
Proteinases and exopeptidases from the phytopathogenic fungus Ustilago maydis
13
327 Mycologia, 95(2), 2003, pp. 327–339. q 2003 by The Mycological Society of America, Lawrence, KS 66044-8897 Proteinases and exopeptidases from the phytopathogenic fungus Ustilago maydis Yuridia Mercado-Flores Ce ´sar Herna ´ndez-Rodrı ´guez Departamento de Microbiologı ´a. Escuela Nacional de Ciencias Biolo ´gicas, IPN. Casco de Santo Toma ´s. Me ´xico DF Jose ´ Ruiz-Herrera Departamento de Ingenierı ´a Gene ´tica, Unidad Irapuato, Centro de Investigacio ´n y Estudios Avanzados, IPN. Irapuato, Guanajuato, Me ´xico Lourdes Villa-Tanaca 1 Departamento de Microbiologı ´a. Escuela Nacional de Ciencias Biolo ´gicas, IPN. Casco de Santo Toma ´s. Me ´xico DF Abstract: The proteolytic system of the phytopath- ogenic and dimorphic fungus Ustilago maydis is not known. In this work, we report the presence of at least four proteases from two haploid strains of U. maydis . Activities of two proteinases pumA and pumB, aminopeptidase pumAPE, and dipeptidylam- inopeptidase pumDAP were measured under several nutritional and morphological conditions, including the yeast-mycelium transition. The activity of pumA was found in the intracellular and extracellular frac- tions, pumAi and pumAe, respectively. The latter ac- tivity was detected only during the yeast-mycelium di- morphic transition induced by growth at acid pH in a medium containing ammonium as the sole nitro- gen source. Activity of pumAe was partially inhibited by Pepstatin A, which also inhibited mycelium for- mation. Activity of pumAi was inhibited by this spe- cific inhibitor of aspartyl-proteases. Activity of pumB was detected in intracellular and extracellular frac- tions, mostly bound to an endogenous inhibitor, which was removed by treatment at acid pH. This fungus contains at least two soluble pumAPE, which might be metallo-proteases, because they were inhib- ited by EDTA and 1–10, phenanthroline. When the fungus was grown in media containing proline or corn infusion as the nitrogen source, an intracellular pumDAP activity was detected. No carboxypeptidase activity was found with N-benzoyl-L-tyrosine-4-nitroan- ilide as substrate in any of the conditions tested in any of the U. maydis strains analyzed. Accepted for publication August 31, 2002. 1 Corresponding author, E-mail: [email protected] Key words: acid proteinase, dimorphic phyto- pathogenic fungus, proteases, Ustilago maydis INTRODUCTION The heterobasidiomycete Ustilago maydis causes corn smut, a common disease in most parts of the world. In this pathosystem, the infection process is associ- ated with sexual development. In its saprophytic phase, the fungus grows in the form of haploid, bud- ding, yeast-like cells (sporidia). Sporidia of opposite mating types might fuse and give rise to the dikary- otic stage, which is the infective form of the fungus. In the host tissues, U. maydis grows as a septate my- celium, which produces diploid teliospores. These spores fill the galls that are characteristic of the dis- ease. Germination of teliospores involves the forma- tion of the promycelium and meiosis with the for- mation of basidiospores. Basidiospores reproduce by budding, completing the life cycle of the fungus (for review see Banuett 1995, Ruiz-Herrera and Martinez- Espinoza 1998). Ustilago maydis requires the maize plant to complete its sexual life cycle. It has been suggested that the plant produces diffusible com- pounds needed for completion of the sexual cycle of the fungus (for review see Banuett 1995). By control of pH in synthetic media, it is possible to control di- morphism of haploid cells of U. maydis. At neutral pH the fungus grows as a homogeneous population of sporidia, whereas at acidic pH the mycelial form developes (Ruiz-Herrera et al 1995). Proteolysis plays an important role in different physiological functions—protein digestion, hormone maturation, immune response, inflammation, coag- ulation, fertilization, germination and other morpho- logical processes (Holzer and Heinrich 1988). Some proteases are important in the formation and ger- mination of spores, in pathogenesis of several micro- organisms and in post-translational regulation (Yuan and Cole 1989, Vartivarian 1992, White and Agabian 1995, Monod et al 1998, Suarez-Rendueles et al 1991). Proteolysis is an essential life process. The study of Saccharomyces cerevisiae has been important in the elucidation of proteinase multiplicity and pro- teinase function in eukaryotic cells. Survival of cells in their natural environment depends highly on their ability to adapt to frequent changes. The ability of
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Proteinases and exopeptidases from the phytopathogenic fungus Ustilago maydis

327
Mycologia, 95(2), 2003, pp. 327–339.q 2003 by The Mycological Society of America, Lawrence, KS 66044-8897
Proteinases and exopeptidases from the phytopathogenic fungusUstilago maydis
Yuridia Mercado-FloresCesar Hernandez-Rodrıguez
Departamento de Microbiologıa. Escuela Nacional deCiencias Biologicas, IPN. Casco de Santo Tomas.Mexico DF
Jose Ruiz-HerreraDepartamento de Ingenierıa Genetica, UnidadIrapuato, Centro de Investigacion y EstudiosAvanzados, IPN. Irapuato, Guanajuato, Mexico
Lourdes Villa-Tanaca1
Departamento de Microbiologıa. Escuela Nacional deCiencias Biologicas, IPN. Casco de Santo Tomas.Mexico DF
Abstract: The proteolytic system of the phytopath-ogenic and dimorphic fungus Ustilago maydis is notknown. In this work, we report the presence of atleast four proteases from two haploid strains of U.maydis. Activities of two proteinases pumA andpumB, aminopeptidase pumAPE, and dipeptidylam-inopeptidase pumDAP were measured under severalnutritional and morphological conditions, includingthe yeast-mycelium transition. The activity of pumAwas found in the intracellular and extracellular frac-tions, pumAi and pumAe, respectively. The latter ac-tivity was detected only during the yeast-mycelium di-morphic transition induced by growth at acid pH ina medium containing ammonium as the sole nitro-gen source. Activity of pumAe was partially inhibitedby Pepstatin A, which also inhibited mycelium for-mation. Activity of pumAi was inhibited by this spe-cific inhibitor of aspartyl-proteases. Activity of pumBwas detected in intracellular and extracellular frac-tions, mostly bound to an endogenous inhibitor,which was removed by treatment at acid pH. Thisfungus contains at least two soluble pumAPE, whichmight be metallo-proteases, because they were inhib-ited by EDTA and 1–10, phenanthroline. When thefungus was grown in media containing proline orcorn infusion as the nitrogen source, an intracellularpumDAP activity was detected. No carboxypeptidaseactivity was found with N-benzoyl-L-tyrosine-4-nitroan-ilide as substrate in any of the conditions tested inany of the U. maydis strains analyzed.
Accepted for publication August 31, 2002.1 Corresponding author, E-mail: [email protected]
Key words: acid proteinase, dimorphic phyto-pathogenic fungus, proteases, Ustilago maydis
INTRODUCTION
The heterobasidiomycete Ustilago maydis causes cornsmut, a common disease in most parts of the world.In this pathosystem, the infection process is associ-ated with sexual development. In its saprophyticphase, the fungus grows in the form of haploid, bud-ding, yeast-like cells (sporidia). Sporidia of oppositemating types might fuse and give rise to the dikary-otic stage, which is the infective form of the fungus.In the host tissues, U. maydis grows as a septate my-celium, which produces diploid teliospores. Thesespores fill the galls that are characteristic of the dis-ease. Germination of teliospores involves the forma-tion of the promycelium and meiosis with the for-mation of basidiospores. Basidiospores reproduce bybudding, completing the life cycle of the fungus (forreview see Banuett 1995, Ruiz-Herrera and Martinez-Espinoza 1998). Ustilago maydis requires the maizeplant to complete its sexual life cycle. It has beensuggested that the plant produces diffusible com-pounds needed for completion of the sexual cycle ofthe fungus (for review see Banuett 1995). By controlof pH in synthetic media, it is possible to control di-morphism of haploid cells of U. maydis. At neutralpH the fungus grows as a homogeneous populationof sporidia, whereas at acidic pH the mycelial formdevelopes (Ruiz-Herrera et al 1995).
Proteolysis plays an important role in differentphysiological functions—protein digestion, hormonematuration, immune response, inflammation, coag-ulation, fertilization, germination and other morpho-logical processes (Holzer and Heinrich 1988). Someproteases are important in the formation and ger-mination of spores, in pathogenesis of several micro-organisms and in post-translational regulation (Yuanand Cole 1989, Vartivarian 1992, White and Agabian1995, Monod et al 1998, Suarez-Rendueles et al1991). Proteolysis is an essential life process. Thestudy of Saccharomyces cerevisiae has been importantin the elucidation of proteinase multiplicity and pro-teinase function in eukaryotic cells. Survival of cellsin their natural environment depends highly on theirability to adapt to frequent changes. The ability of

328 MYCOLOGIA
cells to respond to environmental extremes is vital totheir survival. Proteolysis plays an essential role inthis response to stress. Conditions of cellular stresscan be caused by starvation and physical or chemicalconditions, such as heat, extreme pH values or UVradiation. Nutritional stress probably leads to reor-ganization of cell metabolism and therefore to thereorganization of enzyme composition. The most ob-vious example of this form of stress occurs when adiploid yeast cell is transferred from a glucose-richmedium to a medium with a poor carbon source sup-plied by acetate. Under these conditions, and withremoval of nitrogen, the yeast cell begins to differ-entiate and four spores appear. Here, intensive pro-teolysis of unnecessary vegetative proteins providesthe cell with all amino acids required for new proteinsynthesis and probably for the generation of energy.As a further response to nutritional stress, yeast cellshave been reported to form pseudo-hyphae whenstarved of nitrogen. It has been suggested that thisfilamentous growth enables otherwise sessile cells toforage for nutrients (for review see Hilt and Wolf1992).
Although studies on several phytopathogenic fungisuggest the role of proteases in pathogenesis, the pro-teolytic system in U. maydis is unknown. The phyto-pathogenic fungi in which proteases have been de-scribed include: Botrytis cinerea (Urbanek and Ka-czmare 1985), Fusarium culmorum (Urbanek and Yir-daw 1984), Endothia parasitica (Choi et al 1993, Jaraet al 1996), Glomerella cingulata (Clark et al 1997)and Sclerotinia sclerotiorum (Poussereau et al 2001).
Detailed analyses of proteases in U. maydis mightlet us determine the proteases that this fungus pro-duces and to examine the production of these en-zymes in several nutritional conditions and duringthe dimorphic transition of yeast to mycelia, which isconsidered to be an important factor in the phyto-pathogenicity of this fungus. Moreover, it might bepossible to relate protease synthesis with the life cycleof this fungus and with its response to diffusible com-pounds produced by the host plant, Zea mays.
MATERIALS AND METHODS
Fungal strains. These strains and mating types of U. may-dis were used in this study: FB1 a1b1 and FB2 a2b2. Thesestrains kindly were provided by Dr. Flora Banuett, Universityof California at San Francisco, USA.
Culture media. The strains were maintained at 270 C in50% (v/v) glycerol. The cells were grown on complete Y PDmedium (1% yeast extract, 2% bactopeptone, 2% glucose)or minimal medium (0.17% yeast nitrogen base withoutamino acids and ammonium sulfate, plus 2% glucose) (Sua-rez-Rendueles et al 1991) with different sources of nitrogen:
2% peptone, 2% proline, 0.5% ammonium sulfate or corninfusion (equivalent to 4 mg of protein per mL of medi-um). The corn infusion was obtained by boiling for 20 min500 g of corncobs in 1 L of water and then filtering. Thissolution was supplemented with 0.17% yeast nitrogen basewithout amino acids and ammonium sulfate, plus 2% glu-cose.
Growth conditions. Cells inoculated into liquid mediumpromoted growth as a homogeneous population of bud-ding, yeast-like cells (culture pH 7.0) or as a population inthe mycelial form (pH 3.0) as previously described (Ruiz-Herrera et al 1995).
To measure cell growth, samples of the cultures were cen-trifuged at 20003 g. Sedimented cells were washed twicewith distilled water, and the protein content was measured.Changes in pH were recorded with a pH meter coupled toa recorder.
Enzyme assays. Crude cell-free extracts and soluble ex-tracts were prepared as described by Arbesu et al (1993).Enzymatic activities were determined as described by Hirschet al (1989). These substrates were used for the differentproteases: acid denatured hemoglobin for proteinase A ac-tivity (pumA); Hide Powder Azure (HPA) for proteinase Bactivity (pumB); L-lysyl-4-nitroanilide (Lys-4-NA) for ami-nopeptidase activity (pumAPE); L-alanyl-prolyl-4-nitroani-lide (Ala-pro-4-NA) for dipeptidylaminopeptidase activity(pumDAP); and N-benzoyl-L-tyrosine-4-nitroanilide (N-Bz-Tyr-4-NA) for carboxypeptidase activity (pumCP). Activa-tion of fresh crude extracts was carried out as described bySaheki and Holzer (1975). Lysine amino peptidase activityin polyacrylamide gels was detected with L-lysyl-b-napthy-lamide (Lys-b-NphA) and Fast Garnet as described byGarcıa-Alvarez et al (1991). As a source of crude inhibitorof pumB, we used cell-free extracts of the fungus boiled andcentrifuged as described by Escudero et al (1993). Enolaseactivity was determined with D-glyceric acid 2-phosphate assubstrate as described by Dmitriy and Nowak (1998) andwas used as marker enzyme for the intracellular localization.
Light and scanning electron microscopy. Cells were observedin unstained preparations by interference microscopy. Forscanning electron microscopy (SEM), cells were observedin a JEON model JSM-5800LV, cells were fixed with 1% (v/v) glutaraldehyde in 0.1 M sodium phosphate buffer, pH7.0. They were washed in a solution containing 0.10 MNa2HPO4 and 20% sucrose, pH 7.0, and post-fixed with 2%(w/v) osmium tetroxide. Cells were washed and dehydratedin a graded alcohol series. For SEM, the dehydrated cellswere mounted on specimen holders and coated with gold.The images were captured and digitized by a computer cou-pled to the microscope.
Miscellaneous. Protein content was estimated by the meth-od of Lowry et al (1951). Hemoglobin, Hide Powder Azure(HPA), L-aminoacyl-4-nitroanilides, and L-aminoacyl-b-nap-thylamides, (L-A-4NA; L-A-b-NphA) were used as proteinaseand peptidase substrates. They were obtained from eitherSigma Chemical Co. (St. Louis, MO, USA) or Bachem (Kingof Prussia, PA, USA). D-glyceric acid 2-phosphate (Sigma)was used as the enolase substrate. Growth media were from
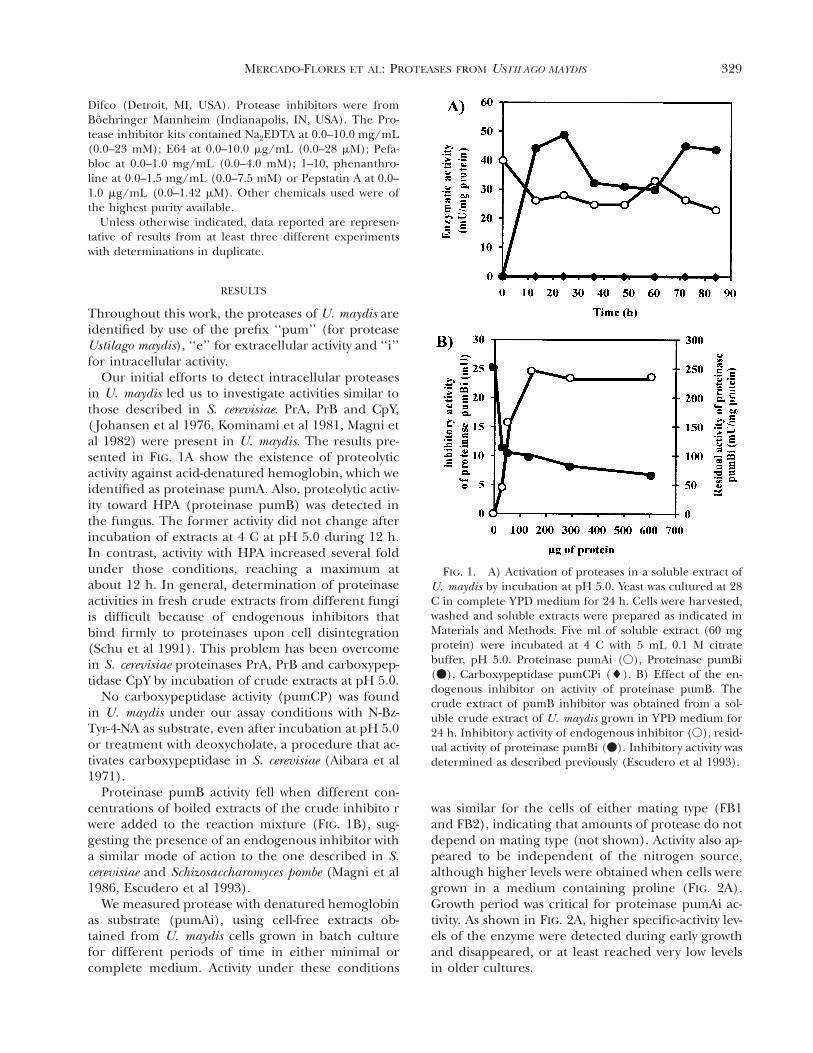
329MERCADO-FLORES ET AL: PROTEASES FROM USTILAGO MAYDIS
FIG. 1. A) Activation of proteases in a soluble extract ofU. maydis by incubation at pH 5.0. Yeast was cultured at 28C in complete Y PD medium for 24 h. Cells were harvested,washed and soluble extracts were prepared as indicated inMaterials and Methods. Five ml of soluble extract (60 mgprotein) were incubated at 4 C with 5 mL 0.1 M citratebuffer, pH 5.0. Proteinase pumAi (V), Proteinase pumBi(v), Carboxypeptidase pumCPi (l). B) Effect of the en-dogenous inhibitor on activity of proteinase pumB. Thecrude extract of pumB inhibitor was obtained from a sol-uble crude extract of U. maydis grown in Y PD medium for24 h. Inhibitory activity of endogenous inhibitor (V), resid-ual activity of proteinase pumBi (v). Inhibitory activity wasdetermined as described previously (Escudero et al 1993).
Difco (Detroit, MI, USA). Protease inhibitors were fromBoehringer Mannheim (Indianapolis, IN, USA). The Pro-tease inhibitor kits contained Na2EDTA at 0.0–10.0 mg/mL(0.0–23 mM); E64 at 0.0–10.0 mg/mL (0.0–28 mM); Pefa-bloc at 0.0–1.0 mg/mL (0.0–4.0 mM); 1–10, phenanthro-line at 0.0–1.5 mg/mL (0.0–7.5 mM) or Pepstatin A at 0.0–1.0 mg/mL (0.0–1.42 mM). Other chemicals used were ofthe highest purity available.
Unless otherwise indicated, data reported are represen-tative of results from at least three different experimentswith determinations in duplicate.
RESULTS
Throughout this work, the proteases of U. maydis areidentified by use of the prefix ‘‘pum’’ (for proteaseUstilago maydis), ‘‘e’’ for extracellular activity and ‘‘i’’for intracellular activity.
Our initial efforts to detect intracellular proteasesin U. maydis led us to investigate activities similar tothose described in S. cerevisiae. PrA, PrB and CpY,( Johansen et al 1976, Kominami et al 1981, Magni etal 1982) were present in U. maydis. The results pre-sented in FIG. 1A show the existence of proteolyticactivity against acid-denatured hemoglobin, which weidentified as proteinase pumA. Also, proteolytic activ-ity toward HPA (proteinase pumB) was detected inthe fungus. The former activity did not change afterincubation of extracts at 4 C at pH 5.0 during 12 h.In contrast, activity with HPA increased several foldunder those conditions, reaching a maximum atabout 12 h. In general, determination of proteinaseactivities in fresh crude extracts from different fungiis difficult because of endogenous inhibitors thatbind firmly to proteinases upon cell disintegration(Schu et al 1991). This problem has been overcomein S. cerevisiae proteinases PrA, PrB and carboxypep-tidase CpY by incubation of crude extracts at pH 5.0.
No carboxypeptidase activity (pumCP) was foundin U. maydis under our assay conditions with N-Bz-Tyr-4-NA as substrate, even after incubation at pH 5.0or treatment with deoxycholate, a procedure that ac-tivates carboxypeptidase in S. cerevisiae (Aibara et al1971).
Proteinase pumB activity fell when different con-centrations of boiled extracts of the crude inhibito rwere added to the reaction mixture (FIG. 1B), sug-gesting the presence of an endogenous inhibitor witha similar mode of action to the one described in S.cerevisiae and Schizosaccharomyces pombe (Magni et al1986, Escudero et al 1993).
We measured protease with denatured hemoglobinas substrate (pumAi), using cell-free extracts ob-tained from U. maydis cells grown in batch culturefor different periods of time in either minimal orcomplete medium. Activity under these conditions
was similar for the cells of either mating type (FB1and FB2), indicating that amounts of protease do notdepend on mating type (not shown). Activity also ap-peared to be independent of the nitrogen source,although higher levels were obtained when cells weregrown in a medium containing proline (FIG. 2A).Growth period was critical for proteinase pumAi ac-tivity. As shown in FIG. 2A, higher specific-activity lev-els of the enzyme were detected during early growthand disappeared, or at least reached very low levelsin older cultures.
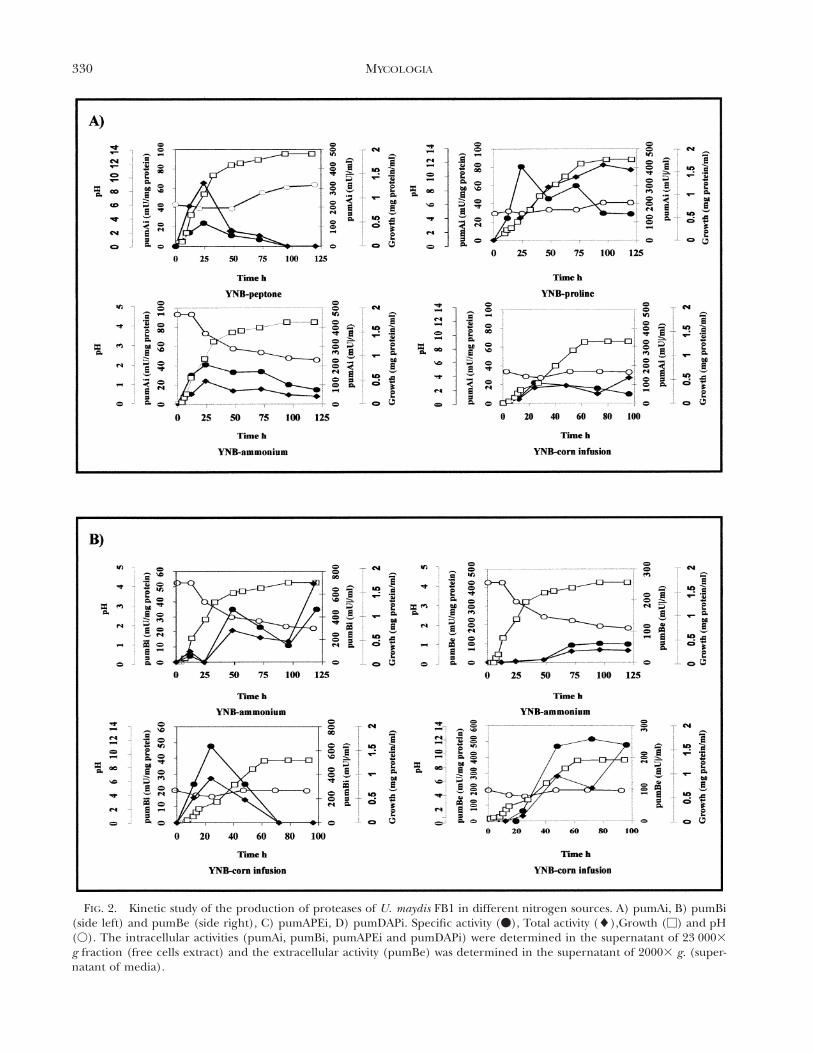
330 MYCOLOGIA
FIG. 2. Kinetic study of the production of proteases of U. maydis FB1 in different nitrogen sources. A) pumAi, B) pumBi(side left) and pumBe (side right), C) pumAPEi, D) pumDAPi. Specific activity (v), Total activity (l),Growth (M) and pH(V). The intracellular activities (pumAi, pumBi, pumAPEi and pumDAPi) were determined in the supernatant of 23 0003g fraction (free cells extract) and the extracellular activity (pumBe) was determined in the supernatant of 20003 g. (super-natant of media).
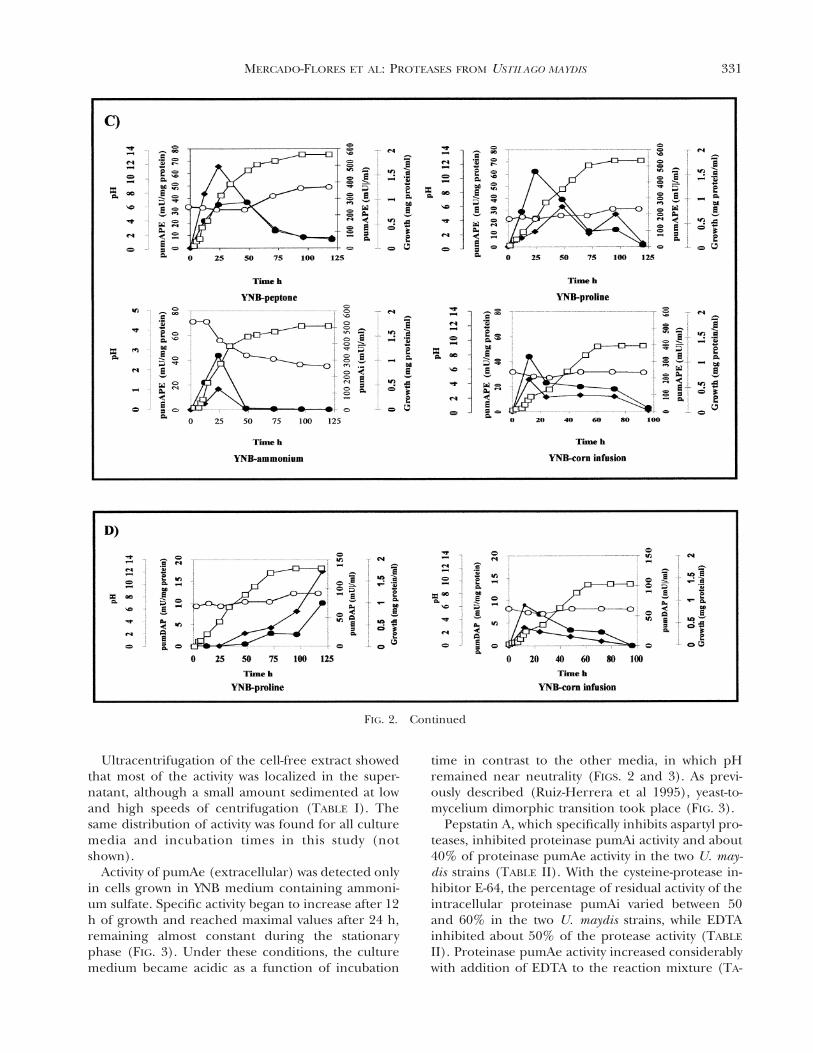
331MERCADO-FLORES ET AL: PROTEASES FROM USTILAGO MAYDIS
FIG. 2. Continued
Ultracentrifugation of the cell-free extract showedthat most of the activity was localized in the super-natant, although a small amount sedimented at lowand high speeds of centrifugation (TABLE I). Thesame distribution of activity was found for all culturemedia and incubation times in this study (notshown).
Activity of pumAe (extracellular) was detected onlyin cells grown in YNB medium containing ammoni-um sulfate. Specific activity began to increase after 12h of growth and reached maximal values after 24 h,remaining almost constant during the stationaryphase (FIG. 3). Under these conditions, the culturemedium became acidic as a function of incubation
time in contrast to the other media, in which pHremained near neutrality (FIGS. 2 and 3). As previ-ously described (Ruiz-Herrera et al 1995), yeast-to-mycelium dimorphic transition took place (FIG. 3).
Pepstatin A, which specifically inhibits aspartyl pro-teases, inhibited proteinase pumAi activity and about40% of proteinase pumAe activity in the two U. may-dis strains (TABLE II). With the cysteine-protease in-hibitor E-64, the percentage of residual activity of theintracellular proteinase pumAi varied between 50and 60% in the two U. maydis strains, while EDTAinhibited about 50% of the protease activity (TABLE
II). Proteinase pumAe activity increased considerablywith addition of EDTA to the reaction mixture (TA-
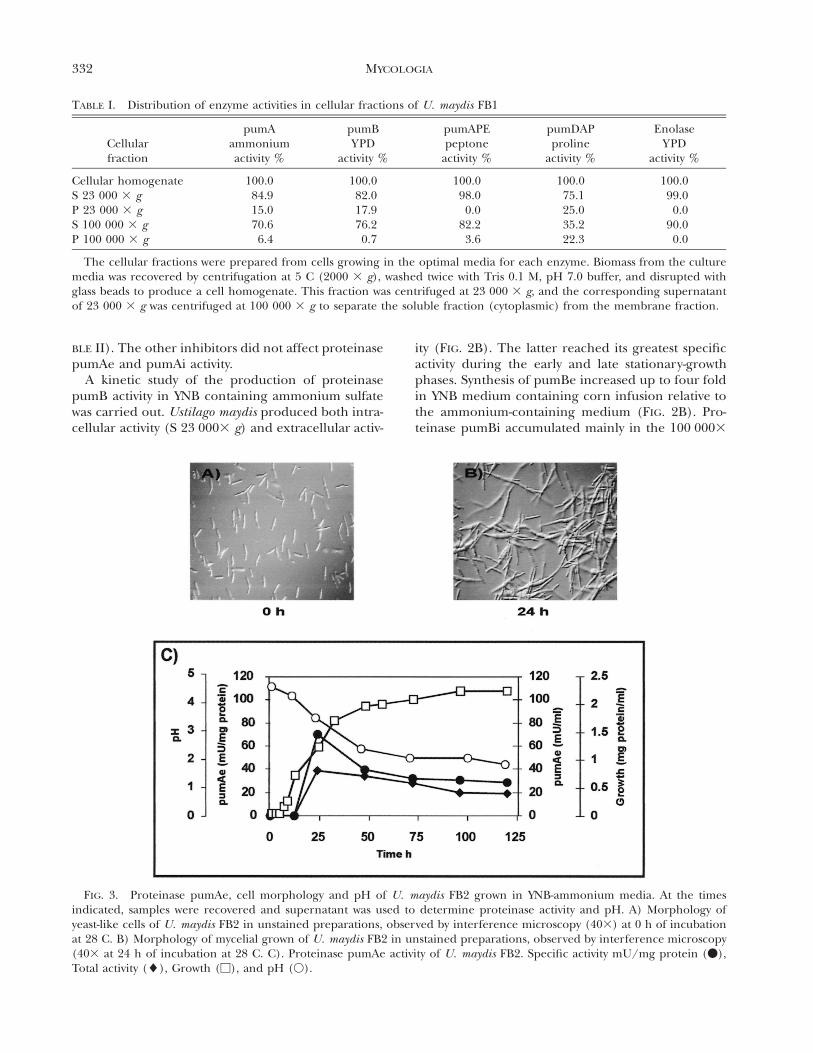
332 MYCOLOGIA
TABLE I. Distribution of enzyme activities in cellular fractions of U. maydis FB1
Cellularfraction
pumAammoniumactivity %
pumBY PD
activity %
pumAPEpeptone
activity %
pumDAPproline
activity %
EnolaseY PD
activity %
Cellular homogenateS 23 000 3 gP 23 000 3 gS 100 000 3 gP 100 000 3 g
100.084.915.070.66.4
100.082.017.976.20.7
100.098.00.0
82.23.6
100.075.125.035.222.3
100.099.00.0
90.00.0
The cellular fractions were prepared from cells growing in the optimal media for each enzyme. Biomass from the culturemedia was recovered by centrifugation at 5 C (2000 3 g), washed twice with Tris 0.1 M, pH 7.0 buffer, and disrupted withglass beads to produce a cell homogenate. This fraction was centrifuged at 23 000 3 g, and the corresponding supernatantof 23 000 3 g was centrifuged at 100 000 3 g to separate the soluble fraction (cytoplasmic) from the membrane fraction.
FIG. 3. Proteinase pumAe, cell morphology and pH of U. maydis FB2 grown in YNB-ammonium media. At the timesindicated, samples were recovered and supernatant was used to determine proteinase activity and pH. A) Morphology ofyeast-like cells of U. maydis FB2 in unstained preparations, observed by interference microscopy (403) at 0 h of incubationat 28 C. B) Morphology of mycelial grown of U. maydis FB2 in unstained preparations, observed by interference microscopy(403 at 24 h of incubation at 28 C. C). Proteinase pumAe activity of U. maydis FB2. Specific activity mU/mg protein (v),Total activity (l), Growth (M), and pH (V).
BLE II). The other inhibitors did not affect proteinasepumAe and pumAi activity.
A kinetic study of the production of proteinasepumB activity in YNB containing ammonium sulfatewas carried out. Ustilago maydis produced both intra-cellular activity (S 23 0003 g) and extracellular activ-
ity (FIG. 2B). The latter reached its greatest specificactivity during the early and late stationary-growthphases. Synthesis of pumBe increased up to four foldin YNB medium containing corn infusion relative tothe ammonium-containing medium (FIG. 2B). Pro-teinase pumBi accumulated mainly in the 100 0003

333MERCADO-FLORES ET AL: PROTEASES FROM USTILAGO MAYDIS
TABLE II. Effect of proteolytic inhibitors on U. maydis FB1 proteases
ProteaseInhibitors/Activators
Concentrationof inhibitor/
activatorPercentactivity
pumAe (S 2000 3 g) Pepstatin A (mg/ml)
EDTA (mg/ml)
0.00.71.00.05.0
10.0
100.060.754.7
100.0141.3213.8
pumAi (S 23 000 3 g) E-64 (mg/ml)
EDTA (mg/ml)
Pepstatin A (mg/ml)
0.05.0
10.00.05.0
10.00.00.71.0
100.072.962.3
100.058.940.9
100.00.00.0
pumAPE (S 10 000 3 g) EDTA (mg/ml)
1–10, phenantroline (mg/ml)
0.05.0
10.00.00.51.5
100.013.80.0
100.00.00.0
pumDAP (P 23 000 3 g) E-64 (mg/ml)
Pefabloc (mg/ml)
EDTA (mg/ml)
1–10, phenantroline (mg/ml)
0.05.0
10.00.00.5
10.00.05.0
10.00.00.51.5
100.074.949.5
100.045.830.1
100.00.00.0
100.00.00.0
pumDAP (P 100 000 3 g) Pefabloc (mg/ml) 0.00.5
10.0
100.022.10.0
Proteolytic activities were determined in different nitrogen sources. pumA (YNB-ammonium); pumAPE (YNB-peptone);pumDAP (YNB-corn infusion).
g supernatant fraction, although considerable activityalso was found in the fraction that sedimented at23 0003 g (TABLE I).
Activity of pumAPE (aminopeptidase) was foundonly in the supernatant fraction (S 23 0003 g) incells grown in all culture media. Similar levels of ac-tivity were found in both U. maydis strains (data notshown). Activity reached maximal values during theexponential growth phase of the fungus for all themedia used (FIG. 2C). Maximal activity was detectedin the 100 0003 g supernatant fraction, as with pumAand pumB (TABLE I). When cell-free extracts weresubjected to PAGE and pumAPE activity was revealed
in gels with the Lys-b-NphA substrate, two bands ofactivity were detected (FIG. 4). Activity of pumAPEwas blocked by two metalloprotease inhibitors, EDTAand 1–10, phenanthroline (TABLE II).
Protease pumDAP activity, detected with the chro-mogenic substrate Ala-pro-4-NA for dipeptidylami-nopeptidase, was observed in U. maydis strains (FB1and FB2). Highest levels of specific activity were ob-served during the late stationary-growth phase inYNB-proline medium (FIG. 2D). In contrast, whenYNB-corn infusion medium was used, highest levelsof activity were reached during the exponentialgrowth phase (FIG. 2D).

334 MYCOLOGIA
FIG. 4. Activity staining of aminopeptidase activitiesfrom U. maydis separated by non-denaturing polyacryl-amide gel electrophoresis. The gels were run at 87 V, andelectrophoresis was carried out for 2 h at 4 C. Aminopep-tidase activity staining was carried out by incubating the gelfor 30 min at 37 C with Tris-HCl 5 mM, pH 7.2, containing0.05% of Lys-b-NphA and 0.1% of Fast Garnet. A) 50 mL ofsoluble extract (S 100 0003 g) of U. maydis FB1 (315 mU/mL). B) 50 mL of soluble extract (S 100 0003 g) of U. may-dis FB2 (535 mU/mL).
In contrast to the other proteases, pumDAP activityfrom cells grown in either YNB-proline or YNB-corninfusion media was localized in the particulate frac-tion that sedimented at 23 0003 g (TABLE I). Activityof pumDAP in the particulate fraction that sedi-mented at 100 0003 g was inhibited by Pefabloc,which is a specific serine protease inhibitor (TABLE
II), but not by E-64, which is a cysteine- protease in-hibitor, EDTA or 1–10, phenanthroline (data notshown). On the other hand, pumDAP in the 23 0003g particulate fraction was only partially inhibited byPefabloc and E-64 and completely inhibited by EDTAor 1–10, phenanthroline (TABLE II).
In contrast to the presence of acid, alkaline andamino-peptidases, no carboxypeptidase activity wasdetected when extracts of either strain of U. maydiswere incubated with the synthetic chromogenic sub-strate N-Bz-Tyr-4-NA.
As shown in FIG. 3, proteinase pumAe was pro-duced only in YNB-ammonium medium after 12 h of
culture, when the fungus acidifies the medium andthe transition from yeast to mycelial form takes place.Levels of proteinase pumAe and pumAi activity weremeasured during development of the mycelial formin U. maydis. Proteinase pumAe in the two U. maydisstrains was formed only under the conditions thatgave rise to the mycelial phase (FIG. 5-2). This eventwas reversible in both strains of U. maydis. When cellswere transferred to a neutral pH medium, mycelialcells reverted to yeast-like growth and levels ofpumAe were reduced (FIG. 5-4).
As indicated above, proteinase pumAe was partiallyinhibited by Pepstatin A (TABLE II). To determinewhether this inhibitor had any effect on the dimor-phic transition of U. maydis, Pepstatin was added tothe culture medium at concentrations of 1 and 2 mg/mL. As a control, medium without inhibitor was in-cluded, as well as a culture in which the yeast phasewas maintained by incubation at pH 7.0. Microscopicobservations and determinations of activities of pro-teinase pumAe and pumAi were performed in allthese cultures. Activity of enolase was determined asan intracellular marker in supernatants of culturemedia; the absence of this enzyme demonstrated theintegrity of the cells (data not shown). Formation ofmycelium was partially inhibited when Pepstatin Awas added to cultures of either strain. Furthermore,the specific activity values of the intracellular and ex-tracellular acid proteinases were reduced comparedto those in the absence of the inhibitor (FIG. 6). Incontrast, cell viability was not affected by addition ofPepstatin A. (data not shown), and levels of pumBi,pumBe, pumAPE and pumDAP did not change (datanot shown). As previously indicated, no pumCP pro-teolytic activity was detected in these experimentswith N-Bz-Tyr-4-NA as substrate.
DISCUSSION
Ustilago maydis has been a useful model for the studyof relationships between phytopathogenic fungi andtheir host plants. It also has been used to study therelationship between dimorphic transition and phy-topathogenicity (Ruiz-Herrera et al 1995, Ruiz-Her-rera and Martinez-Espinoza 1998, Bolker 2001). How-ever, enzymatic activities involved in pathogenesisand the dimorphic transition are largely unknown.Therefore, it is not surprising that we have littleknowledge about the proteases produced by this fun-gus. In fact, the study of proteolytic enzymes in phy-topathogenic microorganisms has been neglected,and only a few reports on this topic have appearedin the recent literature.
Considering the importance of the role of prote-ases in multiple aspects of fungal biology ( Jones
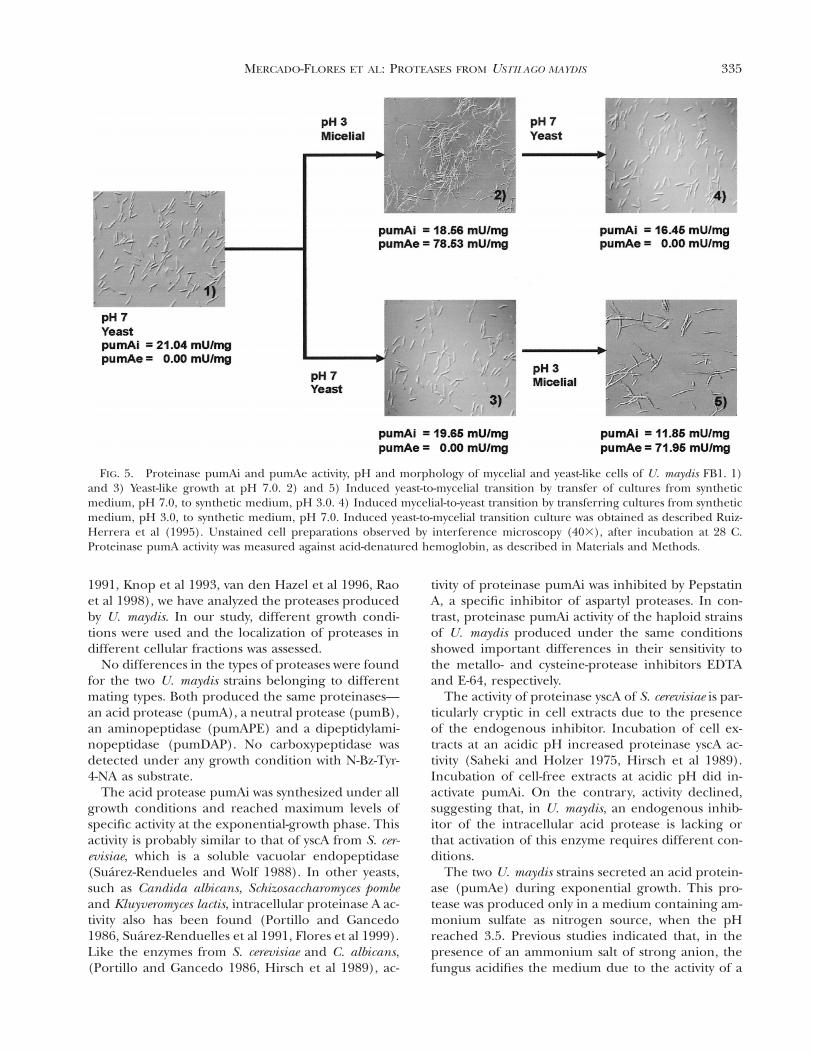
335MERCADO-FLORES ET AL: PROTEASES FROM USTILAGO MAYDIS
FIG. 5. Proteinase pumAi and pumAe activity, pH and morphology of mycelial and yeast-like cells of U. maydis FB1. 1)and 3) Yeast-like growth at pH 7.0. 2) and 5) Induced yeast-to-mycelial transition by transfer of cultures from syntheticmedium, pH 7.0, to synthetic medium, pH 3.0. 4) Induced mycelial-to-yeast transition by transferring cultures from syntheticmedium, pH 3.0, to synthetic medium, pH 7.0. Induced yeast-to-mycelial transition culture was obtained as described Ruiz-Herrera et al (1995). Unstained cell preparations observed by interference microscopy (403), after incubation at 28 C.Proteinase pumA activity was measured against acid-denatured hemoglobin, as described in Materials and Methods.
1991, Knop et al 1993, van den Hazel et al 1996, Raoet al 1998), we have analyzed the proteases producedby U. maydis. In our study, different growth condi-tions were used and the localization of proteases indifferent cellular fractions was assessed.
No differences in the types of proteases were foundfor the two U. maydis strains belonging to differentmating types. Both produced the same proteinases—an acid protease (pumA), a neutral protease (pumB),an aminopeptidase (pumAPE) and a dipeptidylami-nopeptidase (pumDAP). No carboxypeptidase wasdetected under any growth condition with N-Bz-Tyr-4-NA as substrate.
The acid protease pumAi was synthesized under allgrowth conditions and reached maximum levels ofspecific activity at the exponential-growth phase. Thisactivity is probably similar to that of yscA from S. cer-evisiae, which is a soluble vacuolar endopeptidase(Suarez-Rendueles and Wolf 1988). In other yeasts,such as Candida albicans, Schizosaccharomyces pombeand Kluyveromyces lactis, intracellular proteinase A ac-tivity also has been found (Portillo and Gancedo1986, Suarez-Renduelles et al 1991, Flores et al 1999).Like the enzymes from S. cerevisiae and C. albicans,(Portillo and Gancedo 1986, Hirsch et al 1989), ac-
tivity of proteinase pumAi was inhibited by PepstatinA, a specific inhibitor of aspartyl proteases. In con-trast, proteinase pumAi activity of the haploid strainsof U. maydis produced under the same conditionsshowed important differences in their sensitivity tothe metallo- and cysteine-protease inhibitors EDTAand E-64, respectively.
The activity of proteinase yscA of S. cerevisiae is par-ticularly cryptic in cell extracts due to the presenceof the endogenous inhibitor. Incubation of cell ex-tracts at an acidic pH increased proteinase yscA ac-tivity (Saheki and Holzer 1975, Hirsch et al 1989).Incubation of cell-free extracts at acidic pH did in-activate pumAi. On the contrary, activity declined,suggesting that, in U. maydis, an endogenous inhib-itor of the intracellular acid protease is lacking orthat activation of this enzyme requires different con-ditions.
The two U. maydis strains secreted an acid protein-ase (pumAe) during exponential growth. This pro-tease was produced only in a medium containing am-monium sulfate as nitrogen source, when the pHreached 3.5. Previous studies indicated that, in thepresence of an ammonium salt of strong anion, thefungus acidifies the medium due to the activity of a
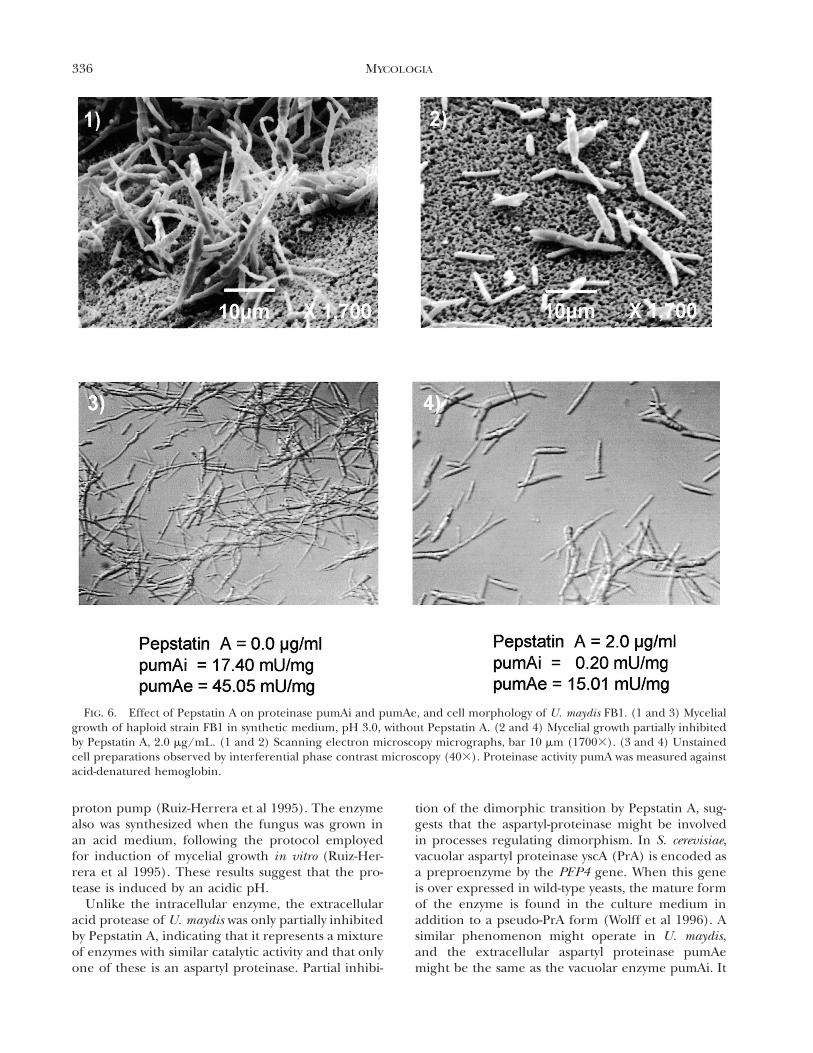
336 MYCOLOGIA
FIG. 6. Effect of Pepstatin A on proteinase pumAi and pumAe, and cell morphology of U. maydis FB1. (1 and 3) Mycelialgrowth of haploid strain FB1 in synthetic medium, pH 3.0, without Pepstatin A. (2 and 4) Mycelial growth partially inhibitedby Pepstatin A, 2.0 mg/mL. (1 and 2) Scanning electron microscopy micrographs, bar 10 mm (17003). (3 and 4) Unstainedcell preparations observed by interferential phase contrast microscopy (403). Proteinase activity pumA was measured againstacid-denatured hemoglobin.
proton pump (Ruiz-Herrera et al 1995). The enzymealso was synthesized when the fungus was grown inan acid medium, following the protocol employedfor induction of mycelial growth in vitro (Ruiz-Her-rera et al 1995). These results suggest that the pro-tease is induced by an acidic pH.
Unlike the intracellular enzyme, the extracellularacid protease of U. maydis was only partially inhibitedby Pepstatin A, indicating that it represents a mixtureof enzymes with similar catalytic activity and that onlyone of these is an aspartyl proteinase. Partial inhibi-
tion of the dimorphic transition by Pepstatin A, sug-gests that the aspartyl-proteinase might be involvedin processes regulating dimorphism. In S. cerevisiae,vacuolar aspartyl proteinase yscA (PrA) is encoded asa preproenzyme by the PEP4 gene. When this geneis over expressed in wild-type yeasts, the mature formof the enzyme is found in the culture medium inaddition to a pseudo-PrA form (Wolff et al 1996). Asimilar phenomenon might operate in U. maydis,and the extracellular aspartyl proteinase pumAemight be the same as the vacuolar enzyme pumAi. It

337MERCADO-FLORES ET AL: PROTEASES FROM USTILAGO MAYDIS
also is possible that culture conditions in our studiesresulted in overexpression and secretion of the en-zyme.
Proteolysis plays an essential role in responses tostress, such as nutrient deficiency, high temperature,extreme pH, UV irradiation or the presence of heavymetal ions or amino acid analogues. Nutritionalstress probably induces a reorganization of cellularmetabolism and existing enzymes (Hilt and Wolf1992). Under the protocol used to obtain the myce-lial phase of U. maydis, it is possible that the cells arestressed and that the acidic pH induces the synthesisof pumAe.
Extracellular aspartyl proteases have been reportedin other phytopathogenic fungi. For example, in Bo-trytis cinerea an aspartyl protease has been found tobe involved in the hydrolysis of proteins in apple cellwalls (Urbanek and Kaczmarek 1985). Likewise, Fu-sarium culmorum produces an acid protease that hy-drolyzes numerous vegetable proteins (Urbanek andYirdaw 1984). Also, Glomerella cingulata has an aspar-tyl protease whose transcript is induced by exogenousproteins and repressed by ammonium salts (Clark etal 1997). Cryphonectria parasitica secretes acid pro-teases, which have been purified and characterized,and their genes have been cloned (eapA, eapB andeapC) (Choi et al 1993, Jara et al 1996). It is possiblethat U. maydis synthesizes more than one extracel-lular acid protease, because as mentioned above itwas only partially inhibited by Pepstatin A.
Proteinase pumAe of the two U. maydis strains test-ed was activated by EDTA. This observation suggeststhe existence of another protease that participates inthe regulation of this enzyme that might be inhibitedby EDTA.
As indicated above, concomitant with the dimor-phic transition of yeast to mycelium, the activity ofthis enzyme increases, and maximum specific activitylevels are reached when only mycelia are found inthe culture. Because an acidic pH develops underthese conditions, we suggest that the enzyme mightbe regulated by pH, as has been reported for otherfungal proteases (for review, see Denison 2000). Nev-ertheless, additional evidence suggests a possible re-lationship between pumAe and the filamentation ofthe fungus. The most appealing observation is that,when Pepstatin A was added to cultures maintainedat pH 3.0, the formation of mycelium was inhibited.
An opposite, but related, example of pH-con-trolled morphogenesis is C. albicans. Unlike U. may-dis, C. albicans grows in the yeast form when culturedin acidic pH; whereas neutral pH favors growth ofmycelium (Soll 1985). Interestingly, however, expres-sion of vacuolar aspartyl proteinase of this yeast isinduced in the yeast form by growth at acidic pH,
but the evidence suggests that this enzyme has nodirect role in the dimorphic transition of the organ-ism (Niimi et al 1997).
On the other hand, the secreted aspartyl protein-ases SAP 4, 5 and 6 that are produced by C. albicans,seem to be implicated in the differentiation that oc-curs during the formation of the germination tube(Hube et al 1994). This could be the case in U. may-dis, in which aspartyl proteinase pumAi is not asso-ciated with the dimorphism events of this fungus,whereas pumAe might be involved in the phenome-non.
Ustilago maydis contains a second extracellular pro-teinase that we called pumBe, which is active againstthe HPA substrate. Highest levels of the enzyme werefound in media containing ammonium salts or corninfusion. Unlike the pH of media containing am-monium, the pH of corn infusion media reaches avalue of 6.0. In a similar way to that described forproteinase yscB of S. cerevisiae (Magni et al 1986,Hirsch et al 1989), pumBi activity in U. maydis is as-sociated with an endogenous inhibitor, because de-tection of activity of this enzyme required incubationof the extracts at pH 5.0. Also in S. pombe, Saccharo-myces carlsbergensis and C. albicans, there are specificendogenous inhibitors of this type of protein (Ma-tern et al 1979, Farley et al 1986, Escudero et al1993).
In both U. maydis strains used, pumBi activity wasfound in YNB-ammonium medium during the earlyand late stationary phases, whereas in the YNB-corninfusion medium, the intracellular activity reached itsmaximum value during the exponential-growthphase. Similarly, in S. cerevisiae expression of thegene coding for the vacuolar proteinase yscB is reg-ulated by nitrogen or carbon sources and dependson the growth phase of the yeast (Naik et al 1997).The activity of proteinase pumB does not seem to beassociated with any events during the dimorphic tran-sition of yeast to mycelia in the two U. maydis strains.
Both U. maydis strains contained at least two sol-uble pumAPEi activities that reached maximal activ-ities during the exponential growth phase and wereinhibited by EDTA and 1–10, phenanthroline. Pro-tease pumAPE, like proteinase pumB, is not clearlyassociated with any of the dimorphic transition phas-es in the U. maydis strains.
Another protease detected in the two U. maydisstrains was pumDAPi. In YNB-proline medium, high-est levels of this enzyme were reached during the latestationary phase, whereas optimal levels were detect-ed during the exponential-growth phase in the YNB-corn infusion medium. This stimulatory effect ofcorn infusion is interesting, because it might be dueto a thermo-stable low molecular-weight compound

338 MYCOLOGIA
from the host. That this effect is involved in the host-pathogen interaction is an attractive possibility.
Activity of pumDAP was localized both in the cy-tosol and the membrane fractions. Activity ofpumDAP in the membrane fraction was inhibited byPefabloc, suggesting that it is a serine protease. It isinteresting to note that the dipeptidylaminopepti-dase of S. cerevisiae, called yscIV, is a membrane ser-ine exopeptidase that participates in the processingof the pheromone (Suarez-Rendueles and Wolf1988). A similar role for that membrane-boundpumDAP in U. maydis would be an intriguing.
The pumDAP in the pellet obtained at low speedwas inhibited by EDTA and 1–10, phenanthrolineand partially inhibited by the serine and cysteine pro-tease inhibitors, Pefabloc and E-64, respectively. Thispattern of inhibition suggests the existence of a mix-ture of enzymes with the same catalytic activity, oneof them a serine-metallo-protease and the other a cys-teine-metallo-protease. S. cerevisiae produces dipep-tidylaminopeptidase yscII, a metalloprotease whoselocation and function are unknown (Suarez-Rendue-les and Wolf 1988).
Activity of pumDAPi was detected in both the yeastand mycelial forms of U. maydis without significantdifferences in their specific activities. This result sug-gests that the enzyme(s) is (are) not involved in thedimorphic transition of the fungus.
At least seven carboxypeptidases have been de-scribed in S. cerevisiae (Suarez-Rendules and Wolf1988). In contrast, no carboxypeptidase activity wasfound in our analysis of U. maydis. This result doesnot appear to be due to the presence of an endoge-nous inhibitor, such as that described for S. cerevisiae(Lenney 1975). Addition of sodium deoxycholate tothe reaction mixture, as well as incubation at an acid-ic pH, failed to reveal any carboxypeptidase activity.Hydrophobic properties of the surface of U. maydisdepend on the proteolysis of Rep1. This protein mustbe processed by proteases to generate small peptidesthat are important in the formation of hydrophobicaerial hyphae, which enhance the contact betweenthe fungus and the plant when infection is initiated(Wosten et al 1996). To date, the only protease de-scribed that is responsible for processing Rep1 is aprotease that is equivalent or similar to Kex2 of S.cerevisiae (Park et al 1994), which has been describedin the processing of the pheromone and killer factorin this yeast. This membrane peptidase cleaves thecarboxyl terminus of synthetic substrates (Suarez-Rendueles and Wolf 1988). Accordingly, either Rep1is processed by a different mechanism in U. maydisor the carboxypeptidase from this fungus is not activeagainst the synthetic substrate used in this study.
ACKNOWLEDGMENTS
The authors acknowledge Dr. Alfredo Martınez-Espinozaand Miss Claudia Leon-Ramırez for their valuable advice.M. C. Yuridia Mercado-Flores was a fellow from CONACyTand PIFI, IPN. This work was partially supported by grantsfrom COFAA and EDD, Instituto Politecnico Nacional toDr. Lourdes Villa-Tanaca and Dr. Cesar Hernandez-Rodrı-guez and from CONACYT to Dr. Jose Ruiz-Herrera
LITERATURE CITED
Aibara S, Hayashi R, Hata T. 1971. Physical and chemicalproperties of yeast proteinase C. Arg Biol Chem 35:658–666.
Arbesu MJ, Valle E, Suarez-Rendueles P. 1993. Purificationand characterization of aminopeptidase yspI from Schi-zosaccharomyces pombe. Yeast 9:637–644.
Banuett F. 1995. Genetics of Ustilago maydis, a fungal path-ogen that induces tumors in maize. Annu Rev Genet29:179–208.
Bolker M. 2001. Ustilago maydis, a valuable model systemfor the study of fungal dimorphism and virulence. Mi-crobiology 147:1395–1401.
Choi GH, Pawlyk DM, Rae B, Shapira R, Nuss DL. 1993.Molecular analysis and overexpression of the gene en-coding endothiapepsin, an aspartic protease from Cry-phonectria parasitica. Gene 125:135–141.
Clark SJ, Templeton MD, Sullivan PA. 1997. A secreted as-partic proteinase from Glomerella cingulata: purifica-tion of enzyme and molecular cloning of the cDNA.Microbiology 143:1395–1403.
Denison SH. 2000. pH regulation of gene expression in fun-gi. Fungal Genet Biol 29:61–71.
Dmitriy AV, Nowak T. 1998. pH dependence of the reactioncatalyzed by yeast Mg-enolase. Biochemistry 37:15238–15246.
Escudero B, Parra F, Suarez-Rendueles P. 1993. Purificationand characterization of the endogenous inhibitor forproteinase B from Schizosaccharomyces pombe. Biochim-ie 75:855–859.
Farley PC, Shepherd MG, Sullivan PA. 1986. The purifica-tion and properties of yeast proteinase B from Candidaalbicans. Biochem J 236:177–184.
Flores MV, Cuellas A, Voget CE. 1999. The proteolytic sys-tem of the yeast Kluyveromyces lactis. Yeast 15:1437–1448.
Garcıa-Alvarez N, Cueva R, Suarez-Rendueles P. 1991. Mo-lecular cloning of soluble aminopeptidase from Saccha-romyces cerevisiae. Sequence analysis of aminopeptidaseyscII, a putative zinc-metallopeptidase. Eur J Biochem202:993–1002.
Hilt W, Wolf DH. 1992. Stress-induced proteolysis in yeast.Mol Microbiol 6:2437–2442.
Hirsch HH, Suarez-Rendueles P, Wolf DH. 1989. Yeast (Sac-charomyces cerevisiae) proteinases: Structure, character-istics and functions. In: Walton EF, Yarranton GT, eds.The molecular and cell biology of yeasts, London:Blackie & Sons Ltd. p 134–200.

339MERCADO-FLORES ET AL: PROTEASES FROM USTILAGO MAYDIS
Holzer H, Heinrich PC. 1988. Control of proteolysis. AnnuRev Biochem 49:63–91.
Hube D, Monod M, Schofield DA, Brown AJP, Gow NAR.1994. Expression of seven members of the gene familyencode secretory aspartyl proteinases in Candida albi-cans. Mol Microbiol 14:87–99.
Jara P, Gilbert S, Delmas P, Guillemot JC, Kaghad M, FerraraP, Loison G. 1996. Cloning and characterization of theeapB y eapC genes of Cryphonectria parasitica encodingtwo new acid proteinases, and disruption of eapC. MolGen Genet 250:97–105.
Johansen JT, Breddam K, Ottesen M. 1976. Isolation of car-boxypeptidase Y by chromatography. Carlsberg ResCommun 41:1–14.
Jones EW. 1991. Three proteolytic system in the yeast Sac-charomyces cerevisiae. J Biol Chem 266:7963–7966.
Knop M, Schiffer HH, Rupp S, Wolf DH. 1993. Vacuolar/lysosomal proteolysis: proteases, substrates, mecha-nisms. Curr Opin Cell Biol 5:990–996.
Kominami E, Hoffschulte H, Holzer H. 1981. Purificationand properties of proteinase B from yeast. BiochimBiophys Acta 661:124–135.
Lenney JF. 1975. Three yeast proteins that specifically in-hibit yeast proteases A, B and C. J Bacteriol 122:1265–1273.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. 1951. Pro-tein measurement with the Folin phenol reagent. J BiolChem 193:265–275.
Magni G, Drewniak M, Santarelli I, Huang CY. 1986. Re-examination of the activation of yeast proteinase B atpH 5: loss of inhibition effect of proteinase B inhibi-tors. Biochem Int 12:557–565.
———, Natalini P, Santarlli I, Vita A. 1982. Baker’s yeastprotease A. Purification and enzymatic and molecularproperties. Arch Biochem Biophys 254:426–433.
Matern H, Weiser U, Holzer H. 1979. Isolation, character-ization and localization of the proteinase B inhibitorIB3 from Saccharomyces carlsbergensis. Eur J Biochem101:325–332.
Monod M, Hube B, Hess D, Sanglard D. 1998. Differentialregulation of SAP8 and SAP9, which encode two newmembers of the secreted aspartic proteinases family inCandida albicans. Microbiology 144:2731–2737.
Naik RR, Nebes V, Jones EW. 1997. Regulation of proteinaseB structural gene PBR1 in Saccharomyces cerevisiae. JBacteriol 179:1469–1474.
Niimi M, Niimi K, Cannon RD. 1997. Temperature expres-sion of the vacuolar aspartic proteinase (APR1) genebeta-N-acetylglucosaminidase (HEX1) gene duringCandida albicans morphogenesis. FEBS Microbiol Lett148:247–254.
Park CM, Bruenn JA, Ganesa C, Flurkey WF, Bozarth RF,Koltin Y. 1994. Structure and heterologous expressionof the Ustilago maydis viral toxin KP4. Mol Microbiol11:155–164.
Portillo F, Gancedo C. 1986. Purification and properties of
three intracellular proteinases from Candida albicans.Biochem Biophys Acta 881:229–235.
Poussereau N, Creton S, Brillon-Grand G, Rascle C, FevreM. 2001. Regulation of acp1, encoding a non-aspartylacid protease expressed during pathogenesis of Sclero-tinia sclerotiorum. Microbiology 147:717–726.
Rao MB, Tanksale AM, Ghatge MS, Deshpnade VV. 1998.Molecular and biotechnological aspects of microbialproteases. Microbiol Mol Biol Rev 62:597–635.
Ruiz-Herrera J, Leon GC, Guevara LO, Carabez AT. 1995.Yeast-mycelial dimorphism of haploid and diploidstrains of Ustilago maydis. Microbiology 141:695–703.
———, Martınez-Espinoza A. 1998. The fungus Ustilagomaydis, from Aztec cuisine to the research laboratory.Int Microbiol 1:149–158.
Saheki T, Holzer H. 1975. Proteolytic activities in yeast.Biochem Biophys Acta 348:203–214.
Schu P, Suarez-Rendueles P, Wolf DH. 1991. The proteinaseyscB inhibitor ( ) gene of yeast and studies on theBI2
function of its protein product. Eur J Biochem 197:1–7.
Soll DR. 1985. Candida albicans. In: Szaniszlo PJ, ed. FungalDimorphism with Emphasis on Fungi Pathogenic forHumans. New York: Plenum Press. p 167–195.
Suarez-Rendueles P, Wolf DH. 1988. Proteinase function inyeast: biochemical and genetic approaches to a centralmechanism of post-translational control in the euka-riotic cell. FEMS Microbiol Lett 54:17–46.
———, Villa L, Arbesu MJ, Escudero B. 1991. The proteo-lytic system of the fission yeast Schizosaccharomyces pom-be. FEMS Microbiol Lett 81:215–220.
Urbanek H, Kaczmarek A. 1985. Extracellular proteinasesof the isolate of Botrytis cinerea virulent to apple tissues.Acta Biochim Pol 32:101–109.
———, Yirdaw G. 1984. Hydrolytic ability of acid proteaseof Fusarium culmorum and role in phytopatogenesis.Acta Microbiol Pol 33:131–136.
van den Hazel HB, Kielland-Brandt MC, Winter JR. 1996.Review: biosynthesis and function of yeast vacuolar pro-teases. Yeast 12:1–16.
Vartivarian SE. 1992. Virulence properties and nonimmunepathogenic mechanism. Clin Infect Dis 14:30–36.
White C, Agabian N. 1995. Candida albicans secreted as-partyl proteinases: isoenzyme pattern is determined byenvironmental factors. J Bacteriol 177:5215–5221.
Wolff AM, Din N, Peterson JG. 1996. Vacuolar and extra-cellular maturation of Saccharomyces cerevisiae pro-teinase A. Yeast 12:823–832.
Wosten ABH, Bohlmann R, Eckerskorn C, Lottspeich F,Bolker M, Kahmann R. 1996. A novel class of smallamphipathic peptides affect aerial hyphal growth andsurface hydrophobicity in Ustilago maydis. EMBO J 15:4274–4281.
Yuan L, Cole GT. 1989. Characterization of proteinase in-hibitors isolated from fungal pathogen Coccidiodes im-mitis. J Biochem 257:729–736.














