
Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi
34
Atta-ur-Rahman (Ed.) Studies in Natural Products Chemistry, Vol. 25 © 2001 Elsevier Science B.V. Allrightsreserved. 293 METABOLISM OF THE TOMATO SAPONIN a-TOMATBVE BY PHYTOPATHOGENIC FUNGI MANUEL RUIZ-RUBIO, ALONSO PEREZ-ESPINOSA, KHALID LAIRINI, TERESA ROLDAN-ARJONA, ANTONIO DIPIETRO AND NURIA ANAYA Departamento de Genetica, Facultad de Ciencias, Universidad de Cordoba, 14071 Cordob a^ Spain ABSTRACT: a-Tomatine is a steroidal glycoalkaioid present in Solanaceoiis plants, principally in a number of Lycopersicon and Solanum species. High concentrations are found in leaves, stems, roots and green fruit of tomato plants, suggesting a possible role in resistance to pathogens. The toxic effects of a-tomatine are attributed to its ability to complex with membrane sterols, causing pore formation and leakage of cell contents. Pathogenic fungi to tomato are usually less sensitive to tomatine than pathogens of other plant species and saprophytes. Fungi have evolved two principal mechanisms to avoid the toxic effect of tomatine, either changing the composition of the cell membrane or producing specific tomatine-detoxifying enzymes, known as tomatinases. Tomatinases have distinct mechanisms of action, and molecular and catalytic properties in different pathogen species. Up to now, the knowledge of a-tomatine metabolism by phytopathogenic fungi is almost exclusively limited to the removal of sugar residues from the molecule, although recent studies in Gibberella pulicaris demonstrate that further hydroxylations of the aglycone moiety tomatidine occur, after the action of tomatinase. Tomatinase-encoding genes have been cloned from Septoria lycopersici and Fusariiim oxysporum. Both genes do not share sequence homology, suggesting that different tomato pathogens have independently developed the ability to cleave tomatine by convergent evolution. Expression of F. oxysporum tomatinase is fully repressed in the presence of glucose, indicating a possible role in nutrition besides detoxification of tomatine, although in BoUytis cinerea glucose repression of tomatinase is not observed. Other genes different to tomatinase are induced by a-tomatine, but the complete significance of this phenomenon is still unknown. INTRODUCTION Plants have evolved different mechanisms to protect themselves against a great variety of invasive pathogens. As a part of the defense, plants produce a number of secondary metabolites with antimicrobial activity. Many of these compounds are produced constitutively, being present in healthy plants as normal metabolic products. VanEtten et al proposed the term "phytoanticipin" to define these preformed antimicrobial
Transcript of Metabolism of the tomato saponin α-tomatine by phytopathogenic fungi

Atta-ur-Rahman (Ed.) Studies in Natural Products Chemistry, Vol. 25 © 2001 Elsevier Science B.V. All rights reserved. 293
METABOLISM OF THE TOMATO SAPONIN a-TOMATBVE BY PHYTOPATHOGENIC FUNGI
MANUEL RUIZ-RUBIO, ALONSO PEREZ-ESPINOSA, KHALID LAIRINI, TERESA ROLDAN-ARJONA, ANTONIO DIPIETRO AND
NURIA ANAYA
Departamento de Genetica, Facultad de Ciencias, Universidad de Cordoba, 14071 Cor dob a^ Spain
ABSTRACT: a-Tomatine is a steroidal glycoalkaioid present in Solanaceoiis plants, principally in a number of Lycopersicon and Solanum species. High concentrations are found in leaves, stems, roots and green fruit of tomato plants, suggesting a possible role in resistance to pathogens. The toxic effects of a-tomatine are attributed to its ability to complex with membrane sterols, causing pore formation and leakage of cell contents. Pathogenic fungi to tomato are usually less sensitive to tomatine than pathogens of other plant species and saprophytes. Fungi have evolved two principal mechanisms to avoid the toxic effect of tomatine, either changing the composition of the cell membrane or producing specific tomatine-detoxifying enzymes, known as tomatinases. Tomatinases have distinct mechanisms of action, and molecular and catalytic properties in different pathogen species. Up to now, the knowledge of a-tomatine metabolism by phytopathogenic fungi is almost exclusively limited to the removal of sugar residues from the molecule, although recent studies in Gibberella pulicaris demonstrate that further hydroxylations of the aglycone moiety tomatidine occur, after the action of tomatinase. Tomatinase-encoding genes have been cloned from Septoria lycopersici and Fusariiim oxysporum. Both genes do not share sequence homology, suggesting that different tomato pathogens have independently developed the ability to cleave tomatine by convergent evolution. Expression of F. oxysporum tomatinase is fully repressed in the presence of glucose, indicating a possible role in nutrition besides detoxification of tomatine, although in BoUytis cinerea glucose repression of tomatinase is not observed. Other genes different to tomatinase are induced by a-tomatine, but the complete significance of this phenomenon is still unknown.
INTRODUCTION
Plants have evolved different mechanisms to protect themselves against a great variety of invasive pathogens. As a part of the defense, plants produce a number of secondary metabolites with antimicrobial activity. Many of these compounds are produced constitutively, being present in healthy plants as normal metabolic products. VanEtten et al proposed the term "phytoanticipin" to define these preformed antimicrobial

294
compounds as opposed to phytoalexins, which are synthesized de novo from separate precursors in response to pathogen attack [1]. In contrast to phytoalexins, phytoanticipins have received comparatively little attention by phytopatologists, although in many cases they may represent one of the first chemical barriers encountered by potential pathogens.
Saponins are found in many major food crops and include a large number of examples of constitutive plant compounds with antimicrobial activity. Their name refers to the property of forming a stable foam when shaken with water, and they consist of triterpenoid, steroid or steroidal glycoalkaloid molecules bearing one or more sugar chains. Excellent general reviews on saponins have been published recently [2-6].
The steroidal glycoalkaloid a-tomatine (from hereon called tomatine) is the major saponin component of tomato (reviewed by Roddick [7]), and has also been found in other Solanaceous plants [7, 8], (reviewed by Maga [9]) The widespread distribution of tomatine in all parts of the tomato plant has stimulated searches for a function of this compound in the general plant metabolism, but apart from the suggestion that it may offer protection against herbivores and pathogens, little evidence exists for a biochemical or physiological function. Tomatine was first isolated from the wild tomato species Lycopersicon pimpinellifolium and from the cultivated variety Lycopersicon esculentum [10]. It is present in all organs of tomato plants: in leaves, stems, roots and green fruit where it is found at high concentrations (up to ImM), whereas in ripe fruit it is almost absent [11-14].
Tomatine is principally known for its inhibitory activity on fungal growth [10, 15-18], although it has also been reported to have insecticidal effects [16, 19-22], to inhibit the growth of nematodes [23], to be embryotoxic in Xenopus [24], to disrupt cell membranes in mammalian intestinal cells [25], and to induce the immune response in mammals [26]. The principal mechanism determining the toxic effect of tomatine is attributed to its interaction with sterols of cell membranes, the formation of complexes, and the resulting loss of membrane integrity and leakage of cells contents [2, 4,27-29].
Due to its antifungal properties it has been suggested that this glycoalkaloid may be important in resistance to fungal pathogens. Nevertheless, evidence for the role of tomatine as a barrier against microbial attack is inconclusive, and some confronting observations make

295
it difficult assigning exclusively a direct involvement in plant resistance. However, one aspect that seems clear is the observation that fungal pathogens of tomato are generally more tolerant to tomatine than pathogens of other plant species or saprophytes [15, 17]. This suggests that tomato pathogens may have evolved specific mechanisms to avoid the toxic effect of this chemical. Tolerance to tomatine is based on two main mechanisms: some fungi are resistant to the compound because they have modifications in their membrane composition, while others produce specific tomatine-detoxifying enzymes, known as tomatinases. These enzymes carry out a number of reactions to cleave tomatine removing either all four sugars or a single sugar from the steroidal glycoalkaloid. Most of them remove all four sugars by cleaving the p,l-linked galactose, thus releasing the tetrasaccharide p-lycotetraose and the aglycone tomatidine. Other seems to degrade tomatine to tomatidine but releasing monosaccharides rather than the tetrasaccharide. Finally, others remove a single sugar, the terminal D-xylose, from tomatine yielding pi-tomatine, or the terminal p,l-2 linked glucose, forming p2"tomatine. In all these cases, deglycosylation appears to be sufficient to destroy the ability of tomatine to complex with membrane sterols and therefore to eliminate or reduce its toxic effect. Detoxification of tomatine may be how these tomato pathogens avoid the glycoalkaloid barrier. The mode of action and the importance of saponin-detoxifying enzymes from fungi in general has been reviewed elsewhere [6, 30, 31].
Two tomatinases-encoding genes have been cloned, one from Septoria lycopersici [32, 33] and the other from Fusarium oxysporum [34]. Unexpectedly, both genes do not share significant sequence homology, suggesting that these tomato pathogens have acquired the ability to cleave tomatine independently through a process of convergent evolution. Furthermore This is supported by the fact that both proteins employ different enzymatic mechanisms achieving the same goal: to detoxify tomatine. Convergent evolution may be extended to other tomatinases since an array of different molecular mass has been found in the enzymes characterized so far [33, 35-37].
The existence of tomatinases in fungal-tomato pathogens supports the idea that tomatine may play a role in resistance to fungal attack because these enzymes seem to act specifically on tomatine. Such a role is also supported by the finding that at least in F. oxysporum f. sp. lycopersici, tomatinase is produced during infection both in roots and stems

296
throughout the complete disease cycle [34, 38]. However, the fact that expression of this tomatinase is fully repressed in the presence of glucose [34] indicates that besides a role in detoxification of tomatine, the enzyme might also have a nutritional purpose.
Most tomatinases characterized so far are induced by tomatine, indicating that they may be part of a fungal defensive mechanism against the compound. The identification of other genes whose expression is activated in the presence of the glycoalkaloid may provide new insights into alternative mechanisms of resistance and further knowledge on the metabolism of tomatine. By applying mRNA differential display analysis to mycelium of F. oxysponim grown on minimal medium either in the absence or in the presence of tomatine, other genes selectively induced in response to the glycoalkaloid have been isolated. The significance and role of these novel genes are unknown, but they may represent adaptive metabolic changes of fungi to tomatine. The knowledge of their physiological role may contribute in the future to the development of new strategies to restrict pathogen propagation inside the plant.
STRUCTURE OF TOMATINE
Tomatine is a steroid alkaloid glycoside consisting of an aglycone moiety (tomatidine), and a tetrasaccharide moiety (p-lycotetraose) composed of two molecules of glucose and one each of galactose and xylose; the four monosaccharides form a branched structure that is attached at the C-3 position of the aglycone (reviewed by Roddick [7], see also references therein). The structure of tomatine is shown in Fig (1).
Table 1.- Structural characteristics of partially hydrolyzed a-tomatine.
Compound
a-tomatine
Pi-tomatine
p2-tomatine
7-tomatine
5-tomatine
Structure
tomatidine + galactose + glucose + glucose + xylose
tomatidine + galactose + glucose + glucose
tomatidine + galactose + glucose + xylose
tomatidine + galactose + glucose
tomatidine + galactose

297
Partial hydrolysis of tomatine yields structures with modified sugar moieties [39]. The form whose sugar moiety is the tetrasaccharide is called a-tomatine (tomatine) and those forms lacking xylose, one glucose, xylose and one glucose, or having only galactose are called Pi-, P2"5 Y" ^^^ 6-tomatine respectively. Table 1 summarizes the glycosidic composition of the different structures originated by partial hydrolysis of tomatine.
p - LYCOTETRAOSE
CH,
TOMATIDINE
Fig. (1). Structure of the steroidal glycoalkaloid a-tomatine showing the aglycone moiety (tomatidine) and the tetrasaccharide moiety (p-lycotetraose).
Although some forms are detected when tomatine is extracted from the plant, these glycosides are probably either products of enzymatic hydrolysis during extraction or natural intermediates in the biosynthesis and/or degradation of tomatine.
DISTRIBUTION OF TOMATINE IN TOMATO PLANTS
Analysis of tomatine contents in tomato plants showed that the glycoalkaloid is present in all organs [7, 40-42]. There is a significant variation in the reported tomatine content of particular plant organs. For example, Tukalo (1958) found, in terms of dry weight, 0.86-1.9% tomatine in leaves, 0.3-0.6% in stems and roots and 0.93-2.2% in fully

298
expanded flowers of tomato plants [43]. Much of this variation appears to be due to factors such as plant variety, the stage of plant development, time of season and culture conditions [44-47].
Recently, new methods have been developed for very precise quantification of tomatine in tomato plants [13, 48-50]. Thus, Friedman and Levin, used HPLC with a pulse amperometric detection method, for the direct analysis of tomatine in different parts of tomato plants. They found 0.134 % in roots, 0.287 % in stems, 0.567% in calyxes, 0.64% in leaves and 0.745% in flowers [13].
Tomatine is not transported into fruits from vegetative organs, but is synthesized by cells in the green fruit. The relative amount of tomatine varies from young green fruits (0.46 %) to red mature ones (0.002%), showing a decrease of 99.5% in the level of the glycoalkaloid. Therefore ripe fruit contained the lowest levels of tomatine relative to any other part of the plant [13]. This observation, that tomatine content decreases dramatically during ripening of the fruit, agrees with the reports of other authors. Thus, Sander showed that young developing fruits accumulated large amounts of tomatine, but when ripening begins alkaloid degradation occurred and concentrations declined [40]. Kajderowicz-Jarosinska recorded high levels of tomatine in green, yellowish and red (ripe) tomato fruits, but when ripe fruits were left on the plant for further 2-3 days, tomatine almost disappeared completely [51]. Finally, Bushway et al. found that only five of one hundred red tomatoes contained levels of tomatine detectable by HPLC [48].
Tomatine has not been detected in dormant seeds but it appears in the radicle during the early stages of germination. In the root, de novo synthesis of tomatine has been established by studies with cultured excised roots of L pimpinellifolium and L esculentum although, in the latter species, alkaloid levels per unit of dry weight were lower [52]. There are indications that the main sites of tomatine biosynthesis in the root are the actively growing regions [7, 52]. Generally it is recognized that the main organ of tomatine synthesis and accumulation is the shoot, and that synthesis takes place principally in meristematic region [40]. On the contrary to other steroids that are synthesized in the cell membrane, tomatine seems to be formed in the soluble phase of the cytoplasm of tomato cells [53].
In spite of the considerable variation in tomatine levels observed between different tomato cultivars [44-47], there is no formal

299
demonstration that correlates saponin content to disease resistance. In fact, tomatine does not appear to be determinant in variety-specific resistance of near-isogenic tomato lines to the vascular wilt pathogen Verticillium albo-atrum, since no particular accumulation of tomatine in the resistant interaction w as found as compared to the compatible one, both before and after infection with the pathogen [54]. Similar conclusions are obtained from studies with F . oxysporum f. sp. lycopersici. In this case, an increase in tomatine concentration in the xylem fluids was found in one cultivar after wounding and inoculating with the pathogen, but again no differences were observed between resistant and compatible interactions [55]. The interpretation of these experiments is complicated by the fact that both V. albo-atrum and F. oxysporum f. sp. lycopersici are able to degrade tomatine enzymatically. A more effective strategy to evaluate the importance of the glycoalkaloid in plant defense, might be the isolation of tomato-plant mutants defective in tomatine biosynthesis [56].
BIOLOGICAL ACTIVITY OF TOMATINE
Mode of action of tomatine Tomatine is known to alter membrane integrity and cause lysis in fungal hyphae [57], animal cells [42] plant organelles [58] and plant tissues [28, 58]. The mode of toxic action of tomatine was earlier attributed to the detergent properties of the glycoalkaloid, but Shultz and Sander demonstrated that tomatine is able to complex in a highly specific manner with 3p-hydroxy-sterols in vitro [59]. Thus, it became evident that binding of the glycoalkaloid to membrane sterols could be the cause of destabilization of the membrane lipid bilayer and this explained the lytic and toxic activity observed. Nevertheless, the possibility that tomatine may have additional effects on fungi other than its membraneolytic action was not excluded.
This primary mode of action of tomatine, that involves the formation of complexes with membrane sterols is similar to that described for polyene antibiotics [2, 4], and results in pore formation and loss of membrane integrity. This mode of action is supported by the reduced activity of tomatine on sterol-free bacteria and Oomycete fungi such as Pythium and Phytophthora [15, 28], and the strongly reduced toxicity of hydrolysis products of the glycoalkaloid which fail to bind sterols [57].

300
In vitro studies by Roddick and Drysdale on the effect of tomatine on the leakage of peroxidases from Hposomes also support the idea that tomatine binds membrane sterols. They detected a significant disruption of the liposome by tomatine, only if the membranes contained sterol and additionally this destabilization was pH-dependent [27]. Later it was demonstrated that liposome membranes containing sterols but lacking 3(3-hydroxy sterol were resistant to tomatine [28].
Since the mechanism of action of tomatine is relatively nonspecific, it is expected to affect all eukaryotic organisms which have free sterols in their membranes. Although it is not clear how plants protect themselves from the lytic effects of their own saponins, it seems that the content of p-hydroxy sterols in the membrane is likely to be a major mechanism [28, 29]. It has been demonstrated that the electrolyte loss from cells incubated in the presence of tomatine was less severe in tissue of tomato (10% free sterols) and potato (12% free sterols) than that of tobacco (50% free sterols) and Nicandra physaloides (54% free sterols) [28]. These observations suggest that tomato tissue is resistant to high concentrations of tomatine because the tomato cell membrane contains a low proportion of free sterol [28]. The same conclusion is drawn for potato tissues, which contain high quantities of a-solanine and a-chaconine [9], glycoalkaloids structurally related to tomatine and analogous in their mode of action [29]. In addition, plants may protect themselves from their own saponins by compartmentalizing them in the vacuole or in other organelles. In the case of tomatine it is stored in the vacuole of tomato cell as biologically active molecule [53]. The membranes of these organelles may avoid lysis due to a low sterol content or to a high proportion of sterols substituted at 3-p-hydroxy 1 position [28].
Carbohydrate side-chain and tomatine activity
The precise way in which saponins becomes incorporated into membranes is still unclear and various models have been proposed [2]. The nature of the aglycone and the oligosaccharide moiety of the tomatine molecule are both likely to contribute to its membraneolytic properties [4]. Initial research led to the hypothesis that the inactivation process of tomatine involves removal of one or all the saccharides of the p-lycotetraose, and that tomatidine or other subproducts formed were

301
nontoxic due to their inability to form a stable complex with membrane-bound sterol [57].
An alternative mechanism of action of tomatine was proposed in which the aglycone, rather than tomatine itself, would act as the membraneolytic agent [60]. According to this hypothesis, the hardly water-soluble aglycone would be the active part of the tomatine molecule and the sugars were regarded as a solubilizing group aiding delivery of the tomatine to the membrane, where membrane-bound glycosidases are then presumed to activate the saponin by converting it to the aglycone [61]. This view, however, has received little support because it is incompatible with evidence showing that tomatidine, the free aglycone of the tomatine molecule, does not bind cholesterol [57] and that tomatine affects the permeability of artificial membranes which lack hydrolytic enzymes [27, 28].
More recently, Blankemeyer et al studied the effect of tomatine and tomatidine on frog embryos and frog skin. They found that tomatine increased membrane permeability in frog embryos and decreased sodium-active transport in frog skin, in contrast to the essentially negative results with tomatidine [62]. This reinforces the hypothesis that the carbohydrate side-chain is essential for the glycoalkaloid activity.
Other studies show that partially hydrolyzed tomatine may be toxic to fungi. Thus, Sandrock et al obtained Aspergillus nidulans transformants, carrying the tomatinase gene from S, lycopersici. A transformed strain was as sensitive to tomatine as the fungus before transformation. Although such behavior could be due to poor expression of the tomatinase gene in Aspergillus it is more likely that p2-tomatine, one of the products originated from tomatine by tomatinase, may be toxic to A, nidulans. In fact, the authors showed that^. nidulans was quite sensitive to p2-tomatine and that the transformed strain was even more so [33]. In addition, it was observed that while p2-tomatine and tomatidine were less toxic to most tomato pathogens than tomatine, these partially hydrolyzed products were inhibitory to some saprophytes and nonpathogens of tomato [17]. The mechanism by which these breakdown compounds exert their toxic action is unknown.

302
MECHANISMS OF RESISTANCE OF FUNGI TO TOMATINE
Correlation between the capacity of pathogens to infect tomatine-containing plants and their resistance to the glycoalkaloid in vitro, suggest that for many fungi, tomatine resistance is a prerequisite for successful infection [6, 17]. Fungi that invade tomato and other Solanaceous plants use different strategies for protecting themselves from the host tomatine.
Avoiding release of tomatine
An obvious way to escape the effect of tomatine is to avoid exposure to the compound. Tomatine is usually stored in the vacuole of tomato cells [53]. Stemphylium solani, is a leaf tomato pathogen very sensitive to tomatine [17], but it grows within the intercellular space and middle lamella of leaf [63], here the advancing hyphae may not come in contact with tomatine. A similar situation is true for the hemibiotrophic tomato pathogen Cladosporiumfulvum, also highly sensitive to tomatine but that avoids its release from the cells by restricting growth to the intercellular spaces of tomato leaves without causing cell damage [64]. In this case, it has been suggested that tomatine may play a role in the variety-specific resistance of tomato to incompatible races of C fulvum, through a release of the saponin from leaf cells in the incompatible interaction, thus killing the pathogen or stopping it from invading the plant [64]. The modes of pathogenicity of 5. solani and Cfulvum differ from other tomato pathogens, such as Septoria lycopersici [65] and Alternaria solani [66], which directly penetrate leaves.
Other tomato pathogen very sensitive to tomatine are Verticillium dahliae and F. albo-atriim. These fungi are wilt pathogens and may avoid tomatine limiting their growth to the xylem tissue [17]. There is evidence, however, that avoidance of plant saponins is not a very frequent strategy in fungal pathogens.
Changing ambient pH to avoid optimal action of tomatine
The membraneolytic action of tomatine is strongly impaired at pH lower than five [27] and evidence exists that some fungi may be able to grow on tomatine-containing tomato tissue by lowering the pH at the infection site

303
[42]. This is the case of A. solani, that may cope with the effects of tomatine by lowering the extracellular pH to levels at which the saponin is ineffective as an antifungal agent [60].
Altering membrane composition
A number of lines of evidence indicate that membrane composition appears to be one of the main mechanisms of resistance of fungi to tomatine.
First, fungal species which not produce 3p-hydroxy sterols are more tolerant to tomatine. Hence, Oomycetes such as Pythium and Phytophthora that lack this kind of membrane sterols are highly resistant to the glycoalkaloid. However, they can incorporate exogenous sterols into their membranes when these are added to the growth medium, acquiring then increased sensitivity to tomatine [28, 67]. Thus, Steel and Drysdale studied electrolyte leakage in Phytophthora megasperma in the presence of tomatine, measuring electrolyte loss due to membrane damage in mycelium grown in the presence of increasing concentrations of cholesterol. The sterol-containing mycelium was definitely more susceptible to tomatine than sterol-free mycelium [28]. Accordingly, fungi that normally contain sterols in their membranes become more resistant to saponins when they are grown in the presence of inhibitors of sterol biosynthesis [68]. A detailed sterol-binding studies with tomatine confirm that binding requires membrane sterols with free 3p-hydroxyl groups [28]. Therefore, susceptibility of membranes to tomatine appears to be mainly influenced by sterol composition.
Second, the importance of membrane composition in determining the ability of fungi to infect tomatine-containing plant, is further emphasized by experiments involving a tomato-attacking isolate of Fusarium solani. The wild-type isolate was pathogenic only to ripe tomato fruits which contain very low levels of tomatine. However, sterol-deficient mutants of a tomato-attacking isolate of F. solani showed increased resistance to the steroidal glycoalkaloid and gained the ability to infect green fruits of tomato, which are particularly rich in tomatine [69, 70]. Analysis of the progeny from crosses between mutant and wild-type fungi showed that pathogenicity to green tomato fruits, low sterol content, and insensitivity to tomatine were always inherited together, revealing the importance of membrane composition in pathogenicity [70].

304
Third, it has been demonstrated, studying tomatine sensitivity of ergosterol mutants of Neurospora crassa, that changes in the type of membrane sterol composition can increase resistance to tomatine [17]. Ergosterol, a 3p-hydroxy sterol, constitutes 80% of the total sterol component of N, crassa [71]. Several mutants affecting sterol biosynthesis and accumulating different sterol intermediates have been isolated [71, 72]. All the mutants became less sensitive to tomatine, and although the interaction of the sterol intermediates of each mutant with the glycoalkaloid is unknown, clearly the diverse types of sterols might interact differently with it, because the different mutants show specific grades of resistance to tomatine [17]. Recently, the erg-S gene from S. lycopersici coding the ergosterol biosynthetic enzyme sterol C-14 reductase has been cloned and has been confirmed that the gene complements the phenotype tomatine-resistant of a N. crassa erg-S mutant [73]. Other research has revealed that the increase of the concentration of glycolipids in cholesterol-containing membrane results in an increase of tomatine sensitivity [29]. Therefore it seems that different alterations of the membrane, not only sterol content, may protect tomato pathogens from the toxic effect of tomatine and other glycoalkaloids.
Tomatine-detoxifying enzymes A second major mechanism of resistance to tomatine involves enzymatic detoxification by tomatinases. Although saponins are very numerous and widely distributed in the plant kingdom, detailed studies on saponin detoxification by fungi have been restricted to pathogens of a few plant species, principally to oat and the Solanaceous tomato and potato. This is because structures and antifungal properties of oat, tomato and potato saponins are well established, and that saponin profiles of these plants are relatively simple, in contrast to other plants like alfalfa, where over 20 predominant different saponins have been identified.
The importance of saponin-detoxifying enzymes in determining the host range of fungal pathogens has been demonstrated for oat- and wheat-attacking isolates of the soil borne fungus Gaeumannomyces graminis. Isolates of G. graminis var. avenae are resistant to avenacin A-1, a saponin predominant in oats. This fungus produces a saponin-detoxifying enzyme, called avenacinase, consisting in a p-glycosyl hydrolase that removes (J, 1-2- and p,l-4-linked terminal D-glucose molecules from avenacin A-1 [74-76]. Isolation of the gene encoding avenacinase allowed
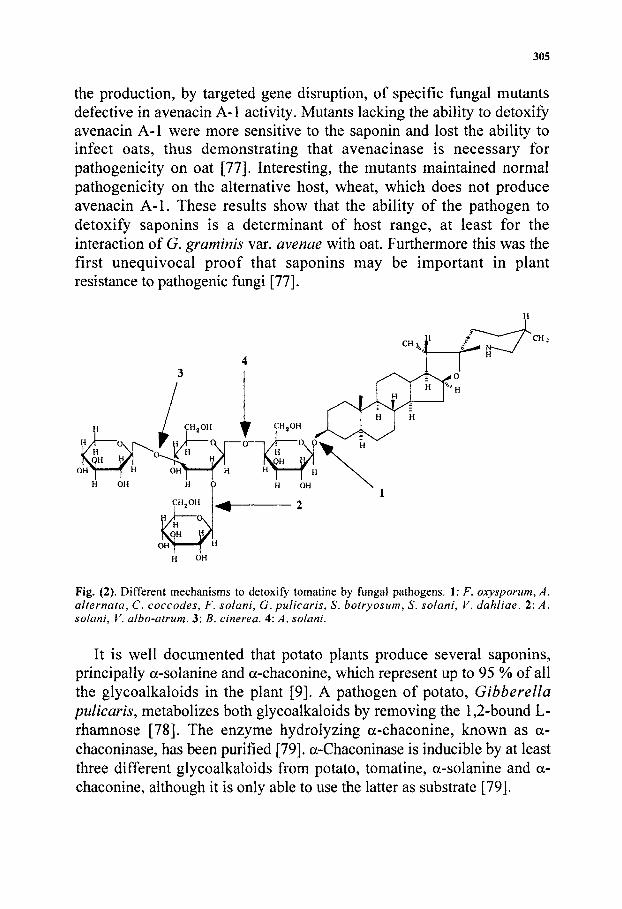
305
the production, by targeted gene disruption, of specific fungal mutants defective in avenacin A-1 activity. Mutants lacking the ability to detoxify avenacin A-1 were more sensitive to the saponin and lost the ability to infect oats, thus demonstrating that avenacinase is necessary for pathogenicity on oat [77]. Interesting, the mutants maintained normal pathogenicity on the alternative host, wheat, which does not produce avenacin A-1. These results show that the ability of the pathogen to detoxify saponins is a determinant of host range, at least for the interaction of G. graminis var. avenae with oat. Furthermore this was the first unequivocal proof that saponins may be important in plant resistance to pathogenic fungi [77].
H / CH^OH y CH^OH
QH> ^ H 0H> r H H > - f H
H OH H O H OH H
CH,OH
4l
OH H OH
Fig. (2). Different mechanisms to detoxify tomatine by fungal pathogens. 1: F. oxysponim, A. alternata, C. coccodes, F. solani, G. pulicaris, S. botryosum, S, solani, V. dahliae. 2: A. solani, V. albo-atrum. 3: B. cinerea. 4: A. solani.
It is well documented that potato plants produce several saponins, principally a-solanine and a-chaconine, which represent up to 95 % of all the glycoalkaloids in the plant [9]. A pathogen of potato. Gibberella piilicaris, metabolizes both glycoalkaloids by removing the 1,2-bound L-rhamnose [78]. The enzyme hydrolyzing a-chaconine, known as a-chaconinase, has been purified [79]. a-Chaconinase is inducible by at least three different glycoalkaloids from potato, tomatine, a-solanine and a-chaconine, although it is only able to use the latter as substrate [79].

306
In contrast to other saponins, in tlie case of tomatine an important number of fungal pathogens of tomato is known to produce specific enzymes known as tomatinases, able to hydrolyze the compound (Table 2). These glycosyl hydrolases remove one or more sugars from the tetrasaccharide moiety of tomatine thus impairing its binding to 3p-hydroxy sterols and avoiding its toxic effect [29, 57]. The metabolites originated by the hydrolysis of tomatine have little or no antifungal activity to most tomato pathogens [17]. Thus, for many phytopathogenic fungi the production of tomatine-detoxifying enzyme may be a determinant to successfully infect tomatine-containing hosts.
The mechanisms of action of tomatinases differ, Fig. (2). Whereas some remove a single sugar, most of them release the intact p-lycotetraose group, yielding tomatidine (Table 2).
Tomatinase from Septoria lycopersici
S. lycopersici is a foliar pathogen of tomato. Arneson and Durbin demonstrated that crude mycelial extract or culture filtrates from this fungus converted a-tomatine to p2-tomatine. Fig (2), by hydrolysis of the p-l,2-D glucosyl bond on the tetrasaccharide moiety, removing a single D-glucose from p-lycotetraose [80]. The enzyme was partially purified and classified as p-l,2-D-glucosidase [81]. Purification to homogeneity was carried out by Sandrock et aL who named the enzyme P2-tomatinase [33] to distinguish it from other fungal tomatinases that release the entire tetrasaccharide moiety or other sugar molecules from tomatine [17, 82, 83]. Although mainly present in extracellular extracts, the enzyme was purified from intracellular extracts to overcome the problem of extracellular melanin-like pigments that hampered the purification.
Avenacinase, an enzyme from G. graminis var. avenae, is related to tomatinase from 5. lycopersici because is able to deglucosilate tomatine by identical mode of action. However, the activity is very low and corresponds to approximately 2% of its activity towards avenacin A-1 [32]. Tomatinase form S, lycopersici, also can cleave avenacin A-1 but has less than 0.01% of activity towards it in comparison to its activity towards tomatine [32]. Therefore, the two enzymes are highly specific for their respective host plant saponins. Purification and characterization of 5. lycopersici tomatinase revealed that this enzyme shares many properties (including immunological cross-reactivity) with avenacinase

Table 2.- Fungal pathogens with tomatinase activity.
307
Species
Altemaria alternata
Alternaha alternata f. sp. lycopersici
Altemaria solani
Botrytis cinerea
Colletotrichum coccodes
Fusarium solani
Fusarium oxysporum f. sp. gladioli^
Fusarium oxysporum f. sp. lycopersici
Fusarium oxysporum f. sp. melonis^
Fusarium oxysporum f. sp. niveum^
Fusarium oxysporum f. sp. radiciS'lycopersici
Fusarium oxysporum f. sp. tuberosi^
Gibberella pulicaris^
Septoria lycopersici
Stemphylium botryosum
Stemphylium solani
Verticillium dahliae
Verticillium albo-atrum
Final enzymatic degradation product
tomatidine
tomatidine
tomatidine, p2-tomatine, 8-tomatine
Pl-tomatine, tomatidine
tomatidine
tomatidine
tomatidine
tomatidine
tomatidine
tomatidine
tomatidine
tomatidine
tomatidine
P2-tomatine
tomatidine
tomatidine
tomatidine
P2-tomatine
Reference
p-^j
[17]
[17]
[17, 36]
[17]
[37]
[89]
[17, 35, 83]
[89]
[89]
[89]
[89]
[91]
[33, 80, 81]
[17]
[17]
[17]
[17, 54]
^ Not pathohenic on tomato.

308
from G. graminis var. avenae [32, 33]. In fact, a cDNA clone encoding tomatinase from S. lycopersici was isolated using avenacinase cDNA as a heterologous probe [32]. The sequence of 5. lycopersici tomatinase revealed that this enzyme shares 68% amino acid similarity with avenacinase.
Glycosyl hydrolases have been grouped into approximately 40 families on the basis of sequence similarity [84, 85]. Within this classification the majority of p-glucosyl hydrolases (EC 3.2.1.21) fall into two families: 1 and 3. The deduced amino acid sequence of the gene encoding tomatinase from 5. lycopersici reveals a high degree of similarity to several other microbial p-glucosidases belonging to family 3 of glucosyl hydrolases [85]. Avenacinase from G. graminis var. avenae belongs to the same family. The members of family 3 most closely related to avenacinase and tomatinase are p-glucosyl hydrolases from fungi that have importance in cellobiose degradation (BGLl from Trichoderma reesei, BGLl and BGL2 from Saccharomyces fibuligera, and BGLS from Candida pelliculosa). The degree of amino acid identity between tomatinase and these other p-glucosidases is 40 to 45%. Avenacinase and tomatinase are more closely related to each other than to other members of family 3, and therefore may represent a subgroup within this family that has the additional ability to degrade saponins [30]. Despite the homology between microbial p-glucosidases of family 3 and tomatinase, it is unknown whether other enzymes from this family, except avenacinase, posses activity towards tomatine or other saponins. The substrate specificity of tomatinase was evaluated by incubating the enzyme with other a- or p-D glycosides. Tomatinase showed a strong preference towards tomatine, having a small activity towards F-gitonin and digitonin, almost undetectable on para-nitrophenyl-p-D-glucopyranoside and no activity at all towards nine other glycosides and/or steroid or triterpenoid compounds similar to tomatine [32, 33]. Many organisms with p-glucosidases found within family 3, such as soil-inhabiting saprophytes, may come in contact with saponins in nature and thus possess these enzymes for detoxification or degradation as a food source. More extensive studies on substrate specificity of these enzymes are required in order to establish whether this is the case.
Tomatinase from S. lycopersici has been introduced in the hemibiotrophic tomato pathogen C.fulvum, that is very sensitive to tomatine. Tomatinase-producing transformants reveal increased

309
sporulation on cotyledons of susceptible tomato cultivars, and they also cause more extensive infection of seedling of resistant tomato lines [86]. These observations suggest that tomatine may contribute to restrict the growth of C.fulvum to some extent in both compatible and incompatible interactions, but indicate that tomatinase activity is not sufficient for pathogenicity.
Tomatinase from Fusarium oxysporum
The tomatinase from the vascular wilt pathogen F. oxysporum f. sp. lycopersici has a different mechanism of action than the S. lycopersici tomatinase, releasing the intact p-lycotetraose group to give the aglycone tomatidine, Fig. (2) [35, 83]. The enzyme is extracellular and inducible by tomatine whereas breakdown products (tomatidine and sugars) fail to increase the level of tomatinase activity [35].
When grown in the presence of tomatine, F. oxysporum f. sp. lycopersici secrets a unique protein species with tomatinase activity. The enzyme is a monomer of 50 kDa. In native conditions consists of at least five isoforms with pis ranging from 4.8 to 5.8, though the same molecular mass. Treatment of pure tomatinase with A^-glycosidase F gives a single band of 45 kDa indicating that the different isoforms are glycosylated [35]. The presence of glycosylation in extracellular enzymes is frequent, it aids to protein folding and to the stabilization of protein conformation, and additionally glycosilation facilitates protein transport and confers resistance to proteolytic degradation [87]. The optimum pH range of tomatinase activity is between 5.5 and 7, in agreement with the fact that tomatine reacts optimally with membrane sterols at pH 7 [27]. Thus, in the plant cellular environment with a pH around 7, the enzyme has maximal activity avoiding presumably membrane disruption.
Tomatinase activity in infected tomato plants has been determined at different stages of infection [38]. No tomatinase activity is detected in leaves at any stage of infection. In stems, activity is higher during the final wilt stage compared to previous stages of infection. In roots, enzyme activity appeared with the first symptoms and is maintained until wilting. Tomatinase activity is not detected in protein extracts from infected plants before appearance of symptoms (1 to 5 days after infection). This may be due to the low quantity of pathogen in the plant and to the inhibitory effect of tomatine on spore germination and mycelial growth. It

310
may also explain the very low tomatinase activity observed in stems of infected tomato plants at stages previous to wilting, where the fungus reaches and proliferates in the stem. Tomatinase activity is also absent in protein extracts from stems and roots of healthy plants, thus ruling out the presence of enzymes with tomatinase activity in non-infected plants. In all cases, the tomatinase activity detected converted the tomatine in tomatidine and p-lycotetraose [38]. Immunoblotting assays using rabbit polyclonal antibodies raised against tomatinase detected a 50 kDa band corresponding to the tomatinase enzyme in protein extracts from roots and stems of plants during the wilt stage of infection [35, 38]. These results demonstrate that the same tomatinase enzyme is induced both in vitro and in vivo. This conclusion is supported by experiments of reverse transcription-polymerase chain reaction (RT-PCR) in planta. Tomatinase gene was indeed expressed throughout the entire disease cycle of F. oxysporum f sp lycopersici both in roots and stems [34]. On the basis of these results it seems clear that expression of tomatinase gene is specifically controlled by the interaction with the host plant.
Although several authors claimed that the glycoalkaloid tomatine is not present in sufficient concentrations in roots or stems to play a major role in resistance to Fusarium oxysporum f. sp. lycopersici [7, 57, 88], the fact that tomatinase is induced in tomato plants during infection, strongly suggests that the enzyme may have a specific role in pathogenicity of tomato plant by F. oxysporum f sp lycopersici.
Tomatinase activity has been traditionally associated to fungi pathogenic on tomato [17, 30]. Recently, Lairini et al. [89], studying tomatinase induction in various ybrma^ speciales of F. oxysporum, found inducible tomatinase activity not only in the tomato pathogens F. oxysporum f sp. lycopersici and radicis-lycopersici, but also in formae speciales not pathogenic to tomato, such as tuberosi, melonis, niveum and gladioli (Table 2). These formae speciales are natural pathogens of other plant species. Some of these are phylogenetically related to tomato, such as the Solanaceous potato, pathogenized by f. sp. tuberosi, whereas others, are not closely related, such as the Cucurbitaceae muskmelon and watermelon pathogenized by f. sp. melonis and f. sp. niveum, respectively, and the Iridaceous monocot Gladiolus sp pathogenized by f sp. gladioli. The tomatinases found in these formae speciales have very similar characteristics to tomatinase from f sp. lycopersici, including the same mode of action hydrolyzing tomatine into tomatidine and p-

311
lycotetraose, the same molecular mass (50 kDa), and they are recognised by polyclonal antibodies raised against f. sp. lycopersici tomatinase [38]. These results suggest that the tomatinases from different formae speciales of F. oxysporum are probably the same enzyme.
The function of these tomatinase in formae speciales that do not pathogenise tomato is unknown. One possible explanation could be the presence of tomatine or similar saponins in their host plant species. However, (i) tomatine has not yet been reported in these plants [2, 7, 9] and (ii) although some of these species contain small amount of tomatine and other saponins structurally related to tomatine (e.g. potato contains a- solanine and a-chaconine [2,4, 9, 90]), these are inactive as inducers of tomatinase and, moreover, tomatinase cannot use any of these glycoalkaloids as substrate [89]. In addition, it is clear that tomatinase is not required for pathogenicity in these isolates, at least in the case of F. oxysporum f. sp. melonis, where some strains that are fully pathogens on muskmelon lack tomatinase activity [89].
If they do not have a function in pathogenicity, the presence of tomatinases in different formae speciales of F. oxysporum and in other fungi not pathogenic to tomato such as G.pulicaris [91], points out a close phylogenetic relationship between these formae speciales and may indicate common forms of evolution for fungal pathogens. Two possible models of evolution of formae speciales in F oxysporum and fungal tomato pathogens can be envisaged in general. Fig. (3). In one model, fungal saprophytes that have evolved into fungal pathogens of tomato may have acquired the ability to degrade tomatine as prerequisite for pathogenicity. Fig. (3A). Once the fungus has gained this ability, it may have evolved further into forms pathogenic to other host plants, maintaining (or not) the ability to degrade tomatine. In the second model. Fig. (3B), fungal pathogens with a broad host range could have started as "general pathogen" such as today are V. dahliae and V. albo-atrum; both of these pathogens cause vascular wilt on a wide variety of host plants range throughout the world, without the occurrence of formae speciales [92]. Thus, a hypothetical pathogen with a wide host range should have acquired several pathogenicity traits such as the ability to detoxify tomatine. On the other hand, hosts may have developed new resistance genes to overcome plant resistance mechanisms, and strains of the pathogen have increasingly evolved to forms with a remarkable degree of host specialization. During this process, the strains may have maintained

312
(or not) some enzymatic characteristics from the ancestral pathogen, such as the ability to detoxify tomatine. These two models are not incompatible and could both have originated the different formae speciales of F, oxysporum and other fungal pathogens that maintain tomatinase activity and are not able to infect tomato.
FUNGAL SAPROPHYTE
tomatinase acquisition
FUNGAL PATHOGEN WITH BROAD HOST RANGE, HAS TOMATINASE AND OTHER
PHYTOANTICIPIN-DEGRADING ENZYMES
FUNGAL TOMATO PATHOGEN
maintains or not tomatinase activity
plant recognises specific avirulence gene product of the pathogen and develops resistance
FUNGAL PATHOGEN OF OTHER HOST PLANT
EVOLUTION TO PATHOGEN FORMS WITH HIGH DEGREE OF
HOST SPECIALIZATION (FORMAE SPECIALES)
Fig. (3). Possible mechanisms of evolution of fungal tomato pathogens and formae speciales. A: Fungal tomato pathogen or formae speciales originated from a non-pathogen. B: Specific tomato pathogen or formae speciales originated from broad host range pathogen.
The tomatinase gene from F. oxysporum f. sp. lycopersici has been cloned recently [34]. This gene encodes a protein that has no sequence homology to any previously described saponinase but which is highly similar to xylanases (family 10 of glycosyl hydrolases) [84, 85, 93-95]. Although F. oxysporum tomatinase does not have any detectable xylanase activity, it remains to be determined whether any of the xylanases listed in this family possesses activity against a-tomatine. In any case, it is

313
conceivable that even highly related enzymes, both structurally and functionally, may have very different substrate specificity.
As mentioned before, the tomatinase gene from the tomato pathogen S. lycopersici [32] encodes a protein closely related to avenacinase saponin-detoxifying enzyme, produced by G, graminis, and that both enzymes are closely related to family 3 of glycosyl hydrolases, which mainly includes p-glucosidases [32]. The fact that F, oxysporum tomatinase shares no sequence homology with tomatinase from S. lycopersici suggests that these different tomato pathogens have developed the ability to cleave tomatine by convergent evolution. This may also explain the different molecular mass of both native tomatinases, 110 kDa for S. lycopersici [33] and 50 kDa for F. oxysporum [35].
The tomatinase from F. oxysporum is encoded by a single gene whose expression is induced by tomatine and fully repressed when mycelium is grown in the presence of glucose. This observation is in contrast to that reported for tomatinase from B. cinerea which is not subject to carbon catabolite repression [36]. Analysis of the tomatinase gene revealed the presence of promoter sequence motifs involved in carbon catabolite repression in yeast and filamentous fungi. Two putative CREA-binding sites [96], suggest that glucose repression may be modulated by CREA. Therefore, it can be concluded that tomatinase production is subject to substrate induction and to catabolite repression. In addition, probably signals derived from the interaction with the plant must regulate expression of this activity since the amount of tomatine found in roots (14 mg/lOOg fresh weight) and in stems (54 mg/lOOg fresh weight) is apparently sufficient to inhibit the growth of F. oxyxporum to some extent [17] and to induce expression of the tomatinase gene [34, 35].
On the other hand, since there is a high sequence homology between tomatinase and glycosyl hydrolases, enzymes presumably required for nutritional purposes, and tomatinase expression is repressed by glucose, it is possible to suggest that, together with its role in detoxification of tomatine, tomatinase might also have a nutritional function. I any case to verify the importance of tomatinase from F. oxysporum f. sp. lycopersici in pathogenicity, the cloned gene is being used to obtain tomatinase-minus mutants by targeted gene disruption, to further study their ability to pathogenise tomato plant.

314
Tomatinase from Botrytis cinerea
The necrotrophic pathogen E. cinerea causes grey mould disease on many fruits, vegetables and flowers. Although some reports indicate that tomatinase from B. cinerea deglycosylates tomatine completely [17, 41], with the same mechanism that tomatinase of F. oxysporum, Quidde et al. analyzing twelve isolates from different geographic areas found that this fungus metabolizes tomatine by removal of the terminal p-1,3-linked D-xylose moiety, yielding Pi-tomatine [36]. Therefore, for most isolates of B. cinerea the mechanism of tomatinase in detoxifying tomatine differs from the tomatinases of S. lycopersici and F. oxysporum [36].
One of the isolates used by Quidde et aL, was unable to metabolize tomatine and was more sensitive to the glycoalkaloid. In addition, this isolate was not able to induce disease symptoms on detached tomato leaves, but could pathogenize bean leaves, indicating that the ability to degrade tomatine may be required for pathogenicity of B. cinerea on tomato [36]. It would be very interesting to transformed this strain with one tomatinase gene to confirm this hypothesis.
The structural relationship of tomatinase from B. cinerea to other tomatinases is unknown, but its molecular mass (70 kDa) is different from the other two enzymes mentioned before, 50 kDa from F. oxysporum [35, 38] and 110 kDa from S. lycopersici [33]. Moreover, when Quidde et al, attempted cloning of the tomatinase gene from 5. cinerea using the tomatinase from S. lycopersici as a probe, they isolated a gene with high sequence homology, whose product had not tomatinase activity but was able to detoxify avenacin A-1 [97], the saponin from oats related to some extent to tomatine.
Tomatinase from Fusarium solani
The occurrence of a tomatine-detoxifying enzyme in F. solani, was studied in liquid medium with or without tomatine. No tomatinase activity was detected in the absence of tomatine, whereas a remarkable increase of tomatinase activity was observed after 12 h of growth in the presence of the glycoalkaloid, the maximal tomatinase activity being observed after 72 h of incubation [37]. Detection of tomatinase activity in F. solani contrasts with the results reported by Defago and Kern [70],

315
which found tomatinase activity in F, oxysporum f. sp. lycopersici but were unable to detect such activity in F. solani (strain ETH 4874).
The mode of action of F. solani tomatinase was studied by thin layer chromatography (TLC) analysis. No degradation of tomatine was observed using as source of the enzyme the crude extract from fungi grown without tomatine. Crude extracts from F. solani grown in the presence of tomatine converted the glycoalkaloid tomatine into two products, tomatidine and p-lycotetraose. The same results were obtained for both F. solani isolates studied. Hence, the mode of action of tomatinase from F, solani is similar to that of tomatinase of F. oxysporum [35, 83].
In order to investigate whether tomatinases from F. oxysporum and F. solani share similar molecular characteristics, F. solani tomatinase was partially purified. Comparative SDS-PAGE analysis of the protein fractions with and without tomatinase activity showed the presence of a 32.5 kDa band in all positive fractions, while this band was absent in fractions without tomatinase activity The apparent molecular mass of tomatinase of F solani differs from that of F oxysporum (50 kDa), S. lycopersici (110 kDa) [33], and Botrytis cinerea (70 kDa)[36]. The F. solani tomatinase presents a very low activity compared with F. oxysporum enzyme [35, 89]. Western blot analysis showed that the two enzymes also differ in their immunological characteristics since the polyclonal antibody against tomatinase of F oxysporum f. sp. lycopersici did not recognize the tomatinase from F solani. These results suggest that the enzyme from F solani is a novel tomatinase species.
When inoculated on tomato fruits, Fusarium solani produced severe rot on the surface of the green fruit one week after inoculation [37]. This result is in contrast to F. solani isolate 4874 used previously by Defago and Kern which did not rot green tomato [70]. To study the possible biological relevance of tomatinase, F oxysporum f sp. lycopersici, with a tomatinase activity five-fold higher than that of F. solani isolates, was used as control. F oxysporum f sp. lycopersici, was not able to grow on the green fruit probably due to the high level of tomatine present; nevertheless, when the fruit began to turn red and its tomatine content decreased, F oxysporum f sp. lycopersici was able to grow into the fruit and produce severe rot. F solani isolates also were more aggressive on red than on green fruit, but whereas F. solani was more aggressive on green

316
fruit than F, oxysporum f. sp. lycopersici, this was more aggressive on the red fruit than F. solani [37].
In conclusion, F. solani isolates appear to be less sensitive to tomatine than F, oxysporum f sp. lycopersici, although they produce much lower tomatinase activity, indicating that they must possess other inherent mechanisms of resistance. One possibility is that F. solani may have a lower content of sterols in the membrane than F. oxysporum. Thus, although tomatinase activity may help F. solani to grow on green tomato tissue it does not seem to be determinant in the resistance to tomatine.
Other fungal tomatinases
The tomatinases known remove either a single sugar or all four sugars from the steroidal tomatine, Fig (2). In a recent study, Sandrock and VanEtten examined twenty-three fungal isolates (12 tomato pathogens, 7 pathogens of other plant hosts, and 2 saprophytic fungi) finding that most tomato pathogens have tomatinase activity [17]. Table 2 lists the fungal pathogens so far known to have the ability to hydrolyze tomatine.
Most fungi yield tomatidine by cleaving the p,l-linked galactose and releasing the tetrasaccharide lycotetraose and tomatidine. The tomato pathogen Alternaria solani was reported to degrade also tomatine to tomatidine but releasing monosaccharides rather than a tetrasaccharide [82]. However, Sandrock and VanEtten have shown recently that culture filtrates from A. solani grown without tomatine were able to convert tomatine to tomatidine[17]. Tomatinase activity increased within the mycelium after exposure to tomatine and in this new activity degraded tomatine through sequential hydrolysis to |32-tomatine, 8-tomatine and tomatidine. This result does not suggest that A, solani has only one tomatinase which hydrolyzes more than one glycosidic bond, as other saponin-degrading enzymes do [98]. On the contrary, the different patterns of tomatine degradation observed in the TLC plate, obtained by culture filtrates and mycelium extracts, indicate that there is probably more than one enzyme involved in degrading tomatine. Therefore, further biochemical characterization is necessary to determine the number of enzymes used by A, solani to metabolize tomatine.
A few tomato pathogens remove a single sugar from tomatine. Besides to S, lycopersici and B. cinerea, (sections 5.4.1 and 5.4.3.), that remove

317
either one glucose to produce p2-tomatine (S. lycopersici) or the terminal D-xylose to yield p i-tomatine, Verticillium albo-atrum cleaves the terminal p,l-2-linked glucose from tomatine [54], originating also P2-tomatine.
Based on these data, it is clear that there is not a common enzymatic mechanism by which tomato pathogens metabolize tomatine. It even seems that the regulation of tomatinase expression follows a different pattern in specific pathogens. Thus, although tomatinase enzymes usually are inducible by tomatine, as reported for F. oxysporum [34, 35, 83], V. albo-atnim [54], B. cinerea [36, 41], 5. lycopersici [33] and F. solani [37], some of them are constitutive [17]. In B, cinerea, a detectable level of constitutive activity is found in culture supernatant of the fungus grown without tomatine, although a considerable increase of activity is detected when tomatine is added [36]. In addition, although it is unequivocal that tomatinase form F, oxysporum is completely repressed by glucose [34], this is not the case of B. cinerea where tomatinase induction is independent of the presence, or not of glucose [36]. All these observations, together with the fact that the molecular masses of native tomatinases show significant differences, and that the two genes sequenced so far (S. lycopersici and F. oxysporum), do not present similarity, suggest that tomatinases have been acquired by phytopathogenic fungi independently during evolution. Therefore they may represent important tools to both become pathogens of tomatine-producing plants and metabolize the glycoalkaloid as saprophyte.
METABOLISM OF THE TOMATINE SUBPRODUCTS: THE AGLYCONE TOMATIDINE
In contrast to the extensive knowledge on the metabolism of tomatine by phytopathogenic fungi, there are only few reports dealing with the final metabolism of the subproducts generated by glycoside cleavage of the glycoalkaloid. Initial reports by Sato and Hayakawa [99-101] showed that the fungus Helicostylium piriforme was able to hydroxylate tomatidine and other related steroids.
More recently, Weltring et al studied the metabolism of tomatine by the potato pathogen Gibberella pulicaris (anamorph Fusarium sambucinum) [91]. This fungus is not pathogenic on tomato plants and it is suggested that probably uses tomatine as a source of nutrients.

318
although it is possible that it evolved from an original tomato pathogen, see Fig. (3). Weltring et al followed both the deglycosilation of tomatine and the further modification of the aglycone moiety tomatidine. When tomatine was incubated in the presence of fungal mycelium, it was completely metabolized in two hours giving lycotetraose and tomatidine. The aglycone was then converted into two more polar products that could be detected after four hours. These products were originated by the introduction of one oxygen atom into tomatidine giving mainly 7-a-hydroxy-tomatidine and the corresponding D^-dehydro 7-a-hydroxy-tomatidenole as secondary metabolite. The structures of these subproducts of tomatine are based on mass spectrometry and nuclear magnetic resonance. These polar compounds were further metabolized after eight hours into at least three compounds with increased polarity, as determined by their migration on TLC plates. All these metabolic products were detected after forty-eight hours of incubation, although at this time tomatidine had completely disappeared [91].
Znidarsic et aL showed that tomatidine induces steroidal hydroxylase activity in the filamentous fungus Rhizopus nigricans [102], and similar activity was proposed to be the cause of hydroxylation of tomatidine in G. pulicaris by a membrane-bound cytochrome P-450 monooxygenase [91]. Cytochrome P-450 monooxygenases are responsible of hydroxylation of progesterone at various positions by Aspergillus species [103, 104], R. nigricans [102, 105], Phycomyces blakesleeanus [106], Sepedonium ampullosporum [107], Botryospheria obtusa [108] and Cochliobolus lunatus [109, 110]. Steroid hydroxylation may reflect defense mechanisms consisting in the removal of hydrophobic steroids toxic to fungal mycelium that may have evolved due to the exposure of fungi to plant secondary metabolites. Further characterizations of fungal cytochrome P-450 monooxygenases by gene cloning and disruption are required to prove their physiological and evolutionary roles.
OTHER GENES INDUCED BY TOMATINE
Identification of other genes induced by tomatine may provide new insights into the mechanisms of resistance to this glycoalkaloid. The physiological adjustments that take place in the fungal cell during exposure to tomatine are largely unknown. Differential display analysis [111] of mRNA from mycelium of F. oxysporum f sp. lycopersici grown

319
on minimal medium either in the absence or in the presence of tomatine led to the isolation of three genes whose expression is selectively induced in response to tomatine (Perez-Espinosa, Roldan-Arjona and Ruiz-Rubio, unpublished results). One of these encodes a predicted cytosolic protein of 313 amino acids with high similarity (about 40 %) to leguminous isoflavone reductases (IFRs). The IFR-like gene is expressed by F, oxysporum during infection of tomato plant, as revealed by RT-PCR analysis with gene-specific primers. Since phytopathogenic fungi are exposed to more than one antifungal metabolite when they invade the plant, it is possible that tomatine could act as a signal to induce not only tomatinase but a number of enzymes able to detoxify different antifungal compounds. This is the case of tomatine and a-solanine in the potato pathogen G. pulicaris. Both glycoalkaloids induce the synthesis of a-chaconinase, an enzyme that metabolizes the potato-saponin a-chaconine but is not active towards tomatine or a-solanine [79]. In tomato, a pathogen invading the plant may encounter for instance the phytoalexin rishitin, and not only tomatine [112, 113]. Thus, the induction of the IFR-like gene may play a role in detoxification of antifungal isoflavone compounds during pathogenesis.
The differential display technique allowed the identification of a second tomatine-induced gene from F, oxysporum. The cDNA sequence predicts a cytosolic protein of 376 amino acids and a molecular mass of 41 kDa, with high homology to microbial and plant pantothenate synthetases. A complementation experiment using this cDNA in a panC auxotroph mutant of Escherichia coli confirmed the pantothenate synthetase gene function. The gene is also expressed constitutively, but its expression increases in the presence of tomatine. Pantothenate is a component of acetyl-coenzyme A. In living organisms, the citric acid cycle consists of a series of reactions that oxidize the acetyl group to two molecules of CO2 in a manner that conserves the liberated free energy for utilization in ATP generation. Acetyl groups enter the citric acid cycle as acetyl-coenzyme A, which is the common product of carbohydrate, fatty acid, and amino acid breakdown. Acetyl-coenzyme A has therefore a critical role in cellular bioenergetics. Besides being a precursor of the phosphopantheteine moiety in Coenzyme A, pantothenate is also part of the acyl carrier protein involved in the biosynthesis of fatty acids. According to the role of pantothenate in ATP generation and fatty acid biosynthesis, it could be important for the fungus to induce pantothenate

320
synthetase during plant infection for two reasons. First, tomatine stimulates sporulation in vitro [55] which implies a high energy demand. Second, the toxic effects of tomatine, as mentioned before, are attributed to its ability to complex with membrane sterols [27-29, 64, 114, 115]. Thus it is reasonable to envisage a scenario where the esterol components of the membrane are being reduced and biosynthesis of fatty acids is increased in order to repair the damages in the cell membrane.
A third tomatine-induced gene from F. oxysporum has been identified that encodes a polypeptide with some similarity to cereal storage proteins. The endosperm of cereal grains serves as a seed storage organ where carbohydrates and proteins are laid down in order to feed the embryo during germination. The putative storage protein from F. oxysporum, as other storage proteins from plant seeds, is rich in glutamine and proline, containing in some regions more than 50% of these amino acids. Genes encoding storage proteins have been isolated from several cereal species [116], but there are no reports on any fungal genes and no formal descriptions of storage proteins in fungal spores exist in the literature. VanEtten et al, described a possible storage protein in dormant spores of the pathogen Botryodiplodia theobromae, which produces fruit rot. [117]. Approximately 23% of the total protein isolated from the spores consisted of a single polypeptide that was degraded during germination [118] .A region of the protein encoded by the F. oxysporum gene has homology to glutenin and y-gliadin from wheat, and to y-hordein from barley. Convergent evolution between seed storage proteins from different plants has been reported [119], as indicated by same structure of reserve proteins coming from different origins. Since spore production is stimulated by tomatine in vitro [55], it is reasonable to consider that the gene induced by tomatine should encode a protein that is stored in the spore and used during germination.
FUTURE PERSPECTIVES
There is increasing evidence that tomatine and other saponins that inhibit fungal growth in vitro act as phytoprotectants in plants. Recent data show that mutants of oat deficient in the saponin avenacin A-1 are compromised in their resistance to a variety of fungal pathogens, suggesting that sensitivity is a direct consequence of saponin deficiency [56]. Fungal mechanisms to resist toxicity of these compounds thus

321
appear to be important for pathogenicity. Among the mechanisms of saponin resistance, enzymatic degradation has received most attention, most likely because characterization of genes encoding degradative enzymes offers a more immediate route to genetic tests of function through targeted gene disruption [31]. In the case of tomatine, most of the data indicate that the capacity to actively degrade tomatine is indeed a prerequisite for pathogenicity on tomato, at least in most cases [17]. Strikingly, several lines of evidence strongly suggest that tomatinase-degrading enzymes have been acquired independently by different tomato pathogens. First, tomatinases of fungal origin differ in their mechanism to remove sugar moieties from the glycoalkaloid. Second, predicted amino acid sequences from the two tomatinase genes cloned so far suggest that these enzymes belong to divergent families of glycosyl hydrolases, sharing few structural features. These results, together with the fact that certain fungal pathogens of other plant species unrelated to tomato, also produce tomatinase activity [89], raise intriguing questions about the nature of the evolutionary mechanisms that may have lead to host plant specificity in pathogenic fungi.
In contrast to enzymatic degradation, nondegradative mechanisms of tomatine tolerance have received relatively little attention so far. The importance in pathogenicity attributed to these mechanisms is likely to increase in the future, as the molecular bases of tolerance will become clearer. Tomatine offers a suitable model for such studies since it is commercially available, tolerated by a considerable number of fungal tomato pathogens and has a mode of action relatively well known. It appears clear that membrane sterol content is a major factor in tomatine resistance [28, 29], but other mechanisms may also play an important role in the capacity of fungi to tolerate high concentrations of the glycoalkaloid. A common phenomenon in fungi involves the appearance of resistance to multiple, structurally unrelated fungitoxic compounds. Thus, mutants of the saprophytic fungus Aspergillus nidulans that were resistant to the fungicide thiazole also showed increased tolerance to tomatine [120]. Resistance to a broad range of antifungal compounds is mediated by ATP-binding cassette (ABC) membrane transport proteins [121]. It is conceivable that phytopathogenic fungi may use this kind of efflux mechanism to prevent buildup of high intracellular concentrations of phytoanticipins [122]. A predicted ABC transporter gene in the saprophytic fungus A, nidulans has recently been isolated and shown to

322
be induced by the pea phytoalexin pisatin [120]. More compellingly, a transformant of the rice blast pathogen Magnaporthe grisea carrying a disrupted copy of an ABC transporter gene has dramatically reduced pathogenicity towards rice plants [123]. It is likely that even more, currently unknown mechanisms exist in fungi to withstand the toxicity of tomatine. The recent identification of several tomatinase-induced genes from F. oxysporum, all of them encoding predicted intracellular polypeptides, suggests that the presence of the saponin triggers major changes in fungal metabolism. To understand these changes and how they contribute to tolerance of tomatine and pathogenicity to plants is an exciting challenge for future research.
ABBREVIATIONS
RT-PCR = Reverse transcription-polymerase chain reaction. TLC = Thin layer chromatography.
ACKNOWLEDGMENTS
This work was supported by grants from The European Commission (BIOTECH, Contract No. BIO2-CT94-3001) and from Junta de Andalucia (group 3084).
REFERENCES
[1] VanEtten, H.D.; Mansfield, J.W.; Bailey, J.A.; Fanner, E.E. Plant Cell, 1994, 9, 1191.
[2] Price, K.R.; Johnson, I.T.; Fenwick, G.R. CRC Crit. Rev. Food Sci. Nutrition, 1987, 26, 27.
[3] Hostettmann, K.; Hostettmann, M.; Marston, A. Method Plant Biochem., 1991,7,435.
[4] Fenwick, G.R.; Price, K.R.; Tsukamato, C; Okubo, K. In Toxic substances in Crop Plants ; D'Mello, J.P., Duffus, CM. & Dufus, J.H., Ed.; The Royal Society of Chemistry: Cambridge, 1992; Vol. pp. 285.
[5] Hostettmann, K.; Marston, A. Saponins., Cambridge University Press: Cambridge, 1995.
[6] Osbourn, A. Trends Plant Sci., 1996, 1, 4. [7] Roddick, J. Phytochemistry, 1974, 13, 9. [8] Friedman, M.; Bautista, F.F.; Stanker, L.H.; Larkin, K.A. J. Agric. Food
C/7ew.,1998,46, 5097.

323
[9] Maga, J.A. Food Rev, International, 1994, 10, 385. [10] Fontaine, T.D.; Irving, G.W.; Ma, R.; Pool, J.B.; Doolittle, S.P. Arch,
Biochem., 194S, 18,467. [11] Eltayeb, E.A.; Roddick, J.G. J. Exp. Bot., 1984, 35, 252. [12] Kozukue, N.; Kozukue, E.; Yamashita, H.; Fujii, S. J, Food ScL, 1994, 59,
1211. [13] Friedman, M.; Levin, C.E. J. Agric. Food Chem,, 1995, 43, 1507. [14] Friedman, M.; Levin, C.E. J. Agric, Food Chem,, 1998, 46, 4571. [15] Ameson, P.A.; Durbin, R.D. Phytopathology, 1968, 58, 536. [16] Jadhav, S.; Sharma, R.P.; Salunkhe, D.K. Crit. Rev, Toxicol,, 1981, 9, 21. [17] Sandrock, R.W.; Vanetten, H.D. Phytopathology, 1998, 88, 137. [18] Lacey, L.A.; Mercadier, G. Mycopathologia, 1998, 142, 17. [19] Gallardo, F.; Boethel, D.J. J. Entomol, Sci,, 1990, 25, 376. [20] Weissenberg, M.; Levy, A.; Svoboda, J.A.; Ishaaya, L Phytochemistry, 1998,
47, 203. [21] Sanford, L.L.; Domek, J.M.; Cantelo, W.W.; Kobayashi, R.S.; Sinden, S.L.
Amer, Potato J,, 1996, 73, 79. [22] Duffey, S.S.; Stout, M.J. Arch, Insect Biochem, Physiol, 1996, 32, 3. [23] Sharma, O.N.; Bajaj, K.L. Plant Disease Res,, 1994, 9, 80. [24] Friedman, M.; Raybum, J.R.; Bantle, J.A. J, Agric. Food Chem., 1992,40,
1617. [25] Gee, J.; Wortley, G.M.; Johnson, I.T.; Price, K.R.; Rutten, A.A.J.J.; Houben,
G.F.; Penninks, A.H. Toxicol in Vitro., 1996, 10, 117. [26] Stanker, L.H.; Kamps-Holtzapple, C; Friedman, M. Agr, Food Chem,, 1994,
42, 2360. [27] Roddick, J.G.; Drysdaie, R.B. Phytochemistry, 1984, 23, 543. [28] Steel, C.C; Drysdaie, R.B. Phytochemistry, 1988, 27, 1025. [29] Keukens, E.A.J.; de Vrije, T.; van de Boom, C ; de Ward, P.; Plasman, H.H.;
Thiei, F.; Chupin, V.; Jongen, W.M.F.; de Kruijff, B.E. Biochim. Biophys, Acta,l995, 1240,216.
[30] Osbourn, A.E. Plant Cell, 1996, 8, 1821. [31] Osbourn, A.E. Fungal Genet, Biol, 1999, 26, 163. [32] Osbourn, A.; Bowyer, P.; Lunness, P.; Clarke, B.; Daniels, M. Mol Plant-
Microbe Interact., 1995, 8, 971. [33] Sandrock, R.W.; Dellapenna, D.; Vanetten, H.D. Mol Plant-Microbe Interact,,
1995, 8, 960. [34] Roldan-Arjona, T.; Perez-Espinosa, A.; Ruiz-Rubio, M. Mol Plant-Microbe
Interact., 1999, 12, 852. [35] Lairini, K.; Perez-Espinosa, A.; Pineda, M.; Ruiz-Rubio, M. Appl Environ,
Microbiol, 1996, 62, 1604. [36] Quidde, T.; Osbourn, A.E.; Tudzynski, P. Physiol Mol Plant Pathol, 1998,
52, 151. [37] Lairini, K.; Ruiz-Rubio, M. Mycol Res., 1998, 102, 1375. [38] Lairini, K.; Ruiz-Rubio, M. Phytochemistry, 1997, 45, 1371. [39] Friedman, M.; Kozukue, N.; Harden, L.A. J, Agric, Food Chem,, 1998, 46,
2096. [40] Sander, H. Planta, 1956, 47, 374. [41] Verhoeff, K. Phytopatol Z , 1975, 82, 333. [42] Schl5sser, E. Z PJlkr, Pfls,, 1975, 82, 476.

324
[43] Tukalo, E.A. Sb. Nauchn. Tr. Dnepropetr. Med. Inst., 1958, 6, 371. [44] Courtney, W.H.; Lambeth, V.N. HorstScience, 1977, 12, 550. [45] Juvick, J.A.; Stevens, M.A. J. Am. Soc. Horde. Sei., 1982, 107, 1061. [46] Juvick, J.A.; Stevens, M.A.; Rick, CM. HortSeienee, 1982, 17, 764. [47] Rick, CM.; Uhlig, J.W.; Jones, A., D. Proe. Natl. Aead. Set. USA, 1994, 91,
12877. [48] Bushway, R.J.; Perkins, L.B.; Paradis, L.R.; Van Derpan, S. J. Agrie. Food
C//e/w., 1994, 42, 2824. [49] Friedman, M.; Levin, C.E.; Mcdonald, G.M. J. Agrie. Food Chem., 1994,
42, 1959. [50] Keukens, E.A.J.; Hop, M.E.C.M.; Jongen, W.M.F. J. Agr. Food Chem., 1994,
42, 2475. [51] Kajderowicz-Jarosinska, D. Aeta Agr. Silvestria Ser. Roln., 1965, 5, 3. [52] Roddick, J.G.; Butcher, D.N. Phytoehemistry, 1972, ,2991. [53] Roddick, J.G. Phytoehemistry, 1976, 15, 475. [54] Pegg, G.F.; Woodward, S. Physiol. Mol. Plant Pathol., 1986, 28, 333. [55] Smith, CA.; MacHardy, W.E. Phytopathology, 1982, 72, 415. [56] Papadopoulou, K.; Melton, R.E.; Legget, M.; Daniels, M.J.; Osboum, A.E.
Proe. Natl. Aead Sei. USA., 1999, 96, 12923. [57] Arneson, P.A.; Durbin, R.D. Plant Physiol., 1968, 43, 683. [58] Roddick, J.G. J. Exp. Botany, 1978, 29, 1371. [59] Schultz, G.; Sander, H. Z Physiol. Chem., 1957, 308, 122. [60] Sch5nbeck, F.; Schl5sser, E. In Physiologieal Plant Pathology; Heitefus, R. &
Williams, P.H., Ed.; Springer-Verlag: Berlin, 1976; pp. 653-678. [61] Segal, R.; Schl5sser, E. Areh. Mierobiol, 1975, 104, 147. [62] Blankemeyer, J.T.; White, J.B.; Stringer, B.K.; Friedman, M. Food Chem.
ToxieoL, 1991, 35, 639. [63] Diener, U.L. Phytopathology, 1955, 45, 654. [64] Dow, J.W.; Callow, J.A. Phytopathol. Z , 1978, 92, 211. [65] Sohi, H.S.; Sokhi, S.S. Indian Phytopathol., 1972, 26, 666. [66] Rotem, J. The genus Alternaria: Biology, Epidemiology, and Pathogenieity,
The American Phytopathological Societ: St. Paul, MN., 1994. [67] Schl5sser, E. Phytopathol. Z , 1972, 92, 211. [68] Olsen, R.A. Physiologia Plantarum, 1973, 27, 202. [69] Defago, G.; Kern, H.; Sedlar, L. Physiol. Plant Pathol., 1983, 22, 39. [70] Defago, G.; Kern, H. Physiol. Plant Pathol., 1983,22, 29. [71] Ellis, S., W.; Rose, M.E.; Grindle, M. J. Gen. Mierobiol., 1991, 137, 2627. [72] Grindle, M. Mol. Gen. Genet., 1973, 120, 283. [73] Apama, K.; Sandrock, R.W.; Kasbekar, D.P. J. Genet., 1998, 77, 71. [74] Turner, E.M. J. Exp. Bot., 1961, 12, 169. [75] Crombie, W.M.L.; Crombie, L.; Green, J.B.; Lucas, J.A. Phytochemistry,
1986, 25, 2075. [76] Osbourn, A.E.; Clarke, B.R.; Dow, J.M.; Daniels, M.J. Physiol. Mol Plant
Pathol.,\99l,3i,3^\. [77] Bowyer, P.; Clarke, B.R.; Lunness, P.; Daniels, M.J.; Osboum, A.E. Science,
1995,267,371. [78] Weltring, K.M.; Vessels, J.; Geyen, R. Phytochemistry, 1997, 46, 1005. [79] Becker, P.; Weltring, K.M. FEMS Microbiol. Letters, 1998, 167, 197. [80] Arneson, P.A.; Durbin, R.D. Phytopathology, 1967, 57, 1358.

325
[81] Durbin, R.D.; Uchytil, T.F. Biochim, Biophys. Acta, 1969, 191, 176. [82] Schlosser, E. Acta PhytopathoL Acad ScL Hung., 1975, 10, 77. [83] Ford, J.E.; McCance, D.J.; Drysdale, R.B. Phytochemistry, 1977, 16, 545. [84] Henrissat, B. Biochem. J., 1991, 280, 309. [85] Henrissat, B.; Bairoch, A. Biochem. J., 1993, 293, 781. [86] Melton, R.E.; Flegg, L.M.; Brown, J.K.M.; Oliver, R.P.; Daniels, M.J.;
Osboum, A.E. Mol Plant-Microbe Interact., 1998, 11, 228. [87] Fiedler, K.; Simons, K. Cell, 1995, 81, 309. [88] Langcake, P.; Drysdale, R.B.; Smith, H. Physiol. Plant Pathol, 1972, 2, 36. [89] Lairini, K.; Perez-Espinosa, A.; Ruiz-Rubio, M. Physiol. Mol. Plant Pathol,
1997, 50, 37. [90] Fewell, A.M.; Roddick, J.G. Phytochemistry, 1993, 33, 23. [91] Weltring, K.M.; Wessels, J.; Pauli, G.F. Phytochemistry, 1998, 48, 1321. [92] Deacon, J.W. Introduction to modern mycology, Blacwell Scientific
Publications: Oxford, 1984. [93] Gilkes, N.R.; Henrissat, B.; Kilbum, D.G.; Miller, R.C.; Warren, R.A.J.
Microbiol Rev., 1991, 55, 303. [94] Henrissat, B.; Bairoch, A. Biochem. J., 1996, 316, 695. [95] Davies, G.; Henrissat, B. Structure, 1995, 3, 853. [96] Kulmburg, P.; Mathieu, M.; Dowzer, C ; Kelly, J.; Felenbok, B. Mol
Microbiol, 1993,7, S41, [97] Quidde, T.; Buttner, P.; Tudzynski, P. European J. Plant Pathol, 1999, 105,
273. [98] Wubben, J.P.; Price, K.R.; Daniels, M.J.; Osboum, A.E. Phytopathology,
1996,86,986. [99] Sato, Y.; Hayakawa, S. J. Organic Chem., 1963, 28, 2739. [100] Sato, Y.; Hayakawa, S. J. Organic Chem., 1963, 28 [101] Sato, Y.; Hayakawa, S. J. Organic Chem., 1964, 29, 198. [102] Znidarsic, P.; Vitas, M.; Komel, R.; Pavko, A. Physiol Mol Plant Pathol,
1999,55,251. [103] Ghosh, D.K.; Samanta, T.B. J. Steroid Biochem., 1981, 14, 1063. [104] Smith, K.E.; Ahmed, F.; Williams, R.A.D.; Kelly, S.L. J. Steroid Biochem.
Mol Biol, 1994,49, 93. [105] Breskvar, K.; Hudnik-Plevnik, T. J. Steroid Biochem., 1981, 14, 395. [106] Smith, K.E.; Latif, S.A.; Kirk, D.N. J. Steroid Biochem. Mol Biol, 1991, 38,
249. [107] Smith, K.E.; White, K.A.; Kirk, D.N. J. Steroid Biochem., 1989, 33, 81. [108] Smith, K.E.; Latif, S.A.; Kirk, D.N. J. Steroid Biochem., 1989, 33, 927. [109] Vitas, M.; Smith, K.E.; Plavec, J.; Kesselmeier, J.; Pajic, T.; Ferlan, A.;
Zigon, D.; Kelly, S.L.; Komel, R. Chemosphere, 1999, 38, 853. [110] Vitas, M.; Smith, K.; Rozman, D.; Komel, R. J. Steroid Biochem. Mol Biol,
1994,48, 87. [111] Liang, P.; Pardee, A.B. Science, 1992, 257, 967. [112] Dharlingue, A.; Mamdouh, A.M.; Malfatti, P.; Soulie, M.C.; Bompeix, G.
Phytochemistry, 1995, 39, 69. [113] Suleman, P.; Tohamy, A.M.; Saleh, A.A.; Madkour, M.A.; Straney, D.C.
Physiol Mol Plant Pathol, 1996,48, 131. [114] Roddick, J.G, Phytochemistry, 1979, 18, 1467.

326
[115] Safe, L.M.; Safe, S.H.; Subden, R.E.; Morris, D.C. Canadian J. Microbiol., 1977, 23, 398.
[116] Colot, V. In Genetic Engineering ; Selow, J.K., Ed.; Plenum Press: New York, 1990; Vol. 12, pp. 225-242.
[117] VanEtten, J.L.; Freer, S.N.; McCune, B.K. J. BacterioL, 1979, 138, 650. [118] Petersen, G.R.; Dahlberg, K.R.; Van Etten, J.L. J, BacterioL, 1983, 155, 601. [119] Chen, L.X.; Fischer, H.J. Plant Physiol,, 1998, 152, 38. [120] Del Sorbo, G.; Andrade, A.C.; Van Nistelrooy, J.G.M.; Van Kan, J.A.L.; Balzi,
E.; De Waard, M.A. MoL Gen, Genet,, 1997, 254, 417. [121] Balzi, E.; Goffeau, A. Biochim, Biophys, Acta, 1994, 1187, 152. [122] De Waard, M.A. Pest, ScL, 1997, 51, 271. [123] Urban, M.; Bhargava, T.; Hamer, J. EMBO J,, 1999, 18, 512.




















