
Influence of the photoperiod on TGF-β1 and p15 expression in hamster Leydig cells

THE ANATOMICAL RECORD 213:448-456 (1985)
Development of Hamster Tracheal Epithelium: I I . Cell Proliferation in the Fetus
ELIZABETH M. McDOWELL, CARNELL NEWKIRK, AND BILL COLEMAN Department of Pathology, University of Maryland School of Medicine (E. M. M., C.N.) and Maryland Znstitute for Emergency Medical Services Systems (B. C.), Baltimore, MD 21201
ABSTRACT Proliferation of epithelial cells in the fetal trachea was studied in hamsters, beginning on the 10th gestational day and ending on the 16th day, shortly after birth. The mean mitotic index (MI) was highest on day 10, with no statistical confirmation of a change between days 10 and 11. The MI fell to about 2% on days 12 and 13, and declined thereafter to about 0.3% on day 16. The MIS for dorsal and ventral surfaces were compared and values were similar except on day 10, when ventral exceeded dorsal, and on day 12, when dorsal exceeded ventral, 2.56% and 1.3%, respectively. On days 10, 11, and 12 the epithelium was simple, composed of poorly differentiated columnar cells that proliferated. On day 13 the epithelium was pseudostratified owing to the presence of a few short cells that did not reach to the lumen. Throughout the fetal period, proliferation of columnar cells predominated but division of short (basal) cells increased from 8% to 40% of the total mitotic activity between day 13 and day 15. Proliferation of basal cells then declined, so that on day 16, 84% of all cells in mitosis were columnar. If basal cells divide to make more of themselves they must proliferate rapidly between day 13 and day 15, because they were virtually absent on day 12 but accounted for about 36% of the ventral and 23% of the dorsal epithelial cells on day 16. Based on the results, a hypothetical model is proposed for the formation of pseudostratified mucociliary epithelium. The model proposes that small granule (endocrine) cells, basal cells, secretory cells, and ciliated cells are established from poorly differentiated columnar cells by day 13. Thereafter short basal cells replicate to produce only basal cells, whereas columnar secretory cells replicate to produce secretory cells and ciliated cells. This model will be tested in future studies.
Studies on various aspects of tracheal development in the fetus have been made in several mammalian species (Leeson, 1961; Dirksen and Crocker, 1965; Cireli, 1966; Kanda and Hilding, 1968; Jeffery and Reid, 1977; Ka- wamata and Fujita 1983) including Syrian golden ham- sters (Emura and Mohr, 1975; Emma et al., 1976, 1977; Mohr et al., 1979; Reznik-Schuller, 1982), but none of these studies has defined the mitotic indices of the dif- ferent epithelial cell types in the ventral and dorsal surfaces during the developmental period.
Prevailing dogma asserts that basal cells are respon- sible for development and maintenance of the tracheo- bronchial epithelium and for restoration of the muco- ciliary state following injury. It is a commonly held the- sis that basal cells of the respiratory epithelium are an- alogous to those of the skin, representing an “undiffer- entiated” or “stem” cell population. Accordingly, trach- eobronchial basal cells are often referred to as “reserve cells,” the division of which gives rise to other cell types in normal development, in maintenance of the adult epithelium, and in regeneration following injury, and to neoplastic cells in tumorigenesis. Evidence against this analogy is mounting. It has been demonstrated that the basal cells of adult human bronchi contain intermediate and high-molecular-weight keratins (Said et al., 1983)
found in the suprabasal differentiating cells but not found in the basal cells of the epidermis (Banks-Schlegel et al., 1981; Thomas et al., 1984). Moreover, several recent reports have described considerable mitotic po- tential in the columnar secretory cells of the tracheo- bronchial epithelium of rodents and primates in normal and abnormal states (Boren and Paradise, 1978; Jeffery et al., 1982; Keenan et al., 1982a,b,c; McDowell et al., 1984a,b; Wilson et al., 1984).
Willis (1962) stressed that regenerating epithelia of mature adult tissues display properties and structures that are essentially similar to those of the corresponding immature fetal tissues. This is certainly true of the trachea, since regeneration of the adult epithelium fol- lowing injury shares many similarities with the normal developmental sequence, as far as it is known (reviewed by McDowell et al., 1984b). Studies in our laboratory have shown that columnar secretory cells play a pivotal role in all stages of regeneration of the adult mucociliary
Received December 10, 1984; accepted May 31, 1985. This is contribution #1763 from the Cellular Pathobiology Labora-
tory of the Department of Pathology, University of Maryland School of Medicine.
0 1985 ALAN R. LISS, INC.

DEVELOPMENT OF HAMSTER TRACHEAL EPITHELIUM 449
tracheal epithelium following mechanical and nutri- tional (vitamin A deficiency) injury. When the epithe- lium is injured, secretory cells divide to form a keratinizing metaplastic epithelium. When the injury is removed, presecretory and preciliated cells arise from division of secretory cells, and shortly thereafter the mucociliary epithelium is restored (Keenan et al., 1982a,b,c, 1983; McDowell et al., 1984a,b).
Although it has been assumed that basal cells act as “reserve cells” from which other cell types arise during development of the tracheobronchial epithelium (Cireli, 1966; Emura et al., 1976), definitive evidence is lacking. In this paper we describe the mitotic indices of the tracheal epithelium during its development in the ham- ster fetus, to learn more about the primordial stem cell(s) and to provide estimates of the relative contributions made by the more specialized cells later in the develop- mental sequence as the mucociliary epithelium is estab- lished. The morphology of the developing tracheal epithelium in fetal hamsters is described in a companion paper (McDowell et al., 1985).
MATERIALS AND METHODS Subjects
The subjects were the same immature 45 hamsters (39 fetal and 6 on the day of birth) that were described in a companion paper (see Materials and Methods, Quanti- tative Morphological Studies, McDowell et al., 1985).
Quantitative Studies Specimen preparation, the number of tracheal sections
examined per fetus, the criteria for cell identification, the landmarks used to categorize the epithelium as dor- sal, ventral and as cranial and caudal, and the total number of cells counted per fetus on each day of devel- opment from gestational day 10 to day 16 were described in a companion paper (McDowell et al., 1985).
Quantitation of cells in mitosis Every cell counted and scored had a visible nucleus
and the number of cells with mitotic nuclei (prophase to telophase) was recorded. The mitotic nuclei were scored according to their geographic location in the epithelium. Mitotic figures were counted in 2-pm-thick glycol meth- acrylate sections under an oil-immersion objective lens by one investigator (E.McD.). They were scored as low if the mitotic figure was located close to the basement membrane and if it was surrounded by cytoplasm that contacted the basement membrane but did not reach to the lumen. Mitotic figures were scored as middle if they were located near the middle or towards the surface of the epithelium and were surrounded 1) by cytoplasm that reached neither to the basement membrane nor to the surface, or 2) by cytoplasm that was seen to reach to the basement membrane but not to the lumen, in that particular section. Mitotic figures were scored as high if they were in epithelial locations similar to those scored as middle but were surrounded by cytoplasm that reach- ed t o the lumen, and in some sections reached also to the basement membrane.
Serial sectioning On each of gestational days 11, 12, and 13,2-pm-thick
serial sections from one tracheal specimen were stained with hematoxylin and eosin and were photographed from
larynx to carina, so as to further characterize cells in which the nuclei had been scored as low, middle, or high. The tracheal sections were prepared sagitally along midline from larynx to carina. Because the tra- chea was rapidly lengthening, this required 3 photo- graphic exposures at day 11,6 exposures at day 12, and 12 exposures at day 13, at an original magnification of 100, for each 2-pm-thick section of trachea. The nega- tives were enlarged by a factor of 3 photographically and montages of the tracheal sections were mounted on boards. Areas of special interest were enlarged further photographically.
Statistical studies We used a linear logistic model to study the data given
in Table 2 and plotted in Figure 2, and the data given in Table 5 . This model is explained in detail in the succeeding companion paper (Coleman and McDowell, 1985).
RESULTS Total Mitotic Indices
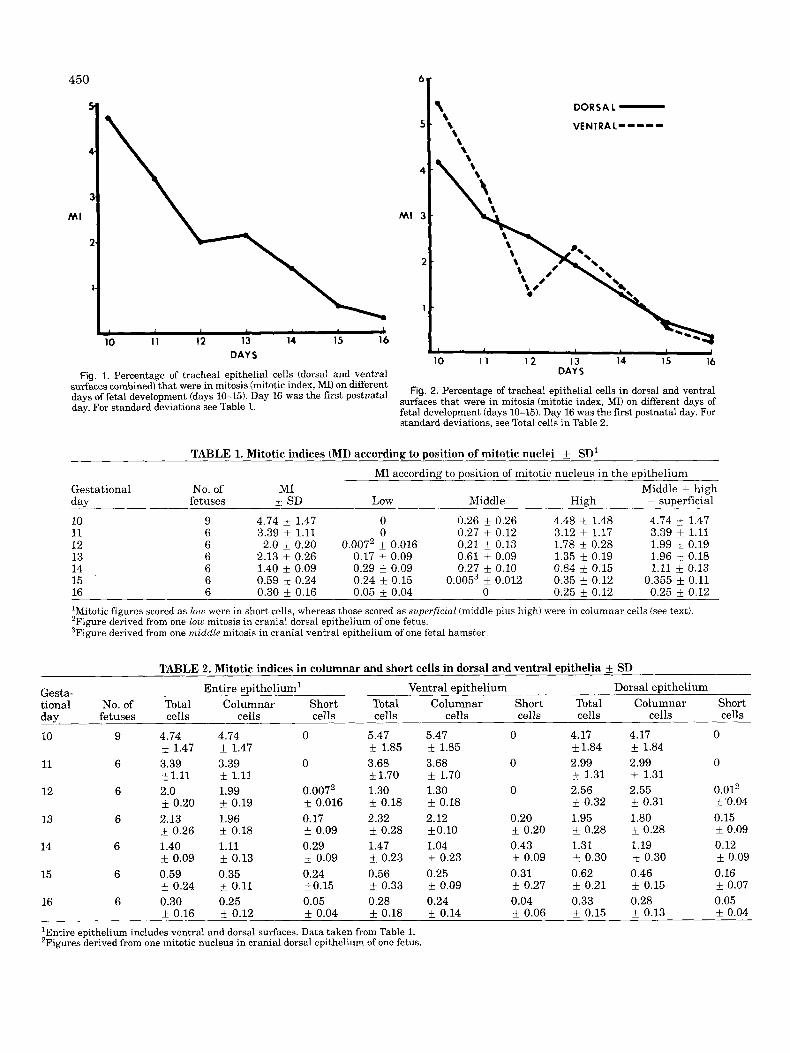
The percentage of cells in the tracheal epithelium (dor- sal and ventral surfaces combined) in mitosis (mitotic index, MI), on different days of development is shown in Figure 1 and Table 1. The mean MI was highest on fetal day 10, about 4.7%, with no statistical confirmation of a change between days 10 and 11. The MI fell to about 2% on days 12 and 13, and thereafter declined to about 0.3% on day 16, the day of birth. On days 12-16, MIS of the cranial and caudal parts of the trachea were compared and the values appeared to be very similar at all times. The MIS for dorsal and ventral epithelial surfaces are shown separately in Figure 2 and Table 2. The values for dorsal and ventral epithelia were similar a t all times except on day 10 (ventral exceeded dorsal) and on day 12, when the dorsal MI exceeded the ventral MI (2.56% and 1.3%, respectively). A detailed statistical analysis is given in the companion paper (Coleman and McDowell, 1985: Tables 2-4).
Mitotic indices according to the position of mitotic nuclei within the epithelium are also shown in Table 1. No mitotic figures were low in the epithelium on days 10 or 11. The onset of pseudostratification was heralded on day 12 by the appearance of one mitotic figure low in the epithelium from a total of 12,096 cells counted, of which 242 were in mitosis. Mitotic nuclei low in the epithelium were invariably surrounded by cytoplasm that contacted the basement membrane but did not reach the lumen. The observation that the cells were short was confirmed in serial sections (Fig. 3A-D). Mitotic nuclei located in the middle of the epithelium were relatively common through day 14. They peaked at day 13 (Table l), a time when the ventral epithelium gave the impression of being stratified in some specimens (McDowell et al., 1985). No mitotic figures were seen in the middle epithelial position on days 15 and 16, with the exception of one mitotic figure in one fetus on day 15. Mitotic figures were scored as middle if they were located about the middle or towards the epithelial sur- face but were surrounded by cytoplasm that did not reach to the surface in that particular section. A study of serial sections confirmed that these mitotic figures also met the criteria for high, in that the cytoplasm
450
MI
6
b 10 1 1 12 13 14 15 16
DAY 5
\ DORSAL - \ VENTRAL----= \
\ \ \
~
10 1 1 12 13 14 IS 16 DAYS Fig. 1. Percentage of tracheal epithelial cells (dorsal and ventral
surfaces combined) that were in mitosis (mitotic index, MI) on different days of fetal development (days 10-15). Day 16 was the first postnatal day. For standard deviations see Table 1.
Fig. 2. Percentage of tracheal epithelial cells in dorsal and ventral surfaces that were in mitosis (mitotic index, MI) on different days of fetal development (days 10-15). Day 16 was the first postnatal day. For standard deviations. see Total cells in Table 2.
TABLE 1. Mitotic indices (MI) according to position of mitotic nuclei f SD' MI according to position of mitotic nucleus in the epithelium
Gestational No. of MI Middle + high day 10 9 4.74 k 1.47 0 0.26 k 0.26 4.48 f 1.48 4.74 k 1.47 11 6 3.39 k 1.11 0 0.27 i 0.12 3.12 f 1.17 3.39 * 1.11 12 6 2.0 f 0.20 0.0072 k 0.016 0.21 & 0.13 1.78 i 0.28 1.99 * 0.19 13 6 2.13 i 0.26 0.17 i 0.09 0.61 k 0.09 1.35 k 0.19 1.96 k 0.18 14 6 1.40 i 0.09 0.29 f 0.09 0.27 k 0.10 0.84 * 0.15 1.11 k 0.13 15 6 0.59 i- 0.24 0.24 f 0.15 0.0053 k 0.012 0.35 +_ 0.12 0.355 i 0.11 16 6 0.30 f 0.16 0.05 f 0.04 0 0.25 f 0.12 0.25 f 0.12 'Mitotic figures scored as low were in short cells, whereas those scored as superficial (middle plus high) were in columnar cells (see text). 'Figure derived from one low mitosis in cranial dorsal epithelium of one fetus. 3Figure derived from one middle mitosis in cranial ventral epithelium of one fetal hamster.
fetuses 3: SD Low Middle High = superficial
TABLE 2. Mitotic indices in columnar and short cells in dorsal and ventral epithelia SD Entire epithelium' Ventral epithelium Dorsal epithelium Gesta-
tional No. of Total Columnar Short Total Columnar Short Total Columnar Short day fetuses cells cells cells cells cells cells cells cells cells
10 9 4.74
11 6 3.39
12 6 2.0
13 6 2.13
14 6 1.40
15 6 0.59
f 1.47
k1.11
* 0.20
0.26
- + 0.09
f 0.24
4.74 f 1.47 3.39 * 1.11 1.99 f 0.19 1.96 f 0.18 1.11 - + 0.13 0.35 * 0.11
0
0
0.0072
0.17 f 0.09 0.29 - + 0.09 0.24 i0.15
0.016
5.47 f 1.85 3.68 f 1.70 1.30 f 0.18 2.32 i 0.28 1.47 i 0.23 0.56 i. 0.33
5.47 k 1.85 3.68 f 1.70 1.30 f 0.18 2.12 fO.10 1.04 f 0.23 0.25 f 0.09
0
0
0
0.20 f 0.20 0.43 f 0.09 0.31 f 0.27
4.17 f1.84 2.99
1.31 2.56 f 0.32 1.95 f 0.28 1.31 f 0.30 0.62 f 0.21
4.17 & 1.84 2.99 f 1.31 2.55 * 0.31 1.80
0.28 1.19
0.30 0.46 f 0.15
0
0
0.012 f 0.04 0.15 i 0.09 0.12 f 0.09 0.16 - + 0.07
16 6 0.30 0.25 0.05 0.28 0.24 0.04 0.33 0.28 0.05 0.16 & 0.12 f 0.04 & 0.18 k 0.14 f 0.06 f 0.15 f 0.13 f 0.04
'Entire epithelium includes ventral and dorsal surfaces. Data taken from Table 1. 'Figures derived from one mitotic nucleus in cranial dorsal epithelium of one fetus.
DEVELOPMENT OF HAMSTER TRACHEAL EPITHELIUM 451
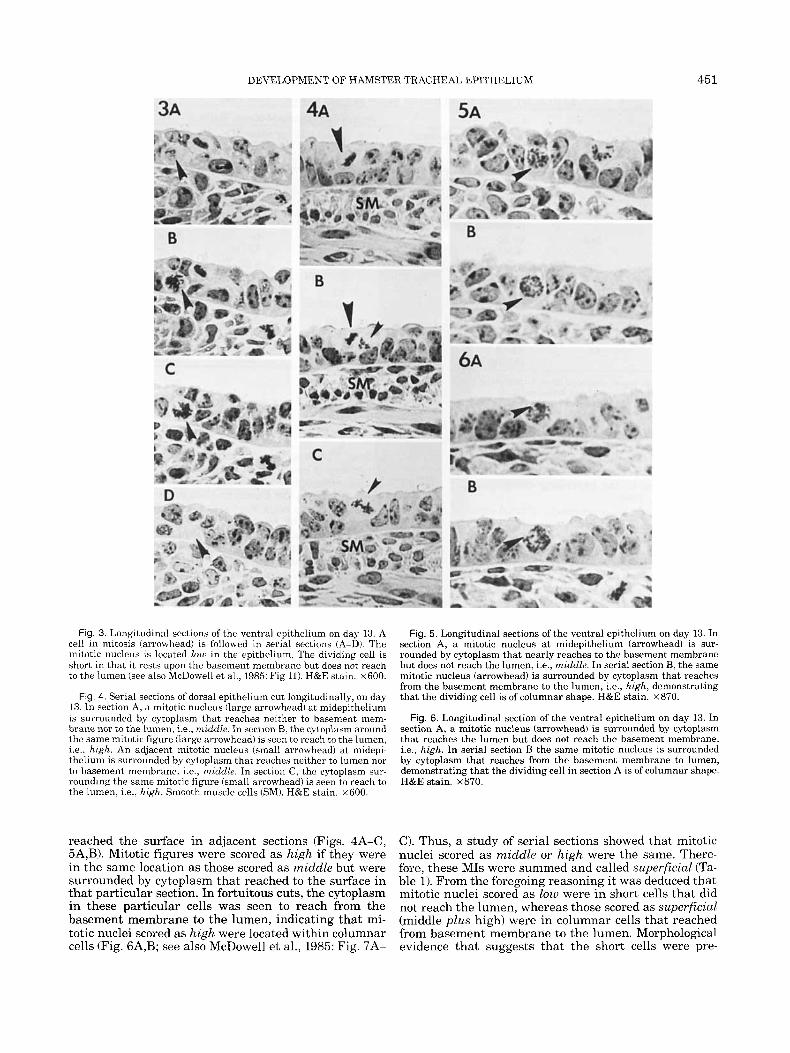
Fig. 3. Longitudinal sections of the ventral epithelium on day 13. A cell in mitosis (arrowhead) is followed in serial sections (A-D). The mitotic nucleus is located low in the epithelium. The dividing cell is short in that it rests upon the basement membrane but does not reach to the lumen (see also McDowell et al., 1985: Fig 11). H&E stain. ~ 6 0 0 .
Fig. 4. Serial sections of dorsal epithelium cut longitudinally, on day 13. In section A, a mitotic nucleus (large arrowhead) at midepithelium is surrounded by cytoplasm that reaches neither to basement mem- brane nor to the lumen, i.e., middle. In sectionB, the cytoplasm around the same mitotic figure (large arrowhead) is seen to reach to the lumen, i.e., high. An adjacent mitotic nucleus (small arrowhead) at midepi- thelium is surrounded by cytoplasm that reaches neither to lumen nor to basement membrane, i.e., middle. In section C, the cytoplasm sur- rounding the same mitotic figure (small arrowhead) is seen to reach to the lumen, i.e., high. Smooth muscle cells (SM). H&E stain. x600.
reached the surface in adjacent sections (Figs. 4A-C, 5A,B). Mitotic figures were scored as high if they were in the same location as those scored as middle but were surrounded by cytoplasm that reached to the surface in that particular section. In fortuitous cuts, the cytoplasm in these particular cells was seen to reach from the basement membrane to the lumen, indicating that mi- totic nuclei scored as high were located within columnar cells (Fig. 6A,B; see also McDowell et al., 1985: Fig. 7A-
Fig. 5. Longitudinal sections of the ventral epithelium on day 13. In section A, a mitotic nucleus at midepithelium (arrowhead) is sur- rounded by cytoplasm that nearly reaches to the basement membrane but does not reach the lumen, i.e., middle. In serial section B, the same mitotic nucleus (arrowhead) is surrounded by cytoplasm that reaches from the basement membrane to the lumen, i.e., high, demonstrating that the dividing cell is of columnar shape. H&E stain. ~ 8 7 0 .
Fig. 6. Longitudinal section of the ventral epithelium on day 13. In section A, a mitotic nucleus (arrowhead) is surrounded by cytoplasm that reaches the lumen but does not reach the basement membrane, i.e., high. In serial section B the same mitotic nucleus is surrounded by cytoplasm that reaches from the basement membrane to lumen, demonstrating that the dividing cell in section A is of columnar shape. H&E stain. ~ 8 7 0 .
C). Thus, a study of serial sections showed that mitotic nuclei scored as middle or high were the same. There- fore, these MIS were summed and called superficial (Ta- ble 1). From the foregoing reasoning it was deduced that mitotic nuclei scored as low were in short cells that did not reach the lumen, whereas those scored as superficial (middle plus high) were in columnar cells that reached from basement membrane to the lumen. Morphological evidence that suggests that the short cells were pre-

452 E.M. McDOWELL, C. NEWKIRK, AND B. COLEMAN
TABLE 3. Proportions of mitotic nuclei in short and columnar cells Proportion of mitotic count, % (No.)
Gesta- - Entire epithelium' Ventral epithelium Dorsal epithelium tional No. of Mitotic Columnar Short Columnar Short Columnar Short day fetuses count1 cells cells cells cells cells cells
10 9 249 100 0 100 0 100 0 (249/249) (01249) (132/132) (OD321 (117/117) (0/117)
11 6 249 100 0 100 0 100 0 (249/249) (01249) (121421) (0/121) (128/128) (01128)
12 6 242 99.59 0.413 100 0 99.42 0.583 (241/242) (1/242) (71/71) (0/71) (1701171) (1/171)
13 6 358 91.62 8.38 91.01 8.99 92.31 7.69 (3281358) (30/358) (172/189) (17/189) (156/169) (13/169)
14 6 272 79.05 20.95 70.7 29.30 90.44 9.56 (2 15/272) (57/272) (111/157) (46/157) (1041115) (11/115)
15 6 110 60 40 45.29 54.71 73.7 26.3 (66/110) (44/110) (24/53) (29/53) (42/57) (1515 7)
16 6 58 84.48 15.52 84.62 15.38 84.38 15.62 (49/58) (9/58) (22/26) (4/26) (27/32) (5/32)
'Total number of cells in mitosis counted a t each time point in all fetuses. 2Entire epithelium includes ventral and dorsal surfaces. 3Figures derived from one mitotic nucleus in the cranial dorsal epithelium of one fetus.
sumptive basal cells (prebasal cells) and that the colum- nar cells were presumptive secretory cells (presecretory cells) is given in the preceding companion paper (Mc- Dowell et al., 1985).
The MIS and the proportions of total mitotic activity due to columnar and short cells in ventral and dorsal epithelia on different days of fetal development are shown in Tables 2 and 3. On days 10 and 11, the epithe- lium was simple columnar (McDowell et al., 1985) and all proliferative activity was due to division of a poorly differentiated columnar cell type. On day 12, more than 99% of all proliferative activity was associated with simple columnar cells. On day 13 the epithelium became pseudostratified and during days 13 and 15 mitotic divi- sion in short cells increased from 8% to 40% of the total. However, on day 16 only about 16% of all mitotic figures were in short cells (see Table 3, Entire Epithelium). Thus, throughout the developmental period, mitotic ac- tivity in columnar cells predominated over that in short cells, when the entire epithelium (dorsal and ventral surfaces collectively) was considered.
On day 13 the proportions of mitotic figures in short and columnar cells in ventral and dorsal epithelia were similar. About 8-9% of dividing cells were short and about 91-92% were columnar, in both surfaces (Table 3). This was not so on day 14, when about 30% of dividing cells were short and 70% were columnar ventrally, whereas about 10% were short and 90% were columnar dorsally. This trend was increased by day 15, so that about 55% of dividing cells were short and 45% were columnar ventrally, whereas about 26% were short and 74% were columnar dorsally. However, by day 16 propor- tions of mitotic figures in ventral and dorsal epithelia in short and columnar cells were similar, and about 16% of dividing cells were short and about 84% were columnar, in both surfaces (Table 3).
In Table 4, the data from Table 3 are shown according to their distribution in cranial and caudal parts of the
trachea. The proportion of mitotic figures in short cells was generally smaller in the caudal than in the cranial trachea, although this was not so in the ventral trachea on day 15, nor in the dorsal trachea on day 16.
Although some cells were short on days 13 and 14, it was impossible to determine the exact proportions of poorly differentiated short and columnar cells by their light microscope morphologies at these times (McDowell et al., 1985). However, by day 15 it was possible to categorize the cells into four categories: basal, presecre- tory, preciliatedkiliated, and small-granule cells (Mc- Dowell et al., 1985; Table 3). Only basal cells and presecretory cells were seen in mitosis; neither small- granule cells nor preciliated-ciliated cells were seen in division, although small-granule cells appeared to be capable of replicating on day 12 (McDowell et al., 1985). Therefore, it was possible to calculate the MI of the basal cell and presecretory cell populations separately on days 15 and 16 (Table 5) and these are analyzed statistically in the following paper (Coleman and McDowell, 1985: Tables 5-7).
DISCUSSION Growth of the Fetal Hamster Trachea
Longitudinal and circumferential growth of the fetal trachea and its epithelium was a dynamic process that involved the interplay between cell differentiation, ma- turation, and related modulations in cell height and linear density (described in the companion paper Mc- Dowell et al., 19851, and cell proliferation, described herein. These relationships are illustrated for the dorsal tracheal epithelium in Figure 7. The mitotic index de- clined, whereas the rate of increase in tracheal diameter and length remained steady or increased during the fetal period. Epithelial cell proliferation did not keep pace with the overall growth of the tracheal epithelium and was compensated for by a decline in linear density and cell height that was displayed morphologically as a

rn
u 2
-
-TRACHEAL LENGTHmrn
C . - . - ' - s MITOTIC INDEX%
TRACHEAL DIAMETER
4--400um t-s-4
l 5 I l6 10 II 12 13 14
DAYS BIRTH
change from tall slender columnar cells on days 10 and 11 to wider shorter cells thereafter. The relationships between cell height, linear density, and tracheal diame-
s .r ter were rather similar to those described by Cireli (1966) e 3 in developing rat trachea. Elongation of the adult tra- - $ E S E m 1 - n n h chea occurs physiologically with stretching, and this k 2 E o? r- we hl* w.& N % also is associated with a marked reduction in epithelial
nation of mitotic figures scored as middle on fetal days c - 15 and 16 (Table 1) coincides with a declining linear z.2 O E Z 3 0 - density and decreased cell height, further confirming & 2 G 8 o$ 25 s ; = j ohl w e as c that mitotic figures scored as middte were representa- g .%
tive of columnar cells that reached from basement mem- 'doe3 brane to lumen. - Development of the Tracheal Epithelium and Progression
From Simple Columnar to the Pseudostratified d - 2:Frna - + r(
4 " Mitotic indices of the developing fetal tracheal epithe- lium (dorsal and ventral epithelia, collectively) were
E @ - - - 7-4- m - r- W * 03% - 2 TFz 2 g 8 og @?$zs 2s zs
n 5 2 ~ ~ g rms lo> cell height (Gatto and Houck, 1983). The virtual elimi-
3
t-s a- as 00s r-6
- N m S w g q s oe
-
2 2 E m - o , E = - - - rn gj 3 % g;;j qz 7 5 oz: 0 2 "u 42 g!z 2% % 3 2 z Mucociliary State
- . a - - .- E Z , " 0 M c - , C : * W - v1 .i
K
m E; iJ - - h
Y L o w 5 5
cd 3 $ 2 **rnr-, .mad-
'- & 3
. - g s g ~ ~ ~ ? 2 $ 2 + od rie +, m e 2s r '-
h .-
reported previously by Emura and Mohr (1975), and with the exception of days 10 and 11 our present findings agree fairly closely with their results. However, in their study the MI at day 11 exceeded all others, whereas in the present study, the mean MI was highest on day 10 with no statistical confirmation of a change between days 10 and 11. As mentioned in the companion paper (McDowell et al., 1985), sectioning the tracheal rudi- ment was most exacting technically on day 10, as it was very small. Vertical sections through the epithelial cells, perpendicular to the basement membrane, were abso- lutely essential for accurate analyses of MI on days 10- 12 because the dividing nuclei were located superficially in the columnar cells (McDowell et al., 1985). Because of -
c3
o i-, S 2 , X N
-3
2 M
mitotic index (%), epithelial cell height (pm), and linear density (num-
through day 16 of hamster fetal development. Data are derived from Table 2 of the companion paper WcDowell et al., 19851, and from
m a, ber of cells per 100 pm) for the dorsal tracheal epithelium from day 10
*

454 E.M. McDOWELL, C. NEWKIRK, AND B. COLEMAN
TABLE 5. Mitotic indices by cell type' f SD
Gestational No. of Entire epithelium Ventral epithelium Dorsal epithelium
15 6 0.75 0.63 0.77 0.43 0.73 0.85
16 6 0.16 0.43 0.13 0.38 0.22 0.49
lMIb, 7 Mitotic index of basal cells (% of basal cell population in mitosis); MI,,, = mitotic index of presecretory cells (% of presecretory cell population in mitosis).
day fetuses MIbc MI,,, MIbc MI,,, M1bc MI,,,
f 0.48 F 0.20 f 0.66 +_ 0.15 i 0.35 f 0.30
+ 0.15 f 0.21 k 0.17 * 0.22 f 0.17 k 0.26
the minute size of the trachea on days 10 and 11, tangen- tial cuts through the columnar cells are obtained quite readily, unless precautions are taken to avoid them. If tangentially sectioned epithelial cells were included in the mitotic counts, the MIS would be inaccurate: Cuts through cell apices would artifically raise the MI, whereas cuts through the columnar cell body below the nucleus would lower it. Cells were tallest on day 10 (McDowell et al., 1985: Table 2), which would favor cuts below the dividing nucleus. This explanation might ac- count for the low MI on day 10 and the higher MI on day 11, as reported by Emura and Mohr (1975).
When reporting mitotic indices in the tracheal epithe- lium, it is important that the regional location be consid- ered. For instance in 5-week-old hamsters (McDowell et al., 1984a) and in 8-week-old mice (Bjork, 19571, dividing cells were most common dorsally at the pars membran- acea. MIS for dorsal and ventral surfaces separately have not been reported previously in the developing trachea. For the most part MIS were similar for both surfaces, but on day 12 the MI for the ventral surface was considerably less than that of the dorsal surface. This disparity was due to a relative depression of the MI of the ventral surface. By day 13 the ventral MI had increased to equate with that of the dorsal surface (Fig. 2). The significance of this disparity is not known, but it may relate to the development of preciliated cells and a disproportionate growth of the dorsal epithelium at this time. By day 13 the epithelium of the pars membrana- cea contained preciliated cells and it was thrown into folds (McDowell et al., 1985: Fig. 91, probably the result of functional activity being initiated in the underlying smooth muscle cells.
Before day 12 all epithelial cells were columnar. On day 13 about 8% of cells in division were short and this proportion increased through day 15, when 40% of the dividing cells were short. Thereafter the proportion fell and by day 16 only 16% of the dividing cells were short. As discussed in the companion paper (McDowell et al., 1985), the short cells, seen in small numbers on day 13, are likely to be presumptive basal cells destined to ma- ture into basal cells over the next 2 days. Assuming that short prebasal cells divide to make more of their own kind, they must proliferate rapidly during days 13 to 15 because by the day of birth (day 16), basal cells ac- counted for about 36% of the ventral and 23% of the dorsal epithelial cells (McDowell et al., 1985: Table 3). The percentage of cells in division that were short was greater in the ventral than in the dorsal epithelium on days 14 and 15, a finding that is consistent with higher wrowortions of basal cells in the ventral eDithelium on
phenomenon is analagous to the appearance of presump- tive basal cells, followed by their rapid proliferation in the developing epididymis, except that in that organ the proliferative events took place postnatally (Sun and Flickinger, 1982). Statistical analysis of the MIS by cell type (Coleman and McDowell, 1985: Table 7) further demonstrated that basal cells were highly proliferative on day 15 in both dorsal and ventral epithelia (about 0.7%) but the MI had declined markedly by day 16. The decrease in MI of the presecretory cells on day 16 was less. During the first postnatal week the total MI re- mained low and the proportions of basal cells in ventral and dorsal epithelia were maintained in proportions es- sentially similar to those present on fetal days 15 and 16 (Otani and McDowell, unpublished observations). These observations indicate that most basal cells of the neonatal epithelium were formed in utero and that they proliferated after birth a t a rate sufficient to maintain their proportion in the growing epithelium.
Preciliated-ciliated cells were present in the dorsal epithelium on day 13, but they could not be quantified at this time. The preciliated-ciliated cells could be quan- tified by day 14 when their highest proportion, about 20%, was in the caudal dorsal epithelium (McDowell et al., 1985: Table 3). At this time, about 97% of mitotic divisions at this location were in the columnar cells (caudal dorsal epithelium, Table 4). Studies of regener- ating tracheal epithelium have indicated that precil- iated cells arise from mitotic division of secretory cells (Keenan et al., 1982c; McDowell et al., 1984b). If mitosis is also required for the production of preciliated cells in the developing tracheal epithelium, the overwhelming number of divisions in columnar cells in the dorsal epi- thelium on days 13 and 14 favors the premise that pre- ciliated cells arose from division of these presumptive secretory cells.
A Model of Tracheal Epithelial Development Based on the sequential appearance of cells with dif-
ferent morphologies (McDowell et al., 19851, coupled with a knowledge of their mitotic activities, a hypothetical model for the formation of pseudostratified mucociliary tracheal epithelium is proposed (Fig. 8).
Small-granule cells were first seen in the epithelium on day 12. These organoid clusters of cells expanded on day 12, a t least in part by mitotic activity (McDowell et al., 1985: Fig. 8) as described in rabbit bronchi (Sorokin et al., 1982). By day 13 a few short cells were recogniz- able by their mitotic nuclei placed low in the epithelium. Moreover, columnar wreciliated and a verv few ciliated
dais 15 and 16 (McDowell et al., 1985: Tible 3). This cells were seen dorsaily.

DEVELOPMENT OF HAMSTER TRACHEAL EPITHELIUM
Iy\y 10 rxf 11 mY 12 M Y 13 mY 14 CYWAFUX
455
--F
pRpxsII;IATED
-- P-
BASAL-
BASAL-
s lALLcaA"E (-1
Fig. 8. Fetal development of the hamster tracheal epithelium-a working hypothesis. The first specialized cell type to arise on day 12 is the small-granule endocrine cell. On day 13 lineages are established for short basal cells and for columnar secretory and ciliated cells (prebasal, presecretory, preciliated cells). It is postulated that basal cells replicate to produce more of themselves, whereas columnar secre-
tory cells replicate to produce secretory and ciliated cells. According to this hypothetical scheme, division of any secretory cell may produce two secretory cells, or two ciliated cells (not shown in Fig. 8 ) or one of each cell type. Factors that determine the phenotype of the progeny of secretory cells are presently unknown.
In studies of development of the tracheal epithelium in hamsters and other species, the formation of ciliated cells was said to precede secretory cells (Leeson, 1961; Cireli, 1966; Emura and Mohr, 1975; Kawamata and Fujita, 1983; Plopper and Alley, 1983). However, in this study it appeared likely that the lineages of basal, secre- tory, and ciliated cells were established simultaneously on day 13 (Fig. 8). Preciliated and ciliated cells were clearly recognizable by their characteristic morpholo- gies, but the lineages of prebasal and presecretory cells were also recognizable in the form of short and columnar cells. By day 14 some short cells became more easily recognizable as basal cells by their hemidesmosomes, and chains of RER developed in some columnar cells, so that by day 15, cells had overtly matured into basal and presecretory cells. The latter contained secretion gran- ules (McDowell et al., 1985).
Mitotic activity was not observed in small-granule cells after day 12, and it was never observed in precil- iated or ciliated cells. From day 13 onwards cells of two lineages were proliferative, the short (prebasal) and the columnar (presecretory) cells. The model (Fig. 8) pro- motes the premise that short basal cells replicate to produce more of themselves, whereas secretory cells rep- licate to produce secretory cells and ciliated cells. Ob- viously further studies are required to substantiate this model.
Comparisons Between Normal Development and Regeneration Following Injury
The developmental model (Fig. 8) is consistent with our model of regeneration of adult tracheal epithelium following mechanical and nutritional injuries. When adult tracheal epithelium is injured by trauma or by vitamin A deficiency, a keratinizing metaplastic epithe- lium is formed that is composed largely of altered secre- tory cells. Reversal of the metaplastic state to a
mucociliary epithelium depends upon the production of newly formed preciliated and secretory cells that arise from divisions of secretory cells (Keenan et al., 1982a,b,c, 1983; McDowell et al., 1984a,b). In a regenerating epi- thelium the immature preciliated cells usually contain secretory granules that have been carried over from the progenitor secretory cells, whereas in the developing fetal epithelium, secretory granules were not observed in the preciliated cells at the ultrastructural level, al- though a few PAS-positive granules were seen apically in preciliated cells by light microscopy (McDowell et al., 1985). Failure to observe mucous granules ultrastruc- turally in the fetal preciliated cells is not surprising, since secretory granules were very scant or even absent from the immature presecretory cells of the fetal tra- chea. However, acidic mucosubstances were abundant in some secretory cells of the larynx on day 16, and similar granules were present in nearby preciliated cells (McDowell et al., 1985: Fig. 37). Alcianophilic apical granules were described in ciliated cells in hamster tra- chea on the first postnatal day (Emura and Mohr, 1975). This phenomenon is not unique to hamsters, and pre- ciliated cells bearing secretory granules have been de- scribed during development of mucociliary respiratory epithelium in chicks (Kalnins and Porter, 1969), Xenopus laevis (Steinman, 19681, rats (Stockinger and Cireli, 1965; Cireli, 1966), and rhesus monkeys (Plopper, personal communication).
During normal development and regeneration of tra- cheal epithelium following injury, preciliated cells bear apical secretory granules but their morphology shows no evidence of mucosubstance synthesis, i.e., neither the rough endoplasmic reticulum nor the Golgi apparatus is well developed. We are of the opinion that during nor- mal development and regeneration following injury, pre- ciliated cells are progeny of the division of secretory cells and that the secretory granules are carried over from the parent cell. In studies of developing tracheas from

456 E.M. McDOWELL, C. NEWKIRK, AND B. COLEMAN
chick and toad, secretory granules were present in cells where ciliogenesis had just begun, but granules were not seen in cells where ciliogenesis was advanced (Stein- man, 1968; Kalnins and Porter, 1969). This further sug- gests that secetory granules were “inherited” from the parent cell and were secreted as ciliary specialization progressed and the ciliated cells matured.
Boren and Paradise (1978) showed that secretory cells gave rise to ciliated cells during normal turnover of adult hamster trachea. We predict that this phenome- non is common to many species and propose that secre- tory cells play the major role in genesis of ciliated cells during fetal and neonatal development, during normal adult epithelial cell turnover, and during regeneration of the injured tracheobronchial epithelium.
ACKNOWLEDGMENTS
We would like to thank our colleagues, Judy M. Strum, Elaine M. Otani, and Phillip Sannes for reading this manuscript and for their constructive criticism and help- ful comments.
This work was supported by National Institutes of Health Grant HL 24722.
LITERATURE CITED
Banks-Schlegel, S., R. Schlegel, and G. Pinkus (1981) Keratin protein domains within the human epidermis. Exp. Cell Res., 136:465-469.
Bjork, H. (1957) Mitotic rate and colchicine sensitivity of the epithe- lium of the esophagus, trachea, and bronchi of the white mouse. Ann. Med. Exp. Biol. Fenniae, 35:223-232.
Boren, H.G., and L.J. Paradise (1978) Cytokinetics of lung. In: Patho- genesis and Therapy of Lung Cancer. C.C. Harris, ed. Marcel Dek- ker, New York, pp. 369-418.
Cireli, E. (1966) Elektronenmikroskopische Analyse der pra- und post- natalen Differenzierung des Epithels der oberen Luftwege der Ratte. 2. Mikrosk. Anat. Forsch., 74:132-178.
Coleman, B., and E.M. McDowell (1985) Development of hamster tra- cheal epithelium. 111. Illustration of statistical methods for propor- tional data in biology. Anat. Rec., 213:457-463.
Dirksen, E.R., and T.T. Crocker (1965) Centriole replication in differ- entiating ciliated cells of mammalian respiratory epithelium. An electron microscopic study. J. Microsc., 5:629-644.
Emura, M., and U. Mohr (1975) Morphological studies on the develop- ment of tracheal epithelium in the Syrian golden hamster. I. Light microscopy. 2. Versuchstierk d., 17:14-26.
Emura, M., H. Reznik-Schuller, and H. Mohr (1976) Morphological studies on development of tracheal epithelium in the Syrian golden hamster. II. Electron microscopy: Changing aspects of chromatin structure. 2. Versuchstierk d., 18:317-324.
Emura, M., H. Reznik-Schiiller, and U. Mohr (1977) Morphological studies on the development of tracheal epithelium in the Syrian golden hamster. 2. Versuchstierk d., 19:47-51.
Gatto, L.A., and B.M. Houck (1983) Changes in epithelial configuration following elongation of rat trachea. Trans. Am. Microsc. Soc., 102:89-90.
Jeffrey, P.K., and L.M. Reid (1977) Ultrastructure of airway epithelium and submucosal gland during development. In: Lung Biopsy in Health and Disease, Vol. 6. Development of the Lung. W.A. Hod. son, ed. Marcel Dekker, New York, pp. 87-134.
Jeffrey, P.K., M. Ayers, and D. Rogers (1982) The mechanisms and control of bronchial mucous cell hyperplasia. In: Mucus in Health and Disease. II. E.N. Chantler, J.B. Elder, M. Elstein, eds. Plenum, New York, pp. 399-409.
Kalnins, V.I., and K.R. Porter (1969) Centriole replication during cili.
ogenesis in the chick tracheal epithelium. 2.Zellforsch. Mikrosk. Anat., 100:l-30.
Kanda, T., and D. Hilding (1968) Development of respiratory tract cilia in fetal rabbits. Acta Otolaryngol., 65:611-624.
Kawamata, S., and H. Fujita (1983) Fine structural aspects of the development and aging of the tracheal epithelium of mice. Arch. Histol. Jpn., 46:355-372.
Keenan, K.P., J.W. Combs, and E.M. McDowell (1982a) Regeneration of hamster tracheal epithelium after mechanical injury. I. Focal lesions: Quantitative morphologic study of cell proliferation. Vir- chows Arch. [Cell Pathol.], 41:193-214.
Keenan, K.P., J.W. Combs, and E.M. McDowell (1982b) Regeneration of hamster tracheal epithelium after mechanical injury. 11. Multi- focal lesions: Stathmokinetic and autoradiographic studies of cell proliferation. Virchows Arch. [Cell Pathol.], 41:215-229.
Keenan, K.P., J.W. Combs, and E.M. McDowell (1982~) Regeneration of hamster tracheal epithelium after mechanical injury. 111. Large and small lesions: Comparative stathmokinetic and single pulse and continuous thymidine labeling autoradiographic studies. Vir- chows Arch. [Cell Pathol.], 41t231-252.
Keenan, K.P., T.S. Wilson, and E.M. McDowell(1983) Regeneration of hamster tracheal epithelium after mechanical injury. IV. Histo- chemical, immunocytochemical and ultrastructural studies. Vir- chows Arch. [Cell Pathol.], 43:213-240.
Leeson, T.S. (1961) The development of the trachea in the rabbit with particular reference to its fine structure. Anat. Am., 110:214-223.
McDowell, E.M., K.P. Keenan, and M. Huang (1984a) Effects of vita- min A-deprivation on hamster tracheal epithelium. Virchows Arch. [Cell Pathol.], 45:197-219.
McDowell, E.M., K.P. Keenan, and M. Huang (1984b) Restoration of mucociliary tracheal epithelium following deprivation of vitamin A. Virchows Arch. [Cell Pathol.], 45:221-240.
McDowell, E.M., C. Newkirk, and B. Coleman (1985) Development of hamster tracheal epithelium. I. A quantitative morphologic study in the fetus. Anat. Rec., 213:429-447.
Mohr, U., H. Reznik-Schiiller, and M. Emura (1979) Tissue differentia- tion as a prerequisite for transplacental carcinogenesis in the ham- ster respiratory system, with specific respect to the trachea. Natl. Cancer Inst. Monogr., 51~117-122.
Plopper, C.G., and J.L. Alley (1983) Differentiation of tracheal epithe- lium in the fetal rhesus monkey. Anat. Rec., 205:155A.
Reznik-Schuller, H.M. (1982) Electron microscopic autoradiography of the trachea in fetal Syrian golden hamsters treated transplacen- tally with N-nitrosodiethylamine. J. Natl. Cancer Inst., 69t927- 931.
Said, J.W., G. Nash, S. Banks-Schlegel, A. Sassoon, S. Murakami, and I.P. Shintaku (1983) Keratin in human lung tumors. Patterns of localization of different molecular weight keratin proteins. Am. J. Pathol., 113:27-32.
Sorokin, S.P., R.F. Hoyt, and M.M. Grant (1982) Development of neu- roepithelial bodies in fetal rabbit lungs. I. Appearance and func- tional maturation as demonstrated by high-resolution light micro- scopy and formaldehyde-induced fluorescence. Exp. Lung Res., 3:237-259.
Steinman, R.M. (1968) An electron microscopic study of ciliogenesis in developing epidermis and trachea in the embryo of Xenopus laevis. Am. J. Anat., 122t19-56.
Stockinger, L., and E. Cireli (1965) Eine bisher unbekannte Art der Zentriolenvermehrung. 2. Zellforsch. Mikrosk. Anat., 68.733-740.
Sun, E.L., and C.J. Flickinger (1982) Proliferative activity in the rat epididymis during postnatal development. Anat. Rec., 203273- 284.
Thomas, P., J.W. Said, G. Nash, and S. Banks-Schlegel(1984) Profiles of keratin proteins in basal and squamous cell carcinomas of the skin. An immunohistochemical study. Lab Invest., 50:36-41.
Willis, R.A. (1962) Regeneration and repair: Resumed embryonic growth, In: The Borderland of Embryology and Pathology, 2nd ed. Butterworths, Washington, pp. 495-518.
Wilson, D.W., C.G. Plopper, and D.L. Dungworth (1984) The response of the macaque tracheobronchial epithelium to acute ozone injury. A quantitative ultrastructural and autoradiographic study. Am. J. Pathol., 116: 193-206.
Copyright © 2022 FDOKUMEN




















