
Topoisomerase I and RecQL1 Function in Epstein-Barr Virus Lytic Reactivation

Induction of Lytic Enzymes by the Interaction
QGu
Fungal Genetics and Biology 29, 145–151 (2000)doi:10.1006/fgbi.2000.1196, available online at http://www.idealibrary.com on
of Ustilago maydis with Zea mays Tissues
C. Cano-Canchola,* L. Acevedo,* P. Ponce-Noyola,* A. Flores-Martınez,*A. Flores-Carreon,* ,† and C. A. Leal-Morales* ,1
*Instituto de Investigacion en Biologıa Experimental (IIBE), Facultad de Quımica, Universidad de Guanajuato,Guanajuato, Mexico; and †Departamento de Genetica y Biologıa Molecular, Centro de Investigacion y EstudiosAvanzados (CINVESTAV), Instituto Politecnico Nacional (IPN), Mexico DF, Mexico
Accepted for publication April 11, 2000
Cano-Canchola, C., Acevedo, L., Ponce-Noyola, P., Expression of xylanase activity coincided with the last
Flores-Martınez, A., Flores-Carreon, A., and Leal-Mo-rales, C. A. 2000. Induction of lytic enzymes by theinteraction of Ustilago maydis with Zea mays tissues.Fungal Genetics and Biology 29, 145–151. The non-pathogenic (FB-2) and pathogenic (FB-D12) strains ofUstilago maydis were grown in medium supplementedwith different carbon sources including monosaccha-rides, polysaccharides, and plant tissues. Both strainswere able to grow on all substrates, with doublingtimes varying from 2 to 25 h depending on the carbonsource. Plant tissues supplied as carbon source in-duced lytic enzymes differentially; pectate lyase andcellulase activities were induced preferentially by api-cal stem meristem in strain FB-D12, whereas leavespreferentially induced xylanase and cellulase activitiesin strain FB2. Stems induced polygalacturonase activ-ity in both strains. All enzyme activities, except cellu-lase in the FB-D12 strain, were detected at a low levelwhen U. maydis was grown on glucose. In planta,chlorosis and production of teliospores were paral-leled by an increase in pectate lyase activity. Antho-cyanin production and formation of galls and telios-pores correlated with polygalacturonase expressionwhereas cellulase activity increased only during thestage of anthocyanin production and gall formation.1 To whom correspondence should be addressed at IIBE, Facultad deuımica, Universidad de Guanajuato, Apartado Postal #187, Guanajuato,to., Mexico, CP 36000. Fax: (473) 2 43 02. E-mail: [email protected].
1087-1845/00 $35.00Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved. 145
stage of teliospore formation. © 2000 Academic Press
Index Descriptors: tissue assay; plant pathology; Zeamays.
Plant pathogens produce a number of extracellular en-zymes capable of degrading plant cell wall components. Ithas been postulated that the lytic enzymes of the pathogenare involved in a variety of processes such as penetration,growth, and development; once the pathogen is inside theplant it obtains nutrients from wall polymers. Thus, tounderstand pathogenesis, it is important to establish therole of the different depolymerases, such as pectinases,cellulases, and xylanases. Due to the structural complexityof the plant cell wall, whose polymers differ considerablyin accessibility, nutrient suitability, and resistance to deg-radation, this role needs to be studied at different stages ofthe infection process (Walton, 1994).
Ustilago maydis, a Basidiomycete fungus, causes cornsmut disease. This disease is characterized by the forma-tion of tumors (galls) on any aerial plant tissue. It has beenobserved that the pathogen colonizes mainly the embry-onic tissue such as the apical meristem (Christensen,1963). The fungus exhibits three types of cells: the non-pathogenic haploid cells with a yeast-like morphology isone. A second type of cells is the filamentous dikaryon thatresults from the mating of two compatible haploids havingdifferent a and b alleles; these cells are parasitic anddifferentiate within tumors. Teliospores, the third cell

type, derive from the above through karyogamy and un-
cpmc1kc
1mp
fso(dcoepdpeua
birchwood, X-0502) (all from Sigma Chemical Co., St.
o
i
146 Cano-Canchola et al.
dergo meiosis to produce haploid sporidia (Christensen,1963; Puhalla, 1970; Banuett and Herskowitz, 1989;Kronstad and Leong, 1990; Banuett, 1991; Gillissen et al.,1992; Spelling et al., 1994). There are other stable diploidells that can be obtained either naturally or after proto-last fusion of compatible haploid sporidia. The latterethod has allowed to elucidate how the mating type loci
ontribute to mating and pathogenicity (Christensen,963; Banuett and Herskowitz, 1989). The resulting cells,nown as solopathogenic strains, are able by themselves toomplete their life and pathogenic cycles in planta. More-
over, the major factors that regulate these cycles havebeen identified at the molecular level (Kronstad and Le-ong, 1989; Schulz et al., 1990; Bolker et al., 1992; Cooper,983). These properties make U. maydis an excellentodel to study the biochemical and molecular bases of
athogenicity.Despite the advantages of U. maydis as a model to study
ungal–plant interactions, little is known about the expres-ion of lytic enzymes and their potential role in the devel-pment of plant symptoms. It is worth noting that Cooper1983) suggested that biotroph organisms, such as U. may-is, produce small amounts of cell wall lytic enzymes. Aellulase gene from U. maydis, whose expression is specificf the filamentous growth, has been cloned (Schauweckert al., 1995). We report here the induction of pectate lyase,olygalacturonase, cellulase, and xylanase activities usingifferent substrates and plant tissues from both a non-athogenic and a solopathogenic strain. Their sequentialxpression during the infective process that follows inoc-lation of the parasite into the apical meristematic tissue islso reported.
MATERIALS AND METHODS
Strains and Growth Curves
The U. maydis nonpathogenic haploid strain FB2 andthe pathogenic diploid strain FB-D12 (Banuett and Her-skowitz, 1989) were obtained from Professor F. Banuett(University of California, San Francisco).
The synthetic culture medium described by Bartnicki-Garcıa and Nickerson (1962) supplemented with eitherone of the following carbon sources was used to propagateU. maydis: 2% glucose, 1% d-galacturonic acid (sodiumsalt), 1% xylose, 1% polygalacturonic acid (from orange,sodium salt, P-1879), 1% CM-cellulose, 1% xylan (from
Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.
Louis, MO), or 1% freeze-dried plant tissue. The pH ofthe medium was adjusted to 4.5 with HCl. Tissues weredissected from 10-day-old corn seedlings. Leaf tissue wasobtained from a 7-cm leaf segment cut off from the tiptoward the base, and stem tissue was obtained from a3-cm-long segment between 3 and 6 cm above the soillevel. To obtain a fraction highly enriched with apical stemmeristem, only four of the layers surrounding the 3-cmsegment above the soil level were cut off. Tissues were cutinto small pieces, lyophilized, and ground to a fine powderin liquid nitrogen with a mortar and pestle. Sporidia werekept on a petri dish in solid minimum medium and whenneeded a loop was inoculated in liquid medium to obtaina preinoculum. Erlenmeyer flasks (250 ml) containing 50ml of the medium supplemented with glucose were inoc-ulated with the preinoculum to a final absorbance of0.03–0.05 at 595 nm (Beckman DU-650) and incubated at28°C in a rotary shaker bath at 100–120 rpm. Sampleswere taken to determine absorbance at 595 nm at differenttime points. Absorbance of culture medium supplementedwith plant tissue was obtained by subtracting the value forthe noninoculated medium. Fungal growth under the var-ious conditions used in this work was determined in atleast three independent experiments.
Inoculation of Plants and Assayf Pathogenicity
For symptom assessment and induction of lytic enzymesduring the pathogenic process, 10-day-old corn seedlings(Golden Bantam variety) were grown under environmen-tal conditions, where temperature fluctuated from 10 to30°C. At least 150 corn seedlings were infected as de-scribed by Puhalla (1968) and inspected daily for theappearance of symptoms in at least four independentexperiments. The pathogenic strain FB-D12 sequentiallyinduced chlorosis, anthocyanin biosynthesis, and tumorson plants over a period of 3 to 20 days.
Induction of Lytic Enzymes in Vitro andn Planta
Lytic enzymes were induced in minimal medium con-taining different carbon sources (see above). Cultureswere harvested at the beginning of the stationary growthphase by filtration through Whatman No. 1 filter paper.Filtrates were dialyzed overnight at 4°C against 50 mMsodium acetate buffer, pH 5.2, and freeze-dried. Prior toassay, samples were resuspended in 2 ml of water.

To determine the lytic activities secreted by U. maydis
a
Tr4aa
at 30°C, activity was determined as described above for
tsmamtacu
u
l
147Lytic Enzymes of Ustilago maydis
during infection, plants were inoculated as describedabove, either with FB2 or FB-D12 strains or with water.Plants were inspected for symptoms, and tissues showingcomparable levels of damage were collected from severalplants each day and lyophilized. For comparison, levels oflytic activities in healthy tissue from infected plants or inequivalent tissues of mock-inoculated plants were alsomeasured. To extract enzymes, 1.0 g of dried tissue wasground as described above and the resulting powder wasresuspended in 4 ml of cold deionized water containing 1mM PMSF and homogenized with a tissue grinder. Thematerial was centrifuged at 12,100g (Rmax) for 20 min, thesediment was reextracted four times under the same con-ditions, and the supernatants were combined and freeze-dried. Prior to assay, samples were resuspended in 2 ml ofwater.
Protein was measured by the Bradford method (Sedmakand Grossberg, 1977) using BSA as standard.
Enzyme Assays
In all assays, reaction mixtures containing 20–50 mgprotein in a final volume of 3 ml were incubated and0.5-ml aliquots were taken at different times to measurereaction products as described below.
Polygalacturonase (PG)2 activity was assayed at 30°C in50 mM acetate buffer, pH 5.3, containing 0.3% (w/v)polygalacturonic acid and 7.5 mM EDTA (Collmer et al.,1988). The amount of reducing groups was determined bythe method of Nelson-Somogyi using d-galacturonic acids standard (Somogyi, 1952).
Pectate lyase (PL) activity was assayed in 100 mMris–HCl buffer, pH 8.5, containing 0.24% polygalactu-
onic acid and 1 mM calcium chloride. The formation of,5-unsaturated uronic acids was followed by absorbancet 232 nm (Beckman DU-650). An increase of 1.73 units ofbsorbance corresponded to the formation of 1 mmol of
unsaturated uronic acid per minute at 30°C (Ching-Hsinget al., 1988).
Cellulase (CEL) activity was assayed in 50 mM acetatebuffer, pH 5.5, containing 0.5% (w/v) CM-cellulose. Ac-tivity was determined after incubation at 30°C as de-scribed for PG, using d-glucose as standard (Amadioha,1993).
Xylanase (XYL) activity was assayed in 50 mM acetatebuffer, pH 5.0, containing 1% (w/v) xylan. After incubation
2 Abbreviations used: CEL, cellulase; PL, pectate lyase; PG, polyga-acturonase; XYL, xylanase.
PG, using d-xylose as standard (Apel et al., 1993).PG activity in plant extracts was assayed by measuring
he release of uronic acid by the carbazole reaction, es-entially as described above for PG. Aliquots of up to 0.5l of the assay mixture were mixed with 6 ml of sulfuric
cid and the tubes were immersed in boiling water for 20in. After cooling, 0.2 ml of carbazole [0.1% (w/v) solu-
ion in ethanol] was added and absorbance was measuredt 530 nm (Bitter and Muir, 1962). An assay mixture,ontaining all the ingredients, was boiled for 3 min andsed as a blank.For all hydrolases, enzyme activity was expressed in
nits. One unit is equal to 1 mmol of the correspondingsugar released after 1 min of incubation under the condi-tions described above. Specific activity was referred to1 mg of protein.
RESULTS
Utilization of Different Carbon Sources
Both FB2 and FB-D12 strains of U. maydis were able togrow on all carbon sources, with glucose and apical mer-istem tissue being the most efficient nutrients. On thesesubstrates, doubling times of 1.9 to 2.2 h (Figs. 1A, 1B, 1E,and 1F) were obtained for both strains compared to thoseof 3 to 5 h observed with xylose or xylan (Figs. 1A–1D). Avalue of 2.5 h was observed for both strains grown onCM-cellulose (Figs. 1C and 1D). Doubling times on ga-lacturonic acid were 25.2 and 21.6 h for strains FB2 andFB-D12, respectively (Figs. 1A and 1B). These resultsshow that both strains can degrade several polysaccharidesand use the corresponding released sugars as a carbonsource. In this point it is important to consider that growthof these strains can be influenced also by componentsother than polysaccharides present in the plant tissues,such as proteins, amino acids, monosaccharides, organicacids, etc.
Production of Lytic Enzymes
To evaluate the induction of lytic enzymes, the funguswas grown on different carbon sources and extracellularenzyme activities were measured early in the stationaryphase.
Pectinases. The induction of PL and PG activities ispresented in Table 1. Both strains showed basal levels of
Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.

stem, which brought about increases of 184- and 232-fold
t
a
t
148 Cano-Canchola et al.
PL activity when they were grown on glucose, correspond-ing to 0.04 and 0.08 U for FB2 and FB-D12, respectively.These activities increased approximately 2-fold over thebasal level when grown on galacturonic acid. With polyga-lacturonic acid, values were either comparable or approx-imately 3-fold lower than the basal levels for strains FB2and FB-D12, respectively. The highest levels of induction,corresponding to 2.3- and 14-fold over the basal level forFB2 and FB-D12 strains, respectively, were obtained withmeristematic tissue.
In the presence of glucose, PG activity was detected atbasal levels. The best inducer of this activity was maize
FIG. 1. Growth of U. maydis FB2 and FB-D12 strains on differentcarbon sources. A series of 250-ml flasks containing 50 ml of minimalculture medium supplemented with 2% glucose, 1% xylose, 1%d-galacturonic acid, 1% CM-cellulose, 1% xylan, 1% polygalacturonicacid, or 1% powdered plant tissue were inoculated with sporidia at a finalabsorbance of 0.03. Samples were withdrawn at various times to deter-mine absorbance at 595 nm. When plant tissue was used as a carbonsource, values were corrected by subtracting the absorbance reading ofthe noninoculated medium. (A, C, and E) Growth of haploid FB2 strain;(B, D, and F) growth of diploid FB-D12 strain. The experiments wererepeated at least three times; the graphs are from a representativeexperiment. (A, B) Glucose, F; xylose, Œ; galacturonic acid, ■. (C, D)CM-cellulose, E; xylan, ‚; polygalacturonic acid, h. (E, F) Stem, ƒ;leaves, 1; apical meristem, .
Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.
in the nonpathogenic and pathogenic strains, respectively,although meristematic tissue was also an effective inducerwith corresponding increases of 149- and 82-fold.
Cellulase. CEL activity of both strains showed basallevels when they were grown on glucose, corresponding to0.02 and 1.28 U for FB2 and FB-D12, respectively. Fol-lowing growth in the presence of CM-cellulose (Table 2),activity increased 467- and 11-fold for the nonpathogenicand pathogenic strains, respectively. The plant tissues thatwere most effective in inducing cellulase activity were notthe same for the two strains; thus, whereas the nonpatho-genic strain was greatly stimulated by leaf tissue (190-fold), the best inducer for the pathogenic strain was apicalmeristem tissue (5.8-fold) (Table 2).
Xylanase. XYL activity was the least induced of allenzymes tested, since only a 5.0-fold increase was ob-tained for the nonpathogenic strain grown on leaf tissue.As in the case of pectinases, the highest levels of XYLactivity were not induced by the corresponding pure en-zyme substrate (Table 2), but instead by leaf tissue, cor-responding to 5.0- and 4.0-fold over the basal level for theFB2 and FB-D12 strains, respectively (Table 2).
Expression of Lytic Enzymes duringhe Infective Cycle of U. maydis
Young corn seedlings are very susceptible to infectionby U. maydis. Accordingly, when a sporidial suspension ofthe solopathogenic strain was injected in the proximity ofthe apical meristem almost 100% of the plants developedthe disease. The tissue around the inoculation site became
TABLE 1
Induction of Pectinolytic Enzymes by Different Carbon Sourcesa
Carbon source
Pectate lyase specificactivityb
Polygalacturonasespecific activityb
FB2 FB-D12 FB2 FB-D12
Glc 0.04 (1.0)c 0.08 (1.0) 0.01 (1.0) 0.02 (1.0)GalA 0.09 (2.3) 0.16 (2.0) 0.03 (3.0) 0.03 (1.5)PGA 0.04 (1.0) 0.03 (0.4) 0.02 (2.0) 0.07 (3.5)Leave 0.08 (2.0) 0.23 (2.5) 0.03 (3.0) 0.12 (6.0)Stem 0.08 (2.0) 0.10 (1.3) 1.84 (184) 4.63 (232)Meristem 0.11 (0.8) 1.09 (14) 1.49 (149) 1.63 (82)
a Results are averages of at least three independent experiments withstandard deviation no greater than 10%.b Specific activity 5 1 mmol sugar released min21 mg protein21.c Numbers in parentheses correspond to activity values relative to
hose obtained in filtrates from glucose-grown cultures.
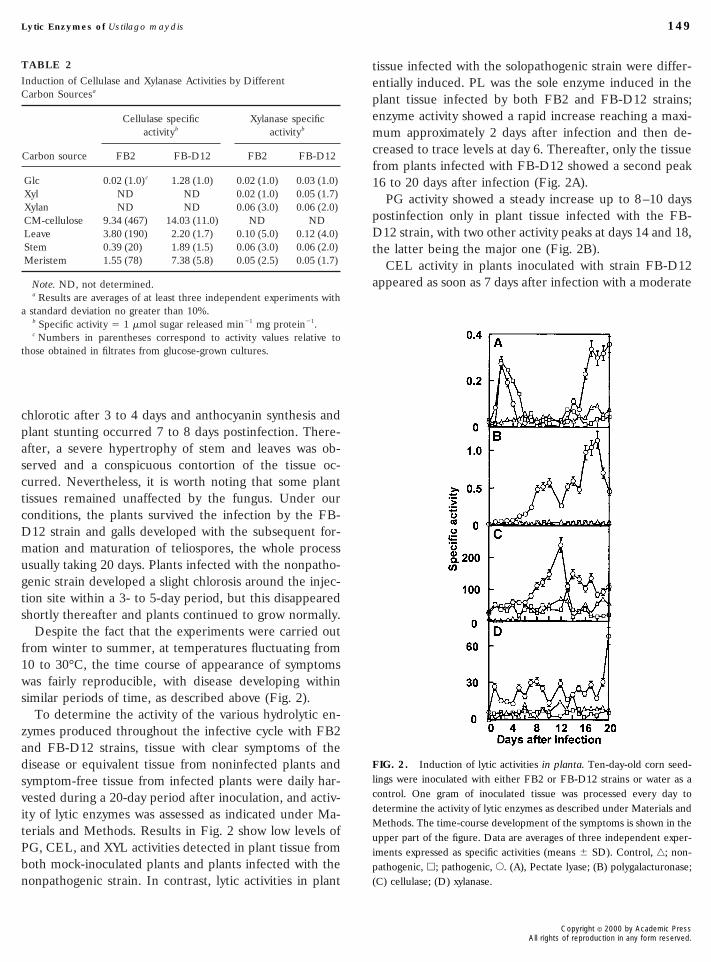
tissue infected with the solopathogenic strain were differ-TABLE 2
a
t
149Lytic Enzymes of Ustilago maydis
chlorotic after 3 to 4 days and anthocyanin synthesis andplant stunting occurred 7 to 8 days postinfection. There-after, a severe hypertrophy of stem and leaves was ob-served and a conspicuous contortion of the tissue oc-curred. Nevertheless, it is worth noting that some planttissues remained unaffected by the fungus. Under ourconditions, the plants survived the infection by the FB-D12 strain and galls developed with the subsequent for-mation and maturation of teliospores, the whole processusually taking 20 days. Plants infected with the nonpatho-genic strain developed a slight chlorosis around the injec-tion site within a 3- to 5-day period, but this disappearedshortly thereafter and plants continued to grow normally.
Despite the fact that the experiments were carried outfrom winter to summer, at temperatures fluctuating from10 to 30°C, the time course of appearance of symptomswas fairly reproducible, with disease developing withinsimilar periods of time, as described above (Fig. 2).
To determine the activity of the various hydrolytic en-zymes produced throughout the infective cycle with FB2and FB-D12 strains, tissue with clear symptoms of thedisease or equivalent tissue from noninfected plants andsymptom-free tissue from infected plants were daily har-vested during a 20-day period after inoculation, and activ-ity of lytic enzymes was assessed as indicated under Ma-terials and Methods. Results in Fig. 2 show low levels ofPG, CEL, and XYL activities detected in plant tissue fromboth mock-inoculated plants and plants infected with thenonpathogenic strain. In contrast, lytic activities in plant
Induction of Cellulase and Xylanase Activities by DifferentCarbon Sourcesa
Carbon source
Cellulase specificactivityb
Xylanase specificactivityb
FB2 FB-D12 FB2 FB-D12
Glc 0.02 (1.0)c 1.28 (1.0) 0.02 (1.0) 0.03 (1.0)Xyl ND ND 0.02 (1.0) 0.05 (1.7)Xylan ND ND 0.06 (3.0) 0.06 (2.0)CM-cellulose 9.34 (467) 14.03 (11.0) ND NDLeave 3.80 (190) 2.20 (1.7) 0.10 (5.0) 0.12 (4.0)Stem 0.39 (20) 1.89 (1.5) 0.06 (3.0) 0.06 (2.0)Meristem 1.55 (78) 7.38 (5.8) 0.05 (2.5) 0.05 (1.7)
Note. ND, not determined.a Results are averages of at least three independent experiments with
standard deviation no greater than 10%.b Specific activity 5 1 mmol sugar released min21 mg protein21.c Numbers in parentheses correspond to activity values relative to
hose obtained in filtrates from glucose-grown cultures.
entially induced. PL was the sole enzyme induced in theplant tissue infected by both FB2 and FB-D12 strains;enzyme activity showed a rapid increase reaching a maxi-mum approximately 2 days after infection and then de-creased to trace levels at day 6. Thereafter, only the tissuefrom plants infected with FB-D12 showed a second peak16 to 20 days after infection (Fig. 2A).
PG activity showed a steady increase up to 8–10 dayspostinfection only in plant tissue infected with the FB-D12 strain, with two other activity peaks at days 14 and 18,the latter being the major one (Fig. 2B).
CEL activity in plants inoculated with strain FB-D12appeared as soon as 7 days after infection with a moderate
FIG. 2. Induction of lytic activities in planta. Ten-day-old corn seed-lings were inoculated with either FB2 or FB-D12 strains or water as acontrol. One gram of inoculated tissue was processed every day todetermine the activity of lytic enzymes as described under Materials andMethods. The time-course development of the symptoms is shown in theupper part of the figure. Data are averages of three independent exper-iments expressed as specific activities (means 6 SD). Control, ‚; non-pathogenic, h; pathogenic, E. (A), Pectate lyase; (B) polygalacturonase;(C) cellulase; (D) xylanase.
Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.

increase throughout, reaching a maximum after 12 days; form polysaccharides of the plant cell wall into metaboliz-
IfiAfip
uettdphiwwteeocHgevopiiqic
150 Cano-Canchola et al.
thereafter, it decreased without returning to basal levels(Fig. 2C). Tissue from plants infected with strain FB-D12invariably showed a much higher CEL activity than thatfrom plants infected with the FB2 strain or that fromhealthy plants.
XYL activity was only slightly induced during infectionwith strain FB-D12 (17 to 22 U/mg protein), although itincreased to 60 U/mg protein 20 days postinfection (Fig.2D).
The sequence of symptoms, which was followed visuallyduring the infection process, was: First, chlorotic spotsappeared after 3–4 days and they seemed to correlate withthe increase in PL activity. Second, a clear evidence ofinfection was the production of anthocyanin and the stunt-ing of the infected plants observed after 7–8 days, whichcorrelated with the expression of PG activity. Third, otherclearly visible symptoms were tissue deformations in thosesites from which the galls eventually emerged, which oc-curred within 10–12 days postinfection coinciding withmaximum CEL activity. Teliospore production and matu-ration occurred later (16–20 days) and correlated with thesecond peak of PL and PG activities.
DISCUSSION
Several ultrastructural and cytochemical studies in thewhole plant have been performed to understand the modeof entry of biotrophic phytopathogenic fungi and the na-ture of the mechanisms involved in this process (Baka andLosel, 1992; Larous and Losel, 1993a,b). The dissolutionof the host cell wall around the point of entrance of afilamentous haustorium of rust fungi is often stronglysuggestive of localized wall-degrading enzymes (Larousand Losel, 1993b), which are believed to be importantfactors in the outcome of plant–fungus interaction. How-ever, little is known on the regulation of induction of theseenzymes during infection by obligatory biotrophic fungi.
It has been proposed that U. maydis invades the hostplant through any aerial meristematic tissue (Christensen,1963) and that the dikaryotic parasitic mycelium does notbecome systemic but spreads rapidly to the vicinity of theinfection site (Snetselaar and Mims, 1994). This allows thepathogen to obtain nutrients from the plant in order tosurvive during initial interaction, penetration, and coloni-zation, and all these processes often involve the degrada-tion of cell wall polymers. Here we demonstrate that bothnoninfective and infective strains of U. maydis can trans-
Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.
able carbon sources. Both strains secreted, though at vari-able extents, lytic activities to the extracellular mediumwith all tested substrates, including glucose, which in-duced low levels of PL, PG, CEL, and XYL activities.Interestingly, the highest induction of these activities wasobtained with different plant tissues, revealing some levelof specificity; accordingly, for both strains, meristem,stems, and leaf tissues preferentially induced PL, PG, andXYL activities, respectively. The best inducers of CELactivity were meristematic and leaf tissues for FB-D12 andFB2 strains, respectively. Tissue factors involved in induc-tion of these activities are thermostable and they can berelated to specific structural characteristics of cell wallpolymers, as has been proposed for the induction of Col-letotrichum lindemuthianum a-galactosidase, a hydrolasespecifically involved in pathogenicity (English et al., 1971).n addition, Schauwecker et al. (1995) isolated a U. maydislament-specific gene that encoded for a cellulase activity.mutation in this gene did not affect either the filament
ormation or the pathogenicity; even so, the role of induc-ble lytic activities in pathogenesis was determined inlanta after infection with Ustilago. We observed that
enzyme induction in planta is sequential and very likelynder complex regulation. However, it was possible tostablish the time-course expression during infection ando correlate the induction pattern with the development ofhe disease in the susceptible tissues. Our results show thaturing infection, lytic activities are expressed only in thelant area damaged by the fungus and not in the adjacentealthy tissue, thus supporting the role of these hydrolases
n the localized destruction of tissue. This is also consistentith the damage observed following inoculation of plantsith culture filtrates of the fungus grown on meristematic
issue (results not shown). Although restriction of lyticnzymes to infected areas and also the induction of thesenzymes by plant tissues strongly support their fungalrigin, further experiments will be necessary to fully dis-ard the plant as the source of some of these activities.owever, whatever their proceedence, i.e., plant or fun-
al, the impact of this communication on the role of lyticnzymes during the infective cycle of U. maydis remainsalid. Moreover, this work not only shows the complexityf the U. maydis–maize interaction, but also opens theossibility to dissect the network of regulatory factors
nvolved in the expression of these activities and their rolen the development of the symptoms, since they are se-uentially induced along the infection process. More stud-
es will be necessary to determine the effect of severalonditions on the induction of polysaccharide-degrading

enzymes by U. maydis grown in culture and to identify the
A
B
B
B
B
C
C
Collmer, A., Reid, L., and Mount, M. S. 1988. Assay methods for pectic
C
E
G
K
K
L
L
P
P
S
S
S
S
S
S
W
151Lytic Enzymes of Ustilago maydis
tissue factors involved in their expression.
ACKNOWLEDGMENTS
This work was supported in part by Grant 5358N-9408 fromCONACyT, Mexico. We express our gratitude to Dr. Everardo Lopez-Romero for helpful discussion and critical review of the manuscript.
REFERENCES
Amadioha, A. C. 1993. Production of cellulolytic enzymes by Rhizopusoryzae in culture and Rhizopus-infected tissue of potato tubers. My-cology 85: 574–578.
pel, P. C., Penaccione, D. G., Holden, F. R., and Walton, J. D. 1993.Cloning and targeted gene disruption of Xyl1, a b1-4 xylanase genefrom the maize pathogen Cochliobolus carbonum. Mol. Plant-MicrobeInteract. 6: 474–473.
Baka, Z. A. M., and Losel, D. M. 1992. Infection of host vascular tissueby Puccina punctiformis and Puccina lagenophorae. Physiol. Mol. PlantPathol. 40: 411–421.
Banuett, F. 1991. Identification of genes governing filamentous growthand tumor induction by plant pathogen Ustilago maydis. Proc. Natl.Acad. Sci. USA 88: 3922–3926.
anuett, F., and Herskowitz, I. 1989. Different a alleles of Ustilagomaydis are necessary for maintenance of filamentous growth but notfor meiosis. Proc. Natl. Acad. Sci. USA 86: 5878–5882.
artnicki-Garcıa, S., and Nickerson W. J. 1962. Nutrition, growth andmorphogenesis of Mucor rouxii. J. Bacteriol. 84: 841–858.
itter, T., and Muir, H. M. 1962. A modified uronic acid reaction. Anal.Biochem. 4: 330–333.
olker, M., Urban, M., and Kahmann, R. 1992. The a mating type locusof Ustilago maydis specifies cell signaling components. Cell 66: 441–450.
hristensen, J. J. 1963. Corn smut caused by Ustilago maydis. In Amer-ican Phytopathology Society Monograph, No. 2, pp. 1–41. Am. Phy-topathol. Soc., St. Paul, MN.
hing-Hsing, L., Hsueh-Yun, H., and Chatterjee, A. K. 1988. An extra-cellular pectate lyase is the pathogenicity factor of the soft-rottingbacterium Pseudomonas virideflava. Mol. Plant–Microbe Interact. 1:199–206.
enzymes. Methods Enzymol. 161: 329–335.ooper, R. M. 1983. The mechanism and significance of enzymaticdegradation of host cell wall by parasites. In Biochemical Plant Pathol-ogy (J. A. Callow, Ed.), pp. 101–136. Wiley, London.
nglish, P. D., Jurale, J. B., and Albersheim, P. 1971. Host–pathogeninteractions. II. Parameters affecting polysaccharide-degrading en-zyme secretion by Colletotrichum lindemuthianum grown in culture.Plant Physiol. 47: 1–6.illissen, B., Bergemann, J., Sandmann, C., Schroeer, B., Bolker, M.,and Kahmann, R. 1992. A two-component regulatory system for self/non-self recognition in Ustilago maydis. Cell 68: 647–657.
ronstad, J. W., and Leong, S. A. 1989. Isolation of two alleles of the blocus of Ustilago maydis. Proc. Natl. Acad. Sci. USA 86: 978–982.
ronstad, J. W., and Leong, S. A. 1990. The b mating-type locus ofUstilago maydis contains variable and constant regions. Genes Dev. 4:1384–1359.
arous, L., and Losel, D. M. 1993a. Vascular infection by Puccinamenthae and other rust fungi. Mycol. Res. 97: 409–414.
arous, L., and Losel, D. M. 1993b. Strategies of pathogenicity inmonokaryotic and dikaryotic phases of rust fungi, with special refer-ence to vascular infection. Mycol. Res. 97: 415–420.
uhalla, J. E. 1968. Compatibility reactions on solid medium and interstrain inhibition in Ustilago maydis. Genetics 60: 461–474.
uhalla, J. E. 1970. Genetic studies on the incompatibility locus ofUstilago maydis. Genet. Res. 16: 229–232.
chauwecker, F., Wanner, G., and Kahmann, R. 1995. Filament-specificexpression of a cellulase gene in the dimorphic fungus Ustilago may-dis. Biol. Chem. Hoppe-Seyler 376: 617–625.
chulz, B., Banuett, F., Dahl, M., Schlesinger, R., Schafer, W., Martin,T., Herskowitz, I., and Kahmann, R. 1990. The b alleles of Ustilagomaydis, whose combinations program pathogenic development, codefor polypeptides containing a homeodomain-related motif. Cell 60:295–306.
edmak, J. J., and Grossberg, S. E. 1977. A rapid and versatile assay forprotein using Coomassie brilliant blue G-250. Anal. Biochem. 79:544–552.
omogyi, M. 1952. Notes on sugar determination. J. Biol. Chem. 195:19–23.
netselaar, K. M., and Mims, C. W. 1994. Light and electron microscopyof Ustilago maydis hyphae in maize. Mycol. Res. 98: 347–355.
pelling, T., Bolker, M., Lottspeich, F., Frank, R. W., and Kahmann, R.1994. Pheromones trigger filamentous growth in Ustilago maydis.EMBO J. 13: 1620–1627.alton, J. D. 1994. Deconstructing the cell wall. Plant Physiol. 104:1113–1118.
Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.
Copyright © 2022 FDOKUMEN




















