
Proteolytic degradation of protein kinase C in the phorbol ester-induced interleukin-2 secreting...
12
ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 267, No. 2, December, pp. 503-514,1988 Proteolytic Degradation of Protein Kinase C in the Phorbol Ester-Induced Interleukin-2 Secreting Thymoma Cells FREESIA L. HUANG,**l PRINCE K. ARORA,? EDGAR E. HANNA,? AND KUO-PING HUANG* *Section on Metabolic Regulation, Endocrinology and Reproduction Research Branch and tSection an Immwwregulation and Cellular Control, Laboratmy of Developmental and Molecular Immunity, National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, Maryland 20892 Received February 18,1988, and in revised form June 1,1988 Effects of phorbol 12-myristate 13-acetate (PMA) on the fate of protein kinase C in two mouse thymoma cell lines, which are either responsive (EL4) or unresponsive (IEL4) to PMA-induced interleukin-2 (IL-2) production, were investigated with polyclonal anti- bodies raised against rat brain enzyme. These antibodies immunoprecipitated com- pletely the protein kinase C from both cell lines and detected mainly an 82-kDa protein by immunoblot analysis of the crude homogenates as well as the partially purified kinase preparations. PMA elicited a time- and dose-dependent redistribution of protein kinase C from cytosol to the particulate fraction and proteolytic degradation of the kinase from both cell lines. The dose of PMA required for half-maximum protein kinase C transloca- tion and degradation was at least five times lower for EL4 than for IEL4. In the presence of 16 nM PMA the rates of protein kinase C translocation and degradation were faster in EL4 than in IEL4, and the half-lives of protein kinase C in EL4 and IEL4 were less than 5 min and greater than 2 h, respectively. Analysis of the tryptic fragments of the immunoprecipitated enzyme, previously phosphorylated in the presence of [-r-32P]ATP, revealed minor structural differences between the protein kinase C from these two cell lines. In neither cell line did the PMA-induced degradation of protein kinase C result in an accumulation of the Ca’+/phospholipid-independent kinase (catalytic unit) as ana- lyzed by immunoblotting and gel filtration chromatography. Thus, activation of protein kinase C through the proteolytic conversion to the effector-independent catalytic unit plays little role in IL-2 production. The role of protein kinase C translocation and degra- dation in the PMA-induced responses in EL4 cells is unknown. However, IL-2 production in EL4 cells was reduced when PMA-induced degradation of protein kinase C was re- tarded by exogenously added protease inhibitors. 0 1988 Academic Press, Inc. Protein kinase C, the Ca2+/phospho- lipid-dependent protein kinase, has been identified as a putative receptor for tumor- promoting phorbol esters (l-4). In viva, this kinase is thought to be activated by di- acylglycerol generated from the signal-in- duced hydrolysis of phosphatidylinositol 4,5-bisphosphate (5, 6). Activation of pro- i To whom correspondence should be addressed at: Building 10, Room 8C407, National Institutes of Health, Bethesda, MD 20892. ’ Abbreviations used: DOG, 1,2-dioleoylglycerol; tein kinase C by either diacylglycerol or tu- mor-promoting phorbol esters has been implicated in the regulation of many cellu- lar functions as well as tumor promotion (7). Protein kinase C is ubiquitously pres- ent in a variety of tissues and cells (8, 9) and its subcellular localization appears to be varied in different cell types. Hormones, growth factors, and tumor-promoting phorbol esters are known to initiate the translocation of protein kinase C from cy- tosol to the particulate fractions (10-14). 503 0003-9861/88 $3.00 Copyright Q 1988 by Academic Press, Inc. All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Proteolytic degradation of protein kinase C in the phorbol ester-induced interleukin-2 secreting...

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 267, No. 2, December, pp. 503-514,1988
Proteolytic Degradation of Protein Kinase C in the Phorbol Ester-Induced Interleukin-2 Secreting Thymoma Cells
FREESIA L. HUANG,**l PRINCE K. ARORA,? EDGAR E. HANNA,? AND KUO-PING HUANG*
*Section on Metabolic Regulation, Endocrinology and Reproduction Research Branch and tSection an Immwwregulation and Cellular Control, Laboratmy of Developmental and Molecular Immunity, National
Institute of Child Health and Human Development, National Institutes of Health, Bethesda, Maryland 20892
Received February 18,1988, and in revised form June 1,1988
Effects of phorbol 12-myristate 13-acetate (PMA) on the fate of protein kinase C in two mouse thymoma cell lines, which are either responsive (EL4) or unresponsive (IEL4) to PMA-induced interleukin-2 (IL-2) production, were investigated with polyclonal anti- bodies raised against rat brain enzyme. These antibodies immunoprecipitated com- pletely the protein kinase C from both cell lines and detected mainly an 82-kDa protein by immunoblot analysis of the crude homogenates as well as the partially purified kinase preparations. PMA elicited a time- and dose-dependent redistribution of protein kinase C from cytosol to the particulate fraction and proteolytic degradation of the kinase from both cell lines. The dose of PMA required for half-maximum protein kinase C transloca- tion and degradation was at least five times lower for EL4 than for IEL4. In the presence of 16 nM PMA the rates of protein kinase C translocation and degradation were faster in EL4 than in IEL4, and the half-lives of protein kinase C in EL4 and IEL4 were less than 5 min and greater than 2 h, respectively. Analysis of the tryptic fragments of the immunoprecipitated enzyme, previously phosphorylated in the presence of [-r-32P]ATP, revealed minor structural differences between the protein kinase C from these two cell lines. In neither cell line did the PMA-induced degradation of protein kinase C result in an accumulation of the Ca’+/phospholipid-independent kinase (catalytic unit) as ana- lyzed by immunoblotting and gel filtration chromatography. Thus, activation of protein kinase C through the proteolytic conversion to the effector-independent catalytic unit plays little role in IL-2 production. The role of protein kinase C translocation and degra- dation in the PMA-induced responses in EL4 cells is unknown. However, IL-2 production in EL4 cells was reduced when PMA-induced degradation of protein kinase C was re- tarded by exogenously added protease inhibitors. 0 1988 Academic Press, Inc.
Protein kinase C, the Ca2+/phospho- lipid-dependent protein kinase, has been identified as a putative receptor for tumor- promoting phorbol esters (l-4). In viva, this kinase is thought to be activated by di- acylglycerol generated from the signal-in- duced hydrolysis of phosphatidylinositol 4,5-bisphosphate (5, 6). Activation of pro-
i To whom correspondence should be addressed at: Building 10, Room 8C407, National Institutes of Health, Bethesda, MD 20892.
’ Abbreviations used: DOG, 1,2-dioleoylglycerol;
tein kinase C by either diacylglycerol or tu- mor-promoting phorbol esters has been implicated in the regulation of many cellu- lar functions as well as tumor promotion (7). Protein kinase C is ubiquitously pres- ent in a variety of tissues and cells (8, 9) and its subcellular localization appears to be varied in different cell types. Hormones, growth factors, and tumor-promoting phorbol esters are known to initiate the translocation of protein kinase C from cy- tosol to the particulate fractions (10-14).
503 0003-9861/88 $3.00 Copyright Q 1988 by Academic Press, Inc. All rights of reproduction in any form reserved.

504 HUANG ET AL.
This ligand-induced redistribution of pro- tein kinase C is generally considered to be an early event necessary in triggering the subsequent cellular responses.
Prolonged exposure of cells to phorbol ester results in a reduction of the binding sites for this ligand and the Ca2+/phospho- lipid-dependent protein kinase activity. This down-regulation has been observed in a variety of cells including mouse and rat keratinocytes (15-17), Swiss 3T3 cells (18), Friend erythroleukemia cells (19), human promyelocytic leukemia cells HL-60 (20), human mammary carcinoma cells (21), rat pituitary tumor cells GH3 (22), and human platelets and neutrophils (23-25). The loss of activity is considered to be due to the proteolytic degradation of the enzyme rather than to an inhibitory mechanism (21-25). In vitro, proteolytic degradation of protein kinase C by Ca2+-dependent neu- tral proteases or trypsin generates a Ca2+/ phospholipid-independent protein kinase, designated M kinase (26-28). Recent stud- ies further demonstrated that tryptic di- gestion of protein kinase C generates not only a kinase fragment of 45-50 kDa but also a phorbol ester-binding fragment of 32-38 kDa (29-31). The resulting phorbol ester-binding fragment has a higher affinity for phosphatidylserine (PS)’ and no longer requires Ca2+ for the binding of phorbol ester (29). However, in intact cells, there is no conclusive evidence presently demonstrating that PMA-mediated trans- location and degradation of the enzyme re- sults in the accumulation of these two functionally competent fragments of pro- tein kinase C.
In order to better define the mechanism of action of tumor-promoting phorbol es- ters and the role of protein kinase C in the regulation of phorbol ester-induced cellu- lar events, we have chosen EL4 mouse thy- moma cells as our model. It has been
a Abbreviations used: DOG, 1,2-dioleoylglycerol; PS, phosphatidylserine; PMA, phorbol 12-myristate 13-acetate; IL-2, interleukin-2; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; PMSF, phenylmethylsulfonyl fluoride; NP-40, Non- idet-P40; EGTA, ethylene glycol bis(P-aminoethyl ether)iV,N’-tetraacetic acid; DTT, dithiothreitol.
shown that PMA causes a loss of cytosolic protein kinase C (10) and secretion of in- terleukin-2 (IL-2) (32) in EL4 cells. It is not clear how protein kinase C is involved in the control of PMA-induced IL-2 secretion, since a second line of thymoma cell, IEL4, which displays similar phorbol ester bind- ing site and PMA-induced reduction of cy- tosolic protein kinase C, does not produce IL-2 (33). So far, the fate of protein kinase C in these two cell lines has not been as- sessed by methods other than measure- ments of the kinase activity and binding of phorbol ester. In addition, the PMA-medi- ated down-regulation of protein kinase C has not been shown in these cells. In this report we have employed polyclonal anti- bodies against rat brain protein kinase C (34) to investigate the effect of PMA on the fate of this kinase. These antibodies are known to recognize all three types of rat and monkey brain protein kinase C iso- zymes and cross-react with the same en- zyme from other species. By using the im- munochemical method we have confirmed the previous findings on the translocation of protein kinase C and demonstrated different PMA dose responses in EL4 and IEL4 cells for protein kinase C transloca- tion and degradation. The greater suscep- tibility of the protein kinase C in EL4 than that in IEL4 to proteolytic degradation may result from the structural differences of the enzymes in these cell lines. The PMA-induced degradation of protein ki- nase C in either cell line, however, does not lead to the accumulation of a Ca’+/phos- pholipid - independent kinase. Further- more, blockage of protein kinase C degra- dation by protease inhibitors resulted in the retardation of IL-2 production in EL4.
EXPERIMENTAL PROCEDURES
Materials. The following materials were obtained from the indicated sources: histone 111s and PMA from Sigma; [Y-~P]ATP, [‘Hlphorbol 12,13-dibuty- rate, [‘?]protein A, and [‘Hlthymidine from New En- gland Nuclear; Pansorbin from Behring Diagnostics; PS and 1,2-dioleoylglycerol (DOG) from Avanti Polar Lipids; TPCK-treated trypsin from Worthington; polyacrylamide gel electrophoresis, immunoblot, and protein determination reagents from Bio-Rad; affin- ity-purified rabbit anti-goat IgG from Cooper Bio-

PROTEOLYTIC DEGRADATION OF PROTEIN KINASE C 505
chemicals; RPM1 1640 medium from GIBCO; and fetal bovine serum from Hyclone.
Cell culture and preparation of cell extracts. EL4 and IEL4 were maintained in RPM1 1640 medium supple- mented with 2 mM glutamine, 100 U/ml of penicillin, 100 pg/ml of streptomycin, and 10% heat-inactivated fetal bovine serum. They were grown as suspension culture for 4 to 7 days before use. Prior to PMA treat- ment, cells were collected and washed twice with fresh medium containing 1% fetal bovine serum and suspended in the same medium at 1 X lo6 cells/ml. Routinely 30 X lo6 cells in 30 ml medium in duplicate were used for each testing. PMA stock solution was diluted with ethanol and added to the cell culture to give a final ethanol concentration of 0.1%. After PMA treatment, cells were quickly chilled in ice and diluted with ice-cold 10 mM Tris-Cl buffer, pH 7.5, containing 150 mM NaCl before centrifugation. Collected cells were washed three times with the same buffered sa- line and the cell pellets were kept frozen at -70°C be- fore processing. Cells were homogenized in Kontes microhomogenizer by hand in 20 mM Tris-Cl buffer, pH 7.5, containing 5 mM EGTA, 2 mM EDTA, 1 mM
DTT, 10% glycerol, 0.5 mM PMSF, and 2 pg/ml leu- peptin (Buffer A). The homogenates were centrifuged at 100,OOOg for 45 min; the supernatant fluid and re- sulting pellet were designated as cytosolic and partie- ulate fractions, respectively. Protein kinase C associ- ated with the particulate fraction was extracted by sonication in Buffer A containing 0.5% NP-40 and fol- lowed by centrifugation at 100,OOOg for 45 min. Whole cell extracts were obtained by sonication of the cells in Buffer A containing 0.5% NP-40 and centrifugation at 100,OOOg for 45 min. The high-speed pellets from the detergent-extracted cells contained negligible amount of protein kinase C as analyzed by immu- noblot.
Partial purification and assay of protein kinose C. The cell extracts derived from 8-10 X 10’ cells were applied to a DEAE-cellulose column (1.5 X 2.5 cm) equilibrated with 20 mM Tris-Cl buffer, pH 7.5, con- taining 0.5 mM EDTA, 0.5 mM EGTA, 1 mM DTT, 10% glycerol, and 0.5 mM PMSF (Buffer B). The column was washed with 30 ml of Buffer B and protein kinase C was eluted from the column with a linear O-O.3 M
KC1 gradient in Buffer B. The enzyme was eluted at a KC1 concentration of approximately 0.08 M. Protein kinase C activity was measured in 30 mM Tris-Cl buffer, pH 7.5, containing 6 mM magnesium acetate, 0.12 mM [y-32P]ATP, 0.4 mM CaClz, 40 pg/ml PS, 8 pg/ ml DOG, 1 mg/ml histone IIIS, and protein kinase. The Ca*+/phospholipid-independent activity was measured under the same conditions without Ca2+ and phospholipid but containing 1.5 mM EGTA.
Immunoprecipitation and immunoblot analysis. Partially purified protein kinase C in Buffer B was in- cubated in a total volume of 30 ~1 with increasing amounts of polyclonal goat anti-rat brain protein ki-
nase C antiserum (34) or preimmune serum; both had previously been dialyzed in Buffer B, at room temper- ature for 30 min. The antigen-antibody complex was removed from the solution after further incubation with Pansorbin (25 ~1 of 35% suspension) at 4°C for 30 min. Aliquots from the resulting supernatant fluid were taken for the measurement of protein kinase C activity. Immunoblot analysis was carried out ac- cording to Towbin et al (35). Polypeptides in each sample were separated by SDS-PAGE (10% gel) fol- lowed by electrophoretic transfer to nitrocellulose membrane. The membrane was incubated succes- sively with 3% gelatin in TBS (20 mM Tris-Cl buffer, pH 7.5, containing 500 mM NaCI) for 30 min, with an- tiserum or preimmune serum (lOOO-fold dilution) in TBS containing 1% gelatin for 90 min, with affinity- purified rabbit anti-goat IgG (2000-fold dilution) for 1 h, and with [‘?lprotein A (0.4 fig, 4-6 &i/20 ml) for 1 h. Following each incubation the membrane was washed extensively with TBS containing 0.05% Tween 20. The immunoreactive bands were visualized by autoradiography. The positions of molecular weight marker proteins were localized by brief stain- ing with 0.1% Amido black in 25% isopropanol con- taining 10% acetic acid.
Peptide mapping analysis. Partially purified pro- tein kinase C of EL4 and IEL4 was autophosphory- lated in the standard protein kinase C assay mixture without histone but containing 0.06 mM [y-32P]ATP. The labeled protein kinase C was isolated by immuno- precipitation using the polyclonal antibodies and sep- arated by SDS-PAGE. The major labeled protein band (82 kDa) was excised from the gel, cut up, treated with acetone/HzO/triethylamine/acetic acid (17/1/1/l, vol) and digested with TPCK-treated tryp- sin in 50 mM NH,HCO,, pH 7.8. The digest was lyophi- lized and analyzed by thin-layer electrophoresis at 1000 V for 30-40 min at pH 6.5 (pyridinelacetie acid/ water, 25/l/225) to allow the tracking dye to migrate 6 cm toward the anode and followed by chromatogra- phy in the second dimension with freshly prepared n-butanol/acetic acid/water (41115). The 32P-labeled peptides were visualized by autoradiography.
Biological assays for IL.2 The culture supernatant was assayed for T cell growth activity as described (32) using an IL-e-dependent mouse CTLL line. CTLL cells (2 X lo* cells/well) were cultured in 96-well flat bottom microtiter plates containing 0.2 ml of medium (RPM1 1640 plus 5% fetal bovine serum) and serially diluted EL4 or IEL4 culture media for 24 h. During the final 6-8 h of incubation the cells were pulsed with [3H]thymidine (1 &i/well). The cells were harvested with a MASH II cell harvester and [3H]thymidine up- take was determined in a liquid scintillation counter.
Other metho&. Sodium dodecyl sulfate-polyacryl- amide gel electrophoresis was carried out according to the method of Laemmli (36) and protein concentra- tion was determined by the dye-binding method of
506 HUANG ET AL.
0 0 2 4 6 8
B
- 45K
s( - 31K
1 2
Immune Serum ( ul )
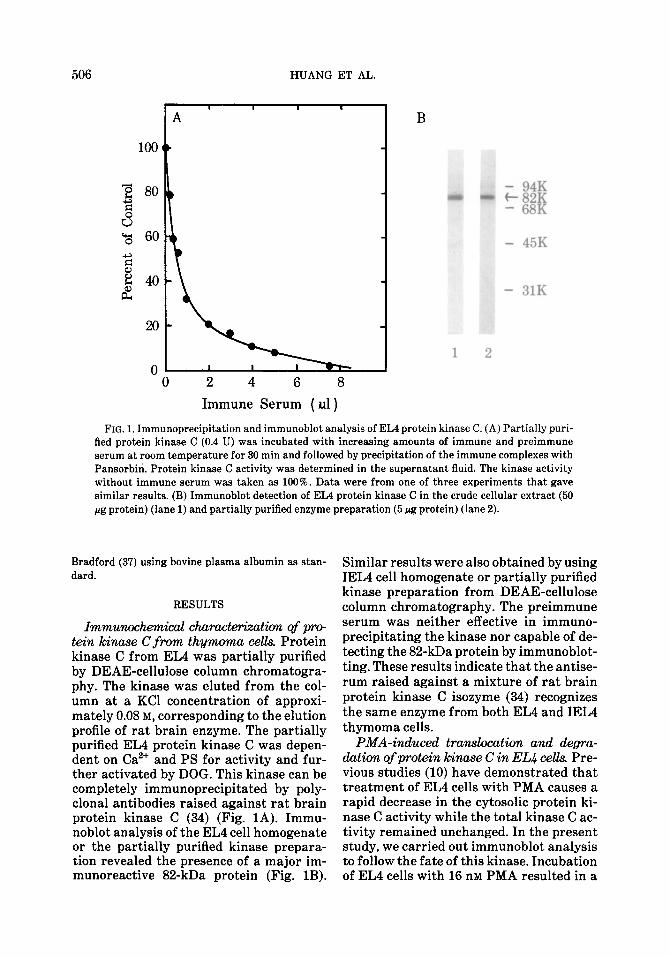
FIG. 1. Immunoprecipitation and immunoblot analysis of EL4 protein kinase C. (A) Partially puri- fied protein kinase C (0.4 U) was incubated with increasing amounts of immune and preimmune serum at room temperature for 30 min and followed by precipitation of the immune complexes with Pansorbin. Protein kinase C activity was determined in the supernatant fluid. The kinase activity without immune serum was taken as 100%. Data were from one of three experiments that gave similar results. (B) Immunoblot detection of EL4 protein kinase C in the crude cellular extract (50 pg protein) (lane 1) and partially purified enzyme preparation (5 pg protein) (lane 2).
Bradford (37) using bovine plasma albumin as stan- dard.
RESULTS
Immunochemical churacterization of pro- tein kinase C frm thymoma cells. Protein kinase C from EL4 was partially purified by DEAE-cellulose column chromatogra- phy. The kinase was eluted from the col- umn at a KC1 concentration of approxi- mately 0.08 M, corresponding to the elution profile of rat brain enzyme. The partially purified EL4 protein kinase C was depen- dent on Ca2+ and PS for activity and fur- ther activated by DOG. This kinase can be completely immunoprecipitated by poly- clonal antibodies raised against rat brain protein kinase C (34) (Fig. 1A). Immu- noblot analysis of the EL4 cell homogenate or the partially purified kinase prepara- tion revealed the presence of a major im- munoreactive 82-kDa protein (Fig. 1B).
Similar results were also obtained by using IEL4 cell homogenate or partially purified kinase preparation from DEAE-cellulose column chromatography. The preimmune serum was neither effective in immuno- precipitating the kinase nor capable of de- tecting the 82-kDa protein by immunoblot- ting. These results indicate that the antise- rum raised against a mixture of rat brain protein kinase C isozyme (34) recognizes the same enzyme from both EL4 and IEL4 thymoma cells.
PMA-induced translocation and degra- dation of protein kinase C in EL4 cells. Pre- vious studies (10) have demonstrated that treatment of EL4 cells with PMA causes a rapid decrease in the cytosolic protein ki- nase C activity while the total kinase C ac- tivity remained unchanged. In the present study, we carried out immunoblot analysis to follow the fate of this kinase. Incubation of EL4 cells with 16 nM PMA resulted in a

PROTEOLYTIC DEGRADATION OF PROTEIN KINASE C 507
94K 82K 68K
45K
31K
+ -
0 2 5 10 30 2 24 0 2 5 10 30 2 24 min hr min hr
FIG. 2. Effect of PMA treatment on the redistribution of EL4 protein kinase C. EL4 cells (30 x 106/ 30 ml) were incubated with 16 nM PMA for the period of time indicated. The cell homogenates were separated into cytosolic and particulate fractions by centrifugation at 100,OOOg for 45 min. Equiva- lent amounts of protein (50 pg) from eytosolic (A) and particulate (B) fractions were analyzed by immunoblotting. Roughly two amounts of cytosolic protein versus one amount of particulate protein were obtained from a given cell homogenate; thus, for quantitation of total enzyme, it was necessary to sum up twice the cytosolic protein kinase C and that of the particulate fraction. Data were from one of four experiments that gave similar results.
rapid loss of protein kinase C molecule from cytosol (Fig. ZA). Concomitant with the loss of cytosolic protein kinase C was an initial increase and followed by a grad- ual decrease in the particulate fraction-as- sociated kinase molecule (Fig. 2B). Before the addition of PMA, over 90% of protein kinase C was cytosolic. The major immu- noreactive protein is the 82-kDa native protein kinase C. We have also detected a minor 45-kDa protein, which appears to be insensitive to the PMA-induced transloca- tion. The molecular weight of this latter protein is similar to that of the proteolytic fragment of rat brain protein kinase C (29); however, identification of this protein has yet to be carried out. Five minutes af- ter the addition of PMA, the 82-kDa pro- tein in the cytosol was reduced by more than 90%, whereas that in the particulate fraction increased to a maximal level. As incubation continued, the particulate frac- tion-associated kinase was also gradually reduced. In fact, degradation occurred si- multaneously as translocation took place, since at 2 min (or even earlier) not all the
protein kinase C that disappeared from cy- tosol could be accounted for in the particu- late fraction. These results demonstrate that PMA causes both the translocation and degradation of protein kinase C. How- ever, the PMA-induced degradation of EL4 protein kinase C does not generate an addi- tional smaller molecular weight immuno- reactive protein in either cytosol or the particulate fraction.
E#ect of PMA on the fate of protein ki- nase C in IEL.4. Previous studies have dem- onstrated that both PMA-responsive EL4 and -unresponsive IEL4 thymoma cells have similar numbers of phorbol ester binding sites with comparable binding affinities (33). Incubation of IEL4 cells with 16 nM PMA also resulted in a translo- cation of protein kinase C from cytosol to the particulate fraction (Fig. 3). However, the rate of protein kinase C loss from the cytosolic fraction of IEL4 (Fig. 3A) was much slower than that observed for EL4 (Fig. 2). Furthermore, the translocated ki- nase seemed to accumulate in the particu- late fraction and was degraded only slowly

508 HUANG ET AL.
-45
- 31
0 2 5 10 30 1 2 6 24 0 2 5 10 30 1 2 6 24 min hr min hr
FIG. 3. PMA-induced redistribution of protein kinase C in IEL4. IEL4 cells (30 X 106/30 ml) were incubated with 16 nM PMA for the period of time indicated. The cell homogenates were separated into cytosolic and particulate fractions. Equivalent amount of protein (50 pg) from cytosolic (A) and particulate (B) fractions were analyzed by immunoblot analysis. For quantitation of total enzyme, see the legend to Fig. 2. Data were from one of three experiments that gave similar results.
(Fig. 3B). The rate of degradation must be very slow as compared to that in EL4. At 2 min, the enzyme accumulated in particu- late fraction of IEL4 was more than that in EL4 in spite of its retarded rate of disap- pearance from cytosol of IEL4. After 30 min of incubation with PMA, over 90% of the translocated kinase still retained in the particulate fraction from several ex- periments. For better quantitation, total protein kinase C contents in the whole cell extract were analyzed (Fig. 4). Significant degradation of the kinase in IEL4 was ob- served only after 2 h of incubation with PMA. The half-life of protein kinase C in the presence of 16 nM PMA was approxi- mately 2-3 h for IEL4 as compared to ap- proximately 5 min for EL4.
Dose response of PMA-induced protein kinase C redistribution. The PMA-induced loss of protein kinase C from the cytosol is dose dependent (Fig. 5), and EL4 is at least five times more sensitive than IEL4. As lit- tle as 0.25-0.5 rig/ml (0.4-0.8 nM) of PMA was sufficient to cause a 50% loss of the ki- nase from cytosol of EL4 in 5 min, whereas at 10 rig/ml, the inactive tumor-promoting phorbol ester, 4-a-phorbol, was not effec- tive. As for IEL4, it required more than 2 rig/ml (3.2 nM) of PMA to produce a 50%
loss of cytosolic kinase C in 5 min. The half-maximal effective dose for either EL4 or IEL4 is less than the previously re- ported value of 4-5 nM for EL4 by measur- ing the changes in the cytosolic protein ki- nase C activity (10). These differences were
80 z * ;
6 60
% .
2 40
8 $ 20
;-I:
Time of Treatment with PMA ( min)
FIG. 4. Comparison of the PMA-induced degrada- tion of protein kinase C from EL4 and IEL4. Whole cell extracts (50 pg protein) from PMA (16 nM)-
treated EL4 (0) and IEL4 (0) were analyzed by im- munoblotting to determine the total protein kinase C in these cells. The amount of immunoreactive 82-kDa enzyme was determined by densitometric measure- ment of the autoradiograph and that at zero time is taken as 100%. Data were from one of three experi- ments that gave similar results.

PROTEOLYTIC DEGRADATION OF PROTEIN KINASE C 509
A. 1 2 3 4 5 6 7 8 9 10 11 12 B. 1 2 3 4 5 6 7
ngiml PMA ngiml PMA
20
0 -10 -9 -8 -7
Log [ PMA (MI]
FIG. 5. Dose response of the PMA-induced loss of protein kinase C from the cytosolic fraction of EL4 and IEL4 cells. EL4 or IEL4 cells (30 X 106/30 ml) were incubated with different concentrations of PMA for 5 min. The cytosolic fractions (each containing 50 yg of protein) were analyzed by immu- noblotting. (A) EL4; (B) IELI. Lane 1 of (A) was incubated with 10 rig/ml 4-a-phorbol. (C) The dose- response curve was determined after densitometric measurement of the autoradiogram. The amount of protein kinase C in cells incubated without PMA was taken as 100%. EL4 (0); IEL4 (0). The experiments were performed three times for EL4 and twice for IEL4, and all give similar results.
probably due to different experimental conditions, as a relatively low cell density (1 X lo6 cells/ml) was used in the present study.
Chromatographic identification of the PMA-induced proteolyzed protein kinase C Incubation of either EL4 or IEL4 with PMA resulted in the degradation of pro-
tein kinase C from these cells. However, we did not detect an accumulation of any im- munoreactive proteolytic fragment of the kinase in either cell line. The polyclonal antibodies employed recognized the tryp- tic fragments of the kinase from rat brain (29) as well as from EL4 and IEL4 (data not shown). In order to determine the deg-
510 HUANG ET AL.
f 120 P
5 80 f
40
n "0 10 20 300 10 20 30
Fraction Number
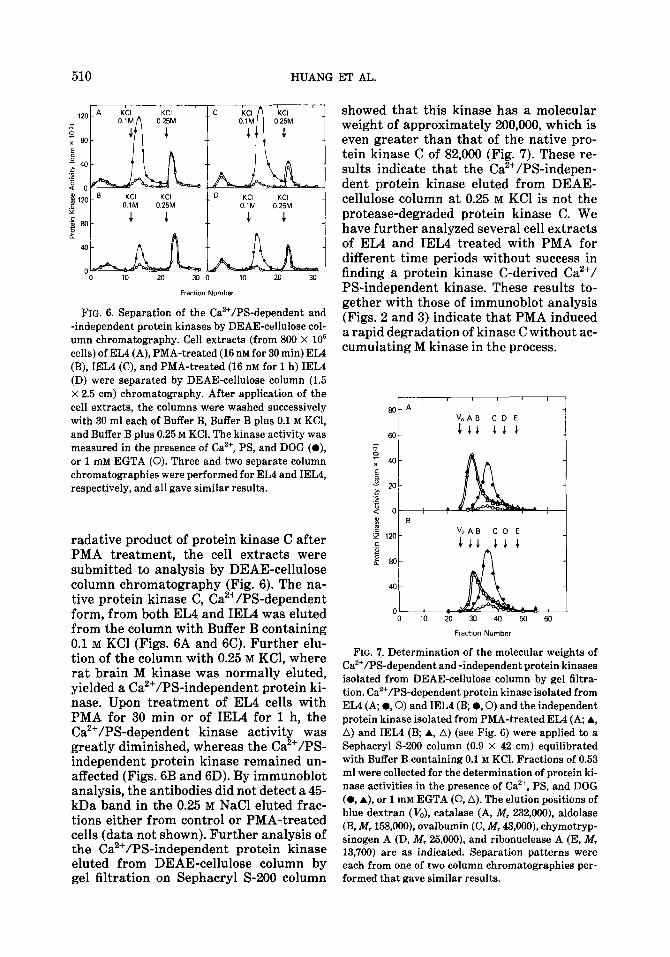
FIG. 6. Separation of the Ca*+/PS-dependent and -independent protein kinases by DEAE-cellulose col- umn chromatography. Cell extracts (from 800 X lo6 cells) of EL4 (A), PMA-treated (16 nM for 30 min) EL4 (B), IEL4 (C), and PMA-treated (16 nM for 1 h) IEL4 (D) were separated by DEAE-cellulose column (1.5 X 2.5 cm) chromatography. After application of the cell extracts, the columns were washed successively with 30 ml each of Buffer B, Buffer B plus 0.1 M KCI, and Buffer B plus 0.25 M KCI. The kinase activity was measured in the presence of Cazf, PS, and DOG (a), or 1 mM EGTA (0). Three and two separate column chromatographies were performed for EL4 and IELI, respectively, and all gave similar results.
radative product of protein kinase C after PMA treatment, the cell extracts were submitted to analysis by DEAE-cellulose column chromatography (Fig. 6). The na- tive protein kinase C, Ca2+/PS-dependent form, from both EL4 and IEL4 was eluted from the column with Buffer B containing 0.1 M KC1 (Figs. 6A and 6C). Further elu- tion of the column with 0.25 M KCl, where rat brain M kinase was normally eluted, yielded a Ca2+/PS-independent protein ki- nase. Upon treatment of EL4 cells with PMA for 30 min or of IEL4 for 1 h, the Ca2+/PS-dependent kinase activity was greatly diminished, whereas the Ca2+/PS- independent protein kinase remained un- affected (Figs. 6B and 6D). By immunoblot analysis, the antibodies did not detect a 45- kDa band in the 0.25 M NaCl eluted frac- tions either from control or PMA-treated cells (data not shown). Further analysis of the Ca2+/PS-independent protein kinase eluted from DEAE-cellulose column by gel filtration on Sephacryl S-200 column
showed that this kinase has a molecular weight of approximately 200,000, which is even greater than that of the native pro- tein kinase C of 82,000 (Fig. 7). These re- sults indicate that the Ca’+/PS-indepen- dent protein kinase eluted from DEAE- cellulose column at 0.25 M KC1 is not the protease-degraded protein kinase C. We have further analyzed several cell extracts of EL4 and IEL4 treated with PMA for different time periods without success in finding a protein kinase C-derived Ca2+/ PS-independent kinase. These results to- gether with those of immunoblot analysis (Figs. 2 and 3) indicate that PMA induced a rapid degradation of kinase C without ac- cumulating M kinase in the process.
I
80 A vo A B CD E
Fraction Number
FIG. ‘7. Determination of the molecular weights of Ca’+/PS-dependent and -independent protein kinases isolated from DEAE-cellulose column by gel filtra- tion. Ca’+/PS-dependent protein kinase isolated from EL4 (A, l , 0) and IEL4 (B; l ,O) and the independent protein kinase isolated from PMA-treated EL4 (A, A, A) and IEL4 (B; A, A) (see Fig. 6) were applied to a Sephacryl S-200 column (0.9 X 42 cm) equilibrated with Buffer B containing 0.1 M KCI. Fractions of 0.53 ml were collected for the determination of protein ki- nase activities in the presence of Ca*+, PS, and DOG (0, A), or 1 mM EGTA (0, A). The elution positions of blue dextran (V,), catalase (A, Al, 232,000), aldolase (B, df, 158,000). ovalbumin (C, Af, 43,000), chymotryp- sinogen A (D, ikf, 25,000), and ribonuclease A (E, M, 13,700) are as indicated. Separation patterns were each from one of two column chromatographies per- formed that gave similar results.

PROTEOLYTIC DEGRADATION OF PROTEIN KINASE C 511
-E 0
FIG. 8. Phosphopeptide mapping analysis of protein kinase C from EL4 and IEL4 cells. Partially purified protein kinase C (after DEAE-cellulose column chro- matography) from EL4 (A) and IEL4 (B) was auto- phosphorylated, immunoprecipitated, and separated on SDS-PAGE. The labeled protein kinase C was ex- cised from gel, digested with trypsin, and analyzed by two-dimensional peptide mapping as described in the text. Arrows on each map denote the spots that are missing from the other map. Missing spots from both maps can be located on a map of the mixed tryptic digest (data not shown). Arrowheads denote the two spots which are prominent in map B but not in map A. Three mapping analyses were performed and all gave similar results.
Peptide mapping of the autophosphory- lated protein kinase C from EL4 and IEL.4. The cause of the differential response of EL4 and IEL4 cells to PMA-induced pro- tein kinase C degradation is unknown. We tested the possibility that protein kinase C from these two cell lines may be structur- ally different. Analysis of the 32P-labeled tryptic peptides derived from the auto- phosphorylated and immunoprecipitated protein kinase C of EL4 and IEL4 showed different separation patterns (Fig. 8). While some peptides were common in both maps, there were at least four minor pep- tides that appeared in the map of EL4 (in-
dicated with arrow) that were not found in the map of IEL4, and two minor peptides in the map of IEL4 (arrow) not in the map of EL4. There were also two prominent peptides in IEL4 (arrowhead) that were relatively less significant in EL4. The differences in the phosphopeptide maps of EL4 and IEL4 were not results of incom- plete tryptic digestion, since more exten- sive digestion with additional trypsin did not alter the number or relative intensity of their phosphopeptides. We interpret the observed differences in the tryptic frag- mentation as a result of structural differ- ences between the kinases of these two cells.
Degradation of protein kinme C and IL2 secretion Since we did not detect the pro- teolyzed Ca’+/ phospholipid - independent protein kinase C fragment in either cell line, we decided to determine the relation- ship of protein kinase C degradation with the subsequent IL-Z secretion. Protease in- hibitors, PMSF and leupeptin, when added simultaneously with PMA, blocked appre- ciably the degradation and subsequently also slowed down the continual transloca- tion of protein kinase C in EL4 as com- pared to cells treated with PMA alone for 30 min (Fig. 9). Such blockage interferred with the IL-Z production analyzed between 5 and 24 h. On the other hand, IEL4, when treated with higher level of PMA (80 or 160 nM), which caused considerable amount of protein kinase C degradation in 30 min (data not shown), did not secrete IL-2 even after 40 h of incubation.
DISCUSSION
Previous studies of Kraft et al. (10) dem- onstrated that PMA induced a rapid loss of protein kinase C activity from the cytosolic fraction of EL4 cells. In subsequent studies using parietal yolk sac (PYS-2) cells these authors had established that the PMA-in- duced loss of cytosolic protein kinase C could be recovered in the particulate frac- tion, in particular the plasma membrane. This redistribution of protein kinase C from cytosol to the particulate fraction has since been observed in several cell types by treatment of PMA as well as by different

512 HUANG
20
w b - 15 X
E
,; 10 .z
2 9 -I - 5
0
x--x---y \ 123456
0 2 4 6 8 10 12
Dilution (2”)
FIG. 9. Effect of protease inhibitors on PMA-in- duced kinase degradation and IL-2 production. EL4 cells (5 X lo6 cells/ml) were treated with 16 nM PMA alone (0, A, and Cl) or PMA with 10 pg/ml leupeptin and 1 mM PMSF (0, A, V, and n ). At 5 h (II, w), 10 h (V), 12 h (A), 16 h (A), and 24 h (0, a), aliquots of cell cultures were withdrawn and centrifuged. Superna- tants were serially diluted and analyzed for IL-2 ac- tivity as described under Experimental Procedures using partially purified rat IL-2 (100 U/ml) as stan- dard (X). After each withdrawal of sample (5,10, and 16 h), the cultures containing protease inhibitors were replenished with leupeptin, 2 pg/ml, and PMSF, 0.2 mM. No IL-2 activity was found in culture superna- tants of untreated cells at 24 h or of IELC treated with either 80 or 160 nM PMA for 24 and 40 h (---). The inset shows the immunoblot analysis of the cytosolic (odd lanes) and particulate (even lanes) protein ki- nase C of the untreated cells (lanes 1 and 2), PMA- and protease inhibitor-treated cells (lanes 3 and 4), and PMA-treated cells (lanes 5 and 6) at 30 min. Quantitation of protein kinase C in the various cell extracts was done as described in the legend to Fig. 2. Data were from one of two experiments performed that gave similar results.
growth factors and hormones (10-14). The ligand-induced translocation of protein ki- nase C was assessed by the changes in the kinase activity, and the total kinase activ- ity reportedly remained constant during incubation with PMA. Recently, Ballester and Rosen (22) employed polyclonal anti- bodies against rat brain protein kinase C
ET AL.
to investigate the translocation of this ki- nase in rat pituitary tumor cells GHa in the presence of PMA and found an accelerated rate of degradation. The fate of protein ki- nase C following degradation, however, could not be monitored because their anti- bodies failed to detect the proteolytic frag- ments of the kinase (22). In the current study we have employed the goat-anti rat brain protein kinase C antibodies (34), which can detect the native kinase as well as the catalytic and phorbol ester binding fragments of the kinase (34), to investigate the fate of protein kinase C in EL4 and IEL4 thymoma cells in response to PMA. Though the antibodies were raised against the rat brain protein kinase C, it immuno- precipitates completely the same enzyme from both thymoma cell lines. It seemed that the immunoreactive determinants of protein kinase C from neuronal and lym- phocytic origins are highly conserved.
Using immunoblot analysis we have pro- vided proof that the PMA-induced loss of cytosolic protein kinase C from EL4 and IEL4 cells is a result of translocation of the enzyme molecule to the particulate frac- tion rather than the generation of inhibi- tor of the kinase. The PMA dose required for half-maximal loss of cytosolic protein kinase C in EL4 cells is lower than that in IEL4 cells. Since both cell lines possess a similar number of phorbol ester-binding sites with similar binding affinity (33), the differences in the rate of protein kinase C translocation from cytosol could be a con- sequence of differential rates of degrada- tion of the membrane-associated kinase. These differential rates of degradation may in part be due to the differences in the primary structure of protein kinase C in EL4 and IEL4 cells. In light of the discov- ery of the various PKC isoforms (39), it is tempting to speculate that the structural differences may due to the presence of different isozymes in these cells. The poly- clonal antibodies employed in the present study recognized all three isozymes (39) and could not differentiate isozymes in these cells.
Proteolytic degradation of protein ki- nase C by Ca’+-dependent protease or tryp- sin in vitro results in the generation of a

PROTEOLYTIC DEGRADATION OF PROTEIN KINASE C 513
kinase and a phorbol ester binding frag- ment (29-31). This degradative pathway has been proposed as a mechanism for the activation of protein kinase C (23-25) in the intact cells. The accumulation of a Ca2+/PS-independent protein kinase of M, 50,000-60,000 has been observed after treatment of platelets with phospholipase C (38) and PMA (23) and of neutrophils with PMA (24,25). However, a positive im- munological identification of this Ca2+/PS- independent protein kinase as the proteo- lytic fragment of protein kinase C have not been given. In this study we have provided proof with immunological and chromato- graphic analyses that the PMA-induced down-regulation of protein kinase C in EL4 and IEL4 cells results in a rapid deg- radation of the enzyme without accumu- lating the Ca2+/PS-independent catalytic fragment. It appears that the PMA-in- duced proteolytic activation of protein ki- nase C in both EL4 and IEL4 thymoma cells may not play a functional role in the PMA-mediated response.
The PMA-induced growth inhibition and IL-2 secretion were observed in EL4 but not in IEL4 cells. We found that the PMA- induced protein kinase C down-regulation in EL4 was much faster than that in IEL4. It is unknown whether or how protein ki- nase C and its down-regulation are related to the PMA response in these cells. The present data indicated that when degrada- tion was blocked by protease inhibitors, IL-2 secretion was also retarded. Pre- viously, Solanki et al. (20) showed that the PMA-induced down-regulation of phorbol ester binding in HL-60 human promyelo- cytic leukemia cells, but not in the resis- tant cell line R-35, was a crucial event in the control of terminal differentiation. In analogy to these findings, perhaps down- regulation of protein kinase C is also im- portant for IL-2 secretion in EL4. It is in- teresting to note that incubation of IEL4 with high level of PMA (80 or 160 nM) also resulted in an enhanced rate of protein ki- nase C degradation, however, without in- ducing IL-2 secretion even after 40 h of in- cubation. It appears that down-regulation of protein kinase C per se is not sufficient to induce IL-2 secretion. Perhaps the
structural difference in the kinase C of these cells affects their abilities to phos- phorylate certain specific substrates. Al- ternatively, the different PMA responses of these two cell lines could also be due to the differences in their target substrates susceptible to phosphorylation by this kinase as was reported by Kramer and Sando (40).
REFERENCES
1. NIEDEL, J. E., KUHN, L. J., AND VANDENBARK, G. R. (1983) Proc. NatL Acd Sci. USA 80,36- 40.
2. LEACH, K. L., JAMES, M. L., AND BLUMBERG, P. M. (1983) Proc. NatL Acd Sci. USA 80,4208-4212.
3. KIKKAWA, U., TAKAI, Y., MINAKUCHI, R., IN- OHARA, S., AND NISHIZUKA, Y. (1983) J. Biol. Ch. 257,13341-13348.
4. ASHENDEL, C. L., STALLER, J. M., AND BOUTWELL, R. K. (1983) Cancer Res. 43,4333-4337.
5. NISHIZUKA, Y. (1984) Nature (London) 308, 693- 698.
6. BERRIDGE, M. J., AND IRVINE, R. F. (1984) Nature (Landon) 312,315-321.
7. NISHIZUKA, Y. (1986) Science 233,305-312. 8. Kuo, J. F., ANDERSON, R. G. G., WISE, B. C.,
MACKERLOVA, L., SALOMONSSON, I., BRACKETT, N. L., LATOH, N., SHOJI, M., AND WRENN, R. W. (1980) Proc. NatL Acad. Sci. USA 77,7039-7043.
9. MINAKUCHI, R., TAKAI, Y., Yu, B., AND NISHI- ZUKA, Y. (1981) J. B&hem. (Tokyo) 89, 1651- 1654.
10. KRAFT, A. S., ANDERSON, W. B., COOPER, H. L., AND SANDO, J. J. (1982) J. BioL Chem. 257, 13193-13196.
11. KRAFT, A. S., AND ANDERSON, W. B. (1983) Nature (London) 301,621-623.
12. FARRAR, W. L., THOMAS, T. P., AND ANDERSON, W. B. (1985) Nature (London) 315,235-237.
13. FARRAR, W. L., AND ANDERSON, W. B. (1985) Na- ture (London) 315.233-235.
14. HIROTA, K., HIROTA, T., AGUILERA, G., AND CATT, K. J. (1985) J. BioL Chem 260,3243-3246.
15. SOLANKI, V., AND SLAGA, T. J. (1983) Proc. NatL Acad. Sci. USA 78,2549-2553.
16. CHIDA, K., AND KUROKI, T. (1983) Cancer Res. 43, 3638-3642.
17. CHIDA, K., KATO, N., AND KUROKI, T. (1986) J. BioL Chem. 261,13013-13018.
18. COLLINS, M. K. L., AND ROZENGURT, E. (1982) J. Cell PhysioL 112,42-50.
19. YAMASAKI, H., DREVON, C., AND MARTEL, N. (1982) Carcinogmesti 3,905-910.
20. SOLANKI, V., SLAGA, T. J., CALLAHAM, M., AND HUBERMAN, E. (1981) Proc. NatL Acad. Sci. USA 78,1722-1725.

514 HUANG ET AL.
21. FABBRO, D., REGAZZI, R., COSTA, S. D., BORNER, C., AND EPPENBERGER, U. (1986) Biochem, Bic- phys. Res. Commun. 135,65-73.
22. BALLESTER, R., AND ROSEN, 0. M. (1985) J. Biol. Chem 260,15194-15199.
23. TAPLEY, P. M., AND MURRAY, A. W. (1984) Biw &em. Biophys. Res. Commun. 122,158-X4.
24. MELLONI, E., PONTREMOLI, S., MICHETTI, M., SACCO, O., SPARATORE, B., AND HORECKER, B. L. (1986) J. Biol Chem. 261,4101-4105.
25. MELLONI, E., PONTREMOLI, S., MICHETTI, M., SACCO, O., SPARATORE, B., SALAMINO, F., AND HORECKER, B. L. (1985) Proc. Nat1 Acad Sci USA 82,5435-6439.
26. TAKAI, Y., KISHIMOTO, A., INOUE, M., AND NISHI- ZUKA, Y. (1977) J. Biol Chem 252,7603-7609.
27. INOUE, M., KISHIMOTO, A., TAKAI, Y., AND NISHI- ZUKA, Y. (1977) J. Biol Chem 252,7610-7616.
28. KISHIMOTO, A., KAJIKAWA, N., SHISTA, M., AND NISHIZUKA, Y. (1983) J. Biol Chem. 258,1156- 1164.
29. HUANG, K.-P., AND HUANG, F. L. (1986) Biochem Biophys. Res. Commun 139,320-326.
30. HOSHIJIMA, M., KIK~CHI, A., TANIMOTO, T., KAI- BUCHI, K., AND TAKAI, Y. (1986) Cancer Rex 46, 3000-3004.
31. LEE, M.-H., AND BELL, R. M. (1986) J. BioL C&m. 261,14867-14870.
32. FARRAR, J. J., FULLER-FARRAR, J., SIMON, P. L., HILFIKER, M. L., STADLER, B. M., AND FARRAR, W. L. (1980) J. Immund 125.2555-2558.
33. SANDO, J. J., HJLFIKER, M. L., PIACXNTINI, M. J., AND LALJFER, T. M. (1982) Cancer Res 42,1676-X80.
34. HUANG, K.-P. AND HUANG, F. L. (1986) J. Biol. Chem 261,14781-14787.
35. TOWBIN, H., STAEHELIN, T., AND GORDON, J. (1979) Proc. Nat1 Acad sci. USA 76,4350-4354.
36. LAEMMLI, U. K. (1970) Nature (London) 222,680- 685.
37. BRADFORD, M. (1976) Anal. B&hem 72,833-842. 38. TAPLEY, P. M., AND MURRAY, A. W. (1984) Bie
them. Biophys. Res. Ccnnmun. 118.835-841. 39. HUANG, K.-P., NAKABAYASHI, H., AND HUANG, F. L.
(1986) Proc Nczd Acud sci USA 83,8535-8539. 40. KRAMER, C. M., AND SANDO, J. J. (1986) Cancer
Rex 46,3040-3045.




















