
Effect of microwave radiation on the permeability of carbonic anhydrase loaded unilamellar liposomes
11
Bioelectromagnetics 15303-31 3 (1994) Effect of Microwave Radiation on the Permeability of Carbonic Anhydrase Loaded Unilamellar Liposomes Alfonsina Ramundo Orlando, Giuseppe Mossa, and Guglielmo D’lnzeo Institute of Experimental Medicine, CNR, 00137 Rome, Italy (A. R.O., G.M.); Depart- ment of Electronic Engineering, University of Rome “La Sapienza, ” 00 184 Rome, Italy (G. D.) The influence of 2.45 GHz microwave exposure (6 mW/g) on the diffusion processes in enzyme-loaded unilamellar liposomes as bioreactors was studied. The enzyme carbonic anhydrase (CA) was entrapped into cationic unilamellar vesicles. Previous kinetic ex- periments showed a very low self-diffusion rate of the substrate p-nitrophenyl acetate (PNPA) across intact liposome bilayer. A twofold increase in the diffusion rate of PNPA through the lipid bilayer was observed after 120 min of microwave radiation compared to temperature control samples. The microwave effect was time dependent. The enzyme activity, as a function of increased diffusion of PNPA, rises over 120 min from 22.3% to 80%. The increase in stearylamine concentration reduces the enzyme activity from 80% to 65% at 120 min. No enzyme leakage was observed. 01994 Wiley-Liss, Inc. Key words: microwaves, liposome, permeability changes, enzyme INTRODUCTION The increasing emphasis on understanding how microwave radiation can produce the enhancement of transmembrane movement in biological membranes [Olcester et al., 1980; Fisher et al., 1982; Cleary et al., 1982; Liburdy and Penn, 1984; Liburdy and Vanek, 19851has induced several research groups to use simple membrane models. The use of liposomes as a model for biological membranes is well known [Bangham, 19781. In this context, an increase in the leakage of entrapped substances from the interior of liposomes induced by microwave exposure (60 mW/g) has been reported [Liburdy and Magin, 19851. The observation of the effect is not unequivocal [Rafferty and Knutson, 1987; Liu and Cleary, 19881, although some differences in the ex- perimental procedure could explain the divergent results. More recently, an increase in such leakage has been demonstrated, using liposomes temperature insensitive Received for review March 15, 1993; revision received October 4, 1993. Address reprints requests to G. D’Inzeo, Department of Electronic Engineering, University of Rome “La Sapienza,” Via Eudossiana 18, 00184 Rome, Italy. 0 1994 Wiley-Liss, Inc.
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Effect of microwave radiation on the permeability of carbonic anhydrase loaded unilamellar liposomes

Bioelectromagnetics 15303-31 3 (1994)
Effect of Microwave Radiation on the Permeability of Carbonic Anhydrase Loaded Unilamellar Liposomes Alfonsina Ramundo Orlando, Giuseppe Mossa, and Guglielmo D’lnzeo
Institute of Experimental Medicine, CNR, 00137 Rome, Italy (A. R.O., G.M.); Depart- ment of Electronic Engineering, University of Rome “La Sapienza, ” 00 184 Rome, Italy (G. D.)
The influence of 2.45 GHz microwave exposure (6 mW/g) on the diffusion processes in enzyme-loaded unilamellar liposomes as bioreactors was studied. The enzyme carbonic anhydrase (CA) was entrapped into cationic unilamellar vesicles. Previous kinetic ex- periments showed a very low self-diffusion rate of the substrate p-nitrophenyl acetate (PNPA) across intact liposome bilayer. A twofold increase in the diffusion rate of PNPA through the lipid bilayer was observed after 120 min of microwave radiation compared to temperature control samples. The microwave effect was time dependent. The enzyme activity, as a function of increased diffusion of PNPA, rises over 120 min from 22.3% to 80%. The increase in stearylamine concentration reduces the enzyme activity from 80% to 65% at 120 min. No enzyme leakage was observed. 01994 Wiley-Liss, Inc.
Key words: microwaves, liposome, permeability changes, enzyme
INTRODUCTION
The increasing emphasis on understanding how microwave radiation can produce the enhancement of transmembrane movement in biological membranes [Olcester et al., 1980; Fisher et al., 1982; Cleary et al., 1982; Liburdy and Penn, 1984; Liburdy and Vanek, 19851 has induced several research groups to use simple membrane models. The use of liposomes as a model for biological membranes is well known [Bangham, 19781.
In this context, an increase in the leakage of entrapped substances from the interior of liposomes induced by microwave exposure (60 mW/g) has been reported [Liburdy and Magin, 19851. The observation of the effect is not unequivocal [Rafferty and Knutson, 1987; Liu and Cleary, 19881, although some differences in the ex- perimental procedure could explain the divergent results. More recently, an increase in such leakage has been demonstrated, using liposomes temperature insensitive
Received for review March 15, 1993; revision received October 4, 1993.
Address reprints requests to G. D’Inzeo, Department of Electronic Engineering, University of Rome “La Sapienza,” Via Eudossiana 18, 00184 Rome, Italy.
0 1994 Wiley-Liss, Inc.

304 Ramundo Orlando et al.
in the fluid phase, that were exposed to 2.45 GHz fields at 60 mW/g [Liburdy and Fingado, 19901 and 38 mW/g [Saalman et al., 19911.
In this study, we analyzed the influence of microwaves (MWs) on the influx rate of the substrate p-nitrophenyl acetate (PNPA) into carbonic anhydrase (CA)- loaded liposomes. The importance of this enzyme in biological systems is well known [Dodgson, 19911. CA is nearly ubiquitous; it is present in many tissues of differ- ent species, in cells, subcellular particles, and membranes. CA-loaded liposomes could be considered as a very simple model of the erythrocyte. In addition, the physicochemical properties of liposomes have been well characterized. The enzyme leakage from the liposome bilayer has been also investigated. On the basis of our results, we confirm that microwave exposure enhances the permeability in liposomes, far from their phase-transition temperature; moreover, for the first time, we report microwave effects on the lipid bilayer in the presence of cholesterol.
MATERIALS AND METHODS
Chemicals
L-a-dipalmitoylphosphatidylcholine (DPPC), cholesterol (Chol), stearylamine (SA), sodium cholate, n-octyl-P-glucopyranoside, PNPA, and bovine CA (catalog No. C7500) were purchased from Sigma (St. Louis, MO). Chloroform, sodium chloride, acetonitrile, and Tris(hydroxymethy1)amino methane were purchased from Farmitalia Carlo Erba.
Liposome Preparation
The vesicles were prepared by controlled detergent dialysis (DIAL liposomes) [Weder and Zumbuehl, 19841 using a Liposomat apparatus (DIANORM GERATE, Munchen, Germany). A chloroform solution ( 5 ml) containing 30 pmol DPPC, 15 pmol cholesterol, and 5 pmol stearylamine (6:3: 1 ratio) or containing 25 pmol DPPC, 15 pmol cholesterol, 10 pmol stearylamine (5:3:2 ratio), and 100 pmol sodium cholate as detergent (lipid/detergent ratio 1:2) was evaporated to dryness in a rotatory evaporator under vacuum at 30 "C. The suspension was made by adding 4 ml0.09 M Tris (pH 7.55)-0.081 NaCl alone or containing 10 mg CA to prepare empty or loaded liposomes, respectively. Then, the liposomes were formed by controlled dialysis of detergent for 18 h at 23 "C. The liposomes were washed six times in Tris-saline, followed by centrifugation at 12000g to remove the free enzyme. The resulting unilamellar liposomes are stable for at least 3-4 days at storage temperature of 4 "C; however, all experiments were carried out as soon as possible, usually 12 h, after the end of the preparation.
Liposome DPPC content was determined according to the method of Stewart [ 19801; Chol was determined with the Farb test (Boehringer Mannheim Kit), and SA was analyzed by gas chromatography-mass spectrometry, as described elsewhere [Passi et al., 19911, Protein content was determined with a modification of the Lowry method [Oyama and Eagle, 19561, with bovine serum albumin as standard. Size distribution measurements of vesicles were determined by negative staining [Gre- gory and Pirie, 19731 observed under a Philips CM12 transmission electron micro- scope (EM) at 80 kV. Liposome diameters were measured directly on the EM films using a Quantimet 570 images analysis system (Cambridge Instruments). The re- sults were obtained as diameters of the equivalent circles.

Microwave Effects on Liposome Permeability 305
Enzyme Activity Measurements
Enzyme activity measurements were performed in 0.09 M Tris-saline at pH 7.55, using PNPA as substrate (from 0.5 to 2 mM) and following the p-nitrophenolate anion appearance at 400 nm wavelength on a UVICON 930 spectrophotometer (KONTRON).
Kinetic studies of CA were performed by adding 0.06 ml free enzyme solu- tion (0.03 mg/ml) or 0.06 ml CA-loaded vesicle suspension to 3 ml Tris-saline. The disruption by detergent was performed as follows. Liposome suspensions (0.5 ml) were incubated for 2 h at 37 "C with n-octyl-P-glucopyranoside (0.5 ml) at 1:6 Fmol lipid/detergent ratio. After incubation, the suspension was centrifuged at 12000g to sediment the lipids. The resultant clear supernatants (0.120 ml) were used for kinetic measurements and for determination of protein contents. Only those lipo- some preparations 6:3:1 and 5:3:2, showing as recovery of CA proteinic values of 0.128 t 0.02 mg and 0.1 55 k 0.03 mg, respectively, were used for the subsequent experiments of microwave exposure. All spectrophotometer measurements were carried out at a constant room temperature of 23 "C; however, between measure- ments the samples were placed in an ice bath. The procedure for assaying the en- zyme leakage from liposomes involved rapidly sedimenting the sample or control liposome suspensions for 5 min in a Beckman L8-55 Ultracentrifuge in a 50-VTi rotor at 35000 rpm (170000g) according to the method of Magin and Weinstein [ 19841. Samples of the resultant clear supernatant were immediately used for the enzyme activity measurements.
Microwave Exposure
In order to expose the samples to a field controlled in amplitude, frequency, and polarization, an S-band exposure chamber was designed (Fig. 1 ) . A coaxial to rectangular waveguide transition was connected to a waveguide with the same trans- verse section, where water was circulated from a thermostated bath placed far from the exposure set-up. Since the characteristic impedance of the water-filled waveguide is high in comparison with the air-filled coaxial-waveguide transition, it has been necessary to insert a quarter-wavelength of dielectric matching material. We choose porcelain as a mechanical stable material with a high relative dielectric constant. The input reflection coefficient was measured using a network analyzer (HP 85 10B). At the working frequency of these experiments (f = 2.45 GHz), about 90% of the power supplied from the generator is delivered in the chamber. No significant modification of the reflection coefficient was measured with insertion in the thermostated water-filled waveguide of the cuvettes containing liposome suspen- sions. The exposure chamber allows the cuvettes to be placed in a Teflon square grid of 6 x 3 in order to have at least three data for each experiment and, at the same time, the possibility of different values of power incident on the same preparation. In this set of experiments, the cuvettes were placed only in the middle of the grid described above to obtain six samples irradiated at the same power.
The SAR of the samples has been evaluated both on the basis of the calcu- lated field at the porcelain-water interface, obtained from the measure on the in- cident and reflected powers and from the values obtained by temperature increase inside a water-filled cuvette in absence of thermostatic control. Both the calcula- tion and the measurement gave the same SAR value. The frequency and the power supplied from the microwave generator (Weinschel Engineering 430B) were con-

306 Ramundo Orlando et al.
EXPOSURE SYSTEM - TRANSVERSAL VIEW
T Z
A=- t I Lsr
COAX-WAVEGUIDE TRANSITION BANDWIDTH INPUT 1.7+2.6 GHz
Fig. 1. Exposure system used to irradiate liposome suspensions.
trolled by means of a spectrum analyzer (HP 8565A) through a high-directivity directional coupler. The sample temperature was continuously monitored using a Luxtron lO0OB (minimum AT = 0.01 "C) fluoroptic thermometer, with the probe placed in the central point of maximum power intensity.
Experimental Procedure
Aliquots (0.06 ml) of CA-loaded or empty liposomes in a final volume of 0.5 ml Tris-saline (pH 7.55) were placed in a 3 cm3 silica cuvette positioned in con- tact with the bottom part of the exposure chamber, as previously described. Dupli- cate liposome samples were exposed to microwaves for 15,30,60,90, or 120 min. The control liposomes were maintained under the same conditions in a water bath kept at the same constant temperature of the MW-exposed samples. At the end of exposure, the cuvettes were removed and the enzyme activity measurements made i mmedi ate1 y.
RESULTS
The enzyme-loaded liposomes, prepared via the detergent dialysis method, were almost 100% unilamellar and homogeneous, while the increase in surface positive charge due to stearylamine, in liposomes with a 5:3:2 lipid ratio, signifi- cantly decreased the size distribution profile of the vesicles. The mean vesicle diameter of liposomes was 61 k 18 nm (Fig. 2a) and 29 rt 11 nm (Fig. 2b; 6:3:1 and 5:3:2, respectively). In the first attempt, we studied the effect of microwave radiation on free CA. As was described in Materials and Methods, we performed

Microwave Effects on Liposome Permeability 307
enzyme activity measurements after MW exposure with the same conditions as were used in the subsequent experiments with CA-loaded liposomes. The results (Table 1) showed that the catalytic activity of CA was not impaired by micro- wave radiation.
As regards liposomes, to evaluate the esterase activity of CA present inside the liposomes, we studied the kinetics of the enzyme on CA-loaded liposomes after disruption by detergent, with disrupted empty liposomes as reference. The overall hydrolysis rate of PNPA (2mM) showed a significant difference in the reaction rate
Field Nunhev 1.6
25 sa 7 5 in0 1.25 158 175 2 E w t i u . Circle Dian. < n a n o n e t r i 1
b Count F i e l d N u n h e r 6 5
X HEClN --I 61.21
[ X MEDIPlN 6 % . 77
X STD DEU 17.78
Y T O l A L
-I I
Fig. 2. a: Vesicle size distribution from EM films of negatively stained DPPC:Chol:SA = 6:3:1 CA- loaded liposomes. A minimum of 450 vesicles was measured. The values of the mean diameter (61.21) and of the standard deviation (17.7) are reported at upper right. b: Vesicle size distribution from EM films of negatively stained DPPC:Chol:SA = 5 : 3 : 2 CA-loaded liposomes. A minimum of 510 vesicles was measured. The values of the mean diameter (28.8) and the standard deviation (10.94) are reported at upper right.

308 Ramundo Orlando et al.
TABLE 1. Effect of 2.45 GHz Radiation on the Activity of Free Carbonic Anhvdrase
Time Enzymatic activity Sample (rnin) (A Ab/min)
Control (n = 5 ) 0 0.0374 k 0.0005 120 0.0407 + 0.0006
Exposed (n = 5) 15 0.0392 + 0.001 30 0.0372 k 0.002 45 0.0378 k 0.001 60 0.0387 f 0.003 90 0.0397 f 0.003
120 0.0388 f 0.002
between empty and CA-loaded liposomes. This difference was equal to the enzy- matic hydrolysis due to the esterase activity of CA liberated from liposomes. Moreover, after liposome disruption, the kinetic data can be fitted by the Michae- lis-Menten equation, with the same K, value as for the free enzyme (data not shown). The esterase activity in loaded liposomes was normalized in different liposome preparations and expressed as 100%. As is shown in Figures 3 and 4, at zero time of exposure, the results showed a nonzero rate of enzymatic activity, perhaps due to CA located on the external surface of intact liposomes. This activity is typical of such intact liposome preparations, showing mean values of 22% f 7% and 27% k 3% in 6:3: 1 and 5:3:2 liposomes, respectively.
After microwave exposure (6 mW/g) times ranging from 90 to 120 min, at a temperature of 20.3 k 0.1 OC, we obtained a significant increase from 41.2% k 7% to 80% f 11 % ( P < 0.05) in the esterase activity. This increment was calculated
* p<0.05 I I I I I I I I 1
0 30 60 90 120 150 time of exposure (min)
Fig. 3. Microwave-stimulated diffusion of PNPA through CA-loaded liposomes (DPPC:Chol:SA = 6:3: 1). The measured SAR was 6 mW/g. The temperature during the experiments was 20.3 f 0.1 "C.
Microwave Effects on Liposome Permeability 309
lo0T 20.3 "C
80
60
4 0
20
Exposed
Control
** p<0.05 * p<o.1
0 I , I I I I I I i
0 30 60 90 120 150
time of exposure (min)
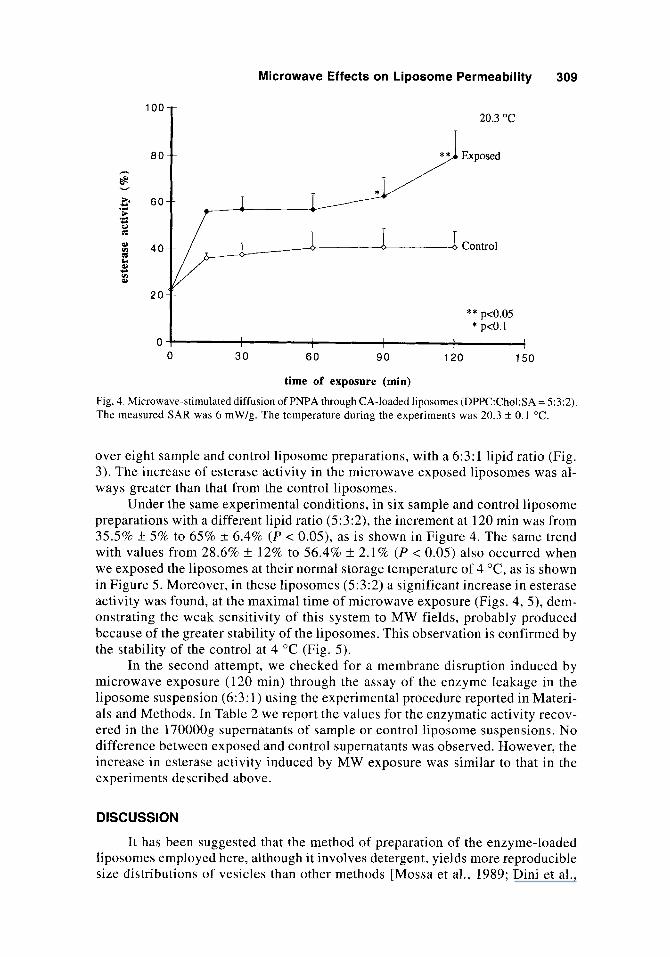
Fig. 4. Microwave-stimulated diffusion of PNPA through CA-loaded liposomes (DPPC:Chol:SA = 5:3:2). The measured SAR was 6 mW/g. The temperature during the experiments was 20.3 ? 0.1 "C.
over eight sample and control liposome preparations, with a 6:3:1 lipid ratio (Fig. 3). The increase of esterase activity in the microwave exposed liposomes was al- ways greater than that from the control liposomes.
Under the same experimental conditions, in six sample and control liposome preparations with a different lipid ratio (5:3:2), the increment at 120 min was from 35.5% k 5% to 65% k 6.4% ( P < 0.05), as is shown in Figure 4. The same trend with values from 28.6% f 12% to 56.4% k 2.1% ( P < 0.05) also occurred when we exposed the liposomes at their normal storage temperature of 4 "C, as is shown in Figure 5. Moreover, in these liposomes (5:3:2) a significant increase in esterase activity was found, at the maximal time of microwave exposure (Figs. 4, 5 ) , dem- onstrating the weak sensitivity of this system to MW fields, probably produced because of the greater stability of the liposomes. This observation is confirmed by the stability of the control at 4 "C (Fig. 5) .
In the second attempt, we checked for a membrane disruption induced by microwave exposure (120 min) through the assay of the enzyme leakage in the liposome suspension (6:3: 1 ) using the experimental procedure reported in Materi- als and Methods. In Table 2 we report the values for the enzymatic activity recov- ered in the 170000g supernatants of sample or control liposome suspensions. No difference between exposed and control supernatants was observed. However, the increase in esterase activity induced by MW exposure was similar to that in the experiments described above.
DISCUSSION
It has been suggested that the method of preparation of the enzyme-loaded liposomes employed here, although it involves detergent, yields more reproducible size distributions of vesicles than other methods [Mossa et a]., 1989; Dini et al.,

310 Ramundo Orlando et al.
100
80 h
5
2 0
4.0 “C
Control
0
0 30 60 90 120 150 time of exposure (min)
Fig. 5. Microwave-stimulated diffusion of PNPA through CA-loaded liposomes (DPPC:Chol:SA = 5:3:2). The measured SAR was 6 mW/g. The temperature during the experiment was 4 f 0.1 “C.
19911. Considerable attention is being given to the chemical composition of lipo- somes, which significantly influences their permeability properties. The bilayer contains phosphatidylcholine, charged lipid (stearylamine), and cholesterol. In this context, the presence of cholesterol on bilayer affects the mobility of the phospholipids so that the lipid bilayer reaches an “intermediate state” i n which the hydrophobic interior has a high viscosity [Demel and de Kruijff, 19761. The membrane viscos- ity directly affects the rates of simple diffusion processes. Thus the increase in cholesterol concentration in liposome bilayer results in a significant reduction in permeability. We used a cholesterol concentration of 30 mol% in the liposome bilayer, which has a marginal effect on the position of the main phase-transition tempera- ture (Tc) [New, 19901. The liposomes undergo a phase transition at approximately 42 “C as reported [Papahadjopoulos et al., 19731 for DDPC vesicles. Moreover, cholesterol phospholipid ratios from 0.1 1 to 0.33 are usually found in plasma and other subcellular membranes [Demel and de Kruijff, 19761. Thus the quality of
TABLE 2. Effect of 2.45 GHz Irradiation on Enzyme Leakage From Liposomes (DPPC:Chol:SA = 6:3:1)
Exposure time Activity A(Exp. - Control) Sample (min) (mean f sd) (mean f sd)
Control 120 21.2 * 7 Exposed 120 58.5 * 2.08 37.2 f 8 Control supern.“ - 17.6 f 2.5 Exposed supema - 19.4 3~ 2.8 1.8 * 0.4 “17000Og supernatants obtained from sedimentation of control or exposed samples after 120 min of treatment.

Microwave Effects on Liposome Permeability 31 1
liposome preparations makes possible a correct comparison between pure lipid bilayers and biomembranes, especially for their permeability properties, in rela- tion to the microwave effects observed.
Although it has been proposed [Liburdy and Magin, 19851 that increased liposome leakage as a result of microwave exposure should occur near the Tc, evidence that increased liposome leakage as a result of microwave exposure occurs far from the Tc has been reported [Liburdy and Fingado, 1990; Saalman et al., 19911. Our results substantiate the latter observations by demonstrating an increase in substrate permeation rate through the liposomes bilayer well below (20 "C) their phase-tran- sition temperature (42 "C). In addition, the lack of enzyme leakage from liposome bilayers exposed to microwaves, as determined by recovery of enzyme in the 170000g supernatant, excludes the possibility that the mechanism responsible for the increase in liposome permeability involves gross membrane disruption, in agreement with previous reports [Liburdy and Magin, 1985; Saalman et a]., 19911.
On the basis of these findings, the mechanism through which the microwave fields induce the increase in membrane permeability is consistent with the forma- tion of pores. In this context we can exclude a simple thermal-based explanation for our results that might involve local heating at the liposome surface, whether for the low conditions of SAR (6 mW/g) employed or for the diffusion of heat from the thin membrane to the surroundings. Diffusion is highly efficient and should prevent local temperature rise, as was recently demonstrated [Saalman et al., 19911. Our results demonstrate that the addition of cholesterol at 30 mol% to the liposome bilayer does not render this system insensitive to microwave fields, although it has been suggested [Liburdy and Vanek, 19851 that the addition of cholesterol obliterates the effect of 2.45 GHz microwave fields on the erythrocyte's permeability increase.
On the other hand, in the vesicles with 5:3:2 lipid ratio, we obtained an over- all decrease of the substrate influx rate compared to that of the 6:3: 1 vesicles. In the 5:3:2 case, the higher cholesterol phospholipid ratio could act to stiffen the membrane and oppose the field, in agreement with the studies mentioned above [Liburdy and Vanek, 19851. At the same time, we should consider that the higher net surface charge in the 5:3:2 vesicles might be involved in the different response of liposomes to microwave fields, since the permeability effect could be dependent on electrostatic charge.
In an attempt to verify this hypothesis, we prepared neutral liposomes (DPPC:Chol = 7:3), but in this case the entrapment of CA was not obtained. We could not evaluate the effect of microwave fields on bilayer devoid of surface charge. However, studies [Liburdy, 19921 conducted on phase-transition liposomes carry- ing either a net negative or a net positive charge have shown a similar effect of mi- crowave fields on the release of entrapped solute, at pre-Tc temperature.
The reduction in diameter of vesicles 5:3:2 (29 k 11 nm) in comparison to 6:3:1 (61 k 18 nm) due to increasing SA concentration could also be a factor re- sponsible for the differences in the MW-induced effects. These results substanti- ate previous observations on the dependence of the permeability increase induced by MW as a function of liposome diameter; in fact, the smaller DPPC:DPPG lipo- somes (200 nm vs. 140 nm) showed a reduced permeability increase (Liburdy, private communication).
Our observations indicate that the microwave fields could induce a rotational motion in the phospholipid acyl chains, with subsequent pore formation in lipids

312 Rarnundo Orlando et al.
in the “intermediate gel” state, formed in the presence of cholesterol, and at tem- peratures well below Tc, where they are not normally leaky.
ACKNOWLEDGMENTS
We thank Prof. A. Finazzi-Agro (Department of Experimental Medicine and Biochemical Science, University “Tor Vergata,” Rome) for stimulating discussion and useful advice throughout this work as well as Dr. R.P. Liburdy (Bioelectromagnetics Research Facility, Berkeley, California) for his valuable comments. We also thank Dr. S. Passi (S. Gallicano Hospital, Rome) for his friendly help in assaying the stearylamine and Dr. A.L. Serafino for the electron micros- copy analysis.
REFERENCES
Bangham AD (1978): Properties and uses of lipid vesicles: An overview. In Papahadjopoulos D (ed): “Liposomes and Their Uses in Biology and Medicine.” Ann NY Acad Sci 308:3-7.
Cleary S, Garber F, Liu LM (1982): Effects of X-band microwave exposure on rabbit erythrocytes. Bioelectromagnetics 3:453-466.
Demel RA, de Kruijff B (1976): The function of sterols in membranes. Biochim Biophys Acta 21 1:373-375.
Dini L, Di Giulio A, Pavan A, Ravagnan G, Mossa G (1991): Size and stability of dipalmitoylphosphatidylcholinelcholesterol unilamellar vesicles are affected by interaction with proteins. Biochim Biophys Acta 1062: 108-1 12.
Dodgson SJ (1991): The carbonic anhydrases. Overview of their importance in cellular physiology and in molecular genetics. In Dodgson SJ, Tashian RE, Gros G, Carter ND (eds): “The Car- bonic Anhydrases.” New York: Plenum Press, pp 3-14.
Fisher PD, Paznasky MJ, Gross WAG (1982): Effect of microwave radiation (2450 MHz) on the active and passive components 14Na+ efflux from human erythrocytes. Radiat Res 92:411-422.
Gregory DW, Pirie BJ (1973): Wetting agents for biological electron microscopy. I. General consid- eration and negative staining. J Microsc 99:2S 1-255.
Liburdy RP (1992): The influence of oscillating electromagnetic fields on membrane structure and function: Synthetic liposomes and the lymphocyte cell membrane with direct application to the controlled delivery of chemical agents. In Nordkn B, Ramel C (eds): “Interaction Mechanisms of Low-Level Electromagnetic Fields in Living Systems.” New York: Oxford University Press,
Liburdy RP, Fingado B (1990): Effects of microwave fields on liposome permeability: Non-phase transition liposome vesicles. Twelfth Annual Bioelectromagnetics Society Meeting, San An- tonio, Texas, 10-14 June, Abstract B-4-5.
Liburdy RP, Magin RL (1985): Microwave-stimulated drug release from liposomes. Radiat Res 103:266-275.
Liburdy RP, Penn A (1984): Microwave bioeffects in the erythrocyte are temperature and PO, depen- dent. Bioelectromagnetics 5:283-291.
Liburdy RP, Vanek PF Jr (1985): Microwaves and cell membrane. 11. Temperature, plasma, and oxygen mediated microwave-induced membrane permeability in the erythrocyte. Radiat Res 102: 190-205.
Liu Li-Ming, Cleary SF (1988): Effects of 2.45 GHz microwave and 100-MHz radiofrequency ra- diation on liposome permeability at the phase transition temperature. Bioelectromagnetics 9:249-257.
Magin RL, Weinstein JN (1984): The design and characterization of temperature- sensitive lipo- somes. In Gregoriadis G (ed): “Liposome Technology,” Vol. 3. Boca Raton, FL: CRC Press, pp 137-155.
Mossa G, Di Giulio A, Dini L, Finaszi-Agr6 A (1 989): Interaction of dipalmitoylphosphatidyIcholine/ cholesterol vesicles with ascorbate oxidase. Biochem Biophys Acta 986:3 10-3 14.
pp 259-279.

Microwave Effects on Liposome Permeability 313
New RRC (1990): Introduction, Chapter 1. In New RRC (ed): “Liposomes. A Practical Approach.” Oxford: IRL Press, pp 1-32.
Olcester RB, Belnian S , Eisenbud M, Mumford WW, Rabinowitz J (1980): The increased passive efflux of sodium and rubidium from rabbit erythrocytes by microwave radiation. Radiat Res 82:244-256.
Oyama VI, Eagle H (1956): Measurements of cell growth in tissue culture with a phenol reagent (Folin- Ciocalteau). Proc SOC Exp Biol Med 91:305-307.
Papahadjopoulos D, Jacobson K, Nir S , Isac T (1973): Phase Transition in phosholipid vesicles. Fluo- rescence polarization and permeability measurement concerning the effect of temperature and cholesterol. Biochem Biophys Acta 3 11:330-348.
Passi S, Picardo M, Morrone A, De Luca C, Ippolito F (1991): Skin surface lipids in HIV sero-posi- tive and HIV sero-negative patients affected with seborrheic ferrothiocyanate. J Dermatol Sci
Rafferty CN, Knutson JR (1987): Effects of pulsed fields on soluble proteins and liposomes. Paper presented at the IEEE/Ninth Annual Conference of the Engineering in Medicine and Biology Society, 0701.
Saalman E, NordCn B, Arvidsson L, Hamnerius Y, Hojevik P, Connell KE, Kurucsev T (1991): Ef- fect of 2.45 GHz microwave radiation on permeability of unilamellar liposomes to 5(6)- carboxyfluorescein. Evidence of non-thermal leakage. Biochim Biophys Acta 1064: 124-1 30.
Stewart JCM (1980): Colorimetric determination of phospholipids with ammonium ferrothiocyanate. Anal Biochem 104: 10-14.
Weder HG, Zumbuehl 0 (1984): The preparation of variably sizes homogeneous liposomes for labo- ratory, clinical, and industrial use by controlled detergent dialysis. In Gregoriadis G (ed): “Liposome Technology,” Vol. 1. Boca Raton, FL: CRC Press, pp 79-107.
2:84-9 I .




















