
An ovine tracheal explant culture model for allergic airway inflammation
Upload
independentCategory
view
2download
0
THE ANATOMICAL RECORD 200:357-370 (1981)
A Method for Culturing Canine Tracheal Smooth Muscle Cells In Vitro: Morphologic and Pharmacologic Observations
BARRY P. AVNER, JUDI DELONGO, STEVE WILSON, AND A.J. LADMAN Departments of Pharmacology and Anatomy, The University of New Mexico, School of Medicine, Albuquerque, New Mexico 87131
ABSTRACT A method of culturing canine tracheal smooth muscle cells in vitro is described. The morphology of these cells is monitored up to 60 days in culture and selected stages are illustrated. The characteristics of these cells are numerous mechanical attachments, the presence of thick filaments in suitably processed cells, and their contractile response to in vitro administration of car- bachol, a cholinomimetic drug. They also possess nexus formations and both thin (actin) filaments and 10-nm filaments. Mitosis is found in the nonconfluent prep- arations up to 16 days after culturing. Cultures of 2 to 8 days appear to be most useful as pharmacological test vehicles. This system will be used to explore the phenomenon of adrenergic beta-2 receptor desensitization in airway smooth mus- cle, to attempt to localize these receptor sites and to determine how receptor affinity and/or number may be regulating cell response to pharmacologic agents.
A variety of human respiratory diseases are characterized by the constriction of airway smooth muscle. Topical or systemic therapy with agents known to relax this smooth muscle is routinely offered as treatment for this con- dition. In the development of our understand- ing of the mechanism by which these agents induce relaxation, it was found that the mem- branes of airway smooth muscle possess ad- renergic beta-2 receptors, the activation of which leads to relaxation.
Beta-2 receptors are known to occur in the membranes of amphibian, avian, and mam- malian peripheral blood cells, and since these are relatively easy to obtain and are abundant sources of receptors, investigations of the prop- erties of beta-2 receptors have routinely used such material. Data obtained from these stud- ies have been applied to considerations of air- way smooth muscle function.
It is reasonable to think that there might be significant differences between the beta-2 re- ceptors of cells so widely divergent as periph- eral blood and airway smooth muscle. Fur- thermore, in dealing with the phenomena of bronchial constriction and relaxation, use of airway smooth muscle seems a more rational approach for examining the effects as well as the mechanisms of a variety of drugs which may be ordered for use in respiratory problems. On this basis, studies were undertaken to de- velop a method for culturing airway smooth
muscle with a view to the eventual examina- tion of the beta-2 receptors in their cellular membranes.
Smooth muscle from vascular and enteric sources has been grown in culture and studied extensively (Chamley-Campbell et al., 1979; Rifas et al., 1979; McLean et al., 1979; Small and Sobieszek, 1980; Ross and Kariya, 1980); that from the trachea and bronchi have not been investigated currently; however, two ear- lier studies by Lewis (1924) and Schopper (1935) indicated that these sources should pro- vide excellent opportunities for study.
The present paper reports a method for the successful culturing of canine tracheal smooth muscle in vitro, describes the morphology of these cells under these conditions, and uses the cholinomimetic drug carbachol to test for the functional integrity of the contractile mecha- nism of these smooth muscle cells in culture.
MATERIALS AND METHODS
General Young (6-8 months) mongrel dogs of either
sex were anesthetized with sodium pentobar- bital (30 mg/kg) and their tracheae were re- moved using clean but not aseptic technique. Each trachea was cut into two or three seg- ments and placed in a wax-lined petri dish con-
Received December 23. 1980; accepted January 12,1981.
0003-276X/81/2003-0357$04.00 0 1981 ALAN R. LISS . INC.

358 BARRY P. AVNER, JUDI DELONGO, STEVE WILSON, AND A.J. LADMAN
taining cold (4°C) oxygenated Krebs-Ringer- Hepes buffer (KRH): NaCl (105 mM), KC1 (5 mM), CaCl, (2 mM), KH2P04 (1 mM), MgSO, (1 mM), glucose (14 mM), 10 pg/ml phenol red, Hepes (25 mM), pH 7.4. The tracheal segments were cleaned and the external layer of smooth muscle was separated from the medial layer of fibrous connective tissue and the cartilage rings with the aid of a dissecting microscope.
Tissue culture The smooth muscle was transferred aseptic-
ally to a fresh petri dish containing sterile KRH buffer and washed two or three times with fresh buffer. The muscle was minced into thin 2-3-mrn squares and centrifuged at 150 x g for 5 minutes at 4°C. The pellet was re- suspended in KRH buffer containing 0.2 mM CaCl, and recentrifuged as before. The washed smooth muscle fragments (approximately 1 gm tissue wet weight) were resuspended in 10 ml of 0.2 mM C a t + KRH buffer containing 19.4 mg Type IV collagenase (Sigma, 350 units/ml); 10.5 mg elastase (Sigma, 100 units/ml); 0.3 mg DNAse (Worthington); and 10 mg soybean trypsin inhibitor (Ives et al., 1978). The smooth muscle fragments and sterile enzyme solution were transferred aseptically to a siliconized, gauze-plugged vial and incubated 70 minutes at 37°C in a shaker water bath set at 120 strokes per minute. After 30 minutes the frag- ments had become visibly turbid. The slurry was then subjected to mild pipetting every 10 minutes for the next 30 minutes with a Pasteur pipette to hasten the release of individual cells. The reaction was then terminated by the ad- dition of 10 ml cold 2 mM Ca++ KRH buffer and the suspension was passed successively through sizes 240,120, and 75 pm Nytex mesh to remove large undigested cell clumps and debris. The filtrate was spun at 150 x g for 10 minutes at 4°C and the enzyme solution was decanted. The cell pellet was then washed two times with fresh KRH buffer as before. Cell counts were determined using a hemocyto- meter and viability was determined by means of the trypan blue exclusion method of Tolnai (1975). Aliquots of the cell suspension contain- ing 2.0 x lo5 viable cells were seeded into prewarmed 100-mm tissue culture dishes con- taining Medium 199 (Gibco) plus 10% heat-in- activated newborn calf serum (Biocell), 50 unitdm1 penicillin, 50 kg/ml streptomycin, and 2.2 gm sodium bicarbonate per liter, pH 7.4. The culture dishes were placed in an in- cubator and maintained at 37°C with humi- dified atmosphere containing 5% COz. After 48
hours, the medium was replaced with fresh culture medium, and thereafter, the culture medium was replaced every 72 hours. The cell yield varied from 1.0 x 106-2.0 x lo7 cells per gram of starting material with a viability of 70-95%. The yield can be substantially in- creased by omitting the final two Nytex filtra- tion steps, although the filtrate will consist of a mixture of single cells and small cell clumps.
Randomly selected cultures were spot checked for Mycoplasma contamination by the technique of Hayflick (1965); no contamination was observed in these cultures.
Morphological Samples from cultures were prepared for ex-
amination by means of light and transmission electron microscopy (TEM) by the following procedures.
Cells were fixed in situ at room temperature for 1-2 hours in 2.5% glutaraldehyde, prepared in 0.075 M cacodylate buffer at pH 7.3 to which 0.1% calcium chloride was added. Cells were postfixed at room temperature in 1% 090, and prepared in the same buffer for approximately 1 hour. They were collected either as pellets by centrifugation or processed in situ, depend- ing upon the experimental need. All prepara- tions were routinely stained en bloc with 1% magnesium uranyl acetate, prepared in saline either overnight at 5°C or for 2 hours at room temperature.
Cells were dehydrated in ascending concen- trations of ethanol, embedded in Epon 812, and cut with diamond knives with a Huxley-Cam- bridge ultramicrotome. One- to two-microme- ter sections of all specimens were routinely examined in order to select those for obser- vation by transmission electron microscopy.
Representative samples of toluidine-blue- stained thick sections of Epon-embedded smooth muscle cells were examined for mitotic activity by two of the authors concurrently, using a double-header binocular microscope. Between 175 and 1,250 cells were counted in random fields of culture samples taken from preparations grown in culture for 5, 8, 9, 11, 17,36, and 60 days, and the number of mitotic figures was recorded as a function of the num- ber of whole nuclei in the field. These data were expressed qualitatively as a mitotic index; i.e., the number of mitoses per 100 cells.
Light microscopic observations of the con- tractile behavior of cultured cells were made with a Zeiss photomicroscope equipped with phase-contrast optics and recorded on Kodak Panatomic-X 35-mm film. Cultures to be ob-

CANINE TRACHEAL MUSCLE IN VITRO 359
served and recorded were handled in the fol- lowing manner. Medium 199, in which the cul- tures were grown, was replaced with KRH buffer containing 2 mM Ca++. This was done in order to control changes in pH which would have occurred during the period of drug ex- posure if the bicarbonate buffer was used. Cells to be photographed were grown on siliconized cover slips for 2 to 8 days. Coverslips were “siliconized by dipping them in Sigmacote (Sigma), allowing them to air dry, rinsing them in distilled water to remove surface im- purities, and finally sterilizing them in prep- aration for tissue culture. The cover slips con- taining the cells were blotted on their free surface and inverted so that the cells were sandwiched between a glass slide and the cover slip with a drop of KRH buffer. The cells were continually bathed in warmed KRH buffer (35°C) by applying bibulous paper to one side of the preparation, which permitted fresh buffer to be drawn across the interface between the glass surfaces by capillary action. The slide preparations were then placed on the micro- scope stage, usually surrounded with a heat curtain to maintain temperature a t about 32-35°C. They were observed for a control pe- riod in KRH buffer, photographed, and then exposed to the cholinomimetic drug carbachol (lo-* M), prepared in KRH buffer, and repho- tographed every 4 minutes for the next 20 minutes. Prints from these exposures were mounted next to each other and each cell in the field was systematically examined for evi- dence of shortening. Volume density changes in cultures were assessed by overlaying a 5- mm-square grid acetate sheet on the above photographs and counting the interception of cells with grid corners.
Cultures of HLM-15 and IMR-90 fibroblasts (courtesy of Dr. R.O. Kelley, University of New Mexico) were bathed in comparable media and were tested for their contractile properties af- ter exposure to carbachol by observing their responses as outlined above for smooth muscle cultures.
Electron microscopic data were collected by use of Hitachi HS-7S and HU-11C instru- ments, using Kodak electron microscopic film 4489 developed in D-19 1:l for 4 minutes a t 20°C with intermittent agitation. Negatives were photographically enlarged to appropriate sizes with a Beseler enlarger.
The kinds and numbers of the 23 cultures examined for this report are: tracheal smooth muscle from intact trachea-three; tracheal smooth muscle after separation of connective
tissue by dissection-one; enzymatically sep- arated cells prior to culture-five; cultured cells after 1 day-three; after 5 days-one; af- ter 8 days-two; after 9 days-one; after 11 days--one; after 16 days-two; after 21 days- one; after 28 daye-one; after 36 days-one; after 60 days-one. Other cultures at these times and at different intervals were used in selective pharmacological tests with or with- out morphological controls. These will be re- ported subsequently elsewhere.
RESULTS
Tracheal smooth muscle dissected clear of almost all surrounding connective tissue ele- ments and fixed by immersion appears struc- turally as described in current textbooks of histology (Bloom and Fawcett, 1975; Gauthier, 1977; Copenhaver et al., 1978). The significant ultrastructural appearances of this tissue are shown in Figures 1-3. Easily noted are cav- eolae of uniform size (approximately 45 nm diameter) localized in variable numbers be- neath the plasmalemma, and cytoplasmic fil- aments and densities which are presumed to represent the contractile apparatus of smooth muscle cells (Fig. 1). When such muscle is fixed in a relaxed condition and sectioned perpen- dicular to the orientation of the contractile proteins, thick and thin filaments can be read- ily seen in the cytoplasm (Figs. 2, 3). Imme- diately beneath the plasmalemma (Fig. 2) is a thin, approximately 300 nm cortical area of cytoplasm with an increased density compared to that exhibited by cytoplasm deeper within the substance of the myoplasm. Such increased peripheral density relates to the filamentous nature of the subplasmalemmal cytoplasm and corresponds to the additive presence of actin and increased numbers of the 10-nm filaments that attach to the thickenings of the plasma membrane known as anchoring densities (Small and Sobieszek, 1980). Such anchoring structures are identified by arrowheads in Fig- ure 2; basal laminae invest these muscle fibers in this figure and appear as a flocculent density external to the plasmalemma. In these isolated specimens of smooth muscle tissue, we have chosen not to illustrate the nexus between ad- jacent cells since this is shown in culture in Figure 11.
After enzymatic treatment, smooth muscle tissue separates into single cells or small ag- gregates of cells (Figs. 4, 5). The cells stain variably with toluidine blue; the darker cells with serrated borders (Fig. 5) as seen by light microscopy are identified as smooth muscle
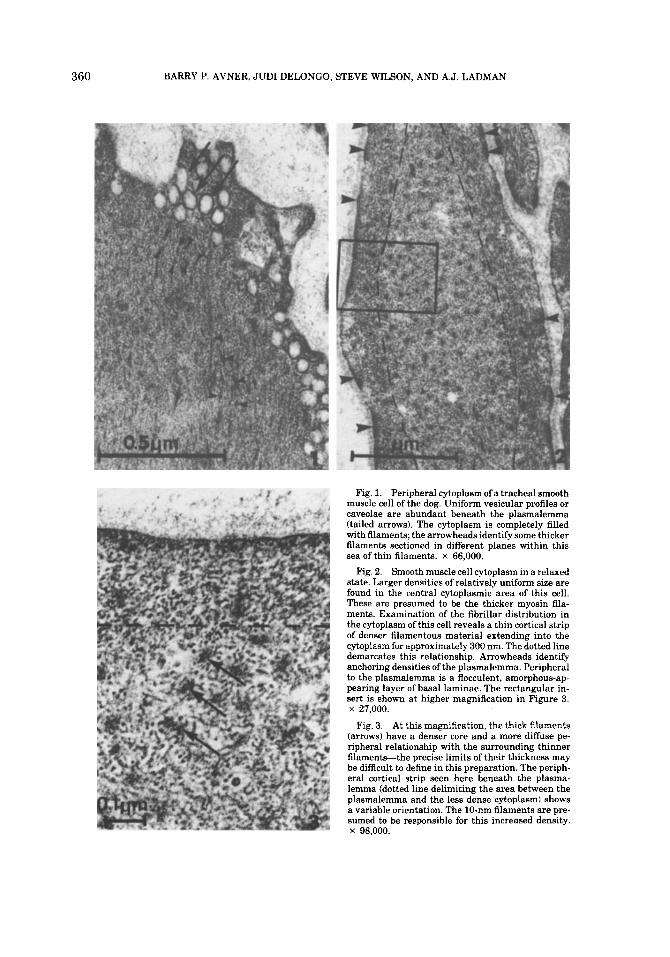
360 BARRY P. AVNER, JUDI DELONGO, STEVE WILSON, AND A.J. LADMAN
Fig. 1. Peripheral cytoplasm of a tracheal smooth muscle cell of the dog. Uniform vesicular profiles or caveolae are abundant beneath the plasmalemma (tailed arrows). The cytoplasm is completely filled with filaments; the arrowheads identify some thicker filaments sectioned in different planes within this sea of thin filaments. x 66,000.
Fig. 2. Smooth muscle cell cytoplasm in a relaxed state. Larger densities of relatively uniform size are found in the central cytoplasmic area of this cell. These are presumed to be the thicker myosin fila- ments. Examination of the fibrillar distribution in the cytoplasm of this cell reveals a thin cortical strip of denser filamentous material extending into the cytoplasm for approximately 300 nm. The dotted line demarcates this relationship. Arrowheads identify anchoring densities of the plasmalemma. Peripheral to the plasmalemma is a flocculent, amorphous-ap- pearing layer of basal laminae. The rectangular in- sert is shown at higher magnification in Figure 3. x 27,000.
At this magnification, the thick filaments (arrows) have a denser core and a more diffuse pe- ripheral relationship with the surrounding thinner filament+the precise limits of their thickness may be difficult to define in this preparation. The periph- eral cortical strip seen here beneath the plasma- lemma (dotted line delimiting the area between the plasmalemma and the less dense cytoplasm) shows a variable orientation. The 10-nm filaments are pre- sumed to be responsible for this increased density. x 98,000.
Fig. 3.

CANINE TRACHEAL MUSCLE IN VITRO 361
cells by TEM. Some of the lighter-staining cells when observed by TEM (Fig. 6) reveal disrup- tion of the plasmalemma; their staining prop- erties are due to presumed leaching of the cy- toplasmic contents into the surrounding medium and their replacement by extracellu- lar fluid (Somlyo and Somlyo, 1975). This is consistent with our observation that 5-30% of the isolated cells obtained after enzyme treat- ment are nonviable. Examination by TEM of the smooth muscle cells identified as dark, ser- rated cells by light microscopy (Fig. 7) reveals an extemely convoluted and pleated plasma membrane with numerous caveolae. Nuclei are darkly staining, mostly asymmetric, and very irregular in outline, with frequent clefts filled with cytoplasm. The margins of the cells are thrown into manifold ruffles and are the equivalent of the serrations seen by light mi- croscopy and indicative of considerable periph- eral activity of these membranes.
After 72 hours in culture, the cells flatten out and display morphologic characteristics of differentiated, contractile smooth muscle cells when viewed with phase-contrast optics; they are spindle or ribbon shaped with a dense cy- toplasm, possess perinuclear mitochondria, and display oval- to sausage-shaped nuclei (Chamley et al., 1977; Gimbrone and Cotran, 1975).
Confluency is usually reached between 10 and 14 days in culture depending upon the con- centration of the initial cell innoculum. Con- fluent monolayer cultures can be identified by the parallel alignment of cells.
After several days to a week or more in vitro, some of these cells appear amoeboid, and me- chanical attachment sites can be found be- tween cells possessing cytoplasm of variable density (Fig. 8). Inspection of the filamentous organization of one of these cells reveals dif- fuse densities interspersed within an area of thin filaments (Fig. 9). Concentration of other organelles, particularly ribosomes, impinges upon the filamentous area and tends to confuse the interpretation of the orientation of the fil- aments as revealed in this plane of section. In cultures of long duration (2 months), some of the morphological characteristics of smooth muscle are still visible. They are filamentous regions of the cytoplasm and multiple me- chanical attachment sites associated with an- choring densities (arrows) (Fig. 10). In this preparation, the lower cell has extensive cor- tical profiles of thin filaments; also present are some lipid droplets and a dilated cisterna of
endoplasmic reticulum (ER) filled with puta- tive proteinaceous precipitate. Its sister cell above possesses some of the same character- istics, in addition to a somewhat overall denser cytoplasm with more free ribosomes. At higher magnification (Fig. ll), another pair of cells show the nature of filamentous material in the cytoplasm, the presence of nexus formation (lower arrow), extracellular matrix (upper ar- rows), and regions of basal laminae (arrow head)-all features of smooth muscle.
The density of cytoplasm, dilated ER cister- nae with precipitate, abundant ribosomes, and filaments in the peripheral cytoplasm have been associated with characteristics of fibro- blasts. Some of these cells may represent a dedifferentiated population of smooth muscle cells more aptly termed myofibroblasts (Guber and Rudolph, 1978). We have been able to show the presence of thick (myosin) filaments in these cells by chemical (trypsin) and physical (mechanical dislodgement) manipulations of sufficient degree to cause the loss of integrity to the plasma membrane. Such a cell from an 8-day culture is shown in Figures 12 and 13. At higher magnification, the thick filaments are seen in transverse as well as longitudinal and oblique sections. In appropriately oriented areas of cells which have their plasmalemmas disrupted, transverse regions of thin filaments as well as thick filaments can be compared in the same field from these freshly prepared en- zymatically treated cells (Fig. 14). When other cells are treated with trypsin and gently dis- lodged from their culture dish after 16 days, no thick filaments are seen in the cytoplasm (Figs. 15, 16). On the basis of the above find- ings, we suggest that thick filaments in situ are extremely short and/or their conforma- tional and organizational state does not permit their morphological disposition in response to our usual dispersal and fixation methods (Small and Sobieszek, 1980).
To demonstrate the functional integrity of the contractile apparatus, a 6-day culture was photographed under phase-contrast condi- tions, exposed to carbachol, and photographed every 4 minutes for the next 20 minutes to assess the effect on the individual cells of the culture. In Figure 17 (control) and Figure 18 (12 minutes after carbachol administration) cells identified alphabetically (as well as oth- ers) can be seen to have shortened significantly during the time interval. Volume density was determined by overlaying a 5-mm-grid acetate sheet on photographs of control and carbachol-

362 BARRY P. AVNER, JUDI DELONGO, STEVE WILSON, AND A.J. LADMAN

CANINE TRACHEAL MUSCLE IN VITRO 363
Fig. 8. A TEM of part of an 11-day culture shows parts of four cells. The denser cell on the left is in contact with the lighter cell at the right by several mechanical attach- ment sites (m.a.). The lighter cell has a greater amount of distended granular endoplasmic reticulum whereas the cy- toplasm of the darker cell is characterized by filamentous areas between areas of free ribosomes and those related to the endoplasmic reticulum. The retangular region is shown in Figure 9. x 8,300.
Fig. 4. Low-magnification light micrograph of enzymat- ically treated tracheal smooth muscle sectioned after being formed into a pellet by mild centrifugation. Cells are dis- tributed either singly or into small aggregates. The rectan- gular area is shown at higher magnification in Figure 5. Stained with Toluidine blue. x 45.
Fig. 5. A light micrograph of cells derived from enzy- matically treated dissected tracheal smooth muscle. The darkly staining cells with highly serrated margins are smooth muscle cells (s.m.c.). Lighter staining cells (d.c.1 also appear in the field, as do red blood cells (r.b.c.1. Stained with Toluidine blue. x 190.
Fig. 9. The fibrillar area of the cytoplasm shown in the rectangle in Figure 8 consists of thin filaments and some densities (d) associated with these areas. Specific thicker myosin filaments cannot be distinguished with assurance. Such dispositions are also common to fibroblasts. x 34,400.
Fig. 6. Low-magnification transmission electron micro- graph of cellular elements as seen in Figures 4 and 5. The darkly staining cells (s.m.c.) are profiles of smooth muscle cells. The light staining cell (d.c.) appears not to be well fixed and may represent a smooth muscle cell which has been disturbed during the processing. The hemoglobin of a red blood cell (r.b.c.) is also seen in this preparation. x 4,400.
Fig. 7. At higher magnification, the multiple foldings and pleating of the plasma membrane with its related cav- eolae are consistent with the interpretation of considerable surface activity of this smooth muscle cell under these con- ditions of culture preparation. x 23,400. Calibration bar, 1 km.

364 BARRY P. AVNER, JUDI DELONGO, STEVE WILSON, AND A.J. LADMAN

CANINE TRACHEAL MUSCLE IN VITRO 365
treated cells and those cells intercepting grid corners were scored. There was a reduction of 30% between the scores of control and treated culture specimens, indicating a net change of cytoplasmic density due to cellular contrac- tion.
Cultures between 2 and 8 days of age are quite responsive to cholinergic stimulation as evidenced by the numbers of cells undergoing shortening. Although cultures older than 8-9 days have not been systematically examined for their responsiveness to cholinergic stimu- lation, in the few cultures tested we noted a diminution in responsiveness with time be- yond 9 days. HLM-15 and IMR-90 fibroblasts when examined under conditions comparable to these smooth muscle cultures did not con- tract when exposed to carbachol.
Mitosis was examined in selected thick sec- tions of samples as a function of age and these data are presented in Table 1. Mitotic rate di- minishes markedly after ll days in culture or when the culture becomes confluent. The mi- totic index between days 5 and ll varies within the limits of 3.2 and 4.8.
DISCUSSION
The overall aim of this study, of which this is the initial stage, is to develop a relatively simple and reproducible in vitro system of cul- turing airway smooth muscle cells that will permit the evaluation of the mechanisms of beta-receptor activation and desensitization. We noted earlier that many studies of smooth
muscle cultures from vascular and enteric sites have appeared; however, since the tissues from which these muscle cells have been derived are relatively unresponsive to the beta-2 receptor stimulant drugs used to treat bronchospasm, we have chosen to work with airway smooth muscle to realize our aim.
Perhaps some brief background would be useful at this point to highlight the signifi- cance of the intended studies which will em- ploy the culture methods described. Until re- cently (Lyons, 1976), very little was known about the mechanism of action of beta-receptor agonist drugs used in the treatment of diseases such as asthma. Inappropriate use of these drugs by patients, based upon the misconcep- tion that if a little is good, more is better, and/ or the marketing of high-potency drug prod- ucts, have been implicated in the rise of mor- tality among asthmatics (Keighley, 1966; Speizer and Doll, 1968a,b; Van Metre, 1969; Reisman, 1970; Heaf, 1970). It has been sug- gested that the beta-agonist bronchodilators may have contributed to this effect through a “desensitization” of the beta-receptors. Desen- sitization is a change in a cell or tissue re- sponse to a drug or agent in which the drug or agent has a negative feedback effect on the receptor system. In the case of tracheal smooth muscle, desensitization with beta-agonist drugs would be identified by a reduced relax- ation to increasing concentrations of admin- istered drug. This phenomenon of decreased sensitivity to beta-agonist bronchodilators is
TABLE 1. Mitosis in tracheal smooth muscle cultures
Age of culture MitosesilOO cells # of cells counted
5 3.2 462
1.0 (Confluent culture) 300 9 0.0 (Confluent culture) 300
11 3.6 600 16 0.04 (Confluent culture) 1,250 36 0.0 (Confluent culture) 300 60 0.0 (Confluent culture) 175
8 4.8 (Explant) 900
Fig. 10. After 2 months of culture, structural character- istics of smooth muscle cell cytoplasm are still present. These consist principally of mechanical attachment sites (arrows) and peripheral distribution of thin filaments. The dilated cisternae of the endoplasmic reticulum, the locali- zation of cytoplasmic fat droplets (0, and general variation of the quality of fixation suggest that these cells may be in a state of lowered responsiveness than much younger cul- tures. Cultures of this age were not tested for contractile competence. N = nucleus. x 10,600.
Fig. 11. After 2 months of culture, the nature of some structural appearances of smooth muscle are evident in this higher-magnification view of the peripheral parts of two cells. The extracellular matrix appears well preserved a t the top of the figure (upper arrows), as does the nexus for- mation (lower arrow) between these two cells. Basal laminae (arrowhead) are also easily identified. The thin filamentous nature of the cytoplasm is also shown to advantage. x 70,000.’

366 BARRY P. AVNER, JUDI DELONGO, STEVE WILSON, AND A.J. LADMAN

CANINE TRACHEAL MUSCLE IN VITRO 367
controversial with respect to our understand- ing of its mechanisms. Beta-receptor desensi- tization has been demonstrated as a result of in vivo and in vitro treatments in rat, guinea pig, dog, and human airway smooth muscle preparations (Lin et al., 1977; Avner et al., 1977; Avner and Noland, 1978; Avner and Jenne, 1977; Jenne et al., 1977; Nelson et al., 1977; Stephen et al., 1980; Avner et al., 1980.) Desensitization of airway smooth muscle is associated with a decrease of beta-receptor af- finity (i.e., the strength of binding of a drug to a membrane site) without an apparent de- crease in beta-receptor number (the quantity of sites represented on the surface membrane being investigated which, when activated, pro- duces maximum relaxation) (Lin et al., 1977; Avner and Noland, 1978). Thus the thrust of our inquiry will be to localize these receptor sites and to determine how these two factors, receptor affinity andlor number, are operating in given circumstances and how these factors can be influenced by assorted treatments with pharmacologic agents.
The method as presented here shows that the structural integrity of the contractile mechanism may display several forms, de- pending upon the age of the culture and the mode of treatment and processing. That these
Fig. 12. A cell from an 8-day culture treated with en- zymes and mechanically insulted to destroy its plasmalem- ma1 integrity. The rectangle defines a field seen in Figure 13. x 12.000,
Fig. 13. In this field, thick myosin filaments (arrows) oriented in various planes are displayed. At the top of the figure, tbey appear in transverse section-some are square or rhomboid in outline. At the left of the figure, these fila- ments appear longitudinally oblique in profile. Thin fila- ments are not well preserved in this field. x 33,500.
Fig. 14. The filamentous disposition within the cyto- plasm of another cell taken from a freshly prepared, enzy- matically treated tissue shows a small, delimited field of thin actin filaments transversely oriented in the upper part of the figure (upper arrow). In the lower part of the figure, thicker myosin filaments (lower arrow) appear in various planes. x 61,500.
Fig. 15. A cell from a 16-day culture which was not treated to violate the integrity of the plasmalemma. This smooth muscle cell is competent to contract and displays good cytoplasmic filaments and other organelles. x 5,500.
Fig. 16. At higher magnification, the rectangular fila- mentous area seen in Figure 15 is shown to better advan- tage. Although many thin filaments are present, the thick filaments cannot be clearly defined after this kind of prep- aratory treatment. x 20,000.
cells in culture do contract to a chemical stim- ulus can be verified and recorded quickly using phase-contrast optics. By the use of carbachol, we are able to distinguish functionally the con- tractility of the smooth muscle cell from the unresponsive fibroblast and myofibroblast (Gabbiani et al., 1972). A suitable technique to identify and record the degree of relaxation of previously contracted cells is in the process of being developed.
The in situ disposition of both thick and thin filaments, the presumed vehicle for contractile behavior in this tissue, as well as the 10-nm filaments that relate to cytoskeletal support, have not been rigorously examined because of the nature of culture conditions. These include a lack of polarity, a relative heterogeneity of the contractile response in this cellular popu- lation, and differences of degree of dediffer- entiation depending upon the time in culture. The reproducibility of our results with a va- riety of drugs, only one of which we have re- ported here, carbachol, indicates that cultures between 2 and 8 days of age are optimal for the purposes of test vehicles.
The morphological features of these cultures up to 16 days appear to be consistent with vig- orous, healthy smooth muscle cells. The pe- ripheral distribution and groupings of thin fil- aments, diffuse densities within the arrays of thin filaments, anchoring densities on the plasma membrane, caveolae, nexus formation, multiple attachment sites, and related basal laminae to plasmalemma all have been ob- served and described herein. Demonstration of thick filaments (presumably myosin) needs special treatment to permit good intracellular hat ion of these proteins. Evidence of intra- cellular localization and functional capacity of thick filaments after 16 days in culture has not been obtained.
In this regard, Rifas et al. (1979) were unable to show the presence of thick myosin filaments in their uterine smooth muscle cells up to 12 days of culture, whereas McLean et al. (1979) report the appearance of both thick and thin filaments in reaggregated vas deferens smooth muscle cells in culture after 9 days. It would be of interest to ascertain whether these thick filaments would be evident in monolayer cul- tures of vas deferens at the same time period as an expression of their conformational be- havior under culture conditions.
Somlyo et al. (1977) observed that profiles of thick filaments as viewed in stereo electron micrographs suggest cross-bridging relation- ships at different levels not only with thin fil- aments but also with presumable other ele-

368 BARRY P. AVNER, JUDI DELONGO, STEVE WILSON, AND A.J. LADMAN
Figs. 17,18. The same field of smooth muscle cells grow- ing in a 6-day culture. Figure 17 is the control and Figure 18 is 12 minutes after exposure of the culture to carbachol
MI. This cholinomimetic drug induces contraction in these cells. Those cells labeled “a” through %,” and others, appear to have been stimulated sufficiently for the observer
to discriminate a shortening of each cell in Figure 18 as compared to Figure 17. Use of this acetylcholine analogue helps to determine the smooth muscle nature of the culture (fibroblasts and myofibroblasts do not respond to this drug) x 100.

CANINE TRACHEAL MUSCLE IN VITRO 369
ments of the cytoplasm. Earlier, Somylo and coworkers (1973) described the thick filaments as irregular circular profiles, 15 nm in diam- eter, whose profiles in electron micrographs formed larger, ribbon-shaped elements repre- senting aggregation products of myosin ele- ments in muscles subjected to excessive shrinkage prior to or during fixation. Both the cross bridges as well as the aggregation prod- ucts cited above may be factors in conferring a square or fluted surface to the profiles of the myosin filaments depicted in our Figure 13.
In summary, the characteristics of our cells in culture might be restated from a compara- tive viewpoint. Structural entities which are shared by both fibroblasts and smooth muscle cells are nexus formations, thin actin fila- ments, and 10-nm filaments. Those elements which appear to be only in the province of smooth muscle structure are numerous me- chanical attachments, thick myosin filaments, and the contractile response to carbachol. The response to carbachol is fundamental to the unequivocal determination of the smooth mus- cle function of our cultured cells.
In both morphologic and pharmacologic cri- teria, the culture method described herein for airway smooth muscle should provide the means to broaden the scope of multidiscipli- nary investigation into this clinically relevant area.
ACKNOWLEDGMENTS
We are indebted to Drs. John A. Trotter and Robert 0. Kelley of this institution for their advice, comment, and criticism of this work during its several stages of progress, and for their substantive help during the preparation of the manuscript. We should like to thank Mr. Charles James of our Department of Microbi- ology for checking our cultures for evidence of Mycoplasma contamination. This work was supported by USPHS grants NIH-HL 20184 and RR-08139.
LITERATURE CITED Avner, B.P., and J.W. Jenne (1977) Comparison of in vitro
isoproterenol induced desensitization in human, dog, rat and rabbit respiratory smooth muscle. Am. Rev. Resp. Dis. (Suppl.), 115: 45 (Abstract).
Avner, B.P., and B.J. Noland (1978) In vivo desensitization to beta-receptor mediated bronchodilator drugs in the r a t Decreased beta-receptor affinity. J. Pharmacol. Exp. Ther., 207: 23-33.
Avner, B.P., B.J. Noland, and J.W. Jenne (1977) Desensi- tization of human bronchial smooth muscle to beta-recep- tor agonists. Proc. West. Pharmacol. Soc., 20: 25-31.
Avner, B.P., S. Wilson, and A.J. Ladman (1980) Beta re- ceptor desensitization in smooth muscle: Past, present and future. Proc. West. Pharmacol. SOC., 23: 477-483.
Bloom, W., and D.W. Fawcett (1975) A Textbook of Histol- ogy, 10th ed. W.B. Saunders Co., Philadelphia, pp. 291-294.
Chamley, J.H., G.R. Campbell, J.D. McConnell, and U. Groschel-Stewart (1977) Comparison of vascular smooth muscle cells from adult human, monkey and rabbit in primary culture and in subculture. Cell Tissue Res., 177: 503-522.
Chamley-Campbell, J., G.R. Campbell, and R. Ross (1979) The smooth muscle cell in culture. Physiol. Rev., 59: 1-61.
Copenhaver, W.M., D.E. Kelly, and R.L. Wood (1978) Bai- ley’g Textbook of Histology. 17th ed. The Williams and Wilkins Co., Baltimore, pp. 240-247.
Gabbiani, G., B.J. Hirschel, G. Ryan, P.R. Statkov, and G. Majno (1972) Granulation tissue as a contractile organ. A study of structure and function. J. Exp. Med., 135: 719-734.
Gauthier, G.F. (1977) In: Histology. 4th ed. L. Weiss and R.O. Greep, eds. McGraw Hill Book Co., New York, pp. 277-280.
Gimbrone, M.A., Jr., and R.S. Cotran (1975) Human vas- cular smooth muscle in culture. Growth and Ultrastruc- ture. Lab. Invest., 33: 16-27.
Guber, S., and R. Rudolph (1978) The myofibroblast. Surg. Gynecol. Obstet. 146: 641-649.
Hayflick, L. (1965) Tissue cultures of mycoplasmas. Tex. Rep. Biol. Med. (Suppl.), 23: 285-303.
Heaf, P.J.D. (1970) Deaths in asthma: A therapeutic mis- adventure? Br. Med. Bull., 26: 245-247.
Ives, H.E., G.S. Schultz, R.E. Galardy, and J.D. Jamieson (1978) Preparation of functional smooth muscle cells from rabbit aorta. J. Exp. Med., 148: 1400-1413.
Jenne, J.W., T.W. Chick, R.D. Strickland, and F.J. Wall (1977) Subsensitivity of beta response during therapy with a long-acting beta-2 preparation. J. Allergy Clin. Immunol., 59: 383-390.
Keighley, J.F. (1966) Iatrogenic asthma associated with ad- renergic aerosols. Ann. Int. Med., 65: 985-995.
Lewis, M.R. (1924) Spontaneous rhythmical contraction of the muscles of the bronchial tubes and air sacs of the chick embryo. Am. J. Physiol., 69: 385-388.
Lin, C-S., L. Hunvitz, J.W. Jenne, and B.P. Avner (1977) Mechanism of isoproterenol-induced desensitization of tracheal smooth muscle. J. Pharmacol. Exp. Ther., 203: 12-22.
Lyons, H.A. (1976) Bronchodilator adrenergic therapy in bronchial asthma mechanisms and therapeutics. In: Bron- chial Asthma, Mechanisms and Therapeutics. E.B. Weiss and M.S. Segal, eds. Little Brown and Co., Boston, pp. 727-738.
McLean, M.J., A. Pelleg, and N. Sperelakis (1979) Electro- physiological recordings from spontaneously contracting reaggregates of cultured smooth muscle cells from guinea pig vas deferens. J. Cell Biol., 80: 539-552.
Nelson, H.S., D. Raine, Jr., H.C. Doner, and W.C. Posey (1977) Subsensitivity to the bronchodilator action of al- buterol produced by chronic administration. Am. Rev. Resp. Dis., 116: 871478.
Reisman, R.E. (1970) Asthma induced by adrenergic aero- sols. J. Allerg., 46: 162-177.
Rifas, L., J. Fant, M.H. Makman, and S. Seifter (1979) The characterization of human uterine smooth muscle cells in culture. Cell Tissue Res., 196: 385-395.
Ross, R., and B. Kariya (1980) Morphogenesis of vascular smooth muscle in atherosclerosis and cell culture. In: Handbook of Physiology. Sect. 2, The Cardiovascular Sys- tem, Vol. 11, Vascular Smooth Muscle. D.F. Bohr, A.P. Somlyo, and H.V. Sparks, Jr., eds. Am. Physiol. Soc., Be- thesda, pp. 69-91.
&hopper, W. (1935) Embryonales und erwachsenes Lun- gengewebe von Meerschweinschen und Huhn in der Kul- tur mit Zeitraffenbeobachtungen on Flimmerepithel sog.

370 BARRY P. AVNER, JUDI DELONGO, STEVE WILSON, AND A.J. LADMAN
Alveolarphagocyten und von Kontraktionen der Bron- chialmuskulatur. Virchows. Arch. Pathol. Anat. Physiol., 295: 623-624.
Small, J.V., and A. Sobieszek (1980) The contractile appa- ratus of smooth muscle. Int. Rev. Cytol., 64: 241-306.
Somlyo, A.P., C.E. Devine, A.V. Somlyo, and R.V. Rice (1973) Filament organization in vertebrate smooth mus- cle. Philos. Trans. R. Soc. Lond. (Biol.), 265: 223-229.
Somlyo, A.P., and A.V. Somlyo (1975) Ultrastructure of smooth muscle. In: Methods in Pharmocology. Vol. 3, Smooth Muscle. E.E. Daniel and D.M. Paton, eds. Plenum Press, New York, pp. 3-46.
Somlyo, A.V., F.T. Ashton, L.F. Lemanski, J . Vallieres, and A.P. Scmlyo (1977) Filament organization and dense bod- ies in vertebrate smooth muscle. In: The Biochemistry of Smooth Muscle. N.L. Stephens, ed. University Park Press,
Baltimore, pp. 445-472. Speizer, F.E., and R. Doll (1968a) Observations on recent
increase in mortality from asthma. Br. Med. J., I: 335-339.
Speizer, F.E., and R. Doll (1968b) Investigation into use of drugs preceding death from asthma. Br. Med. J., I: 339-343.
Stephen, W.C., T.W. Chick, B.P. Avner, and J.W. Jenne (1980) Tachyphylaxis to inhaled isoproterenol and the effect of methylprednisolone in dogs. J. Allergy Clin. Im- munol., 65: 105-109.
Tolnai, S. (1975) A method for viable cell count. Tissue Cult. Assoc. Manual, 1; 37-38.
Van Metre, T.E. (1969) Adverse effects of inhalation of ex- cessive amounts of nebulized isoproterenol in status asth- maticus. J . Allergy, 43: 101-113.
Copyright © 2022 FDOKUMEN




















