
Organizing moving groups during morphogenesis
6
Organizing moving groups during morphogenesis Virginie Lecaudey and Darren Gilmour The directed migration of cells drives the formation of many complex organ systems. Although in this morphogenetic context cells display a strong preference for migrating in organized, cohesive groups, little is known about the mechanisms that coordinate their movements. Recent studies on several model systems have begun to dissect the organization of these migrating tissues in vivo and have shown that cell guidance is mediated by a combination of chemical and mechanical cues. Addresses European Molecular Biology Laboratory, Meyerhofstrasse 1, 69117 Heidelberg, Germany Corresponding author: Gilmour, Darren ([email protected]) Current Opinion in Cell Biology 2006, 18:102–107 This review comes from a themed issue on Cell structure and dynamics Edited by J Victor Small and Michael Glotzer Available online 13th December 2005 0955-0674/$ – see front matter # 2005 Elsevier Ltd. All rights reserved. DOI 10.1016/j.ceb.2005.12.001 Introduction Cell migration is essential to many physiological and disease processes such as embryonic morphogenesis, wound healing and cancer metastasis. Studies on single motile cells in culture have lead to a well-established model whereby cells move via the extension and adhesion of a leading edge pointed in the direction of migration and the retraction and loss of adhesion of the trailing edge at the rear. Here, the forces required for the translocation of the cell body are generated at the points of contact with the flat substrate provided by the Petri dish. While these studies have been crucial in understanding the mechanics of cell motility, it is clear that this controlled experimental environment is very different from what cells experience in the three-dimensional context of living tissues. How- ever, thanks to improvements in microscopy technology it is now possible to observe cells migrating in their natural habitats, such as the intact developing embryo. What has become clear from imaging studies is that during mor- phogenesis cells do not usually travel alone but rather prefer to undertake journeys together, often moving in very large numbers. In some cases cells move as dense streams of freely migrating chemotactic individuals that coalesce at particular locations, with each cell apparently being guided directly by extrinsic cues [1 ]. However, during the morphogenesis of many organ systems it is more common to find cells migrating in some form of adherent group or as tissues. These ‘tissue migrations’ are the focus of this review. Migrating tissues come in many shapes and sizes and show varying degrees of cohesion and organization (for more details see [2]) (Figure 1). These range from rather loose networks, such as chains of neuronal progenitors in the CNS [3,4] and migrating neural crest cells [5 ], to tightly adherent sheets of epithelial cells, where a large number of cells move as a single coherent unit and maintain constant positions throughout [6]. Elsewhere, they can be found as clusters of motile cells, as exempli- fied by Drosophila border cells (BCs) [7,8] or the migrating primordium of the lateral line (LLP) in amphibia and fish [9,10]. Migrating tissues are often employed in sculpting complex three-dimensional forms, including the intricate tubular networks present in the vasculature and the respiratory system. While their forms and functions are diverse, it is clear in all cases that the migratory behaviour of cells within these various tissues must be coordinated to ensure proper movement of the entire group. The aim of this review is to integrate some recent results from several experimental models that shed light on the mechanisms ensuring the concerted movement of tissues during morphogenesis. Because of space limitations, we will not discuss convergent-extension movements during gastrulation, a very important example of collective cell behaviour that has been covered by several excellent recent reviews [11,12]. Getting organized for the journey What guides cell groups on their journey? Genetic studies in a wide range of model systems have shown that tissue migration is regulated by the very same extrinsic chemical cues that guide single cells. Examples include members of the epidermal growth factor and fibroblast growth factor families, which are detected through receptor tyr- osine kinases present in the plasma membrane. As these are known to guide single cells via a chemotactic mechan- ism, it is likely that graded distributions of these factors also determine the directionality of tissue migration in many cases [1 ,13]. An important issue regarding cells moving as a cohesive tissue is the extent to which external gradients penetrate multicellular cohorts to control migra- tion behaviour within. It is becoming clear that extrinsic cues drive the movement of tissues not by acting directly on all members of the group but rather by instructing smaller numbers of peripheral leader cells that in turn are responsible for the guidance of naı¨ve followers. This is Current Opinion in Cell Biology 2006, 18:102–107 www.sciencedirect.com
-
Upload
uni-freiburg -
Category
Documents
-
view
0 -
download
0
Transcript of Organizing moving groups during morphogenesis

Organizing moving groups during morphogenesisVirginie Lecaudey and Darren Gilmour
The directed migration of cells drives the formation of many
complex organ systems. Although in this morphogenetic
context cells display a strong preference for migrating in
organized, cohesive groups, little is known about the
mechanisms that coordinate their movements. Recent studies
on several model systems have begun to dissect the
organization of these migrating tissues in vivo and have shown
that cell guidance is mediated by a combination of chemical
and mechanical cues.
Addresses
European Molecular Biology Laboratory, Meyerhofstrasse 1, 69117
Heidelberg, Germany
Corresponding author: Gilmour, Darren ([email protected])
Current Opinion in Cell Biology 2006, 18:102–107
This review comes from a themed issue on
Cell structure and dynamics
Edited by J Victor Small and Michael Glotzer
Available online 13th December 2005
0955-0674/$ – see front matter
# 2005 Elsevier Ltd. All rights reserved.
DOI 10.1016/j.ceb.2005.12.001
IntroductionCell migration is essential to many physiological and
disease processes such as embryonic morphogenesis,
wound healing and cancer metastasis. Studies on single
motile cells in culture have lead to a well-established
model whereby cells move via the extension and adhesion
of a leading edge pointed in the direction of migration and
the retraction and loss of adhesion of the trailing edge at
the rear. Here, the forces required for the translocation of
the cell body are generated at the points of contact with
the flat substrate provided by the Petri dish. While these
studies have been crucial in understanding the mechanics
of cell motility, it is clear that this controlled experimental
environment is very different from what cells experience
in the three-dimensional context of living tissues. How-
ever, thanks to improvements inmicroscopy technology it
is now possible to observe cells migrating in their natural
habitats, such as the intact developing embryo. What has
become clear from imaging studies is that during mor-
phogenesis cells do not usually travel alone but rather
prefer to undertake journeys together, often moving in
very large numbers. In some cases cells move as dense
streams of freely migrating chemotactic individuals that
coalesce at particular locations, with each cell apparently
Current Opinion in Cell Biology 2006, 18:102–107
being guided directly by extrinsic cues [1��]. However,
during the morphogenesis of many organ systems it is
more common to find cells migrating in some form of
adherent group or as tissues. These ‘tissue migrations’ are
the focus of this review.
Migrating tissues come in many shapes and sizes and
show varying degrees of cohesion and organization (for
more details see [2]) (Figure 1). These range from rather
loose networks, such as chains of neuronal progenitors in
the CNS [3,4] and migrating neural crest cells [5�], totightly adherent sheets of epithelial cells, where a large
number of cells move as a single coherent unit and
maintain constant positions throughout [6]. Elsewhere,
they can be found as clusters of motile cells, as exempli-
fied byDrosophila border cells (BCs) [7,8] or the migrating
primordium of the lateral line (LLP) in amphibia and fish
[9,10]. Migrating tissues are often employed in sculpting
complex three-dimensional forms, including the intricate
tubular networks present in the vasculature and the
respiratory system. While their forms and functions are
diverse, it is clear in all cases that the migratory behaviour
of cells within these various tissues must be coordinated
to ensure proper movement of the entire group. The aim
of this review is to integrate some recent results from
several experimental models that shed light on the
mechanisms ensuring the concerted movement of tissues
during morphogenesis. Because of space limitations, we
will not discuss convergent-extension movements during
gastrulation, a very important example of collective cell
behaviour that has been covered by several excellent
recent reviews [11,12].
Getting organized for the journeyWhat guides cell groups on their journey? Genetic studies
in a wide range of model systems have shown that tissue
migration is regulated by the very same extrinsic chemical
cues that guide single cells. Examples include members
of the epidermal growth factor and fibroblast growth
factor families, which are detected through receptor tyr-
osine kinases present in the plasma membrane. As these
are known to guide single cells via a chemotactic mechan-
ism, it is likely that graded distributions of these factors
also determine the directionality of tissue migration in
many cases [1��,13]. An important issue regarding cells
moving as a cohesive tissue is the extent to which external
gradients penetrate multicellular cohorts to control migra-
tion behaviour within. It is becoming clear that extrinsic
cues drive the movement of tissues not by acting directly
on all members of the group but rather by instructing
smaller numbers of peripheral leader cells that in turn are
responsible for the guidance of naıve followers. This is
www.sciencedirect.com

Organizing moving groups during morphogenesis Lecaudey and Gilmour 103
Figure 1
Cells move in groups of varying shape and size in vivo. (a) The concerted movement of epithelial sheets is a very prevalent feature of morphogenesis,
as demonstrated here during dorsal closure in the Drosophila embryo. (b) The coordinated migration of groups of epithelial cells also drives the
formation of branched tubular networks, such as the Drosophila tracheal system. (c) Chain migration of neuronal precursors in the subventricular
zone of the adult rodent brain. (d,e) Drosophila border cells and the zebrafish lateral line primordium as two examples of cells that migrate in
clusters or cohorts. (Reproduced with permission of (a) Ferenc Jankovics and Damian Brunner; (b) Stefan Luschnig; (c) Arturo Alvarez-Buylla, in [4];
(d) Pernille Rorth, in [18]).
suggested by the fact that in many contexts only a subset
of cells within a tissue display morphological features,
such as filopodia and pseudopodia, characteristic of migra-
tory cells [14]. Further support comes from several studies
where guidance receptor activation is assayed directly
using antibodies that bind specifically to active forms of
receptors or downstream signalling components, allowing
the identification of responsive cells. This approach was
first used with anti-phospho-MAPK (Erk) antibodies to
show that FGF signalling becomes restricted to the tips of
Drosophila tracheal branches soon after they begin to
extend [15]. More recently, anti-phosphotyrosine antibo-
dies have been used as a read-out of guidance receptor
activation to show that during normal migration only a
subset of BCs responds to the cue secreted by the oocyte
[16]. Similarly, during eyelid closure in mouse embryos,
the EGF-like growth factor HB-EGF binds to and acti-
vates the EGF receptor and the downstream ERK signal-
ling cascade only at the leading edge of the migrating
epithelial sheet [17]. The most direct experimental
demonstration that not all cells within migrating tissues
need to respond to cues in vivo comes from genetic mosaic
studies that juxtapose wild-type and migration-defective
mutant neighbours. This approach has been particularly
informative in the case of Drosophila BC clusters, where
wild type cells have been mixed with several different
immotile mutants including slbo, shg (E-cad) and sqh(myosin II). Here the wild-type cells can rescue the
migration of immobile mutant clusters with an efficiency
www.sciencedirect.com
depending on their proportion [18–20]. These combined
findings demonstrate clearly that guidance within tissues
can be non-cell-autonomous, and that groups are com-
posed of cells that respond directly to extrinsic cues and
cells that do not.
Coordinating individual movements withinmoving groupsChemotaxis: tips from a slimy collaborator
How do these leading cells transmit this extrinsic direc-
tional information to the remainder of the group? One
paradigm for how cells within motile groups can organize
each other’s behaviour comes from Dictyostelium slugs,
which are comprised of many thousands of migrating cells
that move collectively [21]. Here, a specialized set of cells
at the tip of the slug, known as the prestalk cells, form an
internal source of the diffusible chemoattractant cAMP
that drives periodic waves of migration throughout the
entire mass. Responding posterior cells are dependent on
this internally generated gradient for their motility; if the
tip region is cut off, it continues to migrate while the
remainder of the slug is rendered immobile [22]. While
tissues moving through embryos are guided by extrinsic
cues, it is possible that leading cells adopt a similar
strategy to organize the migratory behaviour of neigh-
bours through a relay of guidance molecules. It will be
interesting to determine whether the expression of che-
moattractants within moving cohorts provides a mechan-
ism for coordinating their behaviour in vivo.
Current Opinion in Cell Biology 2006, 18:102–107

104 Cell structure and dynamics
Mechanotaxis: can you feel the force?
In many situations it is more likely that leading cells
organize their neighbours by translating extrinsic chemi-
cal guidance cues into directional mechanical force. The
force in question originates at least in part when leading
cells retract their trailing edges, an event that in single
cells is known to be dependent on the activity of myosin
II [23]. Interestingly, Dictyostelium mutants lacking myo-
sin II are able to migrate as single cells but cannot make
slugs, suggesting that force generated through the actin–
myosin network plays an important role in allowingmulti-
cellular movements [24,25]. Likewise, in BC clusters
where the activity of myosin II is reduced, the leading
cell continues to project long cellular extensions (LCEs)
in the direction of migration, suggesting it can respond to
guidance cues, but is unable to translocate its cell body or
the remainder of the cluster, supporting the idea that
contraction of the leading cells generates the force that
initiates group migration [18,26].
An open question in the field is how followers detect the
mechanical tension generated by leading cells in response
to chemical cues. Focal adhesion complexes that form
between cells and the substratum have been shown to
mediate the conversion of external mechanical forces into
biochemical intracellular responses [27]. In the case of
cohesive groups, it is likely that mechanical force is
transduced at points of cell–cell contact, with adherens
junctions (AJs) being the best candidates, a suggestion
supported by recent work on cultured endothelial cells
(ECs). As ECs line the luminal surface of blood vessels
they are constantly exposed to pressure from the passing
flow, termed shear stress, and it is known that hemostatic
pressure influences many aspects of EC behaviour,
including the directionality of their migration [28]. In a
recent paper, Tzima and colleagues have described a
mechanosensory complex present at AJs; this complex
comprises two cell adhesion molecules (PECAM-1 and
vascular endothelial cell cadherin) and a receptor tyrosine
kinase (VEGFR2) and is required for the coordinated
alignment of ECs in response to shear pressure [29��].Cadherins have also previously been shown to mediate
mechanical signalling at cell–cell junctions in fibroblasts
through the activation of stretch-sensitive calcium chan-
nels leading to an increase in actin polymerization [30]
(Figure 2). Calcium signalling has also been shown
recently to modulate cell speed in response to shear stress
in Dictyostelium [31] and to be involved in detachment of
the rear of migrating fish keratocytes [32]. This is parti-
cularly intriguing as it has been proposed that the pro-
pagation of calciumwaves across fields of cells may be one
mechanism for coordinating their movements [33].
How do cells respond when placed under mechanical
stress by their more informed neighbours? Several recent
studies indicate that, rather than simply coming along for
the ride, cells within tissues react in a way that enhances
Current Opinion in Cell Biology 2006, 18:102–107
the mobility of the group. By scratching ‘wounds’ in
cultured MDCK epithelial monolayers, Farooqui and
Fenteany have demonstrated that cells several rows
behind the wound margin extend ‘cryptic’ lamellipodia
basally while maintaining continuous cell–cell contacts
apically, suggesting that these cells are actively involved
in driving collective migration [34�]. Interfering with
either intracellular calcium or gap junctions had no effect
on this behaviour, leading to the proposal that it may be
stimulated through a direction-sensingmechanotransduc-
tion system present at apically located junctions. Using
the same culture system it has recently been shown that
scratching induces two waves of ERK1/2-type MAP
kinase activation propagating from the wound edge to
submarginal cells [35]. The second wave is tightly corre-
lated with the motility of the cells; it is no longer present
as the wound is closing and inhibition of ERK1/2 activa-
tion results in a clear reduction in the rate of migration.
Although the mechanism of propagation is not known, it
is suggested once again that mechanical tension spreading
from the leading edge induces a response in neighbouring
cells — ERK activation in this case — that in turn is
necessary for sheet migration (Figure 2).
Recent papers describe an elegant regulatory mechanism
whereby migration-induced tension influences the status
of the actin cytoskeleton in responding cells via the
activity of the serum response factor (SRF). SRF is a
transcription factor that controls the expression of several
growth-factor-induced genes and is required for cell
motility in several contexts [36–38]. A large body of work,
mainly from Richard Treisman’s laboratory, has shown
that SRF activity is potentiated by situations that stimu-
late actin polymerization, such as Rho GTPase signalling
[39]. The mechanism underlying this regulation
remained obscure until their recent demonstration that
the SRF co-activator MAL moves from the cytoplasm to
the nucleus upon Rho-actin signalling. Once there, it
stimulates the expression of SRF target genes, including
regulators of actin itself. Somogyi and Rorth have recently
described the isolation of mutants in the Drosophilahomologue of MAL (MAL-D, also known as DMRTF),
which show a strong BC migration defect that is appar-
ently due to an inability to organize a robust actin cytos-
keleton [40��]. In this multicellular context, the nuclear
localization of MAL-D appears to be dynamically regu-
lated and shows a clear correlation with cell shape; MAL-
D is observed in the nuclei in a subset of cells in stretched
wild-type clusters whereas it remains cytosolic in all cells
in rounded slbo mutant clusters. Ingenious in vivo ‘pull-
ing’ experiments demonstrate that slbomutants can trans-
locate MAL-D to the nucleus when tugged by motile
wild-type cells, confirming that this is indeed a direct
response to stretching. Given that SRF regulates the
transcription of actin regulators, these data suggest a
positive feedback loop where cytoskeletal changes
induced by pulling forces lead to the nuclear translocation
www.sciencedirect.com
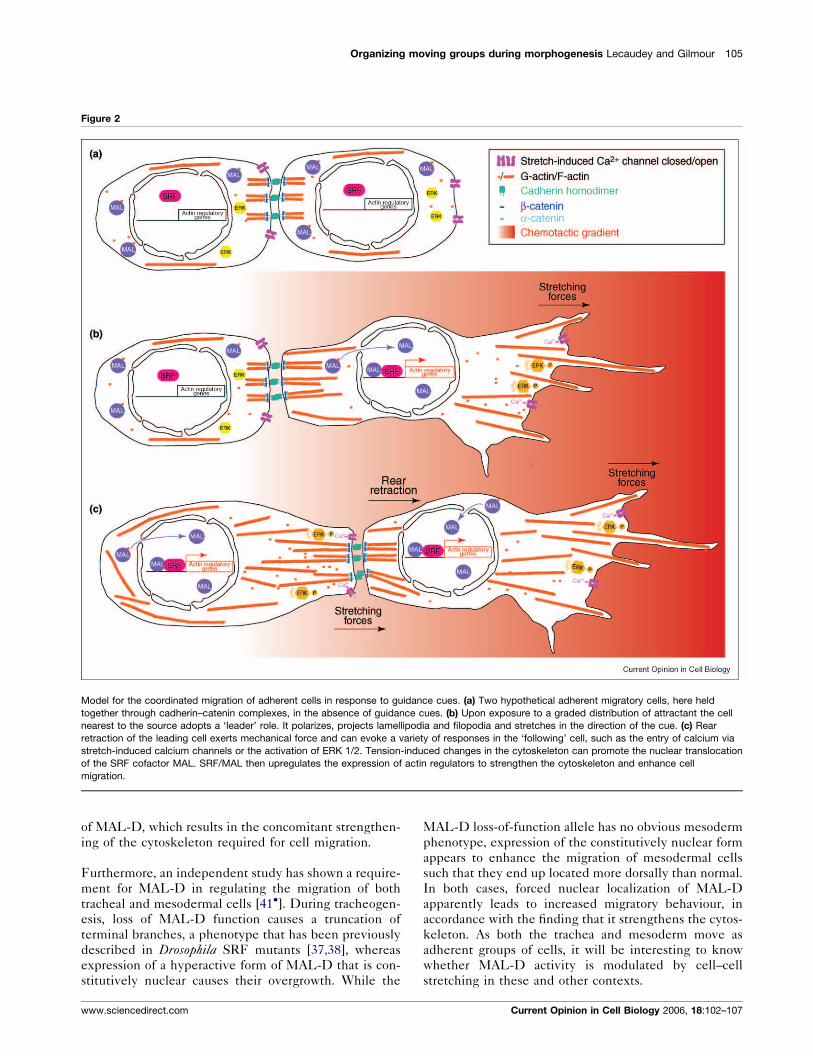
Organizing moving groups during morphogenesis Lecaudey and Gilmour 105
Figure 2
Model for the coordinated migration of adherent cells in response to guidance cues. (a) Two hypothetical adherent migratory cells, here held
together through cadherin–catenin complexes, in the absence of guidance cues. (b) Upon exposure to a graded distribution of attractant the cell
nearest to the source adopts a ‘leader’ role. It polarizes, projects lamellipodia and filopodia and stretches in the direction of the cue. (c) Rear
retraction of the leading cell exerts mechanical force and can evoke a variety of responses in the ‘following’ cell, such as the entry of calcium via
stretch-induced calcium channels or the activation of ERK 1/2. Tension-induced changes in the cytoskeleton can promote the nuclear translocation
of the SRF cofactor MAL. SRF/MAL then upregulates the expression of actin regulators to strengthen the cytoskeleton and enhance cell
migration.
of MAL-D, which results in the concomitant strengthen-
ing of the cytoskeleton required for cell migration.
Furthermore, an independent study has shown a require-
ment for MAL-D in regulating the migration of both
tracheal and mesodermal cells [41�]. During tracheogen-
esis, loss of MAL-D function causes a truncation of
terminal branches, a phenotype that has been previously
described in Drosophila SRF mutants [37,38], whereas
expression of a hyperactive form of MAL-D that is con-
stitutively nuclear causes their overgrowth. While the
www.sciencedirect.com
MAL-D loss-of-function allele has no obvious mesoderm
phenotype, expression of the constitutively nuclear form
appears to enhance the migration of mesodermal cells
such that they end up located more dorsally than normal.
In both cases, forced nuclear localization of MAL-D
apparently leads to increased migratory behaviour, in
accordance with the finding that it strengthens the cytos-
keleton. As both the trachea and mesoderm move as
adherent groups of cells, it will be interesting to know
whether MAL-D activity is modulated by cell–cell
stretching in these and other contexts.
Current Opinion in Cell Biology 2006, 18:102–107

106 Cell structure and dynamics
ConclusionsIt is becoming clear that group migration is regulated by
the combined influence of extrinsic chemical guidance
cues and local mechanical interactions between cells. A
complete understanding of this complex problem will
therefore require a multi-disciplinary approach. Several
recent papers have combined the use of genetics, imaging
and physical ‘tools’ to study the forces involved in
embryonic movements with exciting results. For exam-
ple, atomic force microscopy has been used to measure
the adhesive forces generated by primary zebrafish
embryonic cells plated on cadherin substrates, and these
quantitative findings have been correlated with the cells’
in vivo behaviour [42]. Similarly, a combination of laser
microsurgery and quantitative modelling has been used to
dissect the forces driving morphogenetic movement dur-
ing dorsal closure in Drosophila [43]. On the imaging side,
it will be essential to develop probes that allow the
mechanical activation of biochemical pathways to be
monitored and quantified within living embryos. Once
again, many recent developments show great promise in
this regard, such as FRET probes that can monitor the
dynamics of SRC activation upon mechanical stimulation
[44]. Finally, the identification of biochemical systems
that convert mechanical tension into different types of
migratory behaviour will be a high priority for the future.
AcknowledgementsWe are grateful to Pernille Rorth, Francesca Peri, Petra Haas and StefanEimer for critical reading of the manuscript. We thank Ferenc Jankovics,Damian Brunner, Stefan Luschnig, Arturo Alvarez-Buylla and PernilleRorth for images. V.L. was supported by a fellowship from la Fondationpour la Recherche Medicale.
References and recommended readingPapers of particular interest, published within the annual period ofreview, have been highlighted as:
� of special interest�� of outstanding interest
1.��
Affolter M, Weijer CJ: Signaling to cytoskeletal dynamics duringchemotaxis. Dev Cell 2005, 9:19-34.
A comprehensive review of the signalling pathways controlling cell migra-tion in a wide number of contexts.
2. Friedl P: Prespecification and plasticity: shifting mechanismsof cell migration. Curr Opin Cell Biol 2004, 16:14-23.
3. Murase S, Horwitz AF: Directions in cell migration along therostral migratory stream: the pathway for migration in thebrain. Curr Top Dev Biol 2004, 61:135-152.
4. Doetsch F, Alvarez-Buylla A: Network of tangential pathways forneuronal migration in adult mammalian brain. Proc Natl AcadSci USA 1996, 93:14895-14900.
5.�
Teddy JM, Kulesa PM: In vivo evidence for short- and long-range cell communication in cranial neural crest cells.Development 2004, 131:6141-6151.
Neural crest (NC) cells move in dense streams away from the dorsalhindbrain. Using time-lapse microscopy, it is shown that NC cells withinthese streams make numerous, long-range (up to 100 mm) contactsduring migration. Cell–cell contact may therefore have an importantfunction in ensuring coordinated guidance during this complex process.
6. Schock F, Perrimon N: Molecular mechanisms ofepithelial morphogenesis. Annu Rev Cell Dev Biol 2002,18:463-493.
Current Opinion in Cell Biology 2006, 18:102–107
7. Starz-Gaiano M, Montell DJ: Genes that drive invasion andmigration in Drosophila. Curr Opin Genet Dev 2004, 14:86-91.
8. Rorth P: Initiating and guiding migration: lessons from bordercells. Trends Cell Biol 2002, 12:325-331.
9. Ghysen A, Dambly-Chaudiere C: Development of the zebrafishlateral line. Curr Opin Neurobiol 2004, 14:67-73.
10. Gilmour D, Knaut H, Maischein HM, Nusslein-Volhard C: Towingof sensory axons by their migrating target cells in vivo.Nat Neurosci 2004, 7:491-492.
11. Montero JA, Heisenberg CP: Gastrulation dynamics: cells moveinto focus. Trends Cell Biol 2004, 14:620-627.
12. Keller R: Cell migration during gastrulation. Curr Opin Cell Biol2005, 17:533-541.
13. Dormann D, Weijer CJ: Chemotactic cell movement duringdevelopment. Curr Opin Genet Dev 2003, 13:358-364.
14. Rorth P:Communication by touch: role of cellular extensions incomplex animals. Cell 2003, 112:595-598.
15. Gabay L, Seger R, Shilo BZ: MAP kinase in situ activation atlasduring Drosophila embryogenesis. Development 1997,124:3535-3541.
16. Jekely G, Sung HH, Luque CM, Rorth P: Regulators ofendocytosis maintain localized receptor tyrosine kinasesignalling in guided migration. Dev Cell 2005, 9:197-207.
17. Mine N, Iwamoto R, Mekada E: HB-EGF promotes epithelial cellmigration in eyelid development. Development 2005,132:4317-4326.
18. Fulga TA, Rorth P: Invasive cell migration is initiated by guidedgrowth of long cellular extensions. Nat Cell Biol 2002,4:715-719.
19. Niewiadomska P, Godt D, Tepass U: DE-Cadherin is required forintercellular motility during Drosophila oogenesis. J Cell Biol1999, 144:533-547.
20. Rorth P, Szabo K, Texido G: The level of C/EBP protein is criticalfor cell migration during Drosophila oogenesis and is tightlycontrolled by regulated degradation. Mol Cell 2000, 6:23-30.
21. Dormann D, Weijer G, Dowler S, Weijer CJ: In vivo analysis of3-phosphoinositide dynamics during Dictyosteliumphagocytosis and chemotaxis. J Cell Sci 2004, 117:6497-6509.
22. Dormann D, Weijer CJ: Propagating chemoattractant wavescoordinate periodic cell movement in Dictyostelium slugs.Development 2001, 128:4535-4543.
23. Parent CA: Making all the right moves: chemotaxis inneutrophils andDictyostelium.Curr Opin Cell Biol 2004, 16:4-13.
24. Shelden E, Knecht DA: Mutants lacking myosin II cannot resistforces generated during multicellular morphogenesis.J Cell Sci 1995, 108:1105-1115.
25. Xu XS, Kuspa A, Fuller D, Loomis WF, Knecht DA: Cell–celladhesion prevents mutant cells lacking myosin II frompenetrating aggregation streams of Dictyostelium. Dev Biol1996, 175:218-226.
26. Edwards KA, Kiehart DP: Drosophila nonmuscle myosin II hasmultiple essential roles in imaginal disc and egg chambermorphogenesis. Development 1996, 122:1499-1511.
27. Bershadsky AD, Balaban NQ, Geiger B: Adhesion-dependentcell mechanosensitivity. Annu Rev Cell Dev Biol 2003,19:677-695.
28. Li S, Huang NF, Hsu S: Mechanotransduction in endothelial cellmigration. J Cell Biochem 2005, in press.
29.��
Tzima E, Irani-Tehrani M, Kiosses WB, Dejana E, Schultz DA,Engelhardt B, Cao G, DeLisser H, Schwartz MA: Amechanosensory complex that mediates the endothelial cellresponse to fluid shear stress. Nature 2005, 437:426-431.
As endothelial cells show several coordinated responses to shear stress,such as aligning in the direction of flow, they provide a very nice modelsystem for studying how mechanical force can affect cell behaviour.These authors identify a complex containing three transmembrane
www.sciencedirect.com

Organizing moving groups during morphogenesis Lecaudey and Gilmour 107
proteins that is present at adherens junctions and address the relativecontributions of all three in mediating mechanosensation in this context.
30. Ko KS, Arora PD, McCulloch CA: Cadherins mediateintercellular mechanical signalling in fibroblasts by activationof stretch-sensitive calcium-permeable channels. J Biol Chem2001, 276:35967-35977.
31. Fache S, Dalous J, Engelund M, Hansen C, Chamaraux F,Fourcade B, Satre M, Devreotes P, Bruckert F: Calciummobilization stimulates Dictyostelium discoideum shear-flow-induced cell motility. J Cell Sci 2005, 118:3445-3457.
32. Doyle A, Marganski W, Lee J: Calcium transients inducespatially coordinated increases in traction force during themovement of fish keratocytes. J Cell Sci 2004, 117:2203-2214.
33. Wallingford JB, Ewald AJ, Harland RM, Fraser SE: Calciumsignalling during convergent extension in Xenopus. Curr Biol2001, 11:652-661.
34.�
Farooqui R, Fenteany G: Multiple rows of cells behind anepithelial wound edge extend cryptic lamellipodia tocollectively drive cell-sheet movement. J Cell Sci 2005,118:51-63.
Using a combination of confocal and electron microscopy in a woundclosure model, it is shown that submarginal cells several rows from theleading edge extend cryptic basal lamellipodia under cells in front of themand crawl actively while maintaining apical cell–cell contacts.
35. Matsubayashi Y, Ebisuya M, Honjoh S, Nishida E: ERK activationpropagates in epithelial cell sheets and regulates theirmigration during wound healing. Curr Biol 2004,14:731-735.
36. Alberti S, Krause SM, Kretz O, Philippar U, Lemberger T, CasanovaE, Wiebel FF, Schwarz H, Frotscher M, Schutz G, Nordheim A:Neuronal migration in the murine rostral migratory streamrequires serum response factor. Proc Natl Acad Sci USA 2005,102:6148-6153.
37. Affolter M, Montagne J, Walldorf U, Groppe J, Kloter U, LaRosa M,Gehring WJ: The Drosophila SRF homolog is expressed in asubset of tracheal cells and maps within a genomic regionrequired for tracheal development. Development 1994,120:743-753.
www.sciencedirect.com
38. Guillemin K, Groppe J, Ducker K, Treisman R, Hafen E, Affolter M,Krasnow MA: The pruned gene encodes the Drosophila serumresponse factor and regulates cytoplasmic outgrowth duringterminal branching of the tracheal system. Development 1996,122:1353-1362.
39. Miralles F, Posern G, Zaromytidou AI, Treisman R: Actindynamics control SRF activity by regulation of its coactivatorMAL. Cell 2003, 113:329-342.
40.��
Somogyi K, Rorth P: Evidence for tension-based regulation ofDrosophila MAL and SRF during invasive cell migration.Dev Cell 2004, 7:85-93.
This study uses the experimental strengths of the border cell (BC) modelto address both the function and the regulation of MAL/SRF during cellmigration in vivo. Cell lacking MAL-D show reduced F-actin accumula-tion, which leads to a strong defect in the invasive migration of BCs. BCsmutant for mal-d rupture to produce fragments that are able to migrate ina directional manner, supporting the suggestion that these cells have aweakened cytoskeleton. The authors then exploit the fact that BCs movein a cohesive cluster to show that the nuclear localization of MAL-D isregulated by cell stretching.
41.�
Han Z, Li X, Wu J, Olson EN: A myocardin-related transcriptionfactor regulates activity of serum response factor inDrosophila. Proc Natl Acad Sci USA 2004, 101:12567-12572.
This paper shows that the SRF/MAL-D transcriptional complex isrequired during trachea development and mesodermal migration.Together with the previous paper, these data suggest MAL-D as a verygood candidate to transduce mechanical forces in tissues undergoingextensive cytoplasmic extension or migration.
42. Ulrich F, Krieg M, Schotz EM, Link V, Castanon I, Schnabel V,Taubenberger A, Mueller D, Puech PH, Heisenberg CP: Wnt11Functions in gastrulation by controlling cell cohesion throughRab5c and E-Cadherin. Dev Cell 2005, 9:555-564.
43. Hutson MS, Tokutake Y, Chang MS, Bloor JW, Venakides S,Kiehart DP, Edwards GS: Forces for morphogenesisinvestigated with laser microsurgery and quantitativemodeling. Science 2003, 300:145-149.
44. Wang Y, Botvinick EL, Zhao Y, Berns MW, Usami S, Tsien RY,Chien S: Visualizing the mechanical activation of Src.Nature 2005, 434:1040-1045.
Current Opinion in Cell Biology 2006, 18:102–107




















