
Competition in neuronal morphogenesis and the development of nerve connections

Current Biology, Vol. 14, 2197–2207, December 29, 2004, ©2004 Elsevier Ltd. All rights reserved. DOI 10.1016/j .cub.2004.11.056
Genetic Control of Cell Intercalationduring Tracheal Morphogenesis in Drosophila
nected to their neighbors via adherens junctions (AJs),which maintain the integrity of the tissue. At the sametime, the AJs integrate the function of multiple signals
Carlos Ribeiro,1,2 Marc Neumann,1
and Markus Affolter*Biozentrum der Universitat BaselKlingelbergstrasse 70 into a coordinated response [1].
Epithelial sheets are often remodeled into tubular net-CH-4056 BaselSwitzerland works, allowing for an efficient exchange of fluids or
gases with surrounding tissues [2, 3]. A particularly well-studied example of such a remodeling process occursduring the development of the tracheal system in theSummaryDrosophila embryo [4, 5]. The larval tracheal systemconsists of hundreds of interconnected tubes that trans-Background: Branching morphogenesis transforms anport oxygen and other gases throughout the body. Tra-epithelial sheet into a tubular network with distinct fea-cheal branches are simple tubes consisting of an epithe-tures regarding the length and diameter of individuallial monolayer wrapped around a central lumen. Thetubes. Branching is controlled by several signaling path-development of the trachea is initiated in the early em-ways, but the molecular consequences of these path-bryo upon the determination of ten bilaterally symmetri-ways in the responding cells are poorly understood.cal clusters of approximately 80 tracheal precursor cells.Results: We have undertaken a detailed characteriza-Each cluster, called a tracheal placode, subsequentlytion of cell rearrangements during tracheal branchingundergoes a similar sequence of developmental eventsmorphogenesis in Drosophila embryos with a GFP fu-to generate one segment of the network in the absencesion protein labeling the adherens junctions (AJs) andof further cell divisions. Three key cellular processes arehigh-resolution live imaging. To analyze the branchinginvolved in branch formation: directional cell migration,process at the cellular level, we further developed ancell rearrangements, and cell shape changes. To in-imaging approach that allows us to follow single cellsterconnect the individual metameres, certain tubes ulti-during the branching process. We find that controlledmately fuse with other tubes in neighboring segmentscell intercalation, which requires extensive AJ remodel-[6–8]; during all these processes, tracheal cells maintaining, is key to the formation of tracheal branches of differ-their epithelial integrity.ent cellular complexities. In particular, most branches
A number of signaling systems have been shown toconsist of tubes with individual cells wrapped around theregulate tracheal branch formation during embryogene-lumen. These branches form through cell intercalation,sis, and the challenge is to understand how these differ-which requires the transformation of most of the initialent signals coordinate branching morphogenesis at theintercellular AJs into autocellular AJs. We propose acellular and molecular level. The best understood signalstep-wise model explaining how this AJ remodeling oc-is Branchless/FGF (Bnl/FGF) [9], the ligand of thecurs and use this model to better understand defectsBreathless FGF receptor (Btl/FGFr) [10, 11]. Bnl/FGF isin various mutants. We find that Dpp and Wnt signalingexpressed in discrete groups of cells around the trachealcontrol cell intercalation by regulating the expression ofplacode and attracts tracheal cells via the activation ofSpalt, a zinc finger transcription factor; Spalt inhibitsBtl/FGFr, which is expressed in tracheal cells. Bnl/FGFintercalation, leading to the formation of large, multicel-induces the formation of dynamic filopodia in respondinglular tubes.tracheal cells at the tip of outgrowing branches [12, 13]Conclusion: Tracheal morphogenesis is regulated byand, thus, seems to control cytoskeletal activity andan interplay of different signaling systems that controlmotility. Because tracheal cells are connected with theircell migration and cell intercalation, respectively. Onlyneighbors via AJs throughout the branching process,the combined action of these signaling systems allowsthe migration of a few tip cells away from a sac-likeefficient branch elongation and the formation of mor-invagination under the control of Bnl/FGF signalingphologically distinct branches.leads to the formation of tubular branches consisting ofepithelial cells wrapped around a central lumen.Introduction
In addition to the FGF signaling system, other signalshave been shown to be involved in the development ofEpithelial remodeling is one of the most prominentspecific subsets of branches. Wnt signaling is requiredmechanisms involved in the three-dimensional shapingfor the expression of the transcription factor Sal in theof developing organisms. Remodeling relies on the spa-multicellular dorsal trunk [14, 15], and the sal gene istial regulation of cell division, cell shape changes, andessential for the formation of this particular branch [16].cell rearrangements. Cell rearrangements pose a partic-Dpp signaling is required for the formation of the fineular problem to epithelial tissues because cells are con-dorsal and ventral branches of the tracheal system andacts by inducing the expression of the transcription fac-
*Correspondence: [email protected] tors Kni/Knrl in these branches [17, 18]. In the absence1These authors contributed equally to this work.of Dpp, Bnl/FGF-induced filopodia formation, cell move-2Present address: Institute of Molecular Biotechnology (IMBA), Aus-ment, and dorsal branch outgrowth are initially normal,trian Academy of Sciences, Dr. Bohr-Gasse 3–5, A-1030 Vienna,
Austria. but during later stages, branch outgrowth stalls, and the

Current Biology2198
cells reintegrate into the dorsal trunk, leading to the like structure around the lumen, whereas autocellularAJs appear as single lines lining the lumen (Figure 1A).complete absence of dorsal branches [12]. It is not
As shown in Figures 1B–1F (for interactive 3D models,known what cellular processes necessary for the forma-see the Supplemental Data available with this articletion of distinct branches are controlled by Dpp and Wntonline), the only embryonic tracheal tube containing ex-signaling.clusively intercellular AJs is the dorsal trunk. The trans-To gain insight into the basic cell behavior duringverse connective branch, the structure that links thetracheal morphogenesis, with the ultimate goal to learndorsal trunk to the more lateral and ventral branches,more about the integration of the distinct signaling in-displays a mixed character; it contains zones with multi-puts in epithelial remodeling, we have undertaken a de-ple cells making up the luminal circumference, and thesetailed characterization of the cell rearrangements thatzones are interspersed with regions containing autocel-occur during tracheal tube formation with a GFP fusionlular AJs—especially those cells that link the transverseprotein labeling the AJ and high resolution live imaging.connective to the dorsal trunk adopt a cellular shapeWe describe the different steps of the intercalation pro-with autocellular AJs (see arrow in Figure 1B). All thecess that transforms a cellular arrangement from a side-other branches, including the lateral trunk, the gangli-by-side to an end-to-end configuration, a transformationonic and dorsal branches, and most branches in thethat is accompanied by a dramatic exchange of intercel-head region show a predominance of autocellular AJs,lular AJs with autocellular AJs. Furthermore, we devel-with cells remaining linked to their neighbors only viaoped a method to follow single tracheal cells in vivonarrow, ring-like intercellular AJs (summarized in Figureduring intercalation in order to verify our model at the1G).single-cell level. We then investigate the role of Dpp and
Because the tracheal metameres develop from a sac-Wnt signaling in the regulation of AJ remodeling andlike invagination of the ectoderm, which itself is an epi-cell intercalation during branch formation. We find thatthelial sheet in which all cell-cell AJ contacts are formedthe Wnt-activated gene sal is required and sufficient tobetween adjacent cells, tracheal branching morphogen-inhibit cell intercalation and the formation of autocellularesis relies to a large extent on dramatic cell rearrange-AJs; Sal-expressing cells are therefore part of multicellu-ments that must be accompanied by the exchange oflar branches. In turn, Dpp signaling is required for cellmost of the intercellular AJs with autocellular AJs.intercalation in some branches, but its major function
with regard to intercalation is the downregulation of saltranscription and, therefore, the release of the inhibition
High-Resolution Live Imaging of Cell Rearrangementsof cell rearrangements. Our study integrates the functionduring Branch Morphogenesisof three different signaling systems in the control of cellTo gain insight into possible mechanisms controlling
rearrangements, ultimately leading to the formation ofthese cell rearrangements and the underlying AJ remod-
branches with distinct morphologies.eling events, we used high-resolution live imaging tofollow the dynamics of the AJs of individual branchesduring their outgrowth. We focused our attention on the
Resultsformation of fine tubes with autocellular AJs in the dorsalbranch. We have previously described this process, our
Characterization of the Cellular Architecture focus being the termination of the intercalation processof the Different Tracheal Tubes [21]. At the onset of dorsal branch formation, cells arein the Late Drosophila Embryo aligned in a side-by-side arrangement, and all AJs areThe tracheal system of a late Drosophila embryo con- intercellular. This alignment becomes destabilized, pos-sists of tubes of different cellular complexity and diame- sibly by the pulling force generated by the dorsal move-ter [19, 20]. Three types of tubes have been described: ment of the leading cells toward Bnl/FGF, forcing thelarge, multicellular tubes with intercellular AJs; fine cells to shift their relative position and to intercalate.tubes consisting of single cells wrapped around the Intercalation is accompanied by individual cells reach-luminal space and sealed by autocellular AJs; and intra- ing around the entire lumen and thereby forming thecellular tubes without AJ complexes lining the lumen. first autocellular AJ contacts. As the cells move dorsally
We have previously made use of a D�-Catenin-GFP and elongate, intercellular AJs of adjacent cells are suc-(D�-Cat-GFP) fusion protein specifically expressed in cessively replaced by autocellular AJs of tube-shapedthe developing trachea to outline the cellular architec- cells. Finally, the elongated cells adopt an end-to-endture of this organ [21]. This fusion protein has been arrangement, and the only intercellular AJs that connectshown to be targeted specifically to the apically located individual tube cells are represented by small rings ofcell-cell AJs in living Drosophila embryos [22], and it D�-Cat-GFP around the luminal space (see [21]).therefore permits the visualization of the apical outline From this initial description of the process, it is con-of each individual cell. In order to gain insight into how ceivable that the most critical step in the transition ofthe development of structurally different tubes is con- large, multicellular branches with intercellular AJs to finetrolled, we first determined which of the six main branches with autocellular AJs involves the generationbranches forming the segmental outline of the individual of the first contact of a cell with itself—that is, the forma-tracheal metameres were made up of tubes containing tion of the first autocellular AJ contact. We have followedintercellular AJs and which branches were made up this process at high temporal and spatial resolution,of tubes with autocellular AJs. With D�-Cat-GFP as a and Figure 2A shows individual snapshots from a 4D
confocal microscopy analysis, accompanied by amarker, intercellular AJs appear as a mesh- or ladder-

Cell Intercalation in Tracheal Morphogenesis2199
Figure 1. Adherens Junctions (AJs) in the Late Embryonic Tracheal System
(A) Schematic representation of autocellular (green) and intercellular (red) AJs.(B–F) Three-dimensional reconstructions of living stage 16 embryos expressing D�-Cat-GFP under the control of the trachea-specific btl-Gal4driver. (B) Dorsal, (C) lateral, and (D) ventral views of central metameres are shown. (E) First and (F) tenth tracheal metamere are shownseparately.(G) Schematic overview of the different patterns of AJ in the tracheal system. Intracellular tubes are shown in blue, autocellular AJs in green,and intercellular AJs in red. The TC is dashed in green and red because it can consist of both types of AJs. The following abbreviations wereused: DB, dorsal branch; DT, dorsal trunk; TC, transverse connective; FB, fat body branch; VB, visceral branch; SB, spiracular branch; LT,lateral trunk; GB, ganglionic branch; LG, lateral G branch; LH, lateral H branch; LT, lateral T branch; DB1, dorsal branch of the first metamere;DC, dorsal cephalic branch; SB1, spiracular branch of the first metamere; LC, lateral cephalic branch; VC, ventral cephalic branch; GB0,unusual ganglionic branch; GB1, ganglionic branch of the first metamere; PB, pharyngeal branch; CB, cerebral branch; PSP, posterior spiracle;LP, lateroposterior branch; SB10, spiracular branch of the tenth metamere; and CH, caudal-hindgut branch. For interactive 3D Quick TimeVirtual Reality models of panels (B)–(F), see the Supplemental Data. All images are anterior to the left and dorsal to the top. Nomenclatureand diagram according to [5].
graphic representation of how we interpret what we visu- does so with the distal side, remains open (see alsoDiscussion).alize in vivo (in Figure 2B, we have colored the AJs
of two adjacent cells, whereas in Figure 2C, a graphicrepresentation of these two cells during the intercalation Cell Rearrangements Analyzed
at the Single-Cell Levelprocess is shown). After pairing of adjacent cells (Fig-ures 2A–2C; 0 min), one of the two cells appeared to The model we propose for how cell rearrangements take
place during the formation of tubes with autocellularreach around the lumen with its distal end, bringing itstwo intercellular AJs, which at the onset of the process AJs is based on the analysis of the localization of D�-Cat-
GFP in all tracheal cells and our interpretation of howrun parallel and along the luminal space, closer together,until they met at the opposing side of the lumen (Figures individual cells fit into this meshwork of AJ complexes.
This model makes a number of predictions of how the2A–2C; 12 min, arrow). This close apposition of junc-tional complexes of the same cell must be a prerequisite AJs of an individual cell are remodeled during cell inter-
calation. At the onset of the intercalation process, thefor the formation of autocellular AJ. Soon after the cellreached around the lumen, a single line of AJ complexes AJs of an individual cell in the dorsal branch would form
a single loop because they represent AJ connections toformed out of the contact point where the intercellularAJ first met (Figures 2A–2C; 16.5 min, see arrow). We the neighboring cells (Figure 3C). When this cell reaches
around the lumen to generate the initial autocellular AJinterpret the appearance of this line as the continuationof autocellular AJ formation from the initial contact point. contact, a two-loop structure is formed, consisting of a
small loop and a much larger loop (Figure 3D). DuringThis short stretch of autocellular AJ became longer untilthe entire cell body was sealed up along the tube with the subsequent transformation of intercellular AJs into
autocellular AJs, the part of the large loop that contactsautocellular AJs; only small, ring-like intercellular AJ be-tween adjacent cells stabilized the end-to-end arrange- the small loop is progressively transformed into a single
line, an autocellular AJ (Figure 3E). This process contin-ment (Figure 2C, Termination). The other cell of the pair-wise alignment underwent a similar rearrangement of ues until two small loops of intercellular AJs surrounding
the lumen are left, the loops being connected by a singleAJ, but it reached around the lumen with its proximalend. How the two paired cells decide which one reaches line of autocellular AJs (Figure 3F).
To test these predictions, we developed an assayaround the lumen with the proximal side, and which one
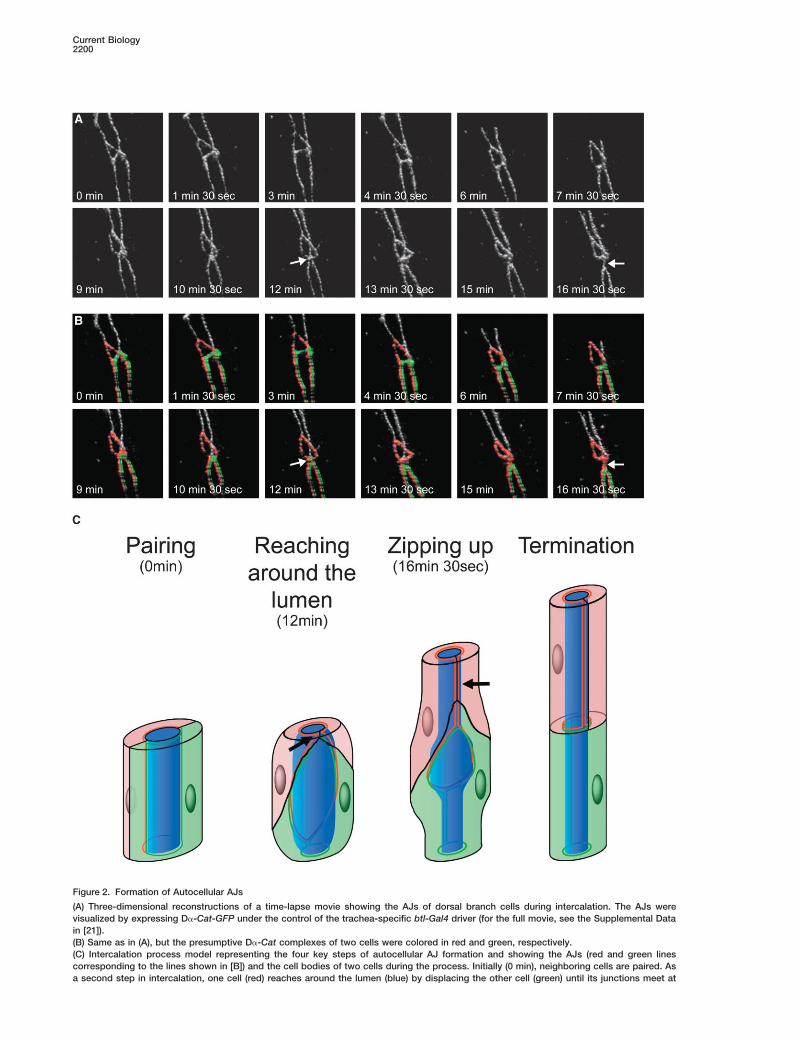
Current Biology2200
Figure 2. Formation of Autocellular AJs
(A) Three-dimensional reconstructions of a time-lapse movie showing the AJs of dorsal branch cells during intercalation. The AJs werevisualized by expressing D�-Cat-GFP under the control of the trachea-specific btl-Gal4 driver (for the full movie, see the Supplemental Datain [21]).(B) Same as in (A), but the presumptive D�-Cat complexes of two cells were colored in red and green, respectively.(C) Intercalation process model representing the four key steps of autocellular AJ formation and showing the AJs (red and green linescorresponding to the lines shown in [B]) and the cell bodies of two cells during the process. Initially (0 min), neighboring cells are paired. Asa second step in intercalation, one cell (red) reaches around the lumen (blue) by displacing the other cell (green) until its junctions meet at
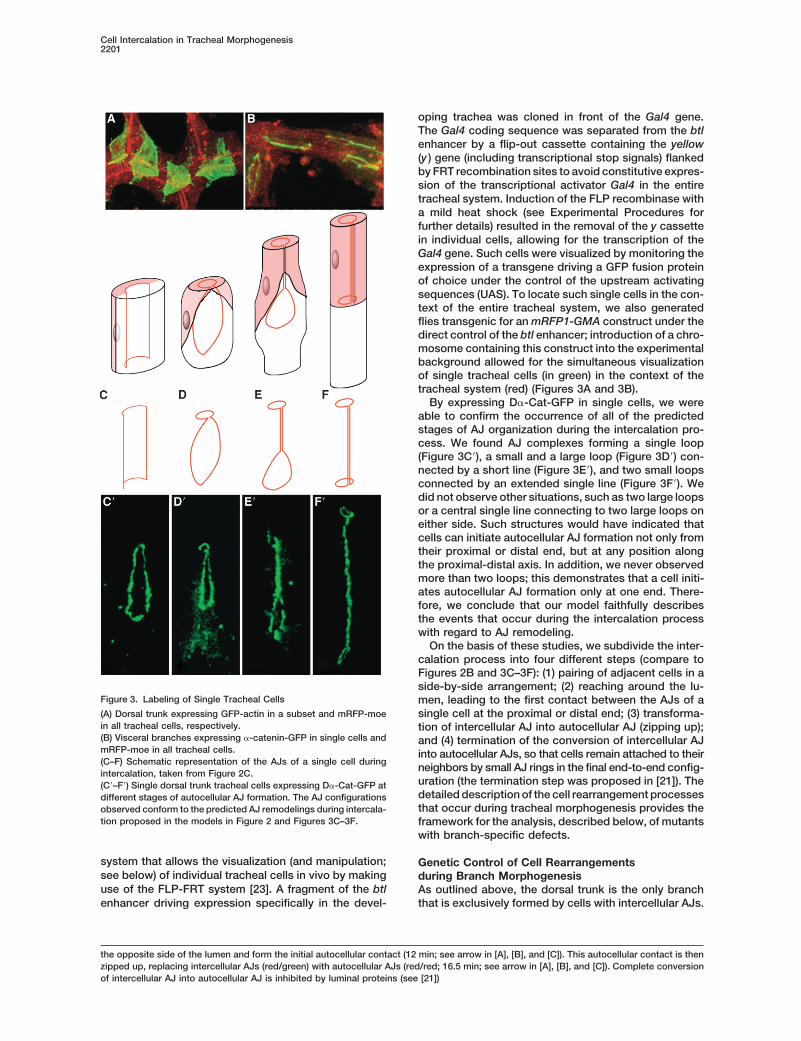
Cell Intercalation in Tracheal Morphogenesis2201
oping trachea was cloned in front of the Gal4 gene.The Gal4 coding sequence was separated from the btlenhancer by a flip-out cassette containing the yellow(y ) gene (including transcriptional stop signals) flankedby FRT recombination sites to avoid constitutive expres-sion of the transcriptional activator Gal4 in the entiretracheal system. Induction of the FLP recombinase witha mild heat shock (see Experimental Procedures forfurther details) resulted in the removal of the y cassettein individual cells, allowing for the transcription of theGal4 gene. Such cells were visualized by monitoring theexpression of a transgene driving a GFP fusion proteinof choice under the control of the upstream activatingsequences (UAS). To locate such single cells in the con-text of the entire tracheal system, we also generatedflies transgenic for an mRFP1-GMA construct under thedirect control of the btl enhancer; introduction of a chro-mosome containing this construct into the experimentalbackground allowed for the simultaneous visualizationof single tracheal cells (in green) in the context of thetracheal system (red) (Figures 3A and 3B).
By expressing D�-Cat-GFP in single cells, we wereable to confirm the occurrence of all of the predictedstages of AJ organization during the intercalation pro-cess. We found AJ complexes forming a single loop(Figure 3C�), a small and a large loop (Figure 3D�) con-nected by a short line (Figure 3E�), and two small loopsconnected by an extended single line (Figure 3F�). Wedid not observe other situations, such as two large loopsor a central single line connecting to two large loops oneither side. Such structures would have indicated thatcells can initiate autocellular AJ formation not only fromtheir proximal or distal end, but at any position alongthe proximal-distal axis. In addition, we never observedmore than two loops; this demonstrates that a cell initi-ates autocellular AJ formation only at one end. There-fore, we conclude that our model faithfully describesthe events that occur during the intercalation processwith regard to AJ remodeling.
On the basis of these studies, we subdivide the inter-calation process into four different steps (compare toFigures 2B and 3C–3F): (1) pairing of adjacent cells in aside-by-side arrangement; (2) reaching around the lu-
Figure 3. Labeling of Single Tracheal Cells men, leading to the first contact between the AJs of a(A) Dorsal trunk expressing GFP-actin in a subset and mRFP-moe single cell at the proximal or distal end; (3) transforma-in all tracheal cells, respectively. tion of intercellular AJ into autocellular AJ (zipping up);(B) Visceral branches expressing �-catenin-GFP in single cells and and (4) termination of the conversion of intercellular AJmRFP-moe in all tracheal cells. into autocellular AJs, so that cells remain attached to their(C–F) Schematic representation of the AJs of a single cell during neighbors by small AJ rings in the final end-to-end config-intercalation, taken from Figure 2C.
uration (the termination step was proposed in [21]). The(C�–F�) Single dorsal trunk tracheal cells expressing D�-Cat-GFP atdetailed description of the cell rearrangement processesdifferent stages of autocellular AJ formation. The AJ configurationsthat occur during tracheal morphogenesis provides theobserved conform to the predicted AJ remodelings during intercala-
tion proposed in the models in Figure 2 and Figures 3C–3F. framework for the analysis, described below, of mutantswith branch-specific defects.
system that allows the visualization (and manipulation; Genetic Control of Cell Rearrangementssee below) of individual tracheal cells in vivo by making during Branch Morphogenesisuse of the FLP-FRT system [23]. A fragment of the btl As outlined above, the dorsal trunk is the only branch
that is exclusively formed by cells with intercellular AJs.enhancer driving expression specifically in the devel-
the opposite side of the lumen and form the initial autocellular contact (12 min; see arrow in [A], [B], and [C]). This autocellular contact is thenzipped up, replacing intercellular AJs (red/green) with autocellular AJs (red/red; 16.5 min; see arrow in [A], [B], and [C]). Complete conversionof intercellular AJ into autocellular AJ is inhibited by luminal proteins (see [21])

Current Biology2202
Figure 4. Genetic Control of AJ Remodeling Events during Tracheal Branch Formation
Three-dimensional reconstruction of living embryos expressing D�-Cat-GFP under the control of the trachea-specific btl-Gal4 driver.(A) Overview of the tracheal AJs of a wild-type embryo.(B) Ectopic, pantracheal expression of sal inhibits the formation of autocellular AJs.(C) In a mutant lacking sal and spalt-related (salr), stretches of autocellular AJs are found throughout the entire tracheal system. Ectopic,pantracheal overexpression of (D) an activated form of tkv or (E) kni induces autocellular AJs in the DT. Insets are magnifications of the boxedregions.(F) In mutants lacking tkv, autocellular AJs are still formed. For interactive 3D Quick Time Virtual Reality models of panels (A)–(F), seeSupplemental Data. All images are anterior to the left and dorsal to the top. The following abbreviations were used: DB, dorsal branch; DT,dorsal trunk; GB, ganglionic branch; TC, transverse connective; and VB, visceral branch.
Previous genetic studies have shown that dorsal trunk around the lumen or that the initial autocellular AJ con-tacts were not stabilized and zipped up.formation requires the nuclear zinc finger protein Sal
[16]. Although Sal is first detected in all dorsal tracheal Conversely, embryos homozygous for a deficiencyremoving sal and spalt-related (salr) displayed autocel-cells, its expression pattern refines during branch forma-
tion, and Sal protein is maintained exclusively in the lular AJs throughout the tracheal system (Figure 4C)and a lack of the multicellular dorsal branch. However,cells of the dorsal trunk through a process that requires
Wnt signaling [14, 15]. Although the removal of Sal re- because the complete removal of sal/salr results in atransformation of the dorsal tracheal placode into a ven-sults in the absence of the large, dorsal trunk structure
[16], ectopic expression of Sal has been shown to inter- tral placode [24] and, therefore, in a lack of dorsalbranches (these branches are transformed to ventralfere with the outgrowth of dorsal branches [18]. There-
fore, we were interested to see whether Sal is required branches), we were looking for a way to reduce salfunction only after the formation of the tracheal placodesfor the formation or maintenance of tubes with intercellu-
lar AJs and whether ectopic expression of Sal would and, thus, after the dorsal/ventral patterning of the plac-ode. It has previously been shown that sal transcriptioninterfere with autocellular AJ formation, a characteristic
feature of those branches that do not express the Sal is repressed by the Kni and Knrl proteins, which areinduced by Dpp signaling in dorsal and ventral trachealproteins.
Indeed, when Sal was ectopically expressed in the branches [18]. Therefore, we asked whether activatingDpp signaling (through the constitutive active Dpp typeentire tracheal system, autocellular AJ formation was
completely blocked, and all branches were transformed I receptor TkvQD; [25]) or expressing Kni in the dorsaltrunk (and thereby repressing sal expression) would re-into tubes with cells organized in a paired arrangement
and connected via intercellular AJs (Figure 4B). Close sult in altered cell rearrangements and lead to the trans-formation of intercellular AJs into autocellular AJs in theinspection of the AJ configuration of these branches
indicated that cells were either not capable of reaching dorsal trunk. Indeed, we found that both conditions,

Cell Intercalation in Tracheal Morphogenesis2203
Figure 5. Modification of Sal Levels in a Sub-set of Tracheal Cells
(A and D) Antibody staining for Sal (red), theluminal antigen 2A12, and GFP (green). Modi-fied cells are labeled with actin-GFP.(B, C, E, and F) Live embryos; modified cellsare labeled with D�-Cat-GFP. All trachealcells are labeled with mRFP-moe. The follow-ing abbreviations were used: DT, dorsal trunk,and TC, transverse connective.(A) Overexpression of sal in single cells re-sults in Sal levels that are comparable to thewild-type expression in the dorsal trunk. Ec-topic expression of sal in (B) a single gangli-onic branch cell or (C) a group of lateral trunkcells is shown. (B) Single cells expressing salectopically still intercalate and form autocel-lular AJs. (C) Small groups of cells expressingsal ectopically generally retain intercellularAJs but are frequently flanked by cells withautocellular AJs at the periphery.(D) Overexpression of tkvQD in a subset of dor-sal trunk cells results in almost complete
downregulation of Sal. The one Sal-expressing cell that appears to be inside the clone is in another focal plane and does not express tkvQD.Ectopic expression of tkvQD in (E) a single dorsal trunk cell or (F) a group of dorsal trunk cells can lead to ectopic formation of autocellularAJs in the dorsal trunk.
ectopic activation of the Dpp signaling pathway (Figure often formed autocellular AJs (in three groups out of theseven groups; an example is shown in Figure 5C). This4D) and ectopic expression of Kni (Figure 4E), resulted
in the formation of fine dorsal trunks containing suggests that individual cells can undergo autocellularAJ formation whenever they are paired with a neighborstretches of autocellular AJs. Taken together, these re-
sults show that Sal is both required and sufficient to that does not express Sal.We then asked whether downregulation of Sal in cellsinhibit the formation of autocellular AJs during tracheal
development. of the dorsal trunk through the expression of TkvQD (Fig-ure 5D) would lead to autocellular AJ formation. Indeed,we observed the formation of autocellular AJs by down-Sal Does Not Act Cell Autonomouslyregulation of Sal in single cells (Figure 5E) or groups ofBecause Sal inhibits cell rearrangements and autocellu-cells (Figure 5F), but only at a low frequency (only 1 outlar AJ formation, we wanted to analyze whether Sal actsof 18 single modified cells displayed autocellular AJscell autonomously, inhibiting a cellular process essentialand only 1 out of 10 groups of cells). This is probablyfor the formation of autocellular AJs, or whether Saldue to the fact that several cells make up the circumfer-influences epithelial junction remodeling in a way thatence of the dorsal trunk, and therefore, in most cases,requires Sal to be present in multiple cells. We usedgroups of nonmodified, Sal-expressing cells interferethe newly developed experimental approach describedwith the formation of autocellular AJs.above and modified Sal levels in single tracheal cells or
Taken together, these results demonstrate that Salin groups of cells and asked whether such alterationsneeds to be expressed in a group of cells to exert itswould result in a modified cellular behavior. We investi-inhibitory effect on autocellular AJ formation.gated whether ectopic Sal expression inhibited autocel-
lular AJ formation in dorsal and ventral branches andwhether downregulation of sal via cell-autonomous acti- Efficient Dorsal Migration of Tracheal Cells Requires
the Elimination of sal Functionvation of the Dpp signaling pathway would result inautocellular AJ formation in the dorsal trunk. We have previously reported that dorsal tracheal cells
respond to Bnl/FGF and form a bud-like outgrowth butWith our modified FLP/FRT system, Sal was ectopi-cally expressed at levels equal or higher than endoge- later reintegrate into the dorsal trunk in the absence of
Dpp signaling [12]. These findings demonstrated that thenous Sal (Figure 5A). When we analyzed the distributionof D�-Cat-GFP, we found that single Sal-expressing lack of a dorsal branch in the absence of Dpp signaling is
due not to interference with Bnl/FGF signaling, but to acells in either dorsal or ventral branches still formedextended autocellular AJs (ten out of ten cases) (Figure different, ill-characterized role of Dpp signaling in dorsal
branch outgrowth. The experiments reported in this5B), demonstrating that Sal does not block a processnecessary for intercalation and autocellular AJ formation study allow for an elegant explanation of this finding. In
the absence of Dpp signaling, Sal protein is present inin a cell-autonomous fashion. When Sal was ectopicallyexpressed in groups of cells in the dorsal or ganglionic all dorsal tracheal cells; because Sal inhibits cell interca-
lation in groups of cells, and its presence does not allowbranch, cells were generally inhibited in their capacityto form autocellular AJs and formed cell aggregates with efficient branch elongation (see above), we speculated
that the removal of Sal might rescue dorsal branch de-intercellular AJs (seven out of seven groups); however,we found that single cells at the periphery of such groups fects in the absence of Dpp signaling; that is, the con-

Current Biology2204
Figure 6. Downregulation of sal Rescues Dorsal Branch (DB) Formation in the Absence of Dpp Signaling
The tracheal system is shown as a three-dimensional reconstruction of (A–C) fixed embryos stained with a lumen-specific antibody (red) anda GFP antibody detecting trachea-specific nuclear GFP expression (green) or (D) living embryos expressing D�-Cat-GFP under the controlof the tracheal btl-Gal4 driver.(A) Expression of the Dpp signaling inhibitor Dad in the tracheal system leads to loss of DB formation.(B) DB formation in the presence of Dad can be rescued by a Wg pathway mutation that results in the downregulation of late-tracheal Salexpression.(C) Highlight of the DB boxed in (B).(D) The cells of the rescued DB intercalate and form autocellular AJs, as visualized with D�-Cat-GFP.(E) Model depicting how different signaling pathways control branch formation. Bnl/FGF-induced pulling forces lead to the outgrowth of aninitial bud. Cell intercalation is inhibited in the dorsal trunk by sal; the expression of sal requires Wnt signaling. Kni expression is induced byDpp signaling in the dorsal-most five to six cells and suppresses sal transcription, thereby allowing cell intercalation and the formation ofautocellular AJs.
comitant removal of both Sal and Dpp signaling might observation that in tkv mutant embryos, fine ventralbranches made up of elongated cells with autocellularrescue the dorsal branch outgrowth defects. As men-
tioned above, the dorsal branches are transformed into AJ were formed despite the absence of Dpp signalingand kni/knrl expression in tracheal cells (Figure 4F).ventral branches in sal mutants, and therefore this mu-
tant background cannot be used to rescue dorsal We conclude from these findings that Sal expressionis required for the formation of multicellular tubes withbranch outgrowth. It has been shown that in armXM19
mutants, sal expression in dorsal trunk cells is strongly intercellular AJs and that it generates such tubes byrepressing cell intercalation and/or the formation of au-reduced or absent, and a dorsal trunk does not form,
whereas the early sal expression that is responsible for tocellular AJs (Figure 6E). The major and possibly solefunction of kni/knrl, with regard to cell rearrangementsthe dorso-ventral patterning of the placode is not af-
fected [14]. Whereas overexpression of Dad, which in- and the formation of autocellular AJs, consists in remov-ing Sal from dorsal branch cells and thereby allowinghibits Dpp signaling, inhibits the outgrowth of the dorsal
branches (Figure 6A), the overexpression of Dad in an them to undergo cell intercalations through the forma-tion of autocellular AJ. Dpp and Sal might have addi-armXM19 mutant, which results in concomitant removal
of Sal and Dpp signaling, results in outgrowth of dorsal tional roles in helping cells to choose their direction ofmigration and in cell fate decisions [17, 18, 26].branches (Figures 6B and 6C), cell intercalation, and
autocellular AJ formation (Figure 6D). These experi-ments demonstrate that Dpp signaling (and therefore Discussionthe expression of the Dpp response genes kni/knrl) indorsal branches is not necessary for intercalation per se Cell Behavior during the Formation of Tubes
Containing Autocellular AJsbut is required for the downregulation of Sal, which wouldotherwise suppress cell intercalation and autocellular AJ Starting from a cell field in which all AJs are shared
among adjacent cells, different tracheal tubes are formedformation. This interpretation is further supported by the

Cell Intercalation in Tracheal Morphogenesis2205
through cell rearrangements and cell intercalation re- rangements are brought about by basolateral protru-sions between pairs of adjacent cells; such protrusionsquiring extensive AJ remodeling. At the end of this pro-
cess, most of the tracheal branches consist of cells in between nonadjacent cells might be extended apicallyby cortical flow, allowing the replacement of compo-an end-to-end arrangement, resulting from a conversion
of intercellular AJs into autocellular AJs. We have under- nents of the circumferential junction belt from below [30,31]. Apical AJs and basal membranes have to be labeledtaken a detailed analysis of the cell rearrangements that
occur during tracheal branching morphogenesis in vivo, simultaneously, but with different fluorescent proteins,to follow their relative movement in time and space indescribe individual steps that accompany cell intercala-
tion, and analyze how rearrangements are regulated ge- order to find out more about how cells reach aroundthe lumen in the developing tracheal branches.netically.
The remodeling of AJ during tracheal branching mor- Once tracheal cells have reached around the lumenand the initial autocellular AJs have formed, cells un-phogenesis can be subdivided into four steps. At the
onset of the process, cells in the larger “precursor” tubes dergo a dramatic junctional remodeling process, duringwhich their intercellular AJs are successively exchangedappear paired [19], and we show here by using D�-Cat-
GFP as a marker that the AJs of adjacent cells align and with autocellular AJs by a mechanism that looks like a“zipping” process. During this process, the retractingform a ring-like structure perpendicular to the tube axis.
This alignment appears to be important to individual neighboring cell might degrade or redistribute its junc-tional complexes. Alternatively, the interaction of apicalbranch cells’ ability to “slip” between each other and
reach around the lumen with their proximal or distal end. junctional complexes between adjacent cells might beextremely dynamic and allow cells to slide along eachIf cells were not aligned in such a manner, the reaching
around the lumen of one cell end would result in a dis- other by junctional remodeling without removal of the AJcomplexes inserted into the apical plasma membrane. Aplacement of the apical side of the neighboring cell away
from the lumen and eventually in the exclusion of the recent study suggests that the activity level of Rac iscrucial for tracheal cell rearrangement by controlling thecell body from the tube; we have never observed such a
phenomenon. However, we have occasionally observed level of E-cadherin [32], but at present, very little isknown about the dynamics of AJ during cell rearrange-that three neighboring cells make up the luminal circum-
ference in the transverse connective or ganglionic ments and cell intercalation at the molecular level, andfurther analyses are required to gain insight into possiblebranches. Such an arrangement does allow intercalation
as long as only one cell reaches around the lumen at a mechanisms. Live-imaging studies such as those de-scribed in this study are clearly indispensable in thegiven time.
The most peculiar behavior of tube cells during the quest for a better understanding.intercalation process is the initial “reaching around thelumen” of individual cells. We do not know whether this
The Formation of Tubes of Different Cellularis an active cellular process, or whether cells give in, byArchitecture is Controlled by Signal-Regulatedintercalating, to the force produced at the tip of theTranscription Factorsoutgrowing branches via cell movement under the con-Many genes have been implicated in tracheal develop-trol of Bnl/FGF. If the process were an active one, thement in previous studies [33–35], and we reanalyzed theforces might be produced in the cell that reaches aroundmost promising mutants for defects in cell rearrange-the lumen. Alternatively, the neighboring cell could gen-ments with D�-Cat-GFP as a marker. Previous studieserate the force by retracting its distal or proximal endimplicated the Dpp and Wnt signaling pathways and theand thereby pulling the adjacent cell along and aroundKni/Knrl and Sal proteins, respectively, in the formationthe lumen. Importantly, paired cells have to coordinateof distinct branches of different cellular complexitiestheir behavior, because at the onset of the process, anand luminal diameter. Sal is required for the formationindividual cell is allowed to reach around the lumenof the large and multicellular dorsal trunk, whereas Kni/either at its proximal or at its distal end; if it simultane-Knrl are expressed in the much finer autocellular AJ-ously reached around the lumen at both ends (a phe-containing dorsal and ventral branches. With gain-nomenon we never observed with single labeled cells;of-function and loss-of-function genetic analyses, wesee Results), the neighboring cell in the paired alignmentfind that Dpp signaling is not strictly required for cellswould eventually lose its luminal contact and would beto intercalate and form autocellular AJs but that it exertsexcluded from the tube.its function by repressing sal transcription in dorsalTwo recent studies have analyzed how epithelial cellsbranch cells. Sal itself is both required and sufficient tointercalate during germ band elongation in the Drosoph-inhibit cell intercalation and autocellular AJ formation,ila embryo [27, 28]. In this particular system, the remod-and therefore its presence leads to the formation ofeling of junctions is not driven by external forces atmulticellular tubes within the tracheal epithelium. Ourtissue boundaries, but depends on local forces at cellresults also suggest that the ability to intercalate is theboundaries. Myosin II is specifically enriched in disas-“ground state” of tracheal cells; it occurs in the absencesembling junctions, and its polarized localization andof Dpp and Wnt signals.activity are required for junction remodeling and cell
To study the cellular function of the Sal transcriptionintercalation [27]. It will be interesting to see whetherfactor, we ectopically expressed Sal in single trachealmyosin II localization and function also contribute tocells of branches that normally undergo cell intercalationbranching morphogenesis of epithelial organs.and asked whether Sal would inhibit intercalation cellSeveral models have been proposed explaining howautonomously, as visualized by an inhibition of autocel-epithelial cells rearrange without disruption of the AJ
complexes [29]. It has been proposed that such rear- lular AJ formation. Surprisingly, we find that single cells

Current Biology2206
mRFP1 [42] was amplified by PCR and cloned upstream of theexpressing Sal do intercalate, but that in small groupsactin binding domain of moesin [43]. This construct was cloned intoof Sal-expressing cells, the process is inhibited. ThispP{CaSper} carrying the P[B123] fragment to generate btlenhancer-might indicate, for example, that the force-producingmRFP1-moe.
cell is the neighboring cell, the cell that does not ectopi-cally express Sal and is therefore not altered. Alterna- Immunostainings and Whole-Mount In Situ Hybridizationtively, Sal expression might elicit a “community effect,” The following primary antibodies were used: a rabbit anti-Sal anti-keeping cells together in a more static fashion compared body [44], a tracheal lumen-specific mouse monoclonal antibody
(IgM 2A12, used at 1:5 dilution; kindly provided by N. Patel), andto that of Sal-nonexpressing cells (e.g., by the expres-mouse anti-GFP IgG antibody (diluted 1:500; Roche). Embryos weresion of homophilic adhesion molecules) so that theyfixed and immunostained as described before [45] with minor modifi-cannot give in to the external force produced by thecations. For confocal analysis, the signal was amplified. As second-
migration of the cells at the tip of the tracheal branches. ary antibodies, we used peroxidase-conjugated antibodies followedThe identification of Sal target genes will be instrumental by biotinylated tyramide (NEN Life Science Product), which is recog-in gaining further insight into the control of branching nized by streptavidin-fluorescein or streptavidin-Texas Red. IgG an-
tibody was used before the 2A12 IgM antibody to minimize cross-morphogenesis and tube diversity in the Drosophila tra-reaction.cheal system.
Time-Lapse Confocal MicroscopyConclusions Embryos expressing the GFP construct of interest were collectedOur study’s view of branching morphogenesis inte- overnight, dechorionated for 3 min with 3%–4% chlorax, andgrates different signaling systems and their effectors at mounted in 10s Voltalef oil with the hanging drop method. Images
were collected on a Leica TCS SP2 confocal system with the Leicathe cellular level (Figure 6E). On the one hand, Bnl/FGFConfocal Software (Version 2.5). For excitation, the 488 nm emissionsignaling is crucial for cell migration and orients theline of an Argon laser and the 568 nm emission line of a Kryptondirection of branch outgrowth [9], presumably via cy-laser were used. The spacing of the focal sections was chosen in
toskeletal effector molecules [12, 13]. On the other hand, order to comply with the critical sampling distance of the objective.Dpp and Wnt signaling control branch outgrowth via thecontrol of cell intercalation by regulating the expression Deconvolution and Three- and Four-Dimensional Reconstructionsof sal. Only the combined action of these signaling sys- Images were deconvoluted with Huygens Essential (Version 2.3.0)
from SVI and subsequently processed with the Volocity 2.61 soft-tems allows efficient branch elongation and the forma-ware (Improvision) or the Imaris 4.0.4 software (Bitplane). Sometion of morphologically distinct branches.images were treated with Photoshop (Adobe) to enhance the con-trast.Experimental Procedures
Supplemental DataDrosophila Strains and GeneticsSeveral interactive 3D models are available at http://www.current-Targeted gene expression was achieved with the Gal4/UAS systembiology.com/cgi/content/full/14/24/2197/DC1/.[36]. A btl-Gal4 strain, which drives Gal4 expression in all tracheal
cells to high levels from late stage 10 onward, was kindly providedAcknowledgmentsby Shigeo Hayashi [37]; tracheal cells were vizualized with UAS
GFP-actin [38]; AJs were visualized with the UAS-D�-cat-GFP linesWe thank N. Patel for generously supplying 2A12 antibody; S. Hay-kindly provided by Hiroki Oda [22]. A recombinant line with btl-Gal4ashi, R. Schuh, H. Oda, T. Tabata, V. Verkhusha, T. Haerry, and M.was generated with the UAS-D�-cat-GFP#8 line. Tracheal nucleiLlimargas for flies; R.Y. Tsien for the mRFP1 cDNA; M. Durrenbergerwere visualized with the UAS-GFPN-lacZ (2-1)/CyO;btl-Gal4 (3-1)/for help with confocal microscopy; and G. Pyrowolakis for criticallyTM3,Sb,Ser line [37]. sal overexpression was achieved with differentreading the manuscript. This study was supported by the SwissUAS-sal lines provided by Reinhard Schuh [16]. Sal knockdown wasNational Science Foundation and by the cantons Basel-Stadt andachieved with UAS kni [18] or UAS tkvQD [25]. Df(5) [39] was usedBasel-Land.for sal/salr loss of function experiments. Dpp signaling was blocked
cell autonomously with a homozygous UAS-Dad line on the secondchromosome [40]. Received: October 1, 2004
hs flp;btlenhancer�y��Gal4,UAS-D�-cat-GFP;btlenhancer- Revised: October 1, 2004mRFP1-moe or hs flp;btlenhancer�y��Gal4,UAS GFP-actin;btlen- Accepted: October 19, 2004hancer-mRFP1-moe (see below) embryos were collected overnight Published: December 29, 2004at 18�C, heat shocked for 30 min at 37�C, and raised at 25�C for5–12 hr. For single cell overexpression, these lines were crossed to ReferencesUAS-sal or UAS tkvQD, and the effect was monitored at least 9 hrafter the heatshock. 1. Perez-Moreno, M., Jamora, C., and Fuchs, E. (2003). Sticky
Embryos lacking Dpp and Wnt signaling in the trachea were gener- business: Orchestrating cellular signals at adherens junctions.ated by first crossing armXM19 (an allele affecting only the signaling Cell 112, 535–548.functions of Arm) virgins to btl-Gal4,UAS-D�-cat-GFP#8 or UAS- 2. Hogan, B.L., and Kolodziej, P.A. (2002). Organogenesis: Molecu-GFPN-lacZ (2-1)/CyO;btl-Gal4 (3-1)/TM3,Sb,Ser males. Virgins car- lar mechanisms of tubulogenesis. Nat. Rev. Genet. 3, 513–523.rying the armXM19 allele and the reporter constructs were then 3. Lubarsky, B., and Krasnow, M.A. (2003). Tube morphogenesis:crossed to males of a homozygous UAS-Dad line. Embryos lacking Making and shaping biological tubes. Cell 112, 19–28.Dpp and Wnt signaling in the trachea were recognized by the expres- 4. Affolter, M., Bellusci, S., Itoh, N., Shilo, B., Thiery, J.P., andsion of the reporter constructs and the lack of dorsal trunk formation. Werb, Z. (2003). Tube or not tube: Remodeling epithelial tissues
by branching morphogenesis. Dev. Cell 4, 11–18.5. Manning, G., and Krasnow, M.A. (1993). Development of theCloning
The P[B123] fragment upstream of the btl gene [41] was amplified by Drosophila tracheal system. In The Development of Drosophilamelanogaster, M. Bate and A. Martınez-Arias, eds. (Cold SpringPCR and subcloned upstream of the Gal4 sequence into pP{Car20.1}
whose ry� gene was deleted. A flip-out cassette containing the y� Harbor, NY: Cold Spring Harbor Laboratory Press), pp. 609–685.6. Samakovlis, C., Manning, G., Steneberg, P., Hacohen, N., Can-gene [23] was then inserted between the enhancer and the Gal4
CDS to generate btlenhancer�y��Gal4. tera, R., and Krasnow, M.A. (1996). Genetic control of epithelial

Cell Intercalation in Tracheal Morphogenesis2207
tube fusion during Drosophila tracheal development. Develop- junction remodeling controls planar cell intercalation and axiselongation. Nature 429, 667–671.ment 122, 3531–3536.
7. Tanaka-Matakatsu, M., Uemura, T., Oda, H., Takeichi, M., and 28. Zallen, J.A., and Wieschaus, E. (2004). Patterned gene expres-sion directs bipolar planar polarity in Drosophila. Dev. Cell 6,Hayashi, S. (1996). Cadherin-mediated cell adhesion and cell
motility in Drosophila trachea regulated by the transcription 343–355.29. Fristrom, D. (1988). The cellular basis of epithelial morphogene-factor Escargot. Development 122, 3697–3705.
8. Lee, S., and Kolodziej, P.A. (2002). The plakin Short Stop and sis. A review. Tissue Cell 20, 645–690.30. Williams-Masson, E.M., Heid, P.J., Lavin, C.A., and Hardin, J.the RhoA GTPase are required for E-cadherin-dependent apical
surface remodeling during tracheal tube fusion. Development (1998). The cellular mechanism of epithelial rearrangement dur-ing morphogenesis of the Caenorhabditis elegans dorsal hypo-129, 1509–1520.
9. Sutherland, D., Samakovlis, C., and Krasnow, M.A. (1996). dermis. Dev. Biol. 204, 263–276.31. Jacobson, A.G., Oster, G.F., Odell, G.M., and Cheng, L.Y. (1986).branchless encodes a Drosophila FGF homolog that controls
tracheal cell migration and the pattern of branching. Cell 87, Neurulation and the cortical tractor model for epithelial folding.J. Embryol. Exp. Morphol. 96, 19–49.1091–1101.
10. Klambt, C., Glazer, L., and Shilo, B.Z. (1992). breathless, a Dro- 32. Chihara, T., Kato, K., Taniguchi, M., Ng, J., and Hayashi, S.(2003). Rac promotes epithelial cell rearrangement during tra-sophila FGF receptor homolog, is essential for migration of tra-
cheal and specific midline glial cells. Genes Dev. 6, 1668–1678. cheal tubulogenesis in Drosophila. Development 130, 1419–1428.11. Reichman-Fried, M., Dickson, B., Hafen, E., and Shilo, B.Z.
(1994). Elucidation of the role of breathless, a Drosophila FGF 33. Affolter, M., and Shilo, B.Z. (2000). Genetic control of branchingmorphogenesis during Drosophila tracheal development. Curr.receptor homolog, in tracheal cell migration. Genes Dev. 8,
428–439. Opin. Cell Biol. 12, 731–735.34. Ghabrial, A., Luschnig, S., Metzstein, M.M., and Krasnow, M.A.12. Ribeiro, C., Ebner, A., and Affolter, M. (2002). In vivo imaging
reveals different cellular functions for FGF and Dpp signaling (2003). Branching morphogenesis of the Drosophila trachealsystem. Annu. Rev. Cell Dev. Biol. 19, 623–647.in tracheal branching morphogenesis. Dev. Cell 2, 677–683.
13. Sato, M., and Kornberg, T.B. (2002). FGF is an essential mitogen 35. Uv, A., Cantera, R., and Samakovlis, C. (2003). Drosophila tra-cheal morphogenesis: Intricate cellular solutions to basicand chemoattractant for the air sacs of the drosophila tracheal
system. Dev. Cell 3, 195–207. plumbing problems. Trends Cell Biol. 13, 301–309.36. Brand, A.H., and Perrimon, N. (1993). Targeted gene expression14. Llimargas, M. (2000). Wingless and its signalling pathway have
common and separable functions during tracheal development. as a means of altering cell fates and generating dominant phe-notypes. Development 118, 401–415.Development 127, 4407–4417.
15. Chihara, T., and Hayashi, S. (2000). Control of tracheal tubulo- 37. Shiga, Y., Tanaka-Matakatsu, M., and Hayashi, S. (1996). A nu-clear GFP �-galactosidase fusion protein as a marker for mor-genesis by Wingless signaling. Development 127, 4433–4442.
16. Kuhnlein, R.P., and Schuh, R. (1996). Dual function of the region- phogenesis in living Drosophila. Dev. Growth Differ. 38, 99–106.38. Verkhusha, V.V., Tsukita, S., and Oda, H. (1999). Actin dynamicsspecific homeotic gene spalt during Drosophila tracheal system
development. Development 122, 2215–2223. in lamellipodia of migrating border cells in the Drosophila ovaryrevealed by a GFP-actin fusion protein. FEBS Lett. 445, 395–401.17. Vincent, S., Ruberte, E., Grieder, N.C., Chen, C.K., Haerry, T.,
Schuh, R., and Affolter, M. (1997). DPP controls tracheal cell 39. Barrio, R., de Celis, J.F., Bolshakov, S., and Kafatos, F.C. (1999).Identification of regulatory regions driving the expression of themigration along the dorsoventral body axis of the Drosophila
embryo. Development 124, 2741–2750. Drosophila spalt complex at different developmental stages.Dev. Biol. 215, 33–47.18. Chen, C.K., Kuhnlein, R.P., Eulenberg, K.G., Vincent, S., Affolter,
M., and Schuh, R. (1998). The transcription factors KNIRPS and 40. Tsuneizumi, K., Nakayama, T., Kamoshida, Y., Kornberg, T.B.,Christian, J.L., and Tabata, T. (1997). Daughters against dppKNIRPS RELATED control cell migration and branch morpho-
genesis during Drosophila tracheal development. Development modulates dpp organizing activity in Drosophila wing develop-ment. Nature 389, 627–631.125, 4959–4968.
41. Ohshiro, T., and Saigo, K. (1997). Transcriptional regulation of19. Samakovlis, C., Hacohen, N., Manning, G., Sutherland, D.C.,breathless FGF receptor gene by binding of TRACHEALESS/Guillemin, K., and Krasnow, M.A. (1996). Development of thedARNT heterodimers to three central midline elements in Dro-Drosophila tracheal system occurs by a series of morphologi-sophila developing trachea. Development 124, 3975–3986.cally distinct but genetically coupled branching events. Devel-
42. Campbell, R.E., Tour, O., Palmer, A.E., Steinbach, P.A., Baird,opment 122, 1395–1407.G.S., Zacharias, D.A., and Tsien, R.Y. (2002). A monomeric red20. Beitel, G.J., and Krasnow, M.A. (2000). Genetic control of epithe-fluorescent protein. Proc. Natl. Acad. Sci. USA 99, 7877–7882.lial tube size in the Drosophila tracheal system. Development
43. Ohashi, T., Kiehart, D.P., and Erickson, H.P. (2002). Dual labeling127, 3271–3282.of the fibronectin matrix and actin cytoskeleton with green fluo-21. Jazwinska, A., Ribeiro, C., and Affolter, M. (2003). Epithelialrescent protein variants. J. Cell Sci. 115, 1221–1229.tube morphogenesis during Drosophila tracheal development
44. Kuhnlein, R.P., Frommer, G., Friedrich, M., Gonzalez-Gaitan, M.,requires Piopio, a luminal ZP protein. Nat. Cell Biol. 5, 895–901.Weber, A., Wagner-Bernholz, J.F., Gehring, W.J., Jackle, H., and22. Oda, H., and Tsukita, S. (1999). Dynamic features of adherensSchuh, R. (1994). spalt encodes an evolutionarily conservedjunctions during Drosophila embryonic epithelial morphogene-zinc finger protein of novel structure which provides homeoticsis revealed by a Dalpha-catenin-GFP fusion protein. Dev.gene function in the head and tail region of the DrosophilaGenes Evol. 209, 218–225.embryo. EMBO J. 13, 168–179.23. Basler, K., and Struhl, G. (1994). Compartment boundaries and
45. Patel, N.H. (1994). Imaging neuronal subsets and other cell typesthe control of Drosophila limb pattern by hedgehog protein.in whole-mount Drosophila embryos and larvae using antibodyNature 368, 208–214.probes. Methods Cell Biol. 44, 445–487.24. Franch-Marro, X., and Casanova, J. (2002). spalt-induced speci-
fication of distinct dorsal and ventral domains is required forDrosophila tracheal patterning. Dev. Biol. 250, 374–382.
25. Haerry, T.E., Khalsa, O., O’Connor, M.B., and Wharton, K.A.(1998). Synergistic signaling by two BMP ligands through theSAX and TKV receptors controls wing growth and patterning inDrosophila. Development 125, 3977–3987.
26. Steneberg, P., Hemphala, J., and Samakovlis, C. (1999). Dppand Notch specify the fusion cell fate in the dorsal branches ofthe Drosophila trachea. Mech. Dev. 87, 153–163.
27. Bertet, C., Sulak, L., and Lecuit, T. (2004). Myosin-dependent
Copyright © 2022 FDOKUMEN




















