Ci/mmol), [3H]N-methyl-scopol- amine ([3H]NMS; 87 Ci/mmol) and r-[32p]ATP (3,000 Ci/mmol) were from Amersham (UK). Fura 2-AM was from Molecular Probes (Eugene, Oreg., USA). Enzymes mad other chemicals were from Sig- ma (St. Louis, Mo., USA).
Animals Mongrel dogs, 10-20 kg, purchased from a local supplier, were
used throughout this study. They were housed in animal facilities under an automatically controlled temperature and light cycle and fed standard laboratory chow and tap water ad libitum. Dogs of eith- er sex were anesthetized with pentobarbitone (30 mg/kg, intrave- nously) and the lungs were ventilated mechanically via an orotra- cheal tube. The trachea was surgically removed.
Isolation of TSMCs The TSMCs were isolated according to methods previously
reported [29]. Briefly, the trachea was cut longitudinally through the cartilage rings and the smooth muscle was dissected. The mus- cle was minced and transferred to the dissociation medium con- taining 0.2% collagenase IV, 0.01% deoxyribonuctease I, 0.01% elastase IV, and antibiotics (100 U/ml penicillin G0 100 ~tg/ml streptomycin, and 250 ng/ml fungizone) in physiological solution. The physiological solution contained: 137 mM NaCt, 5 true/KC1, 1.1 mM CaCI2, 20 mMNaHCO3, 1 mMNaH2PO4, 11 mM glucose and 25 mM HEPES (pH 7.4). The tissue pieces were gently agi- tated at 37°C in a rotary shaker for 1 h. The released cells were collected and the residue was again digested with fresh enzyme solution for an additional 1 h at 37°C. The released cells were washed twice with DMEM/F-12 medium (1:1 vol/vol). The cells, suspended in DME/Vl/F-12 containing 10% FBS, were plated onto a 60-mm culture dish and incubated at 37 °C for 1 h to remove fibroblasts which attached to the dish more rapidly than TSMCs. The cell number was counted and the suspension diluted with DMEM/F-12 to 2 x 105 cells/ml. The cell suspension was plated onto 24-well (0.5 ml/weU), 12-well (1 ml/well), or 6-well (2 ml/well) culture plates containing glass coverstips coated with collagen for receptor-binding assay, IP accumulation and Ca 2+ measurement, respectively. The medium was changed after 24 h and thereafter every 3 days. After 5 days of culture, medium was changed to DMEM/F-12 containing 1% FBS for 24h at 37°C. The cells were then incubated in DMEM/F-12 containing 1% FBS supplemented with IGF-I (10 ng/ml) and insulin (1 gg/ml) for 12-14 days. These culture conditions were adopted to maintain the expression of the receptors on the cultured TSMCs [29].
To characterize the isolated and cultured TSMCs and to exclude contamination by epithelial cells and fibroblasts, the cells were iden- tified by an indirect immunofluorescent staining method using a monoclonal antibody specifically against smooth muscle light-chain myosin [9]. Over 95% of the cell preparation was composed of smooth muscle cells.
Accumulation of lnositol Phosphates (IPs) The effect of carbachol on the hydrolysis of PI was assayed by
monitoring the accumulation of 3H-labelled IPs as described by Bet- ridge et al. [5]. Cultured TSMCs were incubated with 5 gCi/ml of myo-[2-3H]inositol at 37 °C for 2 days. TSMCs were washed twice with PBS and incubated in Krebs-Henseleit buffer (KHS, pH 7.4) containing 117 rru'v/NaC1, 4.7 mM KC1, 1.1 mM MgSO4, 1.2 n'u'¢l KHzPO4, 10 mM NaHCO3, 2.4 mM CaC12, 1 mM glucose, 20 mM HEPES, and 10 mM LiC1 at 37°C for 30 min. After carbachol was added at the concentration indicated, incubation was continued for another 20 rain. When phorbol ester was used, it was added at the time indicated before the addition of carbachol. Reactions were ter- minated by addition of 5% perchloric acid followed by sonication and centrifugation at 3,000 g for 15 rain.
The perchloric-acid-soluble supernatants were extracted four times with ether, neutralized with potassium hydroxide, and applied to a colum of AG1-X8, formate form, 100-200 mesh (Bio-Rad). The resin was washed successively with 5 ml of water and 5 ml of 60 mM ammonium formate/5 mM sodium tetraborate to eliminate free myo-[2-3H]inositol and glycerophosphoinositol. Sequential washes with 5 ml of 0.2M ammonium formate/0.1 M formic acid, 0.4M ammonium formate/0.1 M formic acid, and 1 M ammonium for- mate/0.1 M formic acid were used to elute inositol monophosphate
284 Yang/Hsu/Ong/Hsieh/Tsao/Chen/Luo Protein Kinase C and Signal Transduction

(IP1), inositol bisphosphate (IPz), and IP3, respectively. The amount of [3H]IPs was determined in a radiospectrometer (Beckman TA5000).
Measurement of [Ca2+]i [Ca2+]i was measured in confluent monolayers with the calcium-
sensitive dye fura 2-AM as described by Grynkiewicz et al. [10]. Upon confluence, the cells were cultured in DMEM/F-12 with 1% FBS for 1 day before measurements were made. The monolayers were covered with 1 ml of DMEM~-12 with 1% FBS containing 5 pM fura 2-AM, then incubated at 37 ° C for 45 min. At the end of the incubation, the coverslips were washed twice with the physiologi- cal buffer solution containing: 125 mM NaC1, 5 mM KC1, 1.8 mM CaClz, 2 mM MgClz, 0.5 m_M NaHzPO4, 5 mM NaHCO3, 10 mM HEPES, and 10 mM glucose, pH 7.4. The cells were incubated in PBS for a further 30 min to complete dye de-esterifieation. The cov- erslip was inserted into a quartz cuvette at an angle of approximately 45 ° to the excitation beam and placed in the temperature-controlled holder of an SLM 8000C spectrofluorometer (SLM Aminco, Urbana, IU., USA). Continuous stirring was achieved with a magnetic stirrer.
Fluorescence of CaZ+-bound and unbound fura 2 was measured by rapidly alternating the dual-excitation wavelengths between 340 and 380 nm and electronically separating the resulting fluorescence signals at emission wavelength 510 nm. The autofluorescence of each monolayer was subtracted from the fluorescence data. The ratios (R) of the fluorescence at the two wavelengths were computed and used to calculate changes in [Ca2+]i. The ratios of maximum (Rma~) and minimum (Rmin) fluorescence of fura 2 were determined by adding ionomycin (10 gM) in PBS containing 5 mM Ca 2+ and by adding 5 rm~ r EGTA at pH 8 in Ca2+-free PBS, respectively. The K~ of fura 2 for Ca 2+ was assumed to be 224 nM [10].
PKC Assay Cultured TSMCs were washed with phosphate-buffered saline
(PBS), and then sonicated for 1 rain in 1 ml of Tris-HC1 buffer (20 raM, pH 7.5) containing 0.5 mMEGTA, 1 mM dithiothreitol, 2 mM benzamidine, 1 gg/ml leupeptin, and 0.1 gg/ml pepstatin. The soni- care was centrifuged at 1,000 g for 8 min to remove unbroken cells and nuclei. Soluble and particulate fractions were separated by high- speed centrifugation (100,000 g for 1 h). PKC was assayed at 30 ° C as described elsewhere [3]. The reaction mixture (250 gt) contained 20 mM PIPES buffer (pH 7.0), type III histone (0.5 mg/ml), 100 gM CaC12, 5 m_M MgClz, and 30 gg/ml of protein. The reaction was ini- tiated by the addition of 25 p_M r-[3zP]ATP (approximately 8,000 cpm/gl) and incubated at 37 ° C for 5 min. Additional components of the reaction mixture were: 0.1 mM EGTA, 25 gg/ml phosphatidyl- serine, 0.5 ixg/ml diolein, 1 ~MPMA. Samples (150 lxl) of the reac- tion mixture were added to 4 ml of an ice-cold mixture of 5% trichlo- roacetic acid. The diluted samples were filtered through 0.45-~tm ill- ters and washed eight times with 2 ml of the reaction buffer. Filters and samples (5 gl) of the incubation mixture were treated identically for scintillation counting. Enzyme levels were normalized to protein contents measured by the method of Bradford [6].
Muscarinic Receptor Binding Assay For detection of the effect of PMA treatment on the density and
affinity of mAChRs on TSMCs, [3H]NMS was used as a ligand. The total number of cell surface [3H]NMS-binding sites was determined by incubating cells in PBS containing various concentrations of [3H]NMS ranging from 10 to 1,250 pM. Nonspecific binding of
[3H]NMS was determined in the presence of 1 p,~¢i atropine sulfate under the same incubation conditions. The binding-reaction mixture was incubated at 37 °C for 45 min and the reaction was terminated by removing the mixture and washing the cells three times with PBS. Cells were solubilized in 0.1 N NaOH and counted in a radiospec- trometer. Counting data were corrected for the specific activity of the ligand and quenching. The amount of specific binding was calculated as the total binding minus the binding in the presence of 1 gM atro- pine sulfate. The dissociation constant (K~) and total receptor densi- ty (Bmax) were calculated by the Ligand program, as previously described [29].
Analysis of Data The ECs0 values were estimated by the Graph Pad Program
(Graph Pad, San Diego, Calif., USA). The concentration-effect curves were constructed by noncumulative addition of carbachol and fitted by sigmoid curve (log scale). The data were expressed as the mean + SE of experiments with statistical comparisons based on a two-tailed Student's t test, with p < 0.01 taken as the level of signifi- cance.
Results
Effects of PMA on Carbachol-Stimulated IPs Accumulation To determine whether P K C act ivat ion by phorbol
ester caused a change in carbachol-induced IP3 accumula- tion, the t ime course and concentrat ion-response relation-
ships for P M A inhibit ion of carbachol-s t imulated IP1, IP2
and IP3 accumulat ion were examined in cultured TSMCs. Figure 1 shows that pre t rea tment with 1 g M P M A for 5 min was sufficient to inhibit the subsequent carbachol- s t imulated response by 50%. Following preincubation, max imal inhibit ion ofcarbachol-s t imula ted IP accumula-
t ion was obta ined within 30 rain. P M A was also very potent: 10 n M P M A caused significant inhibit ion (fig. 2), consistent with previous reports in several cell types [ 15,
20, 25, 31]. The inhibitory effect o f P M A was concentra- t ion dependent . P M A induced ha l f -maximal inhibit ion o f
carbachol-st imulated IPx, IP2 and IP3 format ion at 15 -+ 5, 9 + 2 and 7 + 2 n M (n = 4), respectively, but had no effect on the basal levels o f IPs accumulat ion at any of the
concentrat ions tested. The inhibi tory action of P M A ap- peared to result f rom a decrease in the max imal response, ra ther than a shift to the right in the concentration-effect
curve for carbachol- induced IPs accumulat ion (fig. 3). The half max imal values (ECs0) for the s t imulatory effect of carbachol on I P b IPz and IP3 format ion in the presence o f l 0 n M P M A [ 1 4 + 2 ,43 + 12and 19 + 3 n M ( n = 4), respectively] were higher than those in the control cells [10 + 2, 25 _+ 12 and 9 + 1 n M ( n = 4), respectively].
285

Fig. 1. Time course of PMA-induced in- hibition of carbachol-stimulated [3H]IPs ac- cumulation in cultured TSMCs. [3H]inosi- toMabelled TSMCs were preincubated with 1 gM PMA for various times and then ex- posed to 100 gM carbachol for 20min. Reactions were terminated by the addition of 5% perchloric acid and inositol phos- phates were extracted. IP1 (t)), IP2 (11) and IP3 (A) were separated as described in Mate- rials and Methods. Results shown are the mean + SE of triplicate determinations in three separate experiments. Control values were26,300 ± 950,6,100 + 560,and3,700 -+ 430 dpm for IPb IP2 and IP3, respectively.
Fig. 2. Concentration dependence of PMA inhibition of carbachol-stimulated [3H]IPs accumulation. Cells prelabeUed with [3H]inositol were washed with KHS, treated with various concentrations of PMA (0.1 nM-1 gM) for 30 min and then exposed to carbachol (100 gM) for 20 rain. The accu- mulation ofIP1 (e), IP2 (11) and IP3 (A) was determined as described under Materials and Methods. Values are expressed as the mean + SE from four separate experiments determined in triplicate.
Fig. 3. Effect of PMA on concentra- tion-effect curves for carbachol-stimulated [3H]IPs accumulation in cultured TSMCs. Cells prelabelled with [3H]inositol were incu- bated in the absence ((3) or presence (@) of 1 gM PMA for 30 min and then exposed to various concentrations of carbachol for 20 min. The accumulation of IP1 (a), IP2 (b) and IP3 (¢) was determined as described under Materials and Methods. Values are expressed as the mean + SE from four sepa- rate experiments determined in triplicate.
Effects" of PMA on the [Ca2+]i Response to Carbachol Because IP3 is the second messenger involved in the
mobilization o f Ca 2+ from intracellular stores [ 16, 20, 27, 30, 32], one would expect the moblization of Ca 2+ re- sponse to carbachol to correspond inversely with PKC activity. The effect o f carbachol, a constrictor of smooth muscle that operates through calcium-dependent path- ways, on [Ca2+]i was studied in cultured TSMCs. To examine a possible role for PKC in the regulation o f
[Ca2+]i in TSMCs, we tested the effect of PMA, a known activator of PKC, on intracellular Ca 2+ concentration induced by carbachol. Figure 4 shows the t ime course of PMA treatment on carbachol-induced changes in [Ca2+]i up to 24 h. PMA treatment for various periods did not change the resting level of [CaZ+]i. It did, however, signifi- cantly inhibit the transient elevation of [Ca2+]i induced by carbachol in TSMCs treated with PMA for between 30 rain and 2 h (p < 0.001, compared with control cells).
286 Yang/Hsu/Ong/Hsieh/Tsao/Chen/Luo Protein Kinase C and Signal Transduction
T
iiiiiiiiiiiiiii i!ii!i!i~ii!!ii
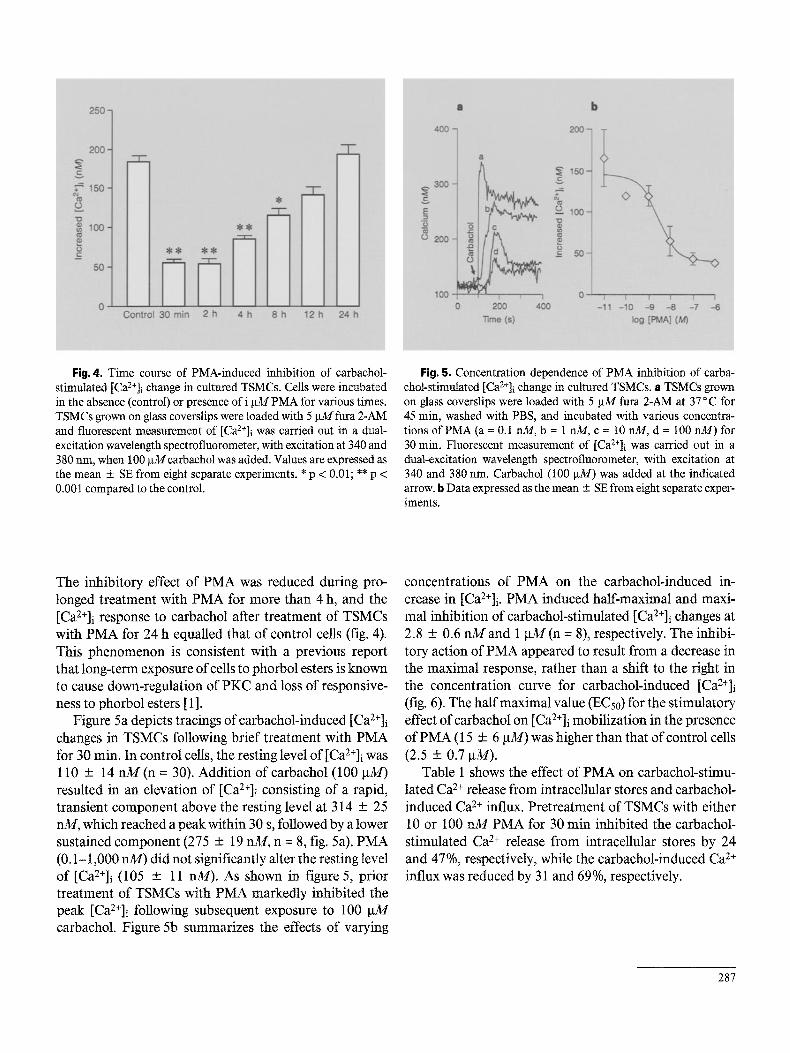
Fig. 4. Time course of PMA-induced inhibition of carbachol- stimulated [Ca2+]i change in cultured TSMCs. Cells were incubated in the absence (control) or presence of i gM PMA for various times. TSMCs grown on glass coverslips were loaded with 5 gM fura 2-AM and fluorescent measurement of [Ca2+]i was carried out in a dual- excitation wavelength spectrofluorometer, with excitation at 340 and 380 rim, when 100 gM carbachol was added. Values are expressed as the mean + SE from eight separate experiments. * p < 0.01; ** p < 0.001 compared to the control.
Fig. 5. Concentration dependence of PMA inhibition of carba- chol-stimulated [Ca2÷]i change in cultured TSMCs. a TSMCs grown on glass eoverslips were loaded with 5 gM fura 2-AM at 37°C for 45 rain, washed with PBS, and incubated with various concentra- tions ofPMA (a = 0.1 nM, b = 1 m~, c = 10 nM, d = 100 nM) for 30 min. Fluorescent measurement of [Ca2+]i was carried out in a dual-excitation wavelength spectrofluorometer, with excitation at 340 and 380 nm. Carbachol (100 gM) was added at the indicated arrow, b Data expressed as the mean + SE from eight separate exper- iments.
The inhibitory effect of PMA was reduced during pro- longed treatment with PMA for more than 4 h, and the [Ca2+]i response to carbachol after treatment of TSMCs with PMA for 24 h equalled that of control cells (fig. 4). This phenomenon is consistent with a previous report that long-term exposure of cells to phorbol esters is known to cause down-regulation of PKC and loss of responsive- ness to phorbol esters [1 ].
Figure 5a depicts tracings of carbachol-induced [Ca2+]i changes in TSMCs following brief t reatment with PMA for 30 min. In control cells, the resting level of [Ca2+]i was 110 + 14 n M ( n = 30). Addition of carbachol (100 gM) resulted in an elevation of [Ca2+]i consisting of a rapid, transient component above the resting level at 314 + 25 nM, which reached a peak within 30 s, followed by a lower sustained component (275 + 19 nM, n = 8, fig. 5a). PMA (0.1-1,000 riM) did not significantly alter the resting level of [Ca2+]i (105 ± 11 nM). As shown in figure 5, prior treatment of TSMCs with PMA markedly inhibited the peak [Ca2+]i following subsequent exposure to 100 g M carbachol. Figure 5b summarizes the effects of varying
concentrations of PMA on the carbachol-induced in- crease in [Ca2+]i. PMA induced half-maximal and maxi- mal inhibition of carbachol-stimulated [Ca2+]i changes at 2.8 + 0.6 n M a n d 1 g M ( n = 8), respectively. The inhibi- tory action of PMA appeared to result from a decrease in the maximal response, rather than a shift to the right in the concentration curve for carbachol-induced [Ca2+]i (fig. 6). The half maximal value (ECs0) for the stimulatory effect ofcarbachol on [Ca2+]i mobilization in the presence of PMA (15 + 6 gM) was higher than that of control cells (2.5 5:0.7 gM).
Table 1 shows the effect of PMA on carbachol-stimu- lated Ca 2+ release from intracellular stores and carbachol- induced Ca 2÷ influx. Pretreatment of TSMCs with either 10 or 100 n M PMA for 30 min inhibited the carbachol- stimulated Ca 2+ release from intracellular stores by 24 and 47%, respectively, while the carbachol-induced Ca 2÷ influx was reduced by 31 and 69%, respectively.
287
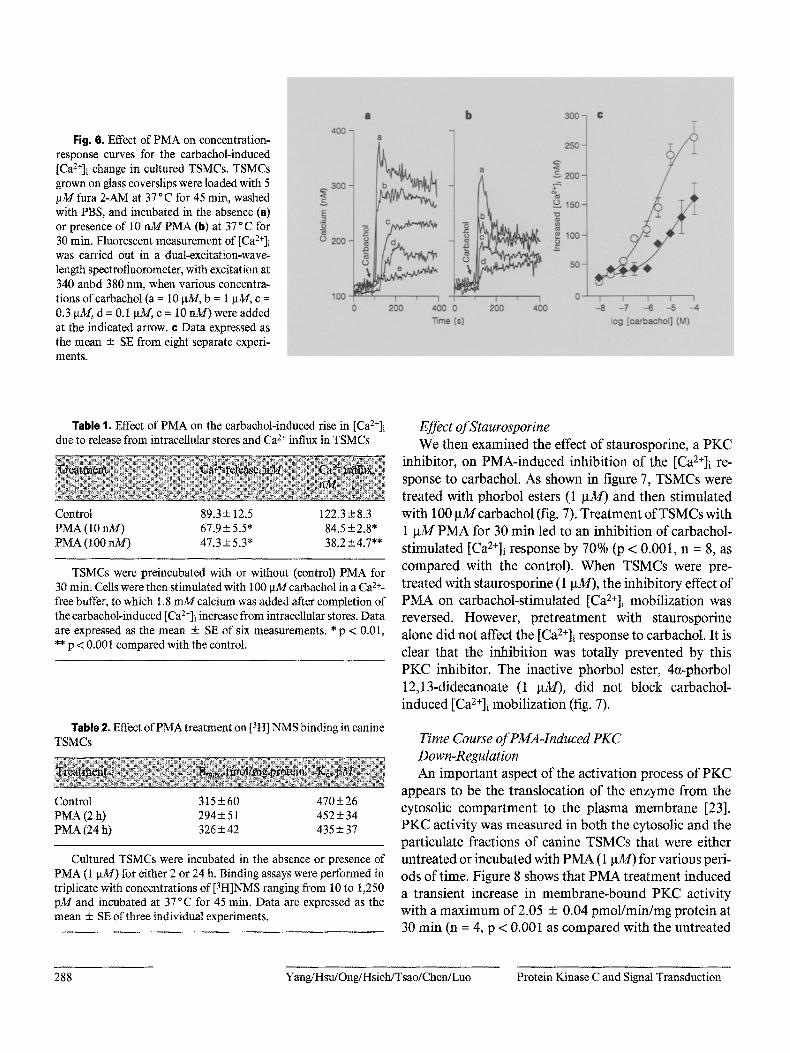
Fig. 6, Effect of PMA on concentration- response curves for the carbachol-induced [Ca2+]i change in cultured TSMCs. TSMCs grown on glass coverslips were loaded with 5 p31 fura 2-AM at 37 °C for 45 min, washed with PBS, and incubated in the absence (e) or presence of 10 nM PMA (b) at 37 ° C for 30 rain. Fluorescent measurement of [Ca2+]i was carried out in a dual-excitation-wave- length spectrofluorometer, with excitation at 340 anbd 380 nm, when various concentra- tions of carbachol (a = 10 p_M, b = 1 la3//, c = 0.3 p~$/, d = 0.1 ~'v/, e = 10 rdd) were added at the indicated arrow. ¢ Data expressed as the mean -+ SE from eight separate experi- ments.
Table 1. Effect of PMA on the carbachol-induced rise in [Ca2+]i due to release from intracellular stores and Ca 2+ influx in TSMCs
Control 89.3 + 12.5 122.3 + 8.3 PMA (l 0 nM) 67.9 + 5.5" 84.5 + 2.8* PMA (100 riM) 47.3 + 5.3* 38.2 + 4.7**
TSMCs were preincubated with or without (control) PMA for 30 min. Cells were then stimulated with 100 pMcarbachol in a Ca z+- free buffer, to which 1.8 n~/calcium was added after completion of the carbachol-induced [Ca2+]i increase from intracellular stores. Data are expressed as the mean + SE of six measurements. * p < 0.01, • * p < 0.001 compared with the control.
Table 2. Effect of PMA treatment on [3HI NMS binding in canine TSMCs
Control 315 + 60 470 + 26 PMA (2 h) 294 + 5 t 452-+ 34 PMA (24 h) 326 + 42 435-+ 37
Cultured TSMCs were incubated in the absence or presence of PMA (1 ~tM) for either 2 or 24 h. Binding assays were performed in triplicate with concentrations of [3H]NMS ranging from 10 to 1,250 pM and incubated at 37 °C for 45 min. Data are expressed as the mean + SE of three individual experiments.
Effect of Staurosporine We then examined the effect o f staurosporine, a P K C
inhibitor, on PMA-induced inhibit ion o f the [Ca2+]i re-
sponse to carbachol. As shown in figure 7, TSMCs were treated with phorbol esters (1 g M ) and then s t imulated with 100 g M carbachol (fig. 7). T rea tmen t of TSMCs with 1 g M P M A for 30 min led to an inhibit ion of carbachol-
st imulated [Ca2+]i response by 70% (p < 0.001, n = 8, as compared with the control). When TSMCs were pre-
treated with staurosporine (1 ~tM), the inhibitory effect o f P M A on carbachol-st imulated [Ca2+]i mobil izat ion was reversed. However, pre t rea tment with staurosporine
alone did not affect the [Ca2+]i response to carbachol. I t is clear that the inhibition was totally prevented by this P K C inhibitor. The inactive phorbol ester, 4a-phorbol
12,13-didecanoate (1 ~tM), did not block carbachol- induced [Ca2+]i mobil izat ion (fig. 7).
Time Course of PMA-Induced PKC Down-Regulation An impor tan t aspect o f the act ivat ion process of P K C
appears to be the translocation of the enzyme f rom the cytosolic compar tmen t to the p lasma m e m b r a n e [23]. P K C activity was measured in both the cytosolic and the particulate fractions o f canine TSMCs that were either untreated or incubated with P M A (1 gM) for various peri- ods o f time. Figure 8 shows that P M A t rea tment induced a transient increase in membrane -bound P K C activity with a m a x i m u m of 2.05 + 0.04 pmol /min /mg protein at 30 min (n = 4, p < 0.00I as compared with the untreated
288 Yang/Hsu/Ong/Hsieh/Tsao/Chen/Luo Protein Kinase C and Signal Transduction
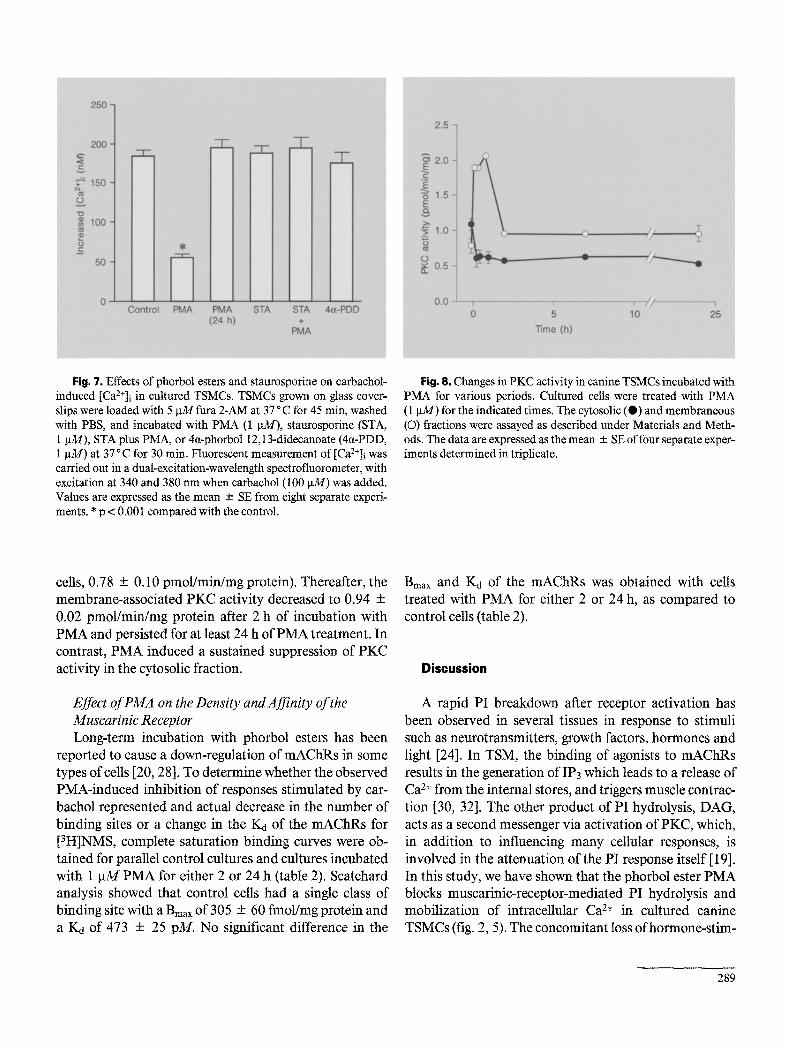
Fig. 7. Effects of phorbol esters and staurosporine on carbachol- induced [Ca2+]i in cultured TSMCs. TSMCs grown on glass cover- slips were loaded with 5 lxM fura 2-AM at 37 °C for 45 min, washed with PBS, and incubated with PMA (1 ~u14), staurosporine (STA, 1 gM), STA plus PMA, or 4ct-phorbol 12,13-didecanoate (4ct-PDD, 1 gM) at 37 ° C for 30 rain. Fluorescent measurement of [Ca2+]i was carried out in a dual-excitation-wavelength spectrofluorometer, with excitation at 340 and 380 nm when carbachol (100 gM) was added. Values are expressed as the mean + SE from eight separate experi- ments. * p < 0.001 compared with the control.
Fig. 8. Changes in PKC activity in canine TSMCs incubated with PMA for various periods. Cuitured cells were treated with PMA (1 gM) for the indicated times. The cytosolic (e) and membraneous (©) fractions were assayed as described under Materials and Meth- ods. The data are expressed as the mean -+ SE of four separate exper- iments determined in triplicate.
cells, 0.78 + 0.10 pmol/min/mg protein). Thereafter, the membrane-associated PKC activity decreased to 0.94 + 0.02 pmol/min/mg protein after 2 h of incubation with PMA and persisted for at least 24 h of PMA treatment. In contrast, PMA induced a sustained suppression of PKC activity in the cytosolic fraction.
Effect of PMA on the Density and Affinity of the Muscarinic Receptor Long-term incubation with phorbol esters has been
reported to cause a down-regulation of mAChRs in some types of cells [20, 28]. To determine whether the observed PMA-induced inhibition of responses stimulated by car- bachol represented and actual decrease in the number of binding sites or a change in the Kd of the mAChRs for [3H]NMS, complete saturation binding curves were ob- tained for parallel control cultures and cultures incubated with 1 gM PMA for either 2 or 24 h (table 2). Scatchard analysis showed that control cells had a single class of binding site with a Bmax of 305 - 60 fmol/mg protein and a K4 of 473 + 25 pM. No significant difference in the
Bmax and Ka of the mAChRs was obtained with cells treated with PMA for either 2 or 24 h, as compared to control cells (table 2).
Discussion
A rapid PI breakdown after receptor activation has been observed in several tissues in response to stimuli such as neurotransmitters, growth factors, hormones and light [24], In TSM, the binding of agonists to mAChRs results in the generation of IP3 which leads to a release of Ca 2+ from the internal stores, and triggers muscle contrac- tion [30, 32]. The other product of PI hydrolysis, DAG, acts as a second messenger via activation of PKC, which, in addition to influencing many cellular responses, is involved in the attenuation of the PI response itself [ 19]. In this study, we have shown that the phorbol ester PMA blocks muscarinic-receptor-mediated PI hydrolysis and mobilization of intracellular Ca 2+ in cultured canine TSMCs (fig. 2, 5). The concomitant loss ofhormone-stim-
289

ulated IPs accumulation and Ca 2+ release induced by short-term PMA treatment supports the causal relation- ship between these responses as suggested by previous studies [7, 19, 20, 23]. In canine TSMCs, IPs accumula- tion is a very early receptor-stimulated event. Our data are consistent with the conclusion that IPs accumulation is a direct consequence of activation of receptors in TSMCs [13, 18], and that it is the inhibition of this response by PMA that prevents mobilization of Ca 2+ from its internal stores [ 13, 18].
Because PKC activation is associated with several cel- lular responses, phorbol-ester-mediated inhibition of IP3 formation might occur at one or several different sites. In a number of cell types, elevation of intracellular Ca 2+ by Ca2+-mobilizing agonists known to act by receptor- mediated stimulation of PI turnover has been shown to be inhibited by phorbol esters [7, 13, 18-201. It has been suggested that protein phosphorylation mediated by in- teraction of phorbol esters with PKC may be the mecha- nism by which PMA modulates hormone-sensitive PI metabolism. According to some reports [8, 14], phorbol esters might attenuate a rise in IP3 through increasing its degradation by activation of a phosphomonoesterase specific for IP3. The activity of this cytosolic enzyme increases after phosphorylation by PKC, which provides a mechanism for inhibiting the agonist-induced rise in IP3 accumulation in platelets. Phorbol esters that do not bind to or activate PKC do not block carbachol-stimulat- ed IPs accumulation [12]. Our finding that PMA rapidly inhibits the carbachol-stimulated Ca 2+ mobilization is consistent with the view that PMA acts through activa- tion of PKC, since staurosporine, a potent PKC inhibi- tor, blocks the inhibitory effect of PMA (fig. 7). More- over, long-term treatment with phorbol ester enhances angiotensin-II-stimulated PI hydrolysis in vascular smooth muscle cells [23]. This may be due to altered total incorporation and uptake of [3H]inositol into cells and inositol phospholipids.
Exposure of the cells to PMA rapidly decreased PKC in the cytosol (from 1.09 ± 0.08 to 0.63 ± 0.09 pmol/min/ mg protein), but increased it transiently in the mem- branes (from 0.78 + 0.10 to 2.05 + 0.04 pmol/min/mg protein), resulting in a decrease of the total PKC activity with exposure longer than 2 h (fig. 8). This result is consis- tent with the described dual action of PMA on PKC activ- ity. Phorbol esters are shown to activate PKC but also increase the rate of degradation [33] with possible differ- ences in sensitivities among isoforms [19].
PMA inhibited carbachol-induced IPs accumulation in cultured canine TSMCs in a time- and concentration-
dependent manner (fig. l, 2). We found that PMA did not affect the basal level of IP3, thus ruling out the possibility that PMA caused its effect by depleting an agonist-sensi- tive pool of membrane PI. In addition, PMA did not change the resting level of [Ca2+]i, as monitored with the fluorescent indicator fura 2. The inhibitory effect of PMA on carbachol-induced Ca 2+ mobilization appears to be directly related to the inhibition of IP3 formation.
One of the targets at which hormone-stimulated PI hydrolysis could be inhibited by PMA is the mAChR. It has been reported that phorbol esters induced phosphory- lation of cq-adrenergic receptors associated with antago- nism of PI breakdown in DDT1-MFz cells and suggested that altered receptor binding may be a mechanism of the PKC-induced inhibitory effect [15]. However, in our study, pretreatment of TSMCs with PMA for either 2 or 24 h did not change the Bmax and Kd of mAChRs in canine TSMCs (table 2). The receptor seems not to be a target for the inhibitory effect of PMA on muscarinic responses. The ability of PMA to block histamine-stimulated IPs accumulation also suggests that the effect of PMA is on a more general component of the PI cycle than on a specific receptor [18]. It is possible that PMA affects an unknown transducer or a known but undefined transducer that cou- ples receptor occupation to the IP/Ca 2+ response. Several lines of evidence suggest that a target distal to the receptor is the best unifying hypothesis for the location of the phor- bol ester inhibitory effect. (1) Phorbol esters block vaso- pressin-induced IP3 accumulation in A10 cells without changing receptor binding and abolish the guanine nu- cleotide shift, indicating that coupling of the receptor to Gp has been altered [t]. (2) We have also found that PMA blocks both agonist- and A1F~-stimulated IPs accumula- tion in canine TSMCs [31]. Since PMA has no effect on the basal level of PI turnover, PKC can uncouple the G protein from PLC. (3) It has been shown that activation of PKC affects the G protein coupling process in neutrophils [ 17] and inhibits the function of Gi protein in platelets [ 11]. In addition, activation of PKC by phorbol esters has been shown to phosphorylate PI-PLC in rat basophilic leukemia cells, indicating an additional mechanism for receptor-PLC uncoupling [4]. Regardless of the precise mechanism, one implication of a target distal to the recep- tor in phorbol ester inhibition is that other agonists that act through PI-PLC stimulation might be uncoupled from cytosolic Ca 2+ release.
The inhibition of the carbachol-stimulated increase of [Ca2+]i by PMA (fig. 5, 6) is in agreement with the inhibi- tory effect of PMA on agonist-induced IPs accumulation and Ca 2+ mobilization in TSMCs [13, 18]. The fact that
290 Yang/Hsu/Ong/Hsieh/Tsao/Chen/Luo Protein Kinase C and Signal Transduction

the same experimental conditions block IP3 accumulation and Ca 2+ release suggest that the mechanism &this PKC- mediated inhibition is not confined to the IP3-sensitive Ca 2+ release site. This is consistent with the observation that the purified IP3 receptor from brain is phosphorylat- ed and not functionally modified by PKC [26].
In conclusion, we have demonstrated that short-term PMA treatment causes a negative feedback regulation on agonist-induced IPs accumulation and Ca 2+ mobilization in canine TSMCs, while long-term PMA treatment is associated with augmenting responses to agonists, per- haps due to down-regulation of PKC and the loss of its inhibitory function. These results suggest that physiologi- cal activation of PKC might serve as a modulator of cellu- lar responses induced by IP3. The site of PMA inhibition appears to be at a postreceptor location and may be
involved in PI-PLC itself, although several sites of PMA antagonism cannot be excluded. To determine how DAG and other second messengers modulate agonist-induced cellular responses is important for clarifying the mecha- nisms underlying bronchial hyperreactivity and manipu- lating the contractile response of the airway in asthma.
Acknowledgments
This work was supported by grant CMRP-340 from Chang Gung Medical Research Foundation and NSC83-0412-B 182-026 from the National Science Council, Republic of China. The authors are greatly indebted to Dr. Anthony Herp at Chang Gung Medical College for his critical reading of the manuscript and suggestions. Appreciation is also expressed to Dr. Delon Wu for his encouragement.
References
1 Aiyar N, Nambi P, Whitman M, Stassen FL, Crooke ST. Phorbol-ester mediated inhibition ofvasopressin and beta-adrenergic responses in a vascular smooth muscle line. Mol Pharmacol 31:180-184;1987.
2 Barnes PJ. Modulation of neurotransmission in airways, Physiol Rev 72:699-729;1992.
3 Barthelson RA, Jacoby DB, Widdicombe JH. Regulation of chloride secretion in dog tracheal epithelium by protein kinase C. Am J Physiol 253:C802-C808;1987.
4 Bennett CF, Crooke ST. Purification and char- acterization of a phosphoinositide-specific phospholipase C from guinea pig uterus: Phos- phorylation by protein kinase C in vitro. J Biol Chem 262; 13789-13797; 1987.
5 Berridge MJ, Dawson RMC, Downes CP, Hes- lop JP, Irvine RF. Changes in the levels of ino- sitol phosphates after agonist-dependent hy- drolysis of membrane phosphoinositides. Bio- chem J 212:473-482; 1983.
6 Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248-254; 1976.
7 Castagna M, Takai Y, Kaibuchi K, Sato K, Kikkiawa U, Nishizuka Y. Direct activation of calcium-activated phospholipid-dependent protein kinase C by tumor promoting phorbol esters. J BiN Chem 257:7847-7851; 1982.
8 Cormolly TM, Lawing JF, Majerus PW. Pro- tein kinase C phosphorylates human platelet inositol trisphosphate 5'-phosphomonoester- ase, increasing the phosphatase activity. Cell 46:951-958;1986.
9 Gown AM, Vogel AN, Gordon D, Lu PL. A smooth muscle-specific monoclonal antibody recognizes smooth muscle actin isozymes. J Cell Biol 100:807-813;1985.
I0 Grynkiewicz G, Poenie M, Tsien RY. A new generation of Ca 2+ indicators with improved fluorescence properties. J Biol Chem 260: 3440-3450; 1985.
11 Katada T, Gilman AG, Watanabe Y, Bauer S, Jacobs KH. Protein kinase C phosphorylates the inhibitory gnanine-nucleotidc-binding reg- ulatory component and apparently suppresses its function in hormonal inhibition of adenyl- ate cyclase. Eur J Biochem 151:431-437; 1985.
12 Kikkawa U, Minakuchi R, Takai Y, Nishizuka Y. Calcium-activated, phospholipid-depen- dent protein kinase (protein kinase C) from rat brain. Methods Enzymo199:288-298; 1983.
13 Kotlikoff MI, Murray RK, Reynolds EE. Hista- mine-induced calcium release and phorboI an- tagonism in cultured airway smooth muscle cells. Am J Physio1253:C561-C565; 1987.
14 Lapetina EG, Reep B, Watson SP. Ionophore A23187 stimulates phosphorylation of the 40,000 dalton protein in human platelets with- out phospholipase C activation. Life Sci 39: 751-759;1986.
15 Leeb-Lundberg LMF, Cotecchia S, Lomasney JW, Debernardis JF, Lefkowitz R J, Caron MG. Phorbol esters promote alpha~-adrenergic re- ceptor phosphorylation and receptor uncou- pling from inositol phospholipid metabolism. Proc Natl Acad Sci USA 82:5651-5655;1985.
16 McCarthy SA, Hallam TJ, Merritt JE. Activa- tion of protein kinase C in human neutrophils attenuates agonist-stimulated rises in cytosolic free Ca 2+ concentration by inhibiting bivalent- cation influx and intmcellular Ca 2+ release in addition to stimulating Ca 2+ efflux. Biochem J 264:357-364;1989.
17 Matsttmoto T, Molski TFP, Pelz C, Kanaho Y, Becker EL, Feinstein MB, Naccahe PH, Sha'afi RI. Treatment of rabbit neutrophils with phor- bol esters result in increased ADP-ribosylation catalyzed by pertussis toxin and inhibition of the GTPase stimulated by fMet-Leu-Phe. FEBS Lett 198;295-300; 1986.
18 Murray RK, Bennett CF, Fluharty S J, Kotli- koff MI. Mechanism of phorbol ester inhibi- tion of histamine-induced IP3 formation in cul- tured airway smooth muscle. Am J Physiol 257:L209-L216; 1989.
19 Nishizuka Y. The molecular heterogeneity of protein kinase C and implications for cellular regulation. Nature 334:661-665; 1988.
20 Orellana SA, Solski PA, Brown JH. Guanosine 5'-O-(thiotrisphosphate)-dependent inositol trisphosphate formation in membranes is in- hibited by phorbol ester and protein kinase C. J Biol Chem 260:5236-5239;1985.
21 Osugi T, Imaizumi T, Mizushima A, Uchida S, Yoshida H. Phorbol ester inhibits bradykinin- stimulated inositol trisphosphate formation and calcium mobilization in neuroblastoma x glioma hybrid NG108-15 cells. J Pharmacol Exp Ther 240:617-622; 1987.
22 Pearce B, Morrow C, Murphy S. Characteris- tics of phorbol ester- and agonist-induced down-regulation of astrocyte receptors coupled to inositol phospholipid metabolism. J Neuro- chem 50:936-944;1988.
291

23 Pfellschifter J, Ochsner M, Whitebread S, De Gasparo M. Down-regulation of protein kinase C potentiates angiotensin II-stimulateed poly- phosphoinositide hydrolysis in vascular smooth-muscle cells. Biochem J 262:285-291; 1989.
24 Rana RS, Hokin LE. Role of phosphoinositides in transmembrane signaling. Physiol Rev 70: 115-164;1990.
25 Somlyo AV, Bond M, Somlyo AP, Scarpa A. Inositol trisphosphate-induced calcium release and contraction in vascular smooth muscle. Proc Natl Acad Sci USA 82:5231-5235; 1985.
26 Supattapone S, Danoff SK, Theibert A, Joseph SK, Steiner J, Snyder SH. Cyclic AMP depen- dent phosphorylation of a brain inositol tris- phosphate receptor decreases its release of caI- cium. Proc Natl Acad Sci USA 85:8747-8750; 1988.
27 Willems PHGM, van Den Broek BAM, van Os CH, De Pont JJHHM. Inhibition of inositol 1,4,5-trisphosphate-induced Ca 2+ release in permeabilized pancreatic acinar cells by hor- monal and phorbol ester pretreatment. J Biol Chem 264:9762-9767; 1989.
28 Xu J, Chuang DM. Muscarinic aeetylcholine receptor-mediated phosphoinositide turnover in cultured cerebellar granule cells: Desensiti- zation by receptor agonists. J Pharmacol Exp Ther 242:238-244;1987.
29 Yang CM, Chou SP, Sung TC. Regulation of functional muscarinic receptor expression in tracheal smooth muscle cells. Am J Physiol 261:C1123-C1129;1991.
30 Yang CM, Chou SP, Wang YY, Hsieh JT, Ong R. Muscarinic regulation of cytosolic free cal- cium in canine tracheal smooth muscle cells: Ca 2+ requirement for phospholipase C activa- tion. Br J Pharmacol 110:1239-1247;1993.
31 Yang CM, Yo YL, Hsieh JT, Ong R. 5- Hydroxytryptamine receptor-mediated phos- phoinositide hydrolysis in canine cultured tra- cheal smooth muscle ceils. Br J Pharmacol 111: 777-786; 1994.
32 Yang CM, Yo YL, Wang YY. Intracellular cal- cium in canine cultured tracheal smooth mus- cle cells is regulated by M3 muscarinic recep- tors. Br J Pharmacol 110:983-988; 1993.
33 Young S, Parker P J, Ullrich A, Stabel S. Down- regulation of protein kinase C is due to an increased rate of degradation. Biochem J 244: 775-779; 1987.
292 Yang/Hsu/Ong/Hsieh/Tsao/Chen/Luo Protein Kinase C and Signal Transduct ion




















