Spatial-frequency- and Orientation Selectivity of Simple and ...
Upload
independentCategory
view
0download
0
Journal of Clinical InvestigationVol. 43, No. 10, 1964
The Selectivity of the HumanPlacenta in the Transfer ofPlasma Proteins from Mother to Fetus *
DAVID GITLIN, JESUS KUMATE, JUAN URRUSTI, ANDCARLOSMORALES
(From the Hospital Infantil de Mexico, the Hospital de Gineco-Obstetricia, Centro MedicoNacional del I.M.S.S., Mexico, D. F., the Children's Hospital Medical Center,
Boston, Mass., and the Children's Hospital, Pittsburgh, Pa.)
The passage of plasma proteins from motherto fetus in humans appears to be highly selective.Although a number of antibodies such as thoseagainst diphtheria (1, 2) and tetanus toxins (3),typhoid H antigen (4), and poliomyelitis virus(5) readily cross from the maternal plasma tothat of the fetus, other antibodies such as thoseagainst typhoid 0 antigen do not (4). This ap-parent variability in the maternofetal transfer ofplasma antibodies has been shown to depend uponthe class of plasma proteins to which the specificantibodies belong (6, 7): antibodies that are 7 Sy2-globulins pass from mother to fetus, but thoseantibodies that are 19 S /1-globulins do so muchless effectively or not at all. That the maternofetalbarrier may also be relatively impermeable to 71A-globulin antibodies and to ceruloplasmin and hap-toglobin as well is suggested by the observation thatthe concentrations of these proteins in the plasmaof the fetus at term are less than 15%o of those inthe maternal plasma (8-10). Although 19 S Yl-globulin is much larger in molecular weight than7 S -y-globulin, ceruloplasmin and haptoglobinhave molecular weights that are similar to orsmaller than that of 7 S y2-globulin. The selectivetransfer of 7 S y2-globulin across the maternofetalbarrier, therefore, may not be determined solelyby molecular size.
The nature of this selective maternofetal trans-fer of 7 S y2-globulin has been studied by Bram-bell and his colleagues in rabbits (11, 12). The7 S /,-globulin molecule can be hydrolyzed by
* Submitted for publication February 24, 1964; acceptedJune 22, 1964.
Presented in part before the American Pediatric So-ciety, Atlantic City, N. J., May 4, 1963. Supported bygrants from the Rockefeller Foundation, the NationalInstitutes of Health, U. S. Public Health Service (AM-00251 and HD-00652), and the Ronnie Miller Fund ofChildren's Hospital of Pittsburgh.
papain into three major polypeptide fragmentsnearly equal in molecular size: two of these frag-ments each contain a single antibody site, whereasthe third fragment has no antibody activity (13).By labeling such fragments obtained from rabbity2-globulin and injecting them into the pregnantdoe, the amount found in the fetus 24 hours latercould be determined; the data suggested that thetransmission of 7 S y2-globulin to the fetus de-pends upon the nonantibody portion of the mole-cule (12).
In this study the selectivity of the transfer of7 S y2-globulin was reinvestigated in the pregnantwoman by using several labeled human proteinsof different molecular weights and fragments of7 S y2-globulin obtained by papain hydrolysis.
Methods
Plan of study. Normal women in their last month ofan uncomplicated pregnancy were given 100 mg of stableKI orally per day. After administration of the KI wasbegun, the women were given a single intravenous in-jection of a specific human plasma protein labeled withI131; the injection was given at a time varying from a fewminutes to 4 weeks before delivery. Between the timeof injection and the time of confinement for the delivery,the women were ambulatory and attended the outpatientdepartment of the Hospital de Gineco-Obstetricia, CentroMedico Nacional del I.M.S.S., for routine prenatal care.The amount of labeled protein given each woman was2 mg or less, and the amount of radioactivity in eachinjection varied from 0.15 uc of 1131 per kg of bodyweight when radioiodinated 7 S 'y2-globulin was in-jected to 0.4 Atc of IP"' per kg of body weightwhen radioiodinated acid glycoprotein was studied.Measurements over the thyroidal area of several of thenewborn infants selected at random in this study indi-cated a level of radioactivity virtually identical to thatover a similar area of the thigh of the same infant with-out evidence of localized concentrations, indicating ef-fective inhibition of thyroidal uptake of radioiodide bythe stable iodide given the mother. The maximalamount of radiation received by the vascular system and
1938

PLACENTALTRANSFEROF PLASMAPROTEINS
its contents, which were the tissues of maximal concen-tration of the radioisotope, was less than 100 milliradsfor the mother and less than 30 millirads for the fetus.
The amount of radioactivity in the maternal plasma10 minutes after injection of the labeled protein at vari-ous intervals before delivery and during the delivery ofthe infant was determined as well as that in the infant'scord plasma at birth. When possible, samples of amnioticfluid were obtained per vagina when the mother's mem-branes ruptured; the amounts of radioactivity in sam-ples of this fluid were determined also.
Proteins studied. The preparations of acid glycopro-tein, albumin, transferrin, 7 S -y2-globulins, and fibrino-gen used in this study were obtained from the pooledplasma of normal adults by using the low temperatureethanol-water fractionation methods of Cohn and hiscolleagues.
1) Three times crystallized acid glycoprotein, or oroso-mucoid, was used.' The protein has a molecular weightof 44,000 (14).
2) The plasma albumin used was preparation decanol10, a twice crystallized albumin that has been character-ized physicochemically and immunochemically in earlierreports (15, 16).
3) The four times crystallized transferrin, or iron-bind-ing globulin,2 used was electrophoretically homogeneous,although traces of albumin and y2-globulin, less than 1%of each, were found by the quantitative immunochemicalprecipitation method. Transferrin has a molecular weightof approximately 90,000 (17, 18).
4) The 7 S 'y2-globulin preparation used had a sedi-mentation coefficient, S20w, of 6.8 S in a 1% solution,and it contained over 98% Y2-globulin as determined elec-trophoretically on paper and by moving boundary. Withthe quantitative precipitin method, the preparation wasfound to contain approximately 1% albumin.
5) The starting material for the purified fibrinogenused in this study was irradiated, Seitz-filtered FractionI of Cohn as prepared by the method of Blomback andBlomback (19); the fibrinogen was isolated from thisfraction by Dr. Edward Hershgold using the method ofMorrison, Edsall, and Miller (20). Approximately 95%of the protein in the final preparation of fibrinogen wasreadily clotted with thrombin, and the clottability of thepreparation was not affected significantly by radioiodina-tion.
6) Macroglobulins were isolated from the plasma of asingle adult donor by Dr. Philip Fireman and Dr. FredRosen: 25 ml of f resh plasma was passed through a2.5-cm by 150-cm column of Sephadex G-200 3 equili-brated with 0.1 M NaCl; the first portion of the firstprotein peak in the effluent was concentrated by nega-tive pressure dialysis and centrifuged in a sucrose gradi-ent (21), 10% at the top of the tube to 40% at the bot-
1 Kindly supplied by Dr. Karl Schmid of Boston Uni-versity.
2 Obtained from Dr. J. L. Oncley of the Universityof Michigan.
3 Pharmacia, Uppsala, Sweden.
tom, at 105,000 times g in a swinging bucket in a Spincomodel L ultracentrifuge for approximately 18 hours.The protein in the bottom 1 cm of the tube was collectedwith the aid of a tube slicer and was used, after dialysisagainst 0.15 M NaCl, without further purification. Theproteins in this preparation had a sedimentation velocity,S20,w, of approximately 19 S, and immunoelectrophoresisrevealed that the proteins present were a2-macroglobulinand y1-macroglobulin; after radioiodination and separa-tion of the radioactive proteins by paper electrophoresis,approximately 55%7 of the total radioactivity was associ-ated with a2-macroglobulin and 45% with y,-macroglobu-lin.
7) Polypeptide fragments were obtained from 7 S 82-globulin by hydrolysis with papain (13) at pH 7.0 in anenzyme to protein ratio of 1:1,000 by weight as describedelsewhere (22). These fragments were each approxi-mately 50,000 in molecular weight with a modificationof the sedimentation equilibrium method of McFar'laneand Kekwick (23), and three fragments were obtainedper hydrolyzed 7 S y2-globulin molecule. Two of thefragments each contained one of the two antibody sitesof the original 7 S ^y2-globulin antibody molecules; thesewere the S fragments (24). The remaining large frag-ment had little or no detectable antibody activity and wasreadily crystallized (23) ; this fragment was the F frag-ment (24). Isolation of these fragments from the papainhydrolysate of 7 S 'y2-globulin was performed by usingcolumns of DEAEcellulose and Sephadex G-100 as de-scribed elsewhere (22). The final preparation of F frag-ment contained less than 0.1% of S fragments asdetermined immunochemically. The preparation of Sfragments contained several immunochemically relatedcomponents, all of which were immunochemically differentfrom the F fragment; if any undegraded 7 S -y2-globulinwas present in this preparation, its concentration was lessthan 0.1% by weight as determined immunochemically.
8) Low molecular weight y-globulins were isolated asdescribed elsewhere (25) from the urine of normal adultmales whose blood had been transfused into sick childrenon several occasions without clinical manifestations ofhepatitis in the recipients. These proteins, which aretermed -y.-globulins in this report, were heterodisperse inthe ultracentrifuge, having an average sedimentation ve-locity of So0w = 1.5, with a range from less than 1 S anda molecular weight of 12,500 to 3 S with a molecularweight of 30,000 (25). The Py.-globulins were immuno-chemically related to the S fragments obtained from 7 Sy2-globulins: rabbit antisera prepared against the 'y-globulins cross-reacted with S fragments, and antiseraagainst S fragments cross-reacted with -y.-globulins.Like the S fragments, the 'y-globulins possessed anti-body activity (25) and were immunochemically distinctfrom the F fragment of 7 S -y2-globulin.
Radioiodiniation. The proteins were labeled with I13'by a modification of the nitrous acid method of Press-man and Eisen (26). Briefly, carrier-free P3' was ob-tained as iodide in bisulfite; 2 Amoles of stable Kb wasadded to the radioiodide for each micromole of protein
1939

D. GITLIN, J. KUMATE, J. URRUSTI, AND C. MORALES
to be iodinated and then oxidized to I2 with nitrous acid.After bringing the radioiodine solution to pH 7, it wasthen added to the protein, which was dissolved in bi-carbonate buffer at pH 9.3. The efficiency of iodinationby this method was approximately 15 to 20%, and the io-dinated protein contained an average of approximately0.5 atoms of iodine per molecule of protein. Nonproteinradioactivity was removed from the labeled protein solu-tions by passage through 1- by 5-cm columns of Dowex-2resin followed by dialysis against several changes of0.15 MNaCl for 24 hours. Sterilization of the final solu-tions was accomplished by pressure filtration through anultrafine sintered glass filter, and bacterial sterility wasconfirmed by culture on blood agar and in infusion broths.
Not more than 2% of the total radioactivity of thefinal preparation of each of the labeled proteins, with theexception of the labeled ay-globulins, remained unpre-cipitated in 10% trichloroacetic acid (TCA). Since alarge proportion, approximately 20%, of the unlabeled yu-globulin was not precipitable in 10% TCA, the amount ofprotein-bound radioactivity in the radioiodinated -y-glob-ulin preparation could not be determined by this method;however, less than 2% of the total radioactivity couldbe removed from the solution of labeled urine 'y-globulinby further dialysis against 0.15 M NaCl for 72 hours.
A single preparation of each of the radioiodinated pro-teins was used in this study with the exception of radio-iodinated 7 S 'y2-globulin. In the case of the latter pro-tein, two preparations of the labeled protein were used,
0~~~~~~~
0~~~~~
rR_
OM_ He . .vcocw W
0 /Io 20o 2x x
7Mt zft/6 m /(r/ON A* LoDaltz- DAVJ-
FIG. 1. DISAPPEARANCE OF RADIOIODINATED ALBUMIN
FROM THE PLASMA OF PREGNANT WOMEN (HOLLOWCIRCLES) AND THE CONCENTRATIONOF THIS PROTEIN IN
CORD PLASMA OF THEIR INFANTS (SOLID CIRCLES) RELA-
TIVE TO THE MATERNALPLASMACONCENTRATION10 MIN-
UTES AFTER INJECTION.
ooS L I a I0 5f /0 Iy 20 Zf )S72vo V77*v=N /IWAv/Or/ONANO /DZr
- DAKV -
FIG. 2. DISAPPEARANCEOF RADIOIODINATED 7 S 'y2-GLOB-ULIN FROM THE PLASMA OF PREGNANTWOMEN(HOLLOWCIRCLES) AND THE CONCENTRATIONOF THIS PROTEIN INCORDPLASMAOF THEIR INFANTS (SOLID CIRCLES) RELATIVETO THE MATERNALPLASMA CONCENTRATION10 MINUTESAFTER INJECTION.
13 women being studied with one and 10 women withthe other.
Estimation of radioactivity. Samples of plasma andamniotic fluid were placed in 4-ml vials, and the radio-activity in each sample was estimated with a 2-inch well-type NaI crystal; counting efficiency for 131 was ap-proximately 50%, and counting standards were used toavoid problems incident to radioactive decay, changes inthe counting efficiency of the system, and differences inthe geometry of the samples. Nonprotein radioactivitywas determined in samples of plasma and amniotic fluidby precipitating the proteins with an equal volume of 10%TCA, centrifuging, and estimating the radioactivity ina sample of the supernatant fluid.
In a few instances, zonal electrophoresis in starch (27),filtration through Sephadex (28), and precipitation withrabbit antiserum were used to determine the nature of theradioactivity in the sera of women given labeled S frag-ment and in the corresponding cord sera. The starchblocks used for electrophoresis measured 4 by 30 cm,and electrophoresis was performed in barbital buffer,pH 8.6, of ionic strength 0.1 at 15 v per cm for 18 hoursat 4° C. The blocks were then cut into segments 1 cm
wide and the proteins eluted with 0.15 M NaCl. Theeluates were assayed for radioactivity and the opticaldensities of the eluates determined in a Beckman spec-trophotometer at a wave length of 280 mo. Two differ-ent columns of Sephadex, each 2.5 cm wide, were usedfor each sample of serum to be studied; elution in eachwas with 0.1 M NaCl, and the eluates were collected in5-ml volumes. The serum was first passed through a
column consisting of a layer of G-25 over a layer ofG-50, each layer being 18 cm high; the optical densitiesof the eluates were determined at a wave length of 280
1940

PLACENTALTRANSFEROF PLASMAPROTEINS
mg and the eluates assayed for total and TCA-precipi-table radioactivity. The proteins excluded by this columnwere then pooled, and a 10-ml sample was passed throughthe second column of Sephadex, which consisted of layersof G-75, G-100, and G-200 f rom top to bottom, eachlayer being approximately 13 cm high. Again opticaldensities and total and TCA-precipitable radioactivityof the eluates were determined. The concentrations ofalbumin and 7 S -y2-globulin in the latter eluates werealso estimated.
The rabbit antiserum used for precipitating S frag-ments was one that was prepared against 7 S 'y2-globulin.The antiserum readily precipitated over 95% of the la-beled S fragment preparation used, whether the latterwas mixed with carrier S fragments or 7 S -y2-globulin;it did not precipitate F fragment.
Ininninochecmical cstimnation of plasna protein CoflCC1Z-trations. The concentrations of albumin, transferrin,7 S y2-globulin, and fibrinogen in maternal and cord sera(Table I) were estimated immunochemically by a uni-dimensional agar diffusion method (29) as described
TABLE I
Concentrations of specific proteins in maternal and cordplasma and in amniotic fluid
PlasmaAmniotic
Protein Mother Newborn fluid
g9/100 ml g/100 ml g/100 ml
Albumin 3.23 3.45 0.200Transferrin 0.401 0.179 0.0067 S 72-globulin 1.75 1.56 0.189Fibrinogen 0.538 0.392 0
elsewhere (30). The same method was used to esti-mate the concentrations of albumin and 7 S 'y2-globulinin the eluates obtained from the Sephadex columns.Briefly, the specific antiserum was incorporated into 0.5%agar in borate buffer of pH 8.6 and placed in glass tubes,10 cm long with an internal diameter of 4 mm, so thatabout half of the tube was filled; the top half of the tubewas filled with the fluid to be studied. After 24 hours,the distance between the agar-fluid interface and the pre-cipitin band that formed in the agar phase was measuredand the concentration of specific protein determined froma standard curve that was obtained with duplicate tubesand known concentrations of the given protein. Albuminand 7 S 'y2-globulin were estimated with horse antisera;transferrin and fibrinogen were determined with rabbitantisera.
ResultsI. The disappearance of radioiodinated proteins
from the plasma. The disappearance of radioio-dinated proteins from the plasma of pregnantwomenduring the last month of gestation is shownin Figures 1 to 4: the relative concentration of la-beled protein remaining in the plasma at the time
I+i
0/0
LK 0-612I0 5 /0 /7 20 Z J9P1-Wt &fTWZZV lNdT/1AND4i l/v[PY
- Da4 VJ -
FIG. 3. PLASMA DISAPPEARANCE OF RADIOIODINATED
19 S MACROGLOBULIN (X), TRANSFERRIN (SOLID DIA-
MONDS), ACID GLYCOPROTEIN (HOLLOW DIAMONDS), ANDFIBRINOGEN (CROSSES) IN PREGNANTWOMEN.
of delivery is given as Cm/CO, where CO is the con-centration of protein-bound radioactivity in theplasma of a given woman 10 minutes after in-jection, and Cm is that in the plasma of the samewoman at the time of the delivery of the infant.The logarithm of Cm/CO obtained for each motherinjected with a given protein was plotted againstthe time that had elapsed between the injection of
'9 0 5 /0 / x /5
-LZ)4k<5J -
FIG. 4. PLASMA DISAPPEARANCE OF RADIOIODINATEDF FRAGMENT (SOLID CIRCLES), S FRAGMENTS (HOLLOWCIRCLES), AND LOW MOLECULARWEIGHT URINE -y-GLOBU-LIN (DOTS) IN PREGNANTWOMEN.
1941

D. GITLIN, J. KUMATE, J. URRUSTI, AND C. MORALES
TABLE II
Half-lives of radioiodinated proteinsin the pregnant women at term
Number Amount Averageof of radio- half-life
mothers activity of proteinProtein studied injected in mother
aC/kg daysAcid glycoprotein 14 0.4 3.8Albumin 18 0.2 14.5Transferrin 16 0.3 1 27 S e2-globulin 23 0.15 20.5Fibrinogen 11 0.2 2.519 S macroglobulin 13 0.4 8.0F fragments 13 0.2 4.0S fragments 10 0.2 0.3
(10% =4.0)yu-Globulins* 15 0.2 0.4
* Low molecular weight urine -y-globulins.
the protein and the delivery. Since each deter-mination of Cm/COrepresents the plasma disappear-ance of a labeled protein in but one person, theaverage plasma disappearance curve thus obtainedfor a given protein represents a composite of thedeterminations of Cm/CO in 10 to 23 differentwomen. The average half-times of turnover or
half-lives, ti, of the proteins studied were estimatedfrom these plasma disappearance curves by simplegraphic analysis; the half-lives thus determined are
listed in Table II.The plasma disappearance of the labeled S frag-
ment preparation (Figure 4) beginning 3 to 4days after injection reached a relatively constantfractional rate representing a half-life of 4 days.If we assume a body distribution of the S frag-ments similar to that obtained for the other pro-
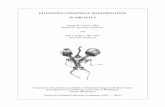
teins and the F fragment, the component witha half-life of 4 days would represent about 10%oof the total S fragment preparation; graphicanalysis of.the disappearance curve then yieldeda tj of plasma disappearance of 0.3 days for theremaining polypeptides in the S fragment prepa-ration during the period from approximately 8hours to 3 days after injection. In accord withthese differences in the tj of disappearance of dif-ferent fractions of the labeled S fragments, thenature of the protein-bound radioactivity presentin the maternal serum appeared to change withtime after injection. The protein-bound radioac-tivity present in the preparation of labeled S frag-ments mixed in vitro with normal serum and thatpresent in maternal serum obtained 10 minutesafter injection of the labeled S fragments had simi-lar properties by starch block electrophoresis and
Sephadex filtration: the protein-bound radioac-tivity had the mobility of a slow y2-globulin onstarch electrophoresis (Figure 5A) ; it was ex-cluded by Sephadex G-50, and it appeared withor just after albumin upon elution from the mixedcolumn of Sephadex G-75, G-100, and G-200(Figure 5E). In maternal serum obtained 8 to12 hours after injection, the mobility of the pro-tein-bound radioactivity appeared to be slightlyfaster than that present 10 minutes after injection(Figure 5B); the protein-bound radioactivity wasexcluded by Sephadex G-50, and it appeared withor somewhat before albumin, but after 7 S -y,-globulin, upon elution from mixed columns ofSephadex G-75, G-100, and G-200 (Figure 5C).Of the total radioactivity present in the maternalserum, variable amounts were included in Sepha-dex G-25 or G-50; the radioactivity in these frac-tions was not precipitable with TCA even in thepresence of added carrier serum.
The maternal plasma disappearance of labeledyu-globulins was followed for only 44 hours, butduring the time from approximately 8 hours afterthe injection until the end of the observation pe-riod, the tj of disappearance from the maternalplasma was 0.4 days; this tj is similar to that ob-tained for the major fraction of labeled S frag-ments during the same interval after injection.
In some instances, the plasma of the infant atbirth contained sufficient amounts of radioactivityto permit following the plasma disappearance ofthe labeled protein in the child over a period of 4to 10 days: the disappearance of radioiodinated7 S yn-globulin transferred from mother to fetuswas studied in 8 newborns, and the disappearanceof radioiodinated albumin transferred from motherto fetus was followed in 6 newborns. For both 7Sy2-globulin and albumin, the fractional rate ofdisappearance of the labeled protein from theplasma in most of the infants studied was almostconstant from birth, even if the infant was bornas soon as 24 hours after injection of the proteininto the mother, indicating that the rate of diffu-sion of these proteins from the infant's plasmainto the infant's extravascular fluids was greaterthan the rate of transfer of the proteins frommother to fetus. The estimated half-lives of theseproteins varied widely in the newborns that couldbe followed, from 7 days to 25 days for transferred
1942
PLACENTALTRANSFEROF PLASMAPROTEINS
labeled 7 S y2-globulin and from 5 days to 15 daysfor transferred labeled albumin.
II. The appearance of radioiodiniated proteinsin fetal plasma. The concentration of labeled al-bumin and 7 S y2-globulin found in the infant atbirth at various intervals after the injection ofthese proteins into the maternal circulation isshown in Figures 1 and 2. The infant's plasmaconcentration of labeled albumin was extremelyvariable: some infants had but 0.2 to 0.4% of theinitial maternal plasma concentration, whereasother infants had plasma concentrations of 1 to
5% of the initial maternal concentration in a simi-lar time interval. On the other hand, the infant'splasma concentration of labeled 7 S y2-globulin atbirth increased quite uniformly with increasingtime after injection of the labeled protein into themother, rising to levels above 10%o of the initialmaternal plasma concentration after 10 days.
Since the infant's plasma concentration of la-beled protein is dependent in part upon the ma-ternal plasma concentration of labeled protein, andsince the maternal concentration of 7 S 72-globu-lin decreased more slowly than that of labeled
/0o-
0a-l-/010
024-
04-
-4
ez4- , ~ -6.4
,044- /-20
-/-- 24*+4-444*+++ -2
4 ///S AS 0O 14
kc
00-
?u -. e*- \O .- .. ,". .
\0
0 *+"0 O.* --
/6' 1. Ao
D.A4N
60-
Jo-~ ~ ~ ~ ~ ~~J'-0- IJ okI
.0* \
0 -4 .',.-Oif /0 / =7
so ff
-200
0
7Vr5, ANUzAf-A'FIG. 5. A ANDB: ELECTROPHORESISOF MATERNALSERA ON STARCHBLOCK; 1-CM
SEGMENTS(ABSCISSA). A: 10 minutes after injection of S fragments. B: 8 hoursafter injection of S fragments. C THROUGHE: 5-ML VOLUMES ELUTED FROM
MIXED COLUMNSOF SEPHADEXG-75, G-100, AND G-200. C: Maternal serum ob-tained 12 hours after injection of S fragments. D: Cord serum obtained 12 hoursafter injection of S fragments. E: Serum from a normal adult to which was
added labeled S fragments in vitro. Symbols: solid line= optical density; crosses
=radioactivity; hollow circles= albumin concentrations (given along ordinate toright of figure in terms of mg per 100 ml X 10-1); solid circles = 7 S 'y2-globulinconcentrations (given along ordinate to right of figure in terms of mg per 100ml). The scale for radioactivity changes inl C through E.
-200 U
-/00»
-o k
-ZLI ts
-26,9%
-/00
1943
/,-I"
Sk
D. GITLIN, J. KUMATE, J. URRUSTI, AND C. MORALES
Q
40,1
E ,20
o
"R I4 a
ao.-0 i /O /5 20 25 .70 JfI I
INrPvAL LOe'irVLt I/VJtCT/ON AND L/rtVtp- DAVJ -
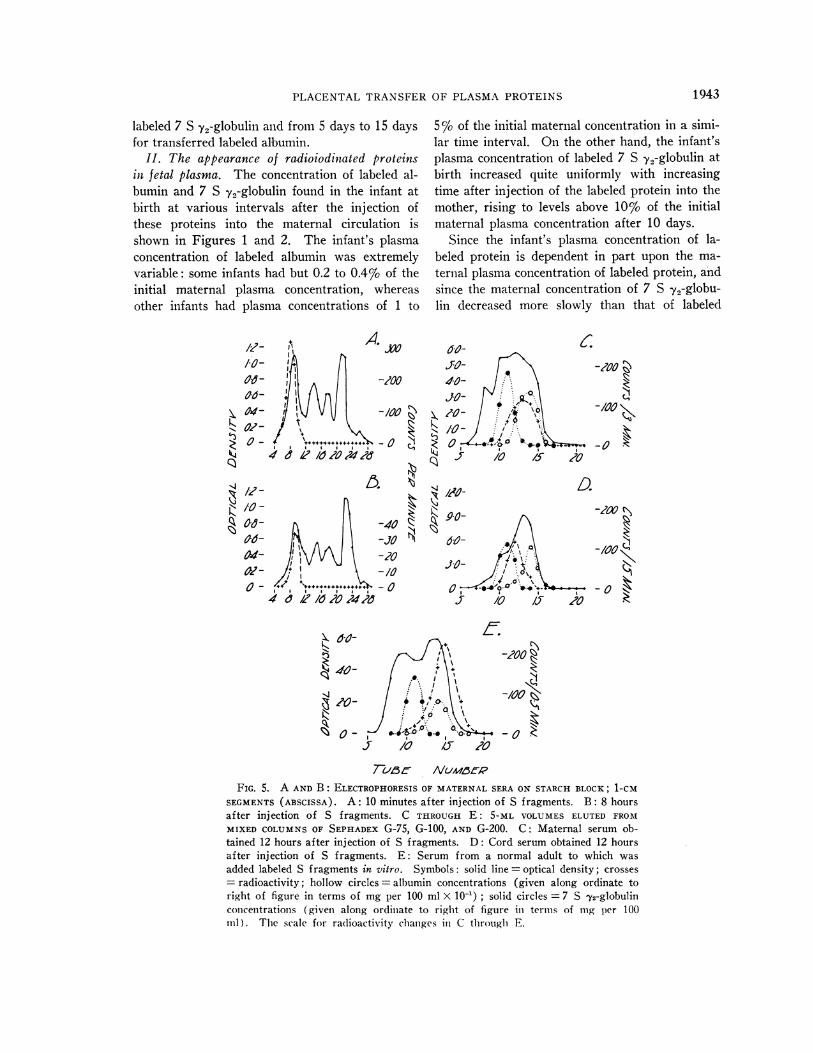
FIG. 6. CONCENTRATIONOF RADIOIODINATED 7 S y2-GLOBULIN (HOLLOW CIRCLES) AND ALBUMIN (SOLIDCIRCLES) IN CORD PLASMAOF THE INFANT AT BIRTH RELA-TIVE TO THE CONCENTRATIONSOF THESE PROTEINS IN THEMATERNALPLASMA AT DELIVERY.
albumin, the concentration of labeled protein in theinfant's plasma at delivery, C1, was expressed interms of the maternal plasma concentration at thattime, or Ci/Cm; the more rapid increase in theinfant's plasma concentration of 7 S 72-globulincompared to the increase found for labeled albu-min is readily apparent (Figure 6). The dataobtained for the other labeled proteins were simi-larly calculated, and the values of Ci/Cm at vari-ous time intervals between injection and deliveryare presented in Figures 7 to 10. The ordinatesin Figures 6 to 10 are linear, and the ordinates inFigures 1 to 4 are logarithmic; the values of Ci
) k 60
.46
~cz2
Ace OO /0 /5 20 25 Jo if
I I I
INfrf/?k4Z A MJ FfV IAAtcr/OAv ANDDZL-I4PV-DAYs
FIG. 7. CONCENTRATIONOF RADIOIODINATED ACID GLY-
COPROTEIN (HOLLOWDIAMONDS) AND TRANSFERRIN (SOLIDDIAMONDS) IN CORD PLASMA OF THE INFANT AT BIRTH
RELATIVE TO THE CONCENTRATIONSOF THESE PROTEINS IN
THE MATERNALPLASMA AT DELIVERY.
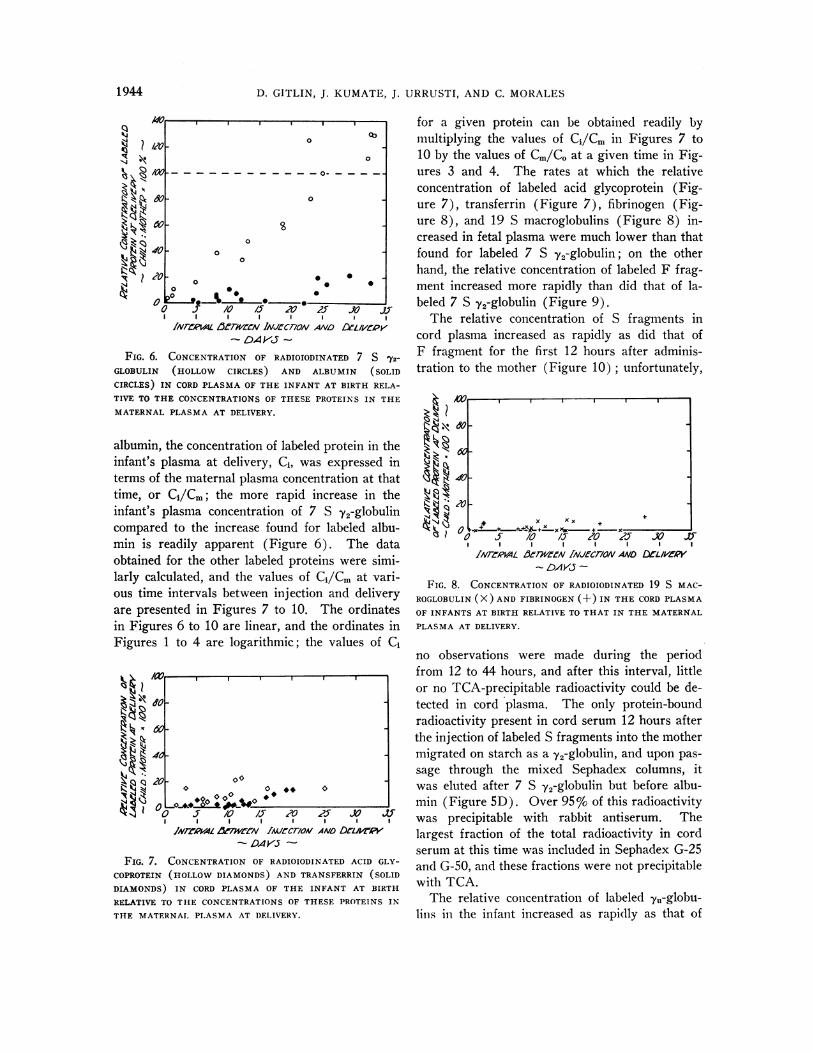
for a given protein can be obtained readily bymultiplying the values of Ci/Cr in Figures 7 to10 by the values of Cm/CO at a given time in Fig-ures 3 and 4. The rates at which the relativeconcentration of labeled acid glycoprotein (Fig-ure 7), transferrin (Figure 7), fibrinogen (Fig-ure 8), and 19 S macroglobulins (Figure 8) in-creased in fetal plasma were much lower than thatfound for labeled 7 S y2-globulin; on the otherhand, the relative concentration of labeled F frag-ment increased more rapidly than did that of la-beled 7 S y2-globulin (Figure 9).
The relative concentration of S fragments incord plasma increased as rapidly as did that ofF fragment for the first 12 hours after adminis-tration to the mother (Figure 10) ; unfortunately,
C 40
k 0013x /O 4 O2F'x5 /0 / 20 25 X7 if
I I
INrFzAWL &1r1'etN /iv1Ffcr/oN AND DtL/veW- DK45 -
FIG. 8. CONCENTRATIONOF RADIOIODINATED 19 S MAC-ROGLOBULIN(X) AND FIBRINOGEN (+) IN THE CORDPLASMAOF INFANTS AT BIRTH RELATIVE TO THAT IN THE MATERNALPLASMAAT DELIVERY.
no observations were made during the periodfrom 12 to 44 hours, and after this interval, littleor no TCA-precipitable radioactivity could be de-tected in cord plasma. The only protein-boundradioactivity present in cord serum 12 hours afterthe injection of labeled S fragments into the mothermigrated on starch as a y2-globulin, and upon pas-sage through the mixed Sephadex columns, itwas eluted after 7 S y2-globulin but before albu-min (Figure 5D). Over 95%o of this radioactivitywas precipitable with rabbit antiserum. Thelargest fraction of the total radioactivity in cordserum at this time was included in Sephadex G-25and G-50, and these fractions were not precipitablewith TCA.
The relative concentration of labeled y11-globou-lins in the infant increased as rapidly as that of
0
0~~~~~
_- - - - - - 0o _ -
0
00~~~~
0
0 0 S0 0 0 0
0- IL *aa
0
%xx +~~~~x +
0 00 10*0
1944
7.
-
-I. ..:E-
1; -'

PLACENTALTRANSFEROF PLASMAPROTEINS
F fragment during the 1-day period of observa-tion (Figure 10), with the exception of one in-fant delivered at 25 hours; no observations of therelative concentration of yu-globulins in cordplasma were made after 25 hours.
The cord plasma concentrations, CI, of labeledF fragment were found to be from 25 to 75% ofthose of 7 S 72-globulins during the period fromapproximately 1 day to 23 days after injectioninto the mother; 23 days was the longest intervalof time between the injection of F fragment anddelivery of the infant. The cord plasma concen-trations of 7,,-globulins and S fragments were verysimilar to each other, as indicated by the data inFigures 4 and 10, and these concentrations wereapproximately 10 to 25% of those for F fragmentduring the initial 12-hour period of observation ofS fragments and the 1-day period of observationof the -yu-globulins.
III. The appearance of radioiodinated proteinsin amniotic fluid. Of the four amniotic fluids ob-tained at the onset of delivery from women givenlabeled acid glycoprotein, the time intervals be-tween injection and procurement of the fluidsranged from 3 days, 11 hours to 9 days, 11 hours.None of these fluids had detectable TCA-precipita-ble radioactivity; the concentration of labeled acidglycoprotein in the corresponding newborn plasmawas 2.8 to 14.8% of that in the maternal plasmaat delivery.
Amniotic fluid was secured at delivery from sixwomengiven labeled albumin; of these, four fluidswere obtained from 3 to 9 hours after injection,and one fluid each was obtained after an intervalof 25 days, 18 hours and 32 days, 14 hours, re-spectively. The latter two fluids had no detectableTCA-precipitable radioactivity, although the new-born plasma contained 14.2 and 15.2%, respec-tively, of the amount present in the maternalplasma at that time. Protein-bound radioactivitywas present in the early four fluids, however, theconcentration being from 0.4 to 0.9%o of that inthe maternal plasma; the corresponding newbornplasma concentrations ranged from 1.4 to 2.7%.The specific activity of the latter four amnioticfluids, or the counts per minute per milligram ofendogenous albumin in a given sample, was ap-proximately 3 to 5 times that in the infant'splasma.
I.0~~~~
,120 fi so 04 60
/40
k440o dD 0 I0-0
0 Y_ /0 LT 20 2J
J1NZk-R04L AJrAZV5v//C7iWVANO DCLI/VLW'- DAVim-
FIG. 9. CONCENTRATIONOF RADIOIODINATED F FRAG-
MENT (SOLID CIRCLES), S FRAGMENTS(HOLLOWCIRCLES),AND LOW MOLECULARWEIGHT URINE -y-GLOBULIN (DOTS)IN THE CORDPLASMA OF INFANTS AT BIRTH RELATIVE TOTHE CONCENTRATIONSOF THESE PROTEINS IN THE MA-
TERNAL PLASMA AT DELIVERY.
Of five amniotic fluids obtained at the deliveryof mothers given labeled transferrin, the intervalbetween injection and delivery ranged from 4 daysto 19 days, 1 hour; two fluids contained no detec-table protein-bound radioactivity, and two con-
zo-
arl)f
x(I
11)ll.
1)k,4 Z,.
.k"T,
/0
a0ao / 2
D24k<
FIG. 10. CONCENTRATIONOF RADIOIODINATED F FRAG-
MENT, S FRAGMENTS, AND URINE -y-GLOBULIN (SYMBOLSAS IN FIGURE 9) IN THE CORD PLASMA OF INFANTS AT
BIRTH RELATIVE TO THE MATERNALPLASMA CONCENTRA-
TION OF THESE PROTEINS AT DELIVERY. This Figure is ain
expansion of the area about the origin of Figure 9,
0
0
0
00.
.00
o,
1945

D. GITLIN, J. KUMATE, J. URRUSTI, AND C. MORALES
tained 0.1 and 0.4%, respectively, of that in thematernal plasma.
Amniotic fluids were obtained from five womengiven labeled 7 S y2-globulin. For four of these,the interval between injection and delivery was4 hours. Two had no detectable protein-boundradioactivity, and two had only 0.1 %of the radio-activity found in the maternal plasma; the in-fants at this time had 1.2 to 1.9% of the concentra-tion of labeled 7 S 72-globulin in the maternalplasma. Of the two infants corresponding to thetwo amniotic fluids that contained protein-boundradioactivity, the plasma concentration of labeledprotein was 19 times that in the amniotic fluid, andthe specific activity of the plasma 7 S y2-globulinwas approximately 2.5 times that in the amnioticfluid. The fluids obtained after an interval of 22days, 22 hours had no detectable protein-boundradioactivity, whereas the concentration in the in-fant's plasma was 125% that of the mother.
Of the three amniotic fluids obtained at an in-terval of 4 hours to 8 days after the injection oflabeled fibrinogen and of the three amniotic fluidsobtained at an interval of 11 days to 16 days, 16hours after the injection of 19 S macroglobulins,none had detectable protein-bound radioactivity.
Discussion
Brambell and his colleagues have presented ex-tensive evidence in rodents to indicate that, in theseanimals, transfer of plasma proteins from motherto fetus is by way of the yolk sac splanchnopleur(11). They inferred from their findings that thepathway for maternofetal transfer of plasma pro-teins in primates might be the amniotic fluid.Wasz-H6ckert, Wager, Hautala, and Widholm(31) indicated that amniotic fluid may not be theroute of transfer of diphtheria antitoxin, a 7 S Y2-globulin antibody, in humans when they observedthat infants with esophageal atresia had amountsof this antibody in their plasma similar to those innormal children. Bangham, Hobbs, and Terry(32) injected radioiodinated homologous seruminto the amniotic fluid in pregnant rhesus monkeysand noted that little, if any, of the labeled proteinwas present in the fetal plasma 3 to 24 hourslater; in the rhesus monkey, the route of trans-fer appeared to be transplacental. Dancis and hisassociates (33) injected radioiodinated albumin
and y-globulin into the circulation of women at3-month gestation who were to undergo therapeu-tic abortion 18 to 24 hours later and observed thatthe amount of TCA-precipitable radioactivity perunit volume of amniotic fluid was less than thatpresent in fetal blood at the time of operation.To exclude a protein-concentrating mechanism bythe fetus, they injected human serum containingtetanus antitoxin directly into the amniotic fluidin women near term who were to undergo Ce-sarean section and found that after 18 to 24 hours,the concentration of tetanus antitoxin in fetal bloodwas less than that in the amniotic fluid. In thestudy reported here, the specific activity of 7 S72-globulin and transferrin in the amniotic fluid atdelivery was always lower than that in the plasmaof the infant, even when the protein had been givento the mother almost 3 weeks or more earlier; inseveral instances no labeled 7 S 72-globulin ortransferrin could be detected in amniotic fluidwhen the plasma of the newborn infant containedsignificant amounts of these proteins. Of themothers given labeled acid glycoprotein, fibrinogen,or 19 S macroglobulins, those amniotic fluids thatwere examined did not contain detectable amountsof these proteins, although the proteins were pres-ent in the plasma of the corresponding infants. Itwould seem quite clear, therefore, that amnioticfluid is not the route of transfer in man for acidglycoprotein, transferrin, 7 S y2-globulin, fibrino-gen, or 19 S macroglobulin. That amniotic fluidis probably not the route of transfer for labeledalbumin as well is indicated by the finding thattwo of the six amniotic fluids examined for thisprotein did not have detectable quantities, al-though significant amounts were present in theinfant. The site of transfer of plasma proteinsbetween mother and fetus in man appears to bethe placenta.
In their study of the selective transfer of 7 S72-globulin in pregnant rabbits, Brambell and hisassociates found that the serum concentration ofFragment III in the fetus 24 hours after placingthe labeled fragments in the uterine lumen was 6 to10 times that of Fragments I and II and only 25%of that of labeled 7 S Y2-globulin (12). In thepresent study in pregnant women, the concentra-tion of labeled F fragment, the analog of FragmentIII, found in cord plasma, Ci, was 4 to 10 timesthat of labeled S fragments, the analog of Frag-
1946

PLACENTALTRANSFEROF PLASMAPROTEINS
ments I and II, during the first 12 hours afteradministration to the mother, and varied from ap-proximately 25 to 75% of the plasma concentra-tion of 7 S y2-globulin over the 23-day period ofobservation of the F fragment. Thus, the resultspresented here, despite the differences in speciesstudied and the route of administration to themother, are actually very similar to those obtainedby Brambell. The yu-globulins, which are im-munochemically similar to S fragments in that thetwo groups of proteins have similar antigenic de-terminants, possess antibody activity, and are im-munochemically very different from F fragments,had cord plasma concentrations similar to those ofS fragments.
On the basis of their data, Brambell and co-workers suggested that the selective maternofetaltransfer of 7 S 79-globulin was attributable to thenonantibody region of the molecule (12). How-ever, the relative rate at which a labeled proteinappears in fetal plasma is not a simple measureof the maternofetal transfer of the protein; theconcentration of a given labeled protein in theplasma of the fetus is, of course, dependent upon1) the rate of transfer from mother to fetus,which in turn is dependent upon the permeabilityof the placenta to that protein as well as the con-centration of labeled protein in the maternalplasma, and 2) the rate of disappearance of theprotein from the fetal circulation, which in turnis dependent upon the rate of degradation of theprotein by the fetus, the net diffusion of labeledprotein from the fetal plasma into the interstitialfluids of the fetus, and the net transfer of the pro-tein from fetus to mother. Although two labeledproteins may cross the maternofetal barrier at thesame rate, if the rates of degradation of the twoproteins are different in either the fetus or in themother, their plasma concentrations in the fetus atany given moment will be different. The serumconcentration of labeled protein found in the rab-bit fetus, when expressed only in terms of theamount placed in the uterine lumen 24 hours ear-lier, as in the study by Brambell and co-workers,is not necessarily a valid measure of the materno-fetal transfer of that protein. Even if there wereno differences in the rates of diffusion of the vari-ous labeled proteins from the uterine lumen andwith the acceptance of Brambell's conclusion thattransfer in the rabbit occurs primarily via the
splanchnopleur so that the maternal concentrationof the protein need not be involved, such data donot reflect any possible differences in the half-livesof different proteins in the fetus. In the presentstudy, the half-life of F fragment in the pregnantwomen was more than 10 times longer than thatof the major portion of the S fragments but only20% of that of 7 S 72-globulin. To take some ac-count of these differences in half-life, the concen-tration of labeled protein in the cord plasma inthis report was expressed in terms of the ma-ternal plasma concentration present at the sametime. Calculated in this manner, the relative con-centration of S fragments in the fetus at term, atleast during the first 12 hours of observation, wassimilar to that of ye-globulins and of F fragment,and these concentrations were higher than thosefor 7 S 19-globulin. To calculate the maternofetaltransfer rates for these proteins, however, it isnecessary to know the rates of degradation ofthe proteins in the fetus and their rates of trans-fer from the fetus to mother, and this informationwas not obtained in the present study nor in thestudy by Brambell and associates.
An explanation for the increase in Ci/Cm forS fragments over the first 12 hours after injectionand then the absence of S fragments in the fetusby 44 hours after injection would seem to lie inthe metabolic heterogeneity of this polypeptidefraction. Approximately 90% of the preparationhad a half-life of 0.3 days, whereas the remainderhad a half-life of 4 days. A component with ahalf-life of 0.3 days would not readily be detectedin the fetus 44 hours after injection, although thecomponent with a 4-day half-life would be foundin the maternal plasma. The evidence obtainedby starch electrophoresis, Sephadex filtration, andimmunochemical precipitation suggests that theprotein-bound radioactivity present in cord plasmawas a component of the S fragment preparationand not a degradation product: it migrated as ay2-globulin, but appeared in fractions differentfrom y2-globulin when passed through Sephadex,and it reacted with specific antiserum. Althoughthe protein-bound radioactivity present in maternaland cord plasma 12 hours after the injection dif-fered considerably in electrophoretic and filtrationcharacteristics from the bulk of the labeled pro-tein present in the S fragment preparation beforeinjection, this is compatible with the demonstrated
1947

D. GITLIN, J. KUMATE, J. URRUSTI, AND C. MORALES
metabolic heterogeneity of the S fragment prepa-ration. Values obtained for Ci/Cm at 44 hoursafter injection and later suggest that the fractionpresent in the fetus had a half-life shorter than 4days or Ci/Cm would not have decreased, sincethe fraction that remained in the maternal plasmahad a half-life of 4 days. The values of Ci/Cmalso suggest that the component of 4-day half-liferemaining in the maternal plasma did not passfrom mother to fetus and that the S fragmentpreparation was heterogeneous as regards transferacross the placenta.
The observation that different heterologousplasma proteins may reach different fetal serumconcentrations after injection into the mother wasnoted by Brambell and co-workers (11) in rab-bits. Bangham and associates (32) clearly pointedout that after injection of labeled homologousproteins in the rhesus monkey, the relative con-centration of labeled albumin found in the fetuswas less than that of labeled y-globulin. Thiswas found to be the case in the human fetus of 3months gestation by Dancis and his associates (33)and in the human infant at term in this study.Apparently, each of the plasma proteins studiedmay pass from mother to fetus during the lastmonth of gestation, although there may be con-siderable variation from individual to individual inthe extent to which a given protein crosses theplacenta, and in some instances, relatively little orno labeled 19 S macroglobulin, transferrin, fibrino-gen, or even albumin appeared in the fetal plasma.The low relative plasma concentration of labeled19 S macroglobulins found in the fetus at term,however, is in accord with the relative deficiencyof this protein in the newborn (34). As statedearlier, however, relative fetal plasma concentra-tions of labeled proteins cannot be translated intotransfer rates from mother to fetus until data areobtained concerning the rates of degradation ofthese proteins in the fetus and their rates of trans-fer from fetus to mother; such information wasnot obtained in¶ this study nor in the earlierstudies.
The relative fetal plasma concentration of la-beled protein was determined as Ci/Cm. If anysignificant fraction of the labeled protein prepara-tions used had been denatured so as to precludematernofetal transfer, while such denatured pro-tein remained in the circulation, Ci/Cm would be
lower than if the denatured component were notpresent and would equal C/ (Cm. + Cd), where Cdrepresents the maternal plasma concentration ofthe denatured protein and Cm' is the maternalplasma concentration of the protein that can passto the fetus. Denaturation either during isolationof the unlabeled protein or during radioiodinationresults in rapid degradation of the labeled pro-tein, usually with almost complete elimination ofthe protein within 24 to 48 hours (35), or resultsin a decrease in the half-life of the protein, orboth (36). Thus, as the denatured protein is re-moved from the circulation, Ci/(Cm' + Cd) ap-proaches Ci/Cm, the same value as when no de-natured nontransferable protein is present in thepreparation. In any event, the average values ofCi/Cm for all of the labeled proteins, with theexception of S fragments, increased gradually overthe entire period of observation, whereas a sud-den increase during the first 24 to 48 hours wasnot observed, suggesting that denaturation seriousenough to result in rapid maternal degradationwithout maternofetal transfer was not a significantfactor in the present study; the reasons for thedecrease in Ci/Cm for S fragments after 12 hourswere discussed earlier. In addition, the averagehalf-life of the labeled 7 S 72-globulin preparationin the pregnant women studied was 20.5 days, avalue that agrees well with the average half-lifeof 23 days obtained by Cohen and Freeman (37)for radioiodinated 7 S y2-globulin in normal adults,the protein in the latter study being isolated byDEAEcellulose column chromatography and ra-dioiodinated by the iodine monochloride method(38). The half-life of transferrin in this studywas 12 days, longer than that obtained for trans-ferrin in normal adults by Katz, i.e., 6.7 to 8.4days, who used a relatively simple I3- method ofiodination (39). The half-life of 14.5 days ob-tained for the radioiodinated albumin preparationin these pregnant women is in accord with thehalf-life of this protein in normal adults reportedby Berson, Yalow, Schreiber, and Post (40) whoused a different method of iodination from thatused here. Although such evidence is not proofthat the labeled protein preparations in this studywere not denatured, at least the preparations ofalbumin, transferrin, and 7 S y2-globulin behavedas did simliar labeled preparations in which theproteins used were isolated or iodinated by meth-
1948

PLACENTALTRANSFEROF PLASMAPROTEINS
ods other than those used in this study. Sincethe S fragments and F fragments were obtainedby enzymatic digestion of 7 S 72-globulin, theyare, of course, degradation products of the parentprotein. The data in man and in animals wouldsuggest that the method of iodination as used inthis report would not significantly modify thehalf-lives of these proteins (41).
Although the transfer of 7 S 79-globulin was
observed for almost 35 days and the transfer ofF fragment was observed for 23 days, a steadystate for labeled protein between mother and in-fant, at which time Ci/Cm would become constant,was not reached. DuPan and his colleagues (42)similarly observed that radioiodinated 7 S y2-glob-ulin rapidly appears in the human fetus after in-jection of the protein into the mother and that thesteady state is reached only slowly.
Since the amount of labeled protein that passes
from mother to fetus was estimated in this studyby measuring the protein-bound radioactivity inthe cord plasma after delivery of the infant, itmight be argued that the radioactivity found inthe cord plasma was the consequence of a sud-den transfer of whole blood from mother to fetusvia the placenta during labor. Under these cir-cumstances, the relative concentration of labeledprotein in the infant's plasma at birth in per centof that found in the maternal plasma at deliverywould be at least [T/(P + T) ] 100 %, where T isthe amount of plasma transfused into the infantand P is the plasma volume of the infant plusplacenta. Therefore, to raise the child's plasmaconcentration only 1 % would require an average
transfusion of approximately 1.8 ml of plasma or
approximately 4.5 ml of blood. Since the average
concentration of each of the labeled proteins inthe infant's plasma at some time after injectionexceeded 5 to 10% of that in the maternal plasmaat delivery, the average amount of blood trans-fused into the fetus at this point would have hadto be approximately 22 to 44 ml of blood. In thecase of F fragment and 7 S 72-globulin where theinfant's concentration of labeled protein exceededthat in the maternal circulation, no amount of sud-den transfusion of blood from mother to fetuscould achieve this result. Thus, sudden transfu-sions from mother to infant during labor wouldnot explain the data obtained.
The occurrence of concentrations of labeled
proteins that are higher in the infant's plasma thanin the maternal plasma is simply the consequenceof tracer kinetics and does not indicate a higherconcentration of specific endogenous protein (41).In infants whose mothers had been given labeled7 S -2-globulin, for example, the plasma concen-tration of labeled protein exceeded that in themother 25 or more days after injection, but theconcentration of endogenous 7 S -y2-globulin inthe infant was equal to or slightly lower than thatin the mother. The injected tracer protein be-haves as do newly synthesized molecules of thatspecific protein entering the maternal vascularsystem.
Summary
Specific human plasma proteins were labeledwith I131 and injected into pregnant women in thelast month of gestation; the thyroids of bothmother and fetus were effectively blocked to theuptake of radioiodide by prior and continuing ad-ministration of stable iodide. Protein-bound ra-dioactivity in the plasma of the mother before andduring delivery was determined as well as thatpresent in the infant's plasma, and amniotic fluidwhen available, at birth. When possible, the pro-tein-bound radioactivity in the infant's plasmawas followed during the first few days of life.In this manner, 133 women and their respectiveinfants were studied with one of the followingproteins: acid glycoprotein, albumin, transferrin,7 S 72-globulin, fibrinogen, 19 S macroglobulin,F fragment from 7 S 72-globulin, S fragmentsfrom 7 S 72-globulin, and 1 S to 3 S urine y-glob-ulin (yu-globulin). The data indicate that: 1)All of the plasma proteins studied appeared inthe fetus after administration to the mother but indifferent amounts. The plasma concentrations oflabeled proteins in the fetus relative to those inthe mother decreased in approximately the fol-lowing order: F fragment, a major fraction ofS fragments and yu-globulins, 7 S y2-globulin,acid glycoprotein, albumin, transferrin, 19 S mac-roglobulin, and fibrinogen. In some instances,little or no labeled 19 S macroglobulin, transferrin,fibrinogen, or even albumin was present in theinfant's plasma at birth. 2) The relative plasmaconcentrations of F fragment, or the nonantibodyportion of the 7 S 72-globulin molecule, a majorfraction of the S fragments, or those fragments
1949

D. GITLIN, J. KUMATE, J. URRUSTI, AND C. MORALES
of the 7 S y2-globulin molecule that possess anti-body activity, and the yu-globulins, which are re-lated immunochemically to the S fragments, weresimilar to each other, but they were higher thanthe relative plasma concentrations of 7 S 'y2-globu-lin.
The problem of translating fetal plasma con-centrations of labeled protein into maternofetaltransfer rates in the absence of data concerningrates of degradation in the fetus and rates oftransfer from fetus to mother is discussed.
Acknowledgments
We express our gratitude to Dr. Benjamin Eguiluz,Director of the Hospital de Gineco y Obstetricia, CentroMedico Nacional del I.M.S.S. and to Dr. FedericoGomez, formerly Director of the Hospital Infantil andnow Director of the Hospital de Pediatria, CentroMedico Nacional del I.M.S.S., Mexico, D. F., for theirkindness, understanding, and encouragement in makingpossible our use of the facilities of those institutions forthis study. We are also indebted to Dr. Fred Rosen ofthe Children's Hospital Medical Center, Boston, Dr.Pablo Yoshida of the Hospital Infantil de Mexico, andMiss Mary Boesman of the Children's Hospital of Pitts-burgh for their tireless assistance in various phases ofthis work.
References
1. Osborn, J. J., J. Dancis, and B. V. Rosenberg.Studies of the immunology of the newborn infant:III. Permeability of the placenta to maternal anti-body during fetal life. Pediatrics 1952, 10, 450.
2. Vahlquist, B. Studies on diphtheria: I. The de-crease of natural antitoxic immunity against diph-theria. Acta paediat. (Uppsala) 1948, 35, 117.
3. TenBroeck, C., and J. H. Bauer. The transmissionof tetanus antitoxin through the placenta. Proc.Soc. exp. Biol. (N. Y.) 1923, 20, 399.
4. Timmerman, W. A. Zur Frage der tYbertragungdes Typhus "H" and "O"-Agglutinin von Mutterauf Kind. Z. Immun.-Forsch. 1931, 70, 388.
5. Perkins, F. T., R. Yetts, and W. Gaisford. Sero-logical response of infants to poliomyelitis vaccine.Brit. med. J. 1958, 2, 68.
6. Kochwa, S., R. E. Rosenfield, L. Tallal, and L. R.Wasserman. Isoagglutinins associated with ABOerythroblastosis. J. clin. Invest. 1961, 40, 874.
7. Gitlin, D., F. S. Rosen, and J. G. Michael. Transient19S gamma1-globulin deficiency in the newborninfant, and its significance. Pediatrics 1963, 31,197.
8. Hitzig, W. H. Das Bluteiweissbild beim gesundenSaugling. Helv. paediat. Acta 1961, 16, 46.
9. Scheinberg, I. H., C. D. Cook, and J. A. Murphy.The concentration of copper and ceruloplasmin in
maternal and infant plasma at delivery (abstract).J. clin. Invest. 1954, 33, 963.
10. Smithies, 0. Zone electrophoresis in starch gels:group variations in the serum proteins of normalhuman adults. Biochem. J. 1955, 61, 629.
11. Brambell, F. W. R., W. A. Hemmings, and M. Hen-derson in Antibodies and Embryos. London, Ath-lone Press, 1951.
12. Brambell, F. W. R., W. A. Hemmings, C. L. Oakley,and R. R. Porter. The relative transmission ofthe fractions of papain hydrolyzed homologous-y-globulin from the uterine cavity to the foetalcirculation in the rabbit. Proc. roy. Soc. B 1960,151, 478.
13. Porter, R. R. The hydrolysis of rabbit y-globulinand antibodies with crystalline papain. Biochem.J. 1959, 73, 119.
14. Smith, E. L., D. M. Brown, H. E. Weimer, andR. J. Winzler. Sedimentation, diffusion, and mo-lecular weight of a mucoprotein from humanplasma. J. biol. Chem. 1950, 185, 569.
15. Cohn, E. J., W. L. Hughes, Jr., and J. H. Weare.Preparation and properties of serum and plasmaproteins. XIII. Crystallization of serum albuminsfrom ethanol-water mixtures. J. Amer. chem.Soc. 1947, 69, 1753.
16. Gitlin, D., and C. A. Janeway. An immunochemicalstudy of the albumins of serum, urine, ascitic fluidand edema fluid in the nephrotic syndrome. J.clin. Invest. 1952, 31, 223.
17. Koechlin, B. A. Preparation and properties of se-rum and plasma proteins. XXVIII. The fi1-metal-combining protein of human plasma. J. Amer.chem. Soc. 1952, 74, 2649.
18. Schultze, H. E., I. G6llner, K. Heide, M. Sch6nen-berger, and G. Schwick. Zur Kenntnis der a-Glob-uline des menschlichen Normalserums. J. Natur-forsch. 1955, 10b, 463.
19. Blomback, B., and M. Blomback. Purification of hu-man and bovine fibrinogen. Arkiv Kemi 1956,10, 415.
20. Morrison, P. R., J. T. Edsall, and S. G. Miller.Preparation and properties of serum and plasmaproteins. XVIII. The separation of purified fibrin-ogen from fraction I of human plasma. J. Amer.chem. Soc. 1948, 70, 3103.
21. Kunkel, H. G., J. H. Rockey, and T. Tomasi.Methods of separation and properties of antibodiesof high molecular weight in Immunochemical Ap-proaches to Problems in Microbiology, M. Heidel-berger and 0. J. Plescia, Eds. New Brunswick,N. J., Rutgers University Press, 1961, p. 30.
22. Hershgold, E. J., F. Cordoba, P. Charache, and D.Gitlin. A crystalline fragment from humangamma-globulin. Nature (Lond.) 1963, 199, 284.
23. McFarlane, A. S., and R. A. Kekwick. Physicalproperties of Bushy Stunt virus protein. Biochem.J. 1938, 32, 1607.
24. Edelman, G. M., J. F. Heremans, M.-Th. Heremans,and H. G. Kunkel. Immunological studies of hu-
1950

PLACENTALTRANSFEROF PLASMAPROTEINS
man y-globulin. Relation of the precipitin linesof whole y-globulin to those of the fragments pro-duced by papain. J. exp. Med. 1960, 112, 203.
25. Merler, E., J. S. Remington, M. Finland, and D.Gitlin. Characterization of antibodies in normalhuman urine. J. clin. Invest. 1963, 42, 1340.
26. Pressman, D., and H. N. Eisen. The zone of locali-zation of antibodies. V. An attempt to saturateantibody-binding sites in mouse kidney. J. Im-munol. 1950, 64, 273.
27. Kunkel, H. G., and R. J. Slater. Zone electrophore-sis in a starch supporting medium. Proc. Soc. exp.Biol. (N. Y.) 1952, 80, 42.
28. Porath, J., and P. Flodin. Gel filtration: a methodfor desalting and group separation. Nature(Lond.) 1959, 183, 1657.
29. Oudin, J., and P. Grabar. e-tude quantitative du sys-teme precipitant ovalubumine-anticorps homologuede lapin. Ann. Inst. Pasteur 1944, 70, 7.
30. Janeway, C. A., and D. Gitlin. The gamma-globu-lins. Advanc. Pediat. 1957, 9, 65.
31. Wasz-H6ckert, O., 0. Wager, T. Hautala, and 0.Widholm. Transmission of antibodies from motherto fetus. A study of the diphtheria antitoxin levelin the newborn with oesophageal atresia. Ann.Med. exp. Fenn. 1956, 34, 444.
32. Bangham, D. R., K. R. Hobbs, and R. J. Terry. Se-lective placental transfer of serum-proteins in therhesus. Lancet 1958, 2, 351.
33. Dancis, J., J. Lind, M. Oratz, J. Smolens, and P.Vara. Placental transfer of proteins in humangestation. Amer. J. Obstet. Gynec. 1961, 82, 167.
34. Hitzig, W. H. Die physiologische Entwicklung der"Immunglobuline" (gamma- und beta2-Globuline).Helv. paediat. Acta 1957, 12, 596.
35. Margen, S., and H. Tarver. Comparative studies onthe turnover of serum albumin in normal humansubjects. J. clin. Invest. 1956, 35, 1161.
36. McFarlane, A. S. Labeling of plasma proteins withradioactive iodine. Biochem. J. 1956, 62, 135.
37. Cohen, S., and T. Freeman. Metabolic heterogeneityof human y-globulin. Biochem. J. 1960, 76, 475.
38. McFarlane, A. S. Efficient trace-labeling of proteinswith iodine. Nature (Lond.) 1958, 182, 53.
39. Katz, J. H. Iron and protein kinetics studied bymeans of doubly labeled human crystalline trans-ferrin. J. clin. Invest. 1961, 40, 2143.
40. Berson, S. A., R. S. Yalow, S. S. Schreiber, and J.Post. Tracer experiments with I13" labeled humanserum albumin: Distribution and degradation stud-ies. J. clin. Invest. 1953, 32, 746.
41. Gitlin, D., and C. A. Janeway. Some isotopic studieson the distribution and metabolism of plasma pro-teins. Advanc. biol. med. Phys. 1960, 7, 249.
42. DuPan, R. M., P. Wenger, S. Koechli, J.-J. Scheideg-ger, and J. Roux. etude du passage de la y-globu-line marquee a travers le placenta humain. Clin.chim. Acta 1959, 4, 110.
1951
Copyright © 2022 FDOKUMEN