
Crystal structures of .alpha.-lytic protease complexes with irreversibly bound phosphonate esters
Upload
independentCategory
view
0download
0
Journal of General Virology (1995), 76, 1401-1408. Printed & Great Britain 1401
Qualitative analysis of the expression of Epstein-Barr virus lytic genes in nasopharyngeal carcinoma biopsies
Domin ique Marte l -Renoir , Virginie Grunewald, Robert Toui tou , Guy S c h w a a b and Irene J o a b *
C N R S U R A 1301, Institut Gustave Roussy, 39 rue Camille Desmoulins, 94800 Villejuif, France
We recently showed that BZLF1, the gene encoding the Epstein-Barr virus (EBV) ZEBRA protein, was ex- pressed in all eight nasopharyngeal carcinoma (NPC) specimens studied. We present here studies on the expression of EBV lyric cycle genes in the same eight NPC biopsies to determine if production of the ZEBRA transactivator could lead to a complete productive cycle. The tumour lesions exhibit a number of different patterns of limited lytic gene expression. In three out of eight tumours neither BRLF1 nor BMLF1 expression could be detected. Otherwise BMLF1 mRNA was
expressed in all the other specimens. Three specimens also expressed BRLF1. Two specimens not only exhibited BZLF1, BMLF1 and BRLF1 transcripts, but also expressed the late gene BLLF1 which encodes the membrane protein gp220. The early gene product BBLF2 could not be detected in any of the eight NPC. However, expression of the late gene encoding the lyric truncated form of LMP1 (D1LMP) was found in seven of the eight NPC biopsies. Thus, it could be suggested that the EBV abortive lytic cycle occurred in most of the NPC studied.
Introduct ion
The Epstein-Barr virus (EBV) is the causative agent of infectious mononucleosis and has been associated with different malignancies: endemic Burkitt's lymphoma, 30% of Hodgkin's disease, B cell lymphomas in immunocompromised individuals, T cell lymphomas, nasopharyngeal carcinoma (NPC), and more recently with gastric carcinoma (Miller, 1990; Chen et al., 1993; Oda et al., 1993). Viral genes expressed in EBV- associated tumours could contribute to carcinogenesis. In NPC, expression of latent viral genes has been observed: EBERs, EBNA1, LMP1 and -2 and a family of mRNAs transcribed rightward through the B a m H I A (Gilligan et aL, 1990, 1991 ; Fahraeus et aL, 1988; Young et al., 1988; Niedobitek et al., 1992; Brooks et al., 1992; Busson et al., 1992; Hit tet al., 1989; Chen et al., 1992).
Serological studies have shown that the clinical onset of NPC is preceded by the appearance of a high antibody titre to EA and VCA. The titres increase with the total tumour burden and the antibodies gradually decline with the response to therapy (Henle et al., 1973; de Th6, 1980). This suggests that the expression of the lytic cycle is associated with the presence of the tumour. We have shown that most NPC patients (87 %) exhibit antibodies to the ZEBRA protein (this protein, also referred as EB1 or Zta, switches EB virus from the latent to the
* Author for correspondence. Fax +33 1 45 59 70 09.
productive cycle) (Joab et al., 1991). We also demonstrated that the gene encoding the ZEBRA protein, BZLFI, was expressed in all NPC specimens studied (Martel-Renoir et al., 1993; Cochet et al., 1993). In order to determine if expression of BZLF1 gives rise to complete or abortive productive cycle, RT-PCR analysis was performed to investigate the expression of early and late genes in eight tumour biopsies. Spliced lytic transcripts, easily distinguishable from genomic DNA were assayed: (1) the immediate early gene B RLF 1 encoding the R transactivator (Manet et al., 1989); (2) the early gene BMLF1 encoding the M transactivator (Buisson et al., 1989); (3) BBLF2-BBLF3 encoding a primase associated factor (Farrell, 1989); and (4) BLLF 1 late gene encoding gp220/350 glycoproteins (Beisel et al., 1985; Tosoni-Pittoni et al., 1989). We also investigated the expression of BNLF1 encoding the latent as well as the truncated form of LMP1, the latter being charac- teristic of the lytic cycle (Hudson et al., 1985). The results presented here suggest that, in most of the cases, an abortive lytic cycle occurs in NPC.
M e t h o d s
Preparation of cDNA. Frozen specimens were pulverized and total RNA prepared using either the guanidinilma thiocyanate method described by Chirgwin et al. (1979) or by the RNAzol B method (Biotecx Laboratories).
The total RNA samples were treated for 1 h at 37 °C with RNase- free RQ 1 DNase (Promega). After heat inactivation at 68 °C for 15 min and ethanol precipitation, the RNA samples were heated for 3 min at
0001-3019 © 1995 SGM

1402 D. Martel-Renoir and others
T a b l e 1. Summary of the different primers used in the PCR amplifications*
Target Sequence (5' to 3 ~) Genomic coordinates
BRLF1 Upstream primer RS1 Downstream primer RAS4 BMLF1 Upstream primer M 1S Downstream primer M2AS BBLF3 Upstream primer F3S Downstream primer F3AS gp220 Upstream primer GPS Downstream primer GPAS LMP Upstream primer LMPS Upstream primer LMP2S Upstream primer LMP2CS Downstream primer LMP1AS Downstream primer LMPEAS Downstream primer LMPGAS
CACACAACACCTCACTACAC CTATGGTGGCAGGAATCATC
TCTCCCGAACTAGCAGCATT AACAGTCCTAGTGACCCAGT
TAGCACAG/GCTTTCCTGCAA CTCGCGTTGATTTCCATAGGA
ACGGCACAGAAA/GTGGTGAA ACTGCAGTACTAGCATGGAG
CTGAGGATGGAACACGACCT ACGCACTTTCTCCTCTTTCC CTAGCGACTCTGCTGGAAAT AATGGAGGGAGAGTCAGTCA GATACCTAAGA/CAAGTAAGCA ACGGGATGACTCATCTCAAC
106138-106157 104775-104794
84250-84269 82768-82787
11722-117515/117386-117375 116819-116838
90664-90653/900061-90054 89558 89577
169480-169461 169133 169152 168373-168392 16809%168118 168955 168965/169042-169051 166869-166888
* The coordinates refer to the positions in the B95-8 genomic map.
80 °c and then rapidly chilled on ice immediately before being reverse transcribed with Moloney murine leukaemia virus reverse transcriptase (M-MLV RT) (GIBCO-BRL) in the presence of ohgo(dT)a 5 as a primer (Promega).
Preparation of probes. Clones of EBV (B95-8) DNA in pBR322 were used (Dambaugh et al.. 1980). Specific probes were prepared from the corresponding plasmids after digestion with the appropriate restriction enzymes. Fragments were purified twxce through 1% Nu Sieve GTG agarose gel (FMC), and then radiolabelled with the Prime-a-Gene system (Promega).
Amplification and analysis of cDNA. The eDNA samples (obtained from 200 ng of total RNA) were subjected to PCR using Taq DNA polymerase (Promega) and specific oligonucleotides (Table 1). The cycle procedure was standardized for all reactions: denaturation for 5 min at 95 °C was followed by the addition of 2.5 U of the enzyme, then denaturation for 30 s at 95 °C, annealing for 1 min at 58 °C, and extension for 2 min at 72 °C for 30 cycles. Cycling was followed by 7 min extension at 72 °C. When specified, 2 ~tl of the PCR product were used for a second round of PCR.
PCR products were analysed by electrophoresis through a 1% agarose gel (Bioprobe systems) and Southern transfer onto Hybond-N membranes (Amersham). Detection of products was achieved by hybridization to a 32P-labelled probe.
When specified, DNA fragments were excised from the agarose gel, purified, made blunt-ended with the Klenow fragment of DNA polymerase, phosphorylated, inserted at the EeoRV site of plasmid pGEM-5ZF + (Promega) and sequenced with the Dyedeoxy terminator cycle sequencing kit (Applied Biosystems) on a model 373A automatic sequenccr.
Results
Analysis of the eDNA preparations
The same c D N A prepara t ions were used in the present s tudy as in the previous report on BZLF1 expression (Cochet et al., 1993). The qual i ty of the c D N A was
.. ;Z.::::N
.,~? ..~.
, . , - , ~ Y ' ~ ~ 4 - 246 bp
Fig. 1. PCR analysis of eDNA of the house-keeping gene hypoxanthine phosphoryltransferase. PCR product (10 ~tl BP 14, 20, 30, 38, 40 and MD NPC biopsies; 20 Ixl for BP 46 and BP 8 biopsies) obtained from 200 ng of total RNA were migrated through 1% agarose gels. The negative control (H20) contained no DNA.
checked by P C R amplifications of the housekeeping gene hypoxanth ine phosphoryl t ransferase, using as the sense pr imer 5' T A T G G A C A G G A C T G A A C G T C and as the antisense pr imer 5' G T T G A G A G A T C A T C T - C C A A C C . The expected band at 246 bp was observed in all biopsy samples (Fig. 1).
Analysis of BRLF1 immediate early transcripts
BRLF1 codes for the R t ransact ivator and is t ranscribed
into several m R N A species: one t ranscript of the entire genomic sequence and another with the 939 bp in t ron sequence spliced out (Mane t et al., 1989). c D N A was amplified using an ups t ream pr imer in exon a (RSI) and a downs t ream primer in exon b (RAS4). Two bands (Fig. 2) were identified in the positive control c D N A of B95- 8 cells "~a ted for 48 h with T P A (12-O-tetradecanoyl-
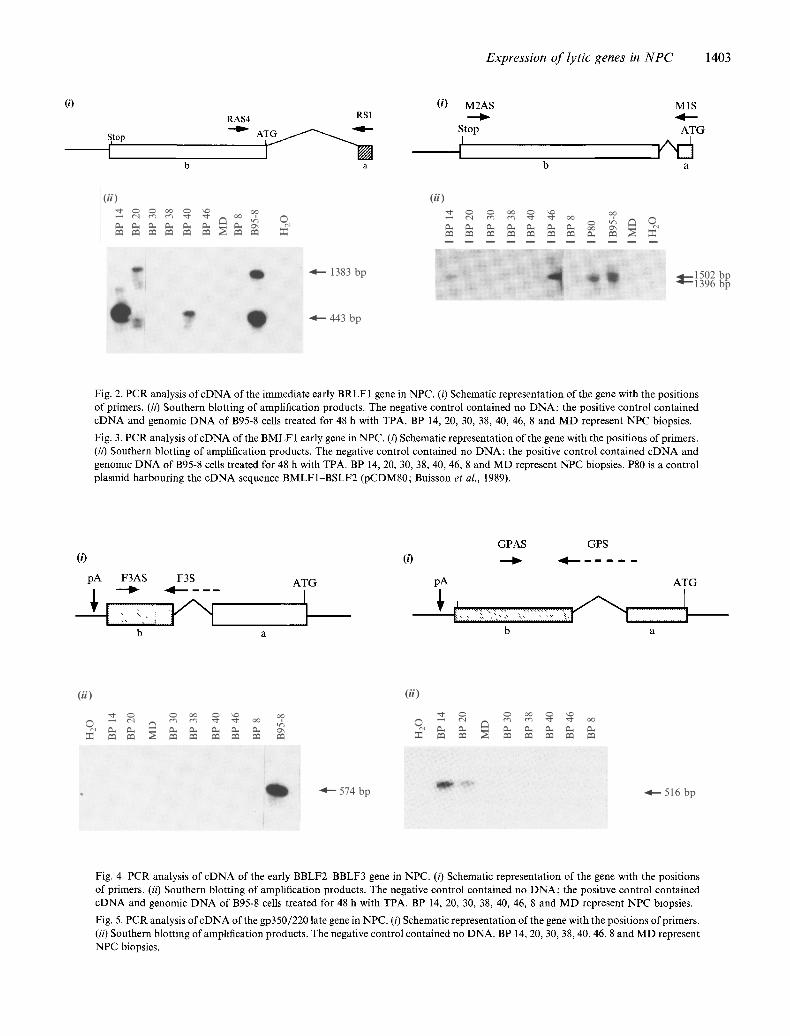
Expression of lytic genes in NPC 1403
(i) (i) M2AS RAS4 RSI - -
Stop Stop
I I b a
M1S
ATG
a
(ii) (ii)
l i d ~ 1383 bp
IL -, e
~ g 1[::::: 1502 bp 1396 bp
Fig. 2 Fig. 3
Fig. 2. PCR analysis of cDNA of the immediate early BRLF 1 gene in NPC. (/) Schematic representation of the gene with the positions of primers. (ii) Southern blotting of amplification products. The negative control contained no DNA; the positive control contained cDNA and genomic DNA of B95-8 cells treated for 48 h with TPA. BP 14, 20, 30, 38, 40, 46, 8 and MD represent NPC biopsies.
Fig. 3. PCR analysis of cDNA of the BMLF 1 early gene in NPC. (i) Schematic representation of the gene with the positions of primers. (ii) Southern blotting of amplification products. The negative control contained no DNA; the positive control contained cDNA and genomlc DNA of B95-8 cells treated for 48 h with TPA. BP 14, 20, 30, 38, 40, 46, 8 and MD represent NPC biopsies. P80 is a control plasmid harbouring the cDNA sequence BMLF1 BSLF2 (pCDM80; Buisson et al., 1989).
(0 (,3
pA F3AS F3S ATG pA 4
[iiiiiiiiii!i ;iiili U h '1 b a
GPAS GPS
4
ATG
b a
(ii) (ii)
4 - - 574 bp I I~ 4 - - 516 bp
Fig. 4 Fig. 5
Fig. 4. PCR analysis of cDNA of the early BBLF2-BBLF3 gene in NPC. (i) Schematic representation of the gene with the positions of primers. (ii) Southern blotting of amplification products. The negative control contained no DNA; the positive control contained cDNA and genomic DNA of B95-8 cells treated for 48 h with TPA. BP 14, 20, 30, 38, 40, 46, 8 and MD represent NPC biopsies.
Fig. 5. PCR analysis of cDNA of the gp350/220 late gene in NPC. (i) Schematic representation of the gene with the positions of primers. (i0 Southern blotting of amplification products. The negative control contained no DNA. BP 14, 20, 30, 38, 40, 46, 8 and MD represent NPC biopsies.
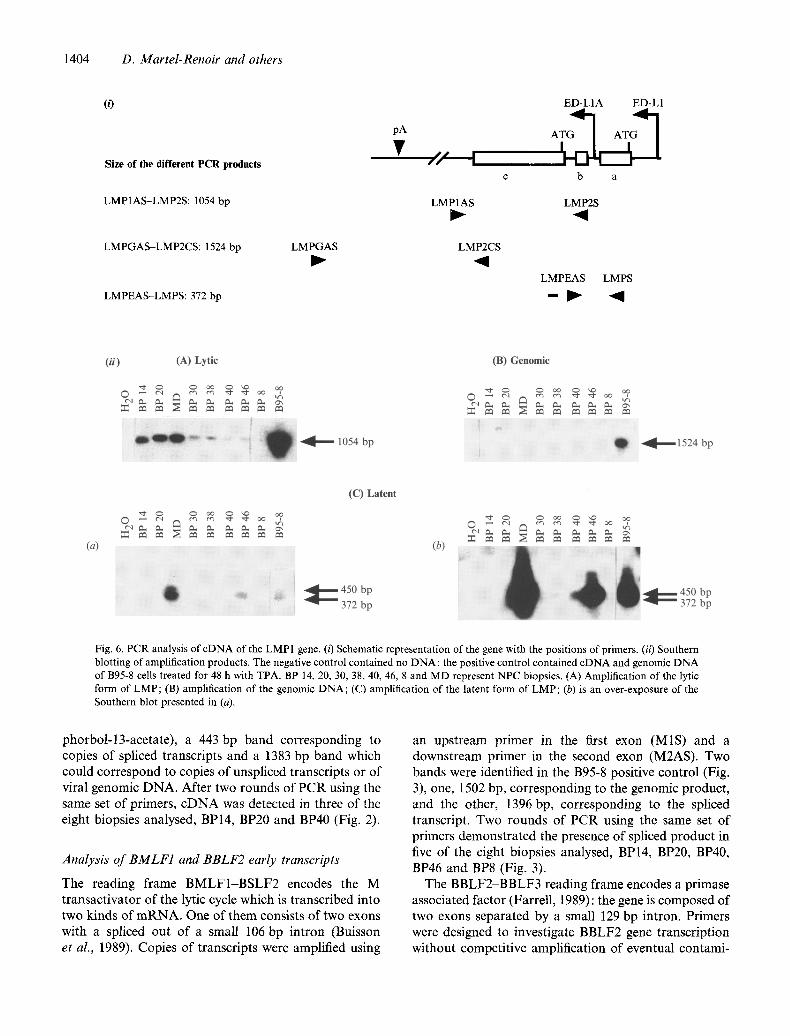
1404 D. Martel-Renoir and others
(0
Size of the different PCR products
LMP1AS-LMP2S: 1054 bp
LMPGAS-LMP2CS: 1524 bp
LMPEAS-LMPS: 372 bp
LMPGAS
pA
• / I ,
LMP1AS
LMP2CS
ED-L1A ED-L1
A'o I b a
LMP2S I
LMPEAS LMPS
- I ~ I I
(ii) (.4,) Lyric (B) Genomic
l l D q ~ . ' ,~" - -1054 bp O -,4,---1524 bp
(a)
(C) Latent
(b)
O ~ 450 bp - ' ~ 372 bp
&
ti
t =:4 0 Fig. 6. PCR analysis of cDNA of the LMP1 gene. (i) Schematic representation of the gene with the positions of primers. (ii) Southern blotting of amplification products. The negative control contained no DNA; the positive control contained cDNA and genomic DNA of B95-8 cells treated for 48 h with TPA. BP 14. 20, 30, 38, 40, 46, 8 and MD represent NPC biopsies. (A) Amplification of the lytic form of LMP; (B) amplification of the genomic DNA; (C) amplification of the latent form of LMP; (b) is an over-exposure of the Southern blot presented in (a).
phorbol-13-acetate), a 443 bp band corresponding to copies of spliced transcripts and a 1383 bp band which could correspond to copies of unspliced transcripts or of viral genomic DNA. After two rounds of PCR using the same set of primers, cDNA was detected in three of the eight biopsies analysed, BP14, BP20 and BP40 (Fig. 2).
Analysis of BMLF1 and BBLF2 early transcripts
The reading frame BMLF1-BSLF2 encodes the M transactivator of the lytic cycle which is transcribed into two kinds of mRNA. One of them consists of two exons with a spliced out of a small 106 bp intron (Buisson et al., 1989). Copies of transcripts were amplified using
an upstream primer in the first exon (MIS) and a downstream primer in the second exon (M2AS). Two bands were identified in the B95-8 positive control (Fig. 3), one, 1502 bp, corresponding to the genomic product, and the other, 1396 bp, corresponding to the spliced transcript. Two rounds of PCR using the same set of primers demonstrated the presence of spliced product in five of the eight biopsies analysed, BP14, BP20, BP40, BP46 and BP8 (Fig. 3).
The BBLF2-BBLF3 reading frame encodes a primase associated factor (Farrell, 1989): the gene is composed of two exons separated by a small 129 bp intron. Primers were designed to investigate BBLF2 gene transcription without competitive amplification of eventual contami-

Expression of lytic genes in NPC 1405
nating genomic DNA. The upstream primer (F3S) described a sequence across the splice junction between the two exons, and the downstream primer (F3AS) was chosen in exon b. A PCR product of 574bp was expected. In Fig. 4, a band of the predicted size is observed in the positive control cDNA of B95-8 TPA- treated cells; however, there was no amplification product in any of the eight NPC tumour biopsies tested. After a second round of PCR, using the same set of primers, no amplification of the BBLF2 gene product could be detected in any of the NPC turnout biopsies (Fig. 4).
Analysis o f late transcripts coding for gp220 and LMP1
The BLLF1 reading frame encodes the principal com- ponents of the outer surface glycoproteins gp350 and gp220 (Beisel et al., 1985; Tosoni-Pittoni et al., 1989). When the 591 bp intron of the BLLF1 transcript is spliced out of the RNA, the RNA codes for gp220. In the unspliced form, the RNA codes for gp350. The gp220 transcripts were amplified using an upstream primer (GPS) across the splice junction between exons a and b and a downstream primer (GPAS) in exon b. A PCR product of 516 bp was expected. After a first round of amplification (not shown), a band was visible only in the B95-8 positive control. After a second round of amplification using the same primers the results (Fig. 5) showed the presence of cDNA encoding gp220 in only two tumour biopsies, BP14 and BP20.
LMP1 is encoded by the BNLF1 reading frame: the gene contains three exons separated by two small introns of 76 and 78 bp. Two kinds of transcripts are expressed according to the state of virus cycle. A transcript containing the three exons is transcribed from the E D - L 1 promoter during the latent cycle, while an unspliced mRNA is transcribed from the ED-L1A promoter during the lytic cycle (Hudson et al., 1985). The resulting late and truncated LMP is functionally distinct from normal LMP (Wang et al., 1988). Primers were designed to distinguish these two types of transcripts.
Transcripts encoding the truncated form of the protein were amplified using the primer LMP2S in the intron between exons a and b downstream of the ED-L1A promoter, and the primer LMP1AS in the untranslated part of the cDNA downstream of exon c; a PCR product of 1054 bp was expected. Fig. 6A shows that after one round of PCR, the transcripts were present in all biopsies except tumour BP8. Since the cDNA encoding the truncated LMP is not spliced, it was important to demonstrate that genomic DNA was not detectable in our preparations. We amplified possible contaminating genomic DNA with an upstream primer in exon c (LMP- 2CS) and a downstream primer (LMPGAS) localized
downstream of the polyadenylation site; an amplified PCR product of 1524 bp was expected and a band of that size was observed when control D N A from B95-8 cells was used (Fig. 6B). This clearly shows that genomic contamination could not account for the 1054bp molecules obtained after PCR amplification in the tumour biopsies. By using serial dilutions of known amounts of DNA and limiting PCR conditions (20 cycles of amplification) we established that amplification of the 1524 bp band is at least as efficient as amplification of the 1054 bp piece (not shown).
The transcripts encoding the latent form of LMP were analysed using a primer located in exon a (LMPS) and a primer (LMPEAS) overlapping the splice junction between exons c and b. After two rounds of PCR (Fig. 6C), transcripts for the latent form of LMP1 (372 bp) were visualized in four of the eight biopsies, MD, BP38, BP40 and BP46. We also visualized a 450 bp band; sequencing of this product demonstrated that one intron was retained in this form.
D i s c u s s i o n
Previous work from our laboratory has demonstrated by in situ hybridization that NPC tumour cells exhibit expression of the BZLF1 gene, and by reverse PCR that spliced transcripts of the BZLF 1 gene were present in all the NPC biopsies analysed (eight specimens) (Martel- Renoir et al., 1993; Cochet et al., 1993). In the present study, by using the same cDNA preparations, we investigated the expression of EBV early and late genes. An overview of the results is shown in Table 2: the tumour lesions exhibit differing patterns of limited lytic gene expression. In three out of eight tumours (BP30, BP38 and ME)) neither BRLF1 nor BMLF1 expression could be detected. Otherwise BMLF1 mRNA was expressed in all the other specimens. Three specimens also expressed BRLF1 (BP14, BP20 and BP40). Two specimens (BP14 and BP20) not only exhibit BZLF1, BMLF1 and BRLF1 transcripts, but also expressed the late gene BLLF1 which encodes the membrane protein gp220. Two rounds of 30 cycles of PCR were necessary to visualize these last cDNAs. Within the samples that we analysed, we did not observe transcripts from BZLF 1
Table 2. Expression of E B V lytic genes in NPC biopsies
BP8 BP14BP20 MD BP30 BP38 BP40BP46
ZEBRA + + + + + + + + BRLF1 - + + - - - + - BMLFI + + + - - - + + BBLF3 . . . . . . . . gp220 - + + . . . . . LMP lytic - + + + + + + +

1406 D. Mar te l -Renoir and others
accompanied by BRLF1 without BMLF1 and we did not detect BLLF1 transcripts without transcription of BZLF1, BMLF1 and BRLF1. However, we don't know if each tumour cell is able to support the expression of all the different EBV lytic mRNAs observed. These results are in agreement with earlier studies on EBV expression in NPC cultivated in nude mice indicating that viral expression was not tightly restricted and that replication or abortive infection occurred in tumour cells (Raab- Traub et al., 1983). In previous work (Cochet et al., 1993), we demonstrated by in situ hybridization that the transcripts encoding ZEBRA were confined to large epithelial tumour cells. We cannot rule out the possibility that the lytic RNAs are expressed both in the infiltrating lymphocytes and in the tumour cells.
All the NPC biopsies did not appear to show transcription of the early BBLF2 BBLF3 reading frame, even after two rounds of PCR, indicating that this gene is not transcribed in NPC, at least not in a sufficient amount to be detected. Another gene, the lytic cycle BHRF1 mRNA, which is abundant in LCL, was also not detected by reverse PCR by Sam et al. (1993) in NPC.
In striking contrast with these observations is the expression of the late gene encoding the lytic truncated form of LMP 1, D 1LMP. Transcripts from this gene were found in seven of the eight NPC biopsies. Since this transcript has no intron, it was of importance to demonstrate that, in spite of the DNase 1 treatment, genomic DNA could not account for the band observed after RT-PCR. Amplification of the DNA with a primer located downstream of the polyadenylation signal showed that genomic contamination was not detectable. The transcripts encoding D1LMP are detectable in the biopsies after a single round of PCR; this could reflect either the abundance of the transcripts or a more effective amplification. Two rounds of PCR are required to visualize the equivalent latent LMP in the same biopsies although BNLF1 transcripts were described as being abundant in NPC biopsies (Gilligan et al., 1990). Similarly, Brooks et al. (1992) also used nested PCR to detect copies of mRNA encoding LMP1 from NPC cultivated in nude mice. Transcripts of latent LMP were visualized in four of the eight biopsies. These observations are in agreement with previous studies on LMP expression in NPC which demonstrate hetero- geneous expression of LMP between individual tumours, the expression varying from 20 to 80 % of cases (Brooks et al., 1992; Niedobitek et al., 1992; Fahraeus et al., 1988). Stewart & Arrand (1993) also reported expression of LMP protein in seven of nine NPC biopsies by probing tissue sections with monoclonal antibodies, CS1-4, raised against a fusion protein between fl- galactosidase and the carboxy terminus of LMP (Rowe et al., 1987). However, these antibodies would also react
with the truncated form of the LMP protein. Since D1LMP protein has no transformation-associated phenotypic effect in Rat-l cells, it is difficult to ascribe to this protein a crucial role in the development of an NPC tumour (Wang et al., 1988; Baichwal & Sugden, 1989).
The findings that emerge from this work are that the EBV lytic cycle is reactivated in NPC, and that this productive cycle is often incomplete. The expression of lytic antigens varied among the individual tumours analysed. The block in the productive cycle could depend either on a defective viral protein, or on the permissivity of the tumour cell itself (Gradoville et al., 1990). However, a feature common to most of the biopsies seems to be expression of the immediate early trans- activator ZEBRA and the lyric form of LMP1.
A number of reports have shown that an abortive lytic cycle occurs in non-Hodgkin's lymphoma of immuno- compromised patients (Katz et al., 1989; Pallesen et al., 1991 a; Bashir et al., 1993). Similar results were observed in Hodgkin and Reed-Sternberg cells (Pallesen et al., 1991b; Bibeau et al., 1994). An abortive lytic cycle has also been reported to occur in gastric UCNT (Rowlands et al., 1993). Our results also suggest that viral reactivation might be a general feature of NPC tumours. The proportion of turnout cells of the EBV related tumours supporting the abortive viral cycle has not been evaluated. Even if viral reactivation concerns only a small population of the primary tumour cells, charac- terization of lytic gene expression remains of interest since abortive lytic cycle in NPC might result in survival of the host cell allowing interaction of viral lytic proteins with cellular processes. It has been recently demonstrated that ZEBRA antigen is able to inhibit p53-dependent transactivation (Zhang et at., 1994) and to functionally and physically interact with the p65 subunit of the cellular factor NF-KB (Gutsch et al., 1994). The BMLF1 lytic gene has been demonstrated to exhibit transforming properties (Corbo et al., 1994). Thus the deregulation of the programmed expression of the EBV lytic antigens could interfere with cellular functions and this may contribute to carcinogenesis.
We are grateful to Martin Lee for helpful reading of the manuscript. We thank Alain Sergeant for providing the pDM80 clone. This investigation was supported by grants from ARC 2017, IGR CRC 92.20 INSERM and FEGEFLUC.
References BAICHWAL, V R. & SUGDEN, B. (1989). The multiple membrane-
spanning segments of the BNLF-I oncogene from Epstei~Barr virus are required for transformation. Oncogene 4, 67-74.
BASHIR, R., LUKA, J., CHELOHA, K., CHAMBERLAIN, M. & HOCHBERG, F. (1993). Expression of Epstein-Barr virus proteins in primary CNS lymphoma m AIDS patients. Neurology 43, 2358-2362.
BEISEL, C., TANNER, J.. MATSUO, T., THORLEY-LAwsON, D., KEZDY, F. & KIEEE, E. (1985). Two major outer envelope glycoproteins of

Expression o f lytic genes in N P C 1407
Epstein-Barr virus are encoded by the same gene. Journal of Virology 54, 665-674.
BIBEAU, F., BROUSSET, P., KNECHT, H., MEGGETTO, F., DROUET, E., RUmN, B. & DELSOL, G. (1994). Epstein-Barr virus replication in Hodgkins disease. Bulletin du Cancer 81, 114-118.
BROOKS, L., YAO, Q.Y., RICKINSON, A. B. & YOUNG, L. S. (1992). Epstei~Barr virus latent gene transcription in nasopharyngeal carcinoma cells: coexpression of EBNA1, LMP1, and LMP2 transcripts. Journal of Virology 66, 2689-2697.
BUISSON, M., MANET, E., TRESCOL-BIEMONT, M.C., GRUFFAT, H., DURAND, B. & SERGEANT, A. (1989). The Epstein-Burr virus (EBV) early protein EB2 is a posttranscrlptional activator expressed under the control of EBV transcription factors EB1 and R. Journal of Virology 63, 5276-5284.
BUSSON, P., McCoY, R., SADLER, R., GILLIGAN, K., TURSZ, T. & RAAB- TRAUB, N. (1992). Consistent transcription of the Epstein-Barr virus LMP2 gene in nasopharyngeal carcinoma. Journal of Virology 66, 3257-3262.
CHEN, C. L., SADLER, R. H., WALLING, D. M., SU, I. J., HSIEH, H. C. & RAAB-TRAuE, N. (1993). Epstein-Barr virus (EBV) gene expression in EBV-positive peripheral T-cell lymphomas. Journal of Virology 67, 6303-6308.
CHEN, H. L., LUNG, M. M. L., SHAM, J. S. T., CHOY, D. T. K., GRIFFIN, B. & NG, M. H. (1992). Transcription of BamHI-A region of the EBV genome in NPC tissues and in B cells. Virology 191, 193-201.
CHIRGWIN, J.M., PRZYBYLA, A. E., MACDONALD, R.J. & RUTTER, W. J. (1979). Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18, 5294-5299.
COCHET, C., MARTEL-RENOIR, D., GRUNEWALD, V., BOSQ, J., COCHET, G., SCHWAAB, G., BERNAUDIN, J. F. & JOAn, I. (1993). Expression of the Epstein-Burr virus immediate early Eerie, BZLF1, in naso- pharyngeal carcinoma tumour cells. Virology 197, 358-365.
CORBO, L., LE Roux, F. & SERGEANT, A. (1994). The EBV early gene product EB2 transforms rodent cells through a signalling pathway involving c-MYC. Oncogene 9, 3299-3304.
DAMBAUGH, T., BEOSEM, C., HUMMEL, M., KING, W., FENNEWALD, S., CHEUNG, A., HELLER, M., RAAB-TRAUB, N. & K1EFF, E. (1980). Epstein-Burr virus (B95-8) DNA VII: molecular cloning and detailed mapping. Proceedings' of the National Academy of Sciences, USA 77, 2999-3003.
DE THt~, G. (1980). The role of Epstein-Barr virus in human diseases infectious mononucleosis (iM), Burkitts iymphomas (BL), and nasopharyngeal carcinoma (NPC). In Viral Oncology, pp. 769-797. Edited by G. Klein. New York: Raven Press.
FAHRAEUS, R., HULl, F., ERNBERG, I., FINKE, J., ROWE, M., KLEIN, G., FALK, K., NILSSON, E., YADAV, M., BUSSON, P., TURZ, T. & KALIING, B. (1988). Expression of Epstei~Barr virus-encoded proteins in nasopharyngeal carcinoma. International Journal of Cancer 42, 329 338.
FARRELL, P. J. (1989). Epstein-Burr virus genome. Advances in Viral Oncology 8, 103-259.
GILLIGAN, K. J., RAJADURAI, P., LIN, J. C., BUSSON, P., ABDEL-HAMID, M., PRASAD, U., TURSZ, T. & RAAB-TRAUB, N. (1991). Expression of the Epstein-Burr virus BamHI A fragment in nasopharyngeal carcinoma: evidence for a viral protein expressed in vivo. Journal of Virology 65, 6252-6259.
GILLIGAN~ K., SATO, I~., RAJADURAI, P., BUSSON, P., YOUNG, L., RICKINSON, A., TURSZ, T. & RAAB-TRAUE, N. (1990). Novel transcription from the Epstein-Burr virus terminal EcoRI fragment, DIJhet, in a nasopharyngeal carcinoma. Journal of Virology 64, 4948-4956.
GRADOVILLE, L., GROGAN, E., TAYLOR, N. & MILLER, G. (1990). Differences in the extent of activation of Epstein-Burr virus replicative gene expression among four nonproducer cell lines stably transformed by oriP/BZLF1 plasmids. Virology 178, 345-354.
GUTSCH, D.E., HOLLEY-GUTHRIE, E.A., ZHANG, Q., STEIN, B., BLANAR, M. A., BALDWIN, A. S. & KENNEY, S. C. (1994). The b-Zip transactlvator of Epstein-Burr virus, BZLFI, functionally and physically interacts with p65 subunit of NFKB. Molecular and Cellular Biology 14, 1939 1948.
HENLE, W., Ho, J. H. C., HENLE, G. • KWAN, H. C. (1973). Antibodies to Epstein-Barr virus related antigens in nasopharyngeal carcinoma.
Comparison to active cases and long term survivors. Journal of the National Cancer Institute 51, 361-369.
HITT, M. M., ALLDAY, M. J., HARA, T., KARRAN, L., JONES, U. D., BUSSON, P., TURSZ, T., ERNBERG, I. & GRIFFIN, B. E. (1989). EBV gene expression in an NPC-related tumour. EMBO Journal 8, 2639-2651.
HUDSON, G., FARRELL, P. J. & BARRELL, B. G. (1985). Two related but differentially expressed potential membrane proteins encoded by the EcoRI Dhet region of Epstein-Barr virus B95-8. Journal of Virology 53, 528-535.
JOAn, I., NICOLAS, J.C., SCHWAAB, G., DE THE, G., CLAUSSE, B., PERRICAUDET, M. & ZENG, Y. (1991). Detection of anti-Epsteln-Barr virus transactivator, ZEBRA, antibodies in sera from patients with nasopharyngeal carcinoma. International Journal of Cancer 48, 647-649.
KATZ, B.Z., RAAB-TRAUB, N. & MILLER, G. (1989). Latent and replicating forms of Epstei~Barr virus DNA in lymphomas and lymphoproliferative diseases. Journal of Infectious Diseases 160, 58%598.
MANET, E., GRUFFAT, H., TRESCOL-BIEMONT, M.C., MORENO, N., CHAMBARD, P, GIOT, J.F. & SERGEANT, A. (1989). Epstein-Barr virus bicistronic mRNAs generated by facultative splicing code for two transcriptional trans-activators. EMBO Journal 8, 1819 1826.
MARTEL-RENoIR, D., GRUNEWALD, V., COCHET, C., BOSQ, J., SCI-IWAAB, G., BERNAUDiN, J. F. & JOAn, I. (1993). Expression of Epstein-Burr virus transactivator, ZEBRA, in nasopharyngeal carcinoma tumour cells. In The Epstein-Burr Virus and Associated Diseases, pp. 489-496. Edited by T. Tursz, J. Pagano, D. V. D. Ablashi, G. de Th6 & G. Lenoir. Paris: John Libbey Eurotext Ltd.
MILLER, G. (1990). Epstei~Barr virus. Biology, pathogenesis and medical aspects. In Virology, pp. 1921 1958. Edited by B. N. Fields & D. M. Knipe. New York: Raven Press.
NIEDOBITEK, G., YOUNG, L. S., SAM, C. K., BROOKS, L., PRASAD, U. & RICKINSON, A. B. (1992). Expression of Epstein-Burr virus genes and of lymphocyte activation molecules in undifferentiated naso- pharyngeal carcinomas. American Journal of Pathology 140, 879 887.
ODA, K., TAMARU, J., TAKENOUCHI, T., MIKATA, A., NUNOMURA, M., SAITOH, N., SARASHINA, H. t~; NAKAJ1MA, N. (1993). Association of Epstein-Burr virus with gastric carcinoma with lymphoid stroma. American Journal of Pathology 143, 1063 1071.
PALLESEN, G., HAMILTON-DUTOIT, S.J., ROWE, M., LISSE, I., RALFKIAER, E., SANDVEJ, K. & YOUNG, L. S. (i99i). Expression of Epstein-Barr virus replicative proteins in AIDS-related non- Hodgkin's lymphoma cells. American Journal of Pathology 165, 289 299.
PALLESEN, G., SANDVEJ, m., HAMILTON-DUTOIX, S.J., ROWE, M. & YOUNG, L. S. (1991). Activation of Epstein-Barr virus replication in Hodgkin and Reed-Sternberg cells. Blood 78, 1162-1165.
RAAB-TRAUB, N., HOOD, R., YANG, C. S., HENRY, B. & PAGANOJ, S. (1983). Epstein-Barr virus transcription in nasopharyngeal car- cinoma. Journal of Virology 48, 580-590.
ROWE, M., EVANS, H. S., YOUNG, L. S., HENNESSY, K., K1EFF, E. & RICKINSON, A.B. (1987). Monoclonal antibodies to the latent membrane protein of Epstein-Burr virus reveal heterogeneity of the protein and inducible expression in virus-transformed ceils. Journal of General Virology 68, 1575-1586.
ROWLANDS~ D. C., ITO, M., MANGHAM, D. C., REYNOLDS, G., HERBST, H., HALLISSEY, M. W., FIELDING, J. W. L., NEWBOLD, K. M., JONES, E. L., YOUNG, L. S. & NmDOmTEK, G. (1993). Epstein-Burr virus and carcinomas - rare association of the virus with gastric adeno- carcinomas. British Journal of Cancer 68, 1014--1019.
SAM, C. K., BROOKS, L. A., NIEDOBITEK, G., YOUNG, L. S., PRASAD, U. ~; RICKINSON, A. B. (1993). Analysis of Epstein-Burr virus infection in nasopharyngeal biopsies from a group at high risk of nasopharyngeal carcinoma. International Journal of Cancer 53, 957 962.
STEWART, J. P. (~; ARRAND, J. R. (1993). Expression of the Epstei~Barr virus latent membrane protein in nasopharyngeal carcinoma biopsy specimens see comments. Human Pathology 24, 239542.
TOSONI-PITTONI, E., JOAB, I., NICOLAS, J. C. & PERRICAUDET, i . (1989). Complete characterization of the gene coding for the Epstei~Barr virus major membrane antigen gp 220/340 and selective expression

1408 D. Martel-Renoir and others
of a secreted form of gp 220. Biochemical and Biophysical Research Communications 158, 676--684.
WANG, D., LIEBOWITZ, D., WANG, F., GREGORY, C., RIfSKINSON, A., LAP, SON, R., SPRINGER, T. • KIEFF, E. (1988). Epstein-Barr virus latent infection membrane protein alters the human B-lymphocyte phenotype: deletion of the amino terminus abolishes activity. Journal of Virology 62, 4173-4184.
Yotm~, L. S., DAWSON, C. W., CLARK, D., RUPANI, H., BUSSON, P., TURSZ, T., JOHNSON, A. & RICKINSON, A. B. (1988). Epstein-Barr virus gene expression in nasopharyngeal carcinoma. Journal of General Virology 69, 1051-1065.
ZHANG, Q., GUTSCH, D. & KENNEY, S. (1994). Functional and physical interaction between p53 and BZLF1 : implications for Epstein-Barr virus latency. Molecular and Cellular Biology 14, 1929-1938.
Copyright © 2022 FDOKUMEN




















