
Immunostimulatory DNA Inhibits Transforming Growth Factor Expression and Airway Remodeling

Our reference: YCIMM 2814 P-authorquery-v8
AUTHOR QUERY FORM
Journal: YCIMM
Article Number: 2814
Please e-mail or fax your responses and any corrections to:
E-mail: [email protected]
Fax: +31 2048 52799
Dear Author,
Please check your proof carefully and mark all corrections at the appropriate place in the proof (e.g., by using on-screen annotation in the PDFfile) or compile them in a separate list.
For correction or revision of any artwork, please consult http://www.elsevier.com/artworkinstructions.
Any queries or remarks that have arisen during the processing of your manuscript are listed below and highlighted by flags in the proof. Clickon the ‘Q’ link to go to the location in the proof.
Location inarticle
Query / Remark: click on the Q link to goPlease insert your reply or correction at the corresponding line in the proof
Q1 This section comprises references that occur in the reference list but not in the body of the text. Pleaseposition each reference in the text or, alternatively, delete it. Any reference not dealt with will be retainedin this section.
Thank you for your assistance.

1
2
3
4
5
67
8
1 0
11121314
151617181920
2 1
45
46
47
48
49
50
Cellular Immunology xxx (2010) xxx–xxx
YCIMM 2814 No. of Pages 10, Model 5G
29 November 2010
Contents lists available at ScienceDirect
Cellular Immunology
journal homepage: www.elsevier .com/locate /yc imm
Induction of immunostimulatory cytokine genes expression in human PBMCsby a novelsemiquinone glucoside derivative (SQGD) isolated from a Bacillus sp. INM-1
Raj Kumar a,⇑, Rakesh Kumar Sharma a, Deen Dayal Bansal a, Dev Dutt Patel a, Saurabh Mishra a,Lyuba Miteva b, Zlatka Dobreva b, Veselina Gadjeva b, Spaska Stanilova b
a Institute of Nuclear Medicine and Allied Sciences, Delhi, Indiab Faculty of Medicine, Trakia University, Stara Zagora, Bulgaria
a r t i c l e i n f o
22232425262728293031
Article history:Received 21 July 2010Accepted 15 November 2010Available online xxxx
Keywords:ImmunostimulationCytokinePBMCsSemiquinone glucoside derivative
3233343536373839404142
0008-8749/$ - see front matter � 2010 Published bydoi:10.1016/j.cellimm.2010.11.005
Abbreviations: C3bgp, C3 binding glycoprotein; DCdiphenyl 2-picrylhydrazyl; EDTA, ethlene diamine teferon-gamma; IL, interleukin; kGy, kilo gray; LPS, lipohistocompatibility antigen; MTCC, microbial type cultfactor kappa beta; NK, natural killer cells; PBMC, pecells; PBS, phosphate buffer saline; PHA, phytoagglutinqRT-PCR, quantitative real-time polymerase chain realiosis homolog A; TLR4, toll like receptor 4; KLH, keyh⇑ Corresponding author. Address: Institute of N
Sciences, Brig. S.K. Mazumdar Road, Delhi 110054, InE-mail address: [email protected] (R. Kum
Please cite this article in press as: R. Kumar etglucoside derivative (SQGD) isolated from a Baci
a b s t r a c t
In the present study, a semiquinone glucoside derivative (SQGD) isolated from a radioresistant bacteriumBacillus sp. INM-1 was evaluated for its immunostimulatory activities. Human peripheral blood mononu-clear cells (PBMCs) were stimulated by different doses (30–90 lg/ml) of SQGD for different time (3–12 h)intervals at 37 �C, and IL-12p40, IL-23p19, IL-10, RelA and c-Jun gene expression analysis was carried outby qRT-PCR method. SQGD dose dependent cytokines protein expression kinetic analysis was carried outusing Western blotting. As the results of SQGD (30 lg/ml) stimulation for 3 h at 37 �C, significant induc-tion in IL-12p40, IL-23p19 and RelA gene expression was observed in PBMCs compared to unstimulatedcontrol cells. However, no such induction in IL-10 and c-Jun gene expression was observed. Time depen-dent protein expression study indicated significant increase in IL-12p40, IL-12p35, IL-23p19 and RelAprotein expression at 3–6 h, which was found decrease at 12 h upon SQGD treatment. In contrast,IL-10 protein expression was found to enhance significantly at 12 h after SQGD treatment to the PBMCs.SQGD dose dependent study showed approximately similar level of induction in IL-12p40, IL-12p35,IL-23p19 and RelA proteins expression at all tested concentration (30–90 lg/ml) compared to control.However, no significant change in the IL-10 and c-Jun protein expression was observed at any SQGDconcentration. SQGD treatment (0.25 mg/kg b wt.) was also found to enhance anti-keyhole LimpetHemocynin (KLH) IgM antibodies significantly in the mice immunized by KLH.
Thus, SQGD fraction stimulates cellular immunity by inducing immunostimulatory cytokines andhumoral immunity by enhancing IgM antibodies and could be a promising immunostimulant. Furtherstudies related to molecular mechanisms offering immunostimulation is underway, will certainly helpfulto unravel its mode of action in the biological system.
� 2010 Published by Elsevier Inc.
43
44
51
52
53
54
55
56
1. Introduction
Cytokines, the key modulators of immune response delivereddiverse functions in both humoral and T cell mediated immunity[1–3]. Among the cytokines, IL-12, IL-23 and IL-10 have beenreported to play a very crucial role in the regulation of pro/anti-
57
58
59
60
61
62
63
64
65
66
67
Elsevier Inc.
s, dendritic cells; DPPH, 1,1-tra acetic acid; IFN-c, inter-polysaccharide; MHC, major
ure collection; NFjb, nuclearripheral blood mononuclearin; PWM, pok weed mitogen;ction; RelA, reticuloendothe-ole limpet hemocyanin.
uclear Medicine and Allieddia. Fax: +91 11 23919509.ar).
al., Induction of immunostimulllus sp. INM-1, Cell. Immunol. (
inflammatory responses induced by microbial infections, oxidativestress, septic shock and ionizing radiation [4–7]. Human IL-12(IL-12p70) is a disulfide-linked heterodimer composed of two sub-units p40 and p35. Earlier studies have defined that IL-12p70 is animportant factor exhibits numerous immune functions includingstimulation of T cells and natural killer (NK) cells, production ofIFN-c and regulation of several cell adhesion molecules [8–10].Various reports demonstrated a vital role of IL-12p70 in the devel-opment of specific immunity against number of intracellularpathogens including Leishmania major, Mycobacterium tuberculosis,Listeria monocytogenes, and Toxoplasma gondii [11–13]. Apart fromthat IL-12 has also been shown to have adjuvant properties, andcan stimulate an effective cellular immune response against micro-bial antigens that are otherwise not immunogenic when adminis-tered alone. Besides immunostimulatory properties, IL-12p70 alsoexhibits anti-tumor activity in several preclinical animal tumormodels [14]. Interleukin-23 (IL-23) is a heterodimeric cytokine
atory cytokine genes expression in human PBMCs by a novel semiquinone2010), doi:10.1016/j.cellimm.2010.11.005

68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
2 R. Kumar et al. / Cellular Immunology xxx (2010) xxx–xxx
YCIMM 2814 No. of Pages 10, Model 5G
29 November 2010
(p40/p19) consisting of a novel p19 subunit and the p40 subunit ofIL-12p70. IL-23 has similar but discrete functions from IL-12p70.IL-23 induces activated memory T cells to proliferate and produceIFN-c. In addition, IL-23 induces production of a pro-inflammatorycytokine, IL-17, from activated T cells [15].
In contrast to IL-12 and IL-23, IL-10 has immunosuppressiveeffect and inhibits Th1 cytokine synthesis from activatedmacrophages [16]. IL-10 is known to produce by a variety of cells,including activated antigen presenting cells, CD4+ Th0 and Th2cells. Up-regulated IL-10 gene expression induced oxidative stressin the septic shock patients [16,17] and lead to mortality. Similarly,higher expression of IL-10 induced immunosupression in thecancer patients [18,19]. It has been reported that the up regulationof IL-10 gene was concomitantly down regulate the activation ofIL-12 and IL-23 gene expression [20–22] and thus lead toimmunosuppression.
Therefore, to combat the IL-10 mediated immunosuppression,induction of IL-12 and IL-23 genes expression are highly desirableto initiate the Th1 cellular immune response in the cancer patientsexposed with radiation during radiotherapy regime. Various natu-ral compounds like lipopolysaccharide of Gram negative bacteria,peptidoglycan, lipoteichoic acid, N-acetyl glucosaminyl glycan,b 1,4 linked N-acetylmuramic acid of gram positive bacteria,C3binding glycoprotein (C3bgp) isolated from Cascuta europea,pokweed mitogen (PWM) a lectin isolated from Phytollacaamericana and phytoagglutinin (PHA) have been identified andevaluated for their abilities to stimulate Th1 mediated immune re-sponse by induction of IL-12 and IL-23 cytokine gene expression[4,17,23–25].
Present study was under taken to evaluate a semiquinone glu-coside derivative rich fraction (SQGD) isolated from Bacillus sp.INM-1 for its immunomostimultory activities using human PBMCs.Modulation of IL-12p40 and IL-23p19 genes expression was stud-ied and compared with immunosuppressive cytokine IL-10 geneexpression using qRT-PCR methods. The effect of SQGD on Th1 im-mune response, regulatory transcription factor RelA and c-Jun geneexpression was evaluated and compared with LPS and C3bgp med-iated gene expression. To further validate the qRT-PCR based geneexpression profile of IL-12p40, IL-12p35, IL-23p19, RelA, IL-10 andc-Jun, time dependent protein expression studies also carried out.Effect of SQGD on keyhole limpet hemocynin (KLH) stimulated hu-moral response was also evaluated in vivo using Swiss Albino malemice.
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
2. Material and methods
2.1. Reagent
Lipopolysaccharides (LPS) from Escherichia coli serotype 026:B6,Antibodies, Histopaque-1077, RPMI-1640 medium, PHA, PWM etc.were procured from Sigma, USA and all cell culture plastic warewere purchased from Nunc, Denmark. C3 binding glycoprotein(C3bgp) was isolated at Trakia University, Stara Zagora, Bulgariafrom a water extract of the seeds of the parasitic plant Cuscutaeuropea by affinity chromatography as described [26].
SQGD, an endotoxin free semiquinone glucoside derivative(SQGD) rich fractions was isolated from Bacillus sp. INM-1 (MTCCNo 1026) at our laboratory at INMAS, Delhi, India.
179
180
181
182
183
2.2. PBMC isolation
The blood samples were collected from five healthy volunteersafter obtaining consent form each of them and diluted (1:1 ratio)with sterile phosphate buffer saline (PBS). The diluted blood sam-ples were carefully transferred to histopaque-1077 followed by
Please cite this article in press as: R. Kumar et al., Induction of immunostimulglucoside derivative (SQGD) isolated from a Bacillus sp. INM-1, Cell. Immunol. (
centrifugation at 1800 rpm at 4 �C for 20 min. The interface ofhistopaque-1077 and diluted blood samples containing peripheralblood mononuclear cells (PBMC) was gently harvested and washedtwice with sterile cold serum-free RPMI-1640 medium. Afterwashing, cell pellet of PBMC was resuspended in RPMI-1640medium and cells viability was tested with Trypan blue exclusionmethod.
2.3. Cell culture and stimulation
PBMCs (1 � 106) were cultured in sterile 24-well polystyreneplates containing 2.0 ml RPMI-1640 media supplemented with100 U/ml penicillin, 100 lg/ml gentamicin and 0.3 mg/ml L-gluta-mine. The cultures were stimulated with 30 lg/ml SQGD, 30 lg/mlC3 binding glycoprotein (C3bgp), and 30 lg/ml LPS. Both stimulatedand unstimulated cultures were incubated at 37 �C for 3 h.
2.4. RNA isolation
PBMCs (1 � 106 cells) stimulated with SQGD, C3bgp, or LPS for3 h, were used for RNA isolation. Unstimulated cells were used ascalibrators. After incubation, cells were washed and resuspendedin lysis solution RL supplied with a column-based In-Prep bloodRNA isolation kit (AJ Roboscreen, Leipzig; Germany). Total RNA iso-lation was performed according to the manufacturer’s instructions.To remove traces of genomic DNA, total RNAs (1 lg) were treatedwith RNase-free DNase I (Fermentas, USA) as per the manufacture’sinstruction.
2.5. Reverse transcription
Synthesis of cDNA was performed manually as per the instruc-tions supplied by manufacturers with high-capacity cDNA Archivekit (Applied Biosystems, USA) that uses random primers andMultiScribeTM MuLV reverse transcriptase enzyme. Incubate for10 min at 25 �C followed by 2 h at 37 �C.
2.6. Quantitative real-time polymerase chain reaction (qRT-PCR)
Quantitative real-time polymerase chain reaction (qRT-PCR) wasperformed on a 7500 Real-Time PCR System (Applied Biosystems,CA, USA). The following validate PCR primers and TaqManMGB probes (6FAM-labeled) were used: IL-12p40 (assay ID:Hs00233688_m1), IL-23p19 (Hs00372324_m1), IL-10 (Hs00174086_m1), RelA (Hs00153294_m1) and c-Jun (Hs99999141_s1).Eukaryotic 18S ribosomal RNA (Hs99999903_m1) was used asendogenous control. An aliquot of 5 ll of the RT reaction was ampli-fied in duplicate in final volume of 20 ll of TaqMan Universal PCRmaster mix and gene expression assay mix, containing specific for-ward and reverse primers and labeled probes for target genes andendogenous control (Applied Biosystems, USA).The thermocyclingconditions were applied as followed: initial 10 min incubation at95 �C followed by 40 cycles of denaturation for 15 s at 95 �C andextension for 1 min at 60 �C. PCR data were collected with SequenceDetection System software, version 1.3.1. Relative quantitative eval-uation of cytokine mRNAs was performed by the comparative DDCtmethod. The mean DCt obtained in unstimulated PBMC for eachcytokine mRNA was used as calibrator, after normalization toendogenous control-18S rRNA. The results are presents as an n-folddifference relative to calibrator (RQ = 2�DDCt).
2.7. Kinetics of cytokines protein expression at different SQGDconcentrations
PBMCs were stimulated with different doses (30–90 lg/ml) ofSQGD at 37 �C temperature for 3, 6 and 12 h. After completion of
atory cytokine genes expression in human PBMCs by a novel semiquinone2010), doi:10.1016/j.cellimm.2010.11.005
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
R. Kumar et al. / Cellular Immunology xxx (2010) xxx–xxx 3
YCIMM 2814 No. of Pages 10, Model 5G
29 November 2010
the incubation, cells were harvested by centrifugation at 5000 rpmfor 10 min. Cells were then sonicated (two cycles of 50 Hz) in PBSand total soluble proteins were extracted by centrifugation(5000 rpm, 20 min) at 4 �C.
2.7.1. Protein estimationTotal soluble protein in the cell lysate was estimated by
Bradford method (28) and the amount (lg) of protein was quanti-fied using BSA standard curve.
2.7.2. SDS–PAGE and western blot analysisFor SDS–PAGE and western blot analysis of IL-12p40, IL-12p35,
IL-23p19, RelA, IL-10 and c-Jun protein expression was carried outusing the protocols standardized in our laboratory (29). IL-12p40,IL-12p35, IL-23p19, RelA, IL-10 and c-Jun expression levels wereanalyzed by probing with respective mouse monoclonal antibodies(1:1000 dilution) having cross reactivity with human proteins. Theprotein bands obtained were further subjected to densitometric
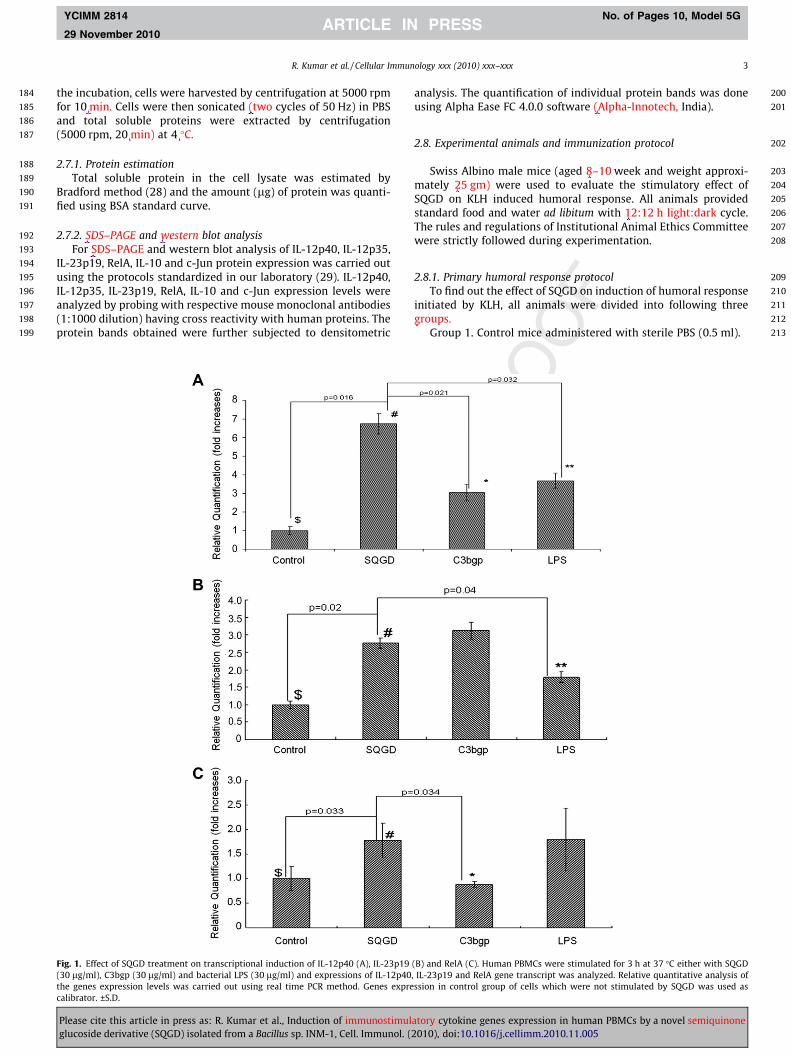
Fig. 1. Effect of SQGD treatment on transcriptional induction of IL-12p40 (A), IL-23p19(30 lg/ml), C3bgp (30 lg/ml) and bacterial LPS (30 lg/ml) and expressions of IL-12p40the genes expression levels was carried out using real time PCR method. Genes exprecalibrator. ±S.D.
Please cite this article in press as: R. Kumar et al., Induction of immunostimulglucoside derivative (SQGD) isolated from a Bacillus sp. INM-1, Cell. Immunol. (
analysis. The quantification of individual protein bands was doneusing Alpha Ease FC 4.0.0 software (Alpha-Innotech, India).
2.8. Experimental animals and immunization protocol
Swiss Albino male mice (aged 8–10 week and weight approxi-mately 25 gm) were used to evaluate the stimulatory effect ofSQGD on KLH induced humoral response. All animals providedstandard food and water ad libitum with 12:12 h light:dark cycle.The rules and regulations of Institutional Animal Ethics Committeewere strictly followed during experimentation.
2.8.1. Primary humoral response protocolTo find out the effect of SQGD on induction of humoral response
initiated by KLH, all animals were divided into following threegroups.
Group 1. Control mice administered with sterile PBS (0.5 ml).
(B) and RelA (C). Human PBMCs were stimulated for 3 h at 37 �C either with SQGD, IL-23p19 and RelA gene transcript was analyzed. Relative quantitative analysis ofssion in control group of cells which were not stimulated by SQGD was used as
atory cytokine genes expression in human PBMCs by a novel semiquinone2010), doi:10.1016/j.cellimm.2010.11.005
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
4 R. Kumar et al. / Cellular Immunology xxx (2010) xxx–xxx
YCIMM 2814 No. of Pages 10, Model 5G
29 November 2010
Group 2. The first injection of KLH antigen (50 lg) was given atday 0 and booster dose (50 lg) of the KLH antigen in saline wasadministered (i.p.) to the mice on days 7.
Group 3. Mice administered with primary (at day 0) and sec-ondary (at day 7) doses of KLH antigen (50 lg) were subsequentlyadministered with different (250, 500 and 750 lg) concentrationsof SQGD.
Seven days after final treatment, all the mice were sacrificed.Blood was collected and anti-KLH IgM and IgG antibody titer inthe serum were estimated.
240
241
242
243
244
245
246
2.8.2. ELISA for serum anti-KLH antibodies estimationThe surface of 96-well Maxisorb immunoplates (Nunc,
Danmark) were coated with 10 lg KLH per ml of 0.5 M sodiumcarbonate buffer (pH 9.6) and incubated overnight at 4 �C. Then,plates were washed, kept at room temperature and blocked for1 h with PBS containing 1% BSA and subjected for two washes withPBS. The plates were then incubated with a 1:100 dilution of
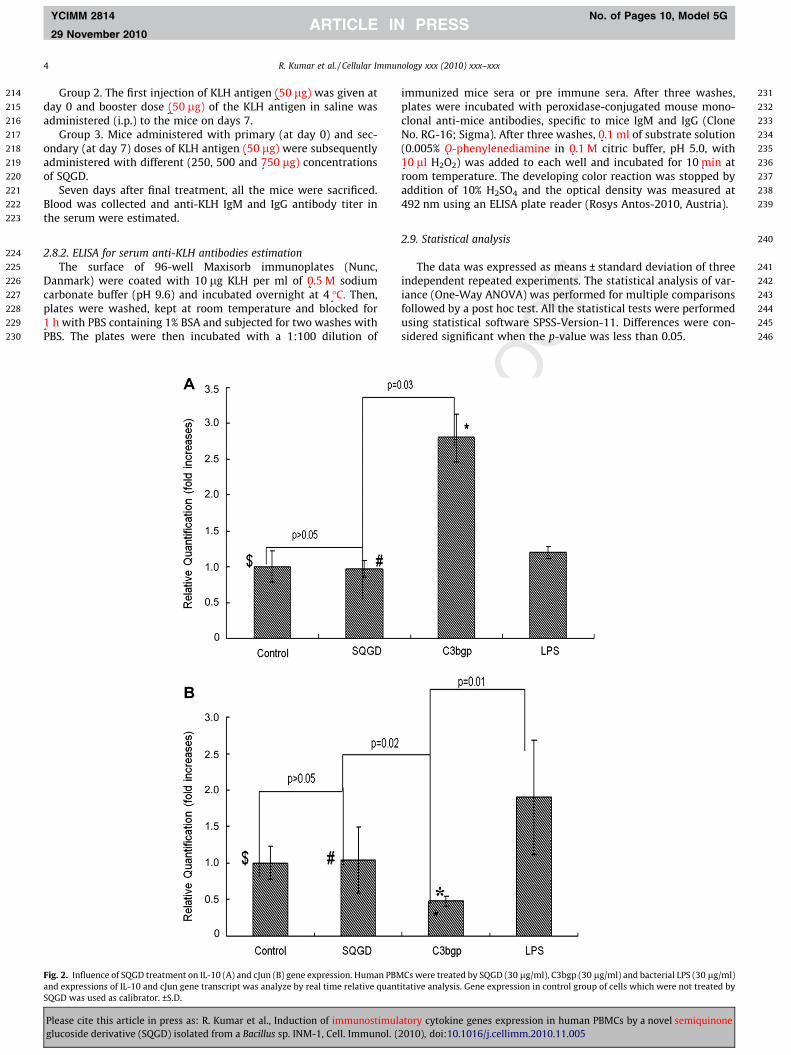
Fig. 2. Influence of SQGD treatment on IL-10 (A) and cJun (B) gene expression. Human PBMand expressions of IL-10 and cJun gene transcript was analyze by real time relative quantSQGD was used as calibrator. ±S.D.
Please cite this article in press as: R. Kumar et al., Induction of immunostimulglucoside derivative (SQGD) isolated from a Bacillus sp. INM-1, Cell. Immunol. (
immunized mice sera or pre immune sera. After three washes,plates were incubated with peroxidase-conjugated mouse mono-clonal anti-mice antibodies, specific to mice IgM and IgG (CloneNo. RG-16; Sigma). After three washes, 0.1 ml of substrate solution(0.005% O-phenylenediamine in 0.1 M citric buffer, pH 5.0, with10 ll H2O2) was added to each well and incubated for 10 min atroom temperature. The developing color reaction was stopped byaddition of 10% H2SO4 and the optical density was measured at492 nm using an ELISA plate reader (Rosys Antos-2010, Austria).
2.9. Statistical analysis
The data was expressed as means ± standard deviation of threeindependent repeated experiments. The statistical analysis of var-iance (One-Way ANOVA) was performed for multiple comparisonsfollowed by a post hoc test. All the statistical tests were performedusing statistical software SPSS-Version-11. Differences were con-sidered significant when the p-value was less than 0.05.
Cs were treated by SQGD (30 lg/ml), C3bgp (30 lg/ml) and bacterial LPS (30 lg/ml)itative analysis. Gene expression in control group of cells which were not treated by
atory cytokine genes expression in human PBMCs by a novel semiquinone2010), doi:10.1016/j.cellimm.2010.11.005
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
R. Kumar et al. / Cellular Immunology xxx (2010) xxx–xxx 5
YCIMM 2814 No. of Pages 10, Model 5G
29 November 2010
3. Results
3.1. Induction of IL-12p40 cytokine gene expression by SQGD
The quantitative real time gene expression analysis of IL-12p40was carried out using human PBMCs. Cells were stimulated withSQGD for 3 h, and level of IL-12p40 gene transcription analyzedand compared with the untreated control cells as well as withthe cells stimulated with known immunostimulatory agents (i.e.,LPS and C3bgp). The results were calculated and presented as thefold increase compared to unstimulated controls. As the resultsof SQGD treatment, maximum (p = 0.016) induction in the tran-scription of IL-12p40 (5.74-fold increase) gene was observed in hu-man PBMCs as compared to unstimulated control (Fig. 1A).Similarly, significant induction in IL-12p40 gene expression wasobserved in the group of cells stimulated with SQGD as comparedto the cells stimulated with C3bgp (decreased by 2.7-fold;p = 0.021) and LPS (decreased by 1.7-fold; p = 0.032), respectively(Fig. 1A).
280
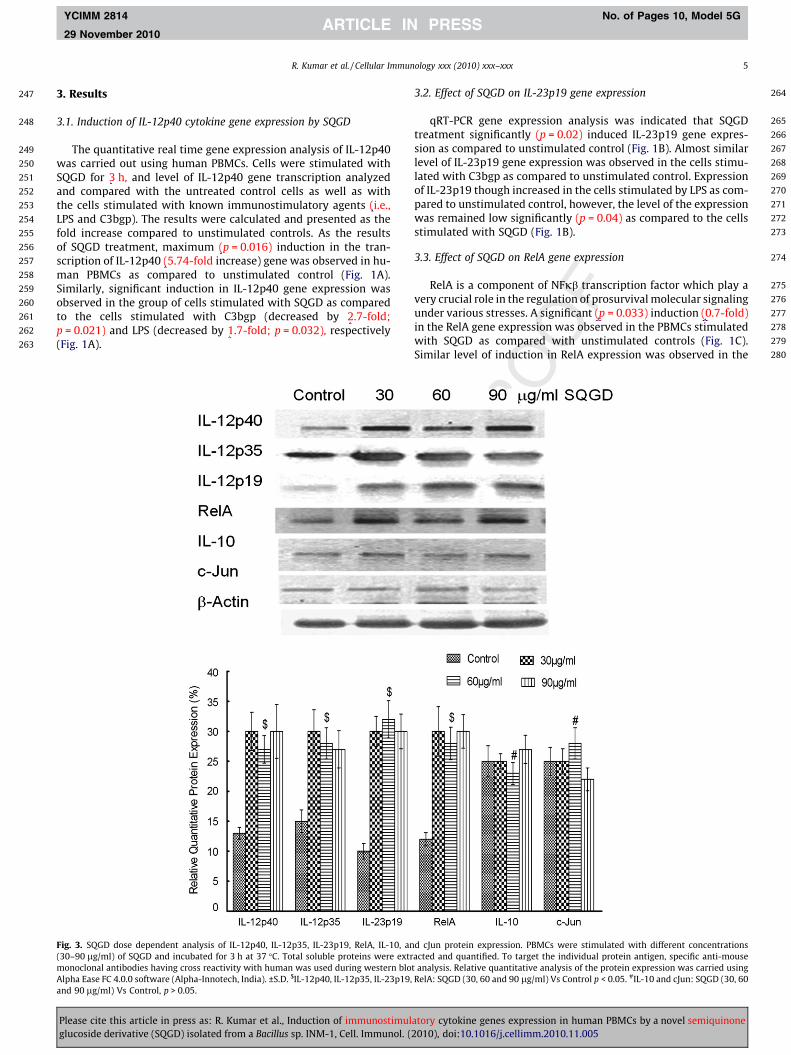
Fig. 3. SQGD dose dependent analysis of IL-12p40, IL-12p35, IL-23p19, RelA, IL-10, an(30–90 lg/ml) of SQGD and incubated for 3 h at 37 �C. Total soluble proteins were extrmonoclonal antibodies having cross reactivity with human was used during western bloAlpha Ease FC 4.0.0 software (Alpha-Innotech, India). ±S.D. $IL-12p40, IL-12p35, IL-23p19,and 90 lg/ml) Vs Control, p > 0.05.
Please cite this article in press as: R. Kumar et al., Induction of immunostimulglucoside derivative (SQGD) isolated from a Bacillus sp. INM-1, Cell. Immunol. (
3.2. Effect of SQGD on IL-23p19 gene expression
qRT-PCR gene expression analysis was indicated that SQGDtreatment significantly (p = 0.02) induced IL-23p19 gene expres-sion as compared to unstimulated control (Fig. 1B). Almost similarlevel of IL-23p19 gene expression was observed in the cells stimu-lated with C3bgp as compared to unstimulated control. Expressionof IL-23p19 though increased in the cells stimulated by LPS as com-pared to unstimulated control, however, the level of the expressionwas remained low significantly (p = 0.04) as compared to the cellsstimulated with SQGD (Fig. 1B).
3.3. Effect of SQGD on RelA gene expression
RelA is a component of NFjb transcription factor which play avery crucial role in the regulation of prosurvival molecular signalingunder various stresses. A significant (p = 0.033) induction (0.7-fold)in the RelA gene expression was observed in the PBMCs stimulatedwith SQGD as compared with unstimulated controls (Fig. 1C).Similar level of induction in RelA expression was observed in the
d cJun protein expression. PBMCs were stimulated with different concentrationsacted and quantified. To target the individual protein antigen, specific anti-mouset analysis. Relative quantitative analysis of the protein expression was carried usingRelA: SQGD (30, 60 and 90 lg/ml) Vs Control p < 0.05. #IL-10 and cJun: SQGD (30, 60
atory cytokine genes expression in human PBMCs by a novel semiquinone2010), doi:10.1016/j.cellimm.2010.11.005
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
6 R. Kumar et al. / Cellular Immunology xxx (2010) xxx–xxx
YCIMM 2814 No. of Pages 10, Model 5G
29 November 2010
cells stimulated with LPS. However, significant (p = 0.034) decreasein the RelA expression was noticed in the cells stimulated withC3bgp as compared to SQGD (p = 0.034). Therefore, results of thepresent study indicated that SQGD was equally induce the transcrip-tion of RelA gene as LPS and thus may share the similar mechanismof RelA or NFjb activation as adopted by LPS.
3.4. Influence of SQGD on IL-10 and c-Jun genes expression
No significant (p > 0.05) change in the level of IL-10 and c-Jungene expression was observed in the cells after 3 h, stimulationwith SQGD as compared to unstimulated controls (Fig. 2A, B).Though expression of IL-10 was found to be increased (�1.7-fold)significantly (p = 0.03) in the cells stimulated with C3bgp as com-pared to SQGD stimulated cells. However, the expression of c-Jungene was observed to be higher in the LPS stimulated cells as com-pared to the cells treated with C3bgp and SQGD (Fig. 2B).
3.5. Kinetics of cytokines expression after stimulation of PBMCs by SQGD
To evaluate the effect of different concentrations (30–90 lg/ml)of SQGD on cytokines protein expression, western blot analysis
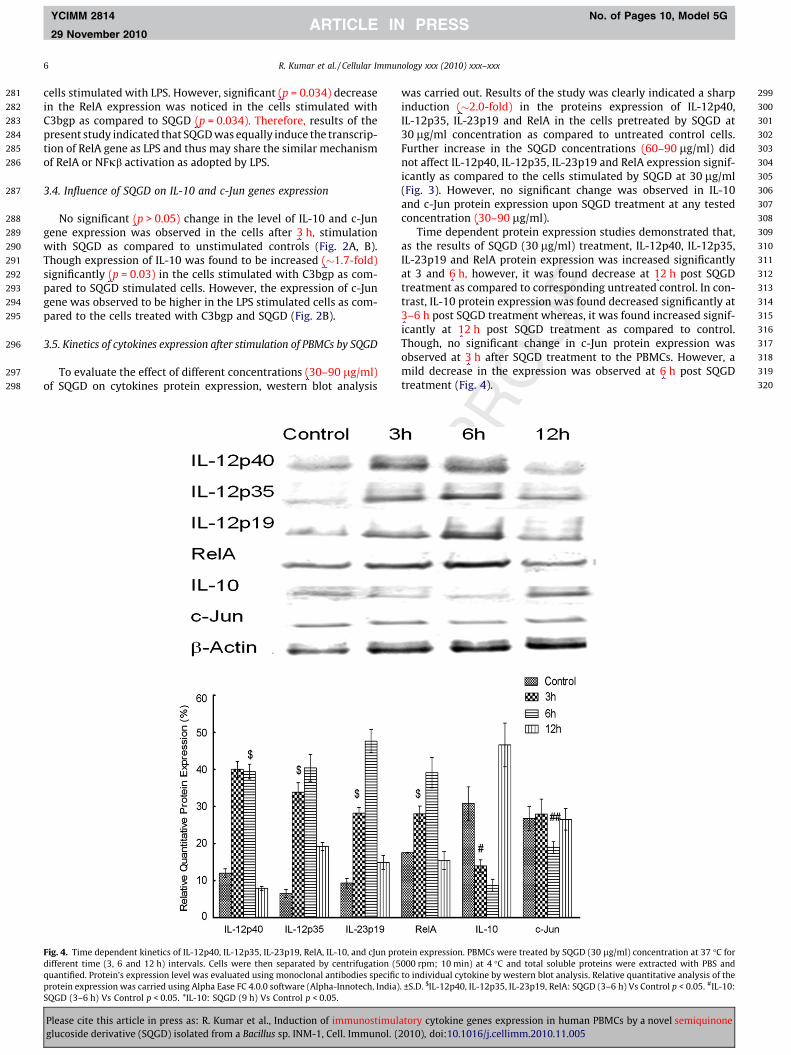
Fig. 4. Time dependent kinetics of IL-12p40, IL-12p35, IL-23p19, RelA, IL-10, and cJun prodifferent time (3, 6 and 12 h) intervals. Cells were then separated by centrifugation (5quantified. Protein’s expression level was evaluated using monoclonal antibodies specificprotein expression was carried using Alpha Ease FC 4.0.0 software (Alpha-Innotech, India)SQGD (3–6 h) Vs Control p < 0.05. *IL-10: SQGD (9 h) Vs Control p < 0.05.
Please cite this article in press as: R. Kumar et al., Induction of immunostimulglucoside derivative (SQGD) isolated from a Bacillus sp. INM-1, Cell. Immunol. (
was carried out. Results of the study was clearly indicated a sharpinduction (�2.0-fold) in the proteins expression of IL-12p40,IL-12p35, IL-23p19 and RelA in the cells pretreated by SQGD at30 lg/ml concentration as compared to untreated control cells.Further increase in the SQGD concentrations (60–90 lg/ml) didnot affect IL-12p40, IL-12p35, IL-23p19 and RelA expression signif-icantly as compared to the cells stimulated by SQGD at 30 lg/ml(Fig. 3). However, no significant change was observed in IL-10and c-Jun protein expression upon SQGD treatment at any testedconcentration (30–90 lg/ml).
Time dependent protein expression studies demonstrated that,as the results of SQGD (30 lg/ml) treatment, IL-12p40, IL-12p35,IL-23p19 and RelA protein expression was increased significantlyat 3 and 6 h, however, it was found decrease at 12 h post SQGDtreatment as compared to corresponding untreated control. In con-trast, IL-10 protein expression was found decreased significantly at3–6 h post SQGD treatment whereas, it was found increased signif-icantly at 12 h post SQGD treatment as compared to control.Though, no significant change in c-Jun protein expression wasobserved at 3 h after SQGD treatment to the PBMCs. However, amild decrease in the expression was observed at 6 h post SQGDtreatment (Fig. 4).
tein expression. PBMCs were treated by SQGD (30 lg/ml) concentration at 37 �C for000 rpm; 10 min) at 4 �C and total soluble proteins were extracted with PBS andto individual cytokine by western blot analysis. Relative quantitative analysis of the. ±S.D. $IL-12p40, IL-12p35, IL-23p19, RelA: SQGD (3–6 h) Vs Control p < 0.05. #IL-10:
atory cytokine genes expression in human PBMCs by a novel semiquinone2010), doi:10.1016/j.cellimm.2010.11.005
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
R. Kumar et al. / Cellular Immunology xxx (2010) xxx–xxx 7
YCIMM 2814 No. of Pages 10, Model 5G
29 November 2010
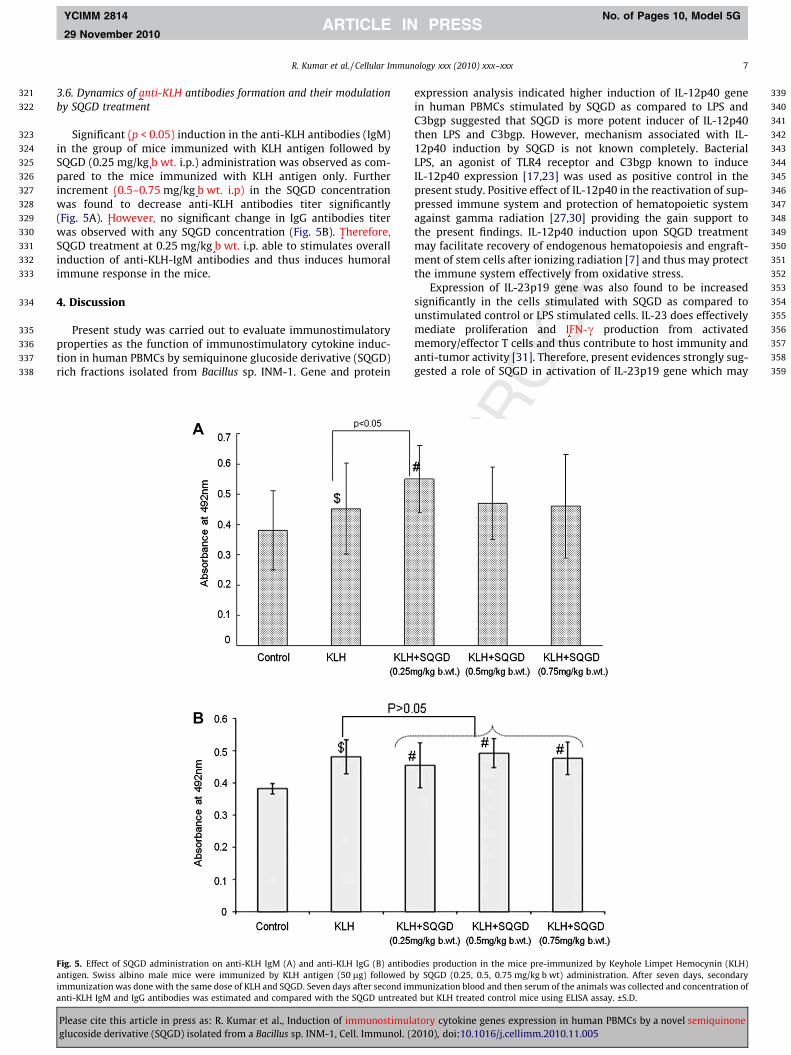
3.6. Dynamics of anti-KLH antibodies formation and their modulationby SQGD treatment
Significant (p < 0.05) induction in the anti-KLH antibodies (IgM)in the group of mice immunized with KLH antigen followed bySQGD (0.25 mg/kg b wt. i.p.) administration was observed as com-pared to the mice immunized with KLH antigen only. Furtherincrement (0.5–0.75 mg/kg b wt. i.p) in the SQGD concentrationwas found to decrease anti-KLH antibodies titer significantly(Fig. 5A). However, no significant change in IgG antibodies titerwas observed with any SQGD concentration (Fig. 5B). Therefore,SQGD treatment at 0.25 mg/kg b wt. i.p. able to stimulates overallinduction of anti-KLH-IgM antibodies and thus induces humoralimmune response in the mice.
4. Discussion
Present study was carried out to evaluate immunostimulatoryproperties as the function of immunostimulatory cytokine induc-tion in human PBMCs by semiquinone glucoside derivative (SQGD)rich fractions isolated from Bacillus sp. INM-1. Gene and protein
Fig. 5. Effect of SQGD administration on anti-KLH IgM (A) and anti-KLH IgG (B) antiboantigen. Swiss albino male mice were immunized by KLH antigen (50 lg) followed bimmunization was done with the same dose of KLH and SQGD. Seven days after second imanti-KLH IgM and IgG antibodies was estimated and compared with the SQGD untreate
Please cite this article in press as: R. Kumar et al., Induction of immunostimulglucoside derivative (SQGD) isolated from a Bacillus sp. INM-1, Cell. Immunol. (
expression analysis indicated higher induction of IL-12p40 genein human PBMCs stimulated by SQGD as compared to LPS andC3bgp suggested that SQGD is more potent inducer of IL-12p40then LPS and C3bgp. However, mechanism associated with IL-12p40 induction by SQGD is not known completely. BacterialLPS, an agonist of TLR4 receptor and C3bgp known to induceIL-12p40 expression [17,23] was used as positive control in thepresent study. Positive effect of IL-12p40 in the reactivation of sup-pressed immune system and protection of hematopoietic systemagainst gamma radiation [27,30] providing the gain support tothe present findings. IL-12p40 induction upon SQGD treatmentmay facilitate recovery of endogenous hematopoiesis and engraft-ment of stem cells after ionizing radiation [7] and thus may protectthe immune system effectively from oxidative stress.
Expression of IL-23p19 gene was also found to be increasedsignificantly in the cells stimulated with SQGD as compared tounstimulated control or LPS stimulated cells. IL-23 does effectivelymediate proliferation and IFN-c production from activatedmemory/effector T cells and thus contribute to host immunity andanti-tumor activity [31]. Therefore, present evidences strongly sug-gested a role of SQGD in activation of IL-23p19 gene which may
dies production in the mice pre-immunized by Keyhole Limpet Hemocynin (KLH)y SQGD (0.25, 0.5, 0.75 mg/kg b wt) administration. After seven days, secondarymunization blood and then serum of the animals was collected and concentration ofd but KLH treated control mice using ELISA assay. ±S.D.
atory cytokine genes expression in human PBMCs by a novel semiquinone2010), doi:10.1016/j.cellimm.2010.11.005

360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422423424425426427428429430431432433434435436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484485486487488489490491492493494495496497498499500501502503504
Q1
8 R. Kumar et al. / Cellular Immunology xxx (2010) xxx–xxx
YCIMM 2814 No. of Pages 10, Model 5G
29 November 2010
delivered diverse functions to enhance the cellular immunity andcell proliferation in the surrounding normal tissues damagedduring radiotherapy of the cancer patients without effecting tumorregression rate [32,33].
RelA (p65) is a subunit of NFjb, form a heterodimer with sisterpeptide p50 and act as transcription factor. Expression of RelA geneand corresponding protein was found to be increased significantlyin the cells expressing higher level of IL-12p40 and IL-23p19 geneafter SQGD stimulation (Figs. 1A–C, 3 and 4). This observation wassuggested an involvement of SQGD in activation of prosurvivalNFjb signal transduction pathway. NFjb has already been knownfor its selective up regulation of pro-inflammatory cytokine genes[34,35] further provided a gain support to the present investiga-tion. Though, expression of IL-12p40 and IL-23p19 was increased(Fig.1A, B) significantly, while, no significant change in IL-10 geneexpression was noticed in the PBMCs stimulated by SQGD at 3 h(Fig. 2A). However, significant decrease in IL-10 protein expressionat 6 h after SQGD treatment (Fig. 4) suggested immunostimulatorypotential of SQGD. Down regulation of IL-10 synthesis acceleratesIL-12 secretion by induction of IL-12 secreting cell population[36]. Hoffmann and co-workers [37] reported that in the absenceof IL-10, mice exposed with radiation attenuated larvae developedTh1 type cytokine (IL-12) profile and high level of T cells mediatedimmunity against Schistosoma mansoni infection. Similar resultswere obtained in our previous study conducted on the septic shocksurvivor’s, which, revealed significant reduction in IL-10 expres-sion along with higher level of IL-12p40 expression in humanPBMCs stimulated by C3bgp compared to non survivors [17] pro-vides a gain support to the present investigation. Apart from cellmediated primary immune response, SQGD was also evaluatedfor its secondary immune response stimulatory properties. Resultsof the study showed significant induction in the level of anti-KLHantibodies in the serum of the mice treated with KLH antigenfollowed by SQGD administration as compared to only KLH immu-nized mice (Fig. 5A, B) suggested that SQGD effectively potentiatethe secondary immune response in the mice immunized with KLHantigen. Similar results were also reported by Stanilova [38] withC3bgp plant derive antigen in the rabbit provided a gain supportto the present investigation.
In conclusion, present study provided strong evidence thatsemiquinone glucoside derivative (SQGD) rich fraction stimulatesprimary immune response by inducing IL-12p40, IL-12p35,IL-23p19 and RelA cytokine genes and corresponding proteinwithout significantly effecting IL-10 cytokine gene expressionand protein in the human PBMCs. SQGD was also found to acceler-ate secondary immune response by inducing anti-KLH IgMantibodies in the mice. Thus in future, this novel biologically activesemiquinone glucoside derivative may qualify as a potentialimmunostimulatory agent of therapeutic significance.
5. Uncited references
Refs. [28,29].
Acknowledgments
The work was carried out under the aegis of Indo–Bulgariacollaborative project (B-57) financially supported by the Depart-ment of Science and Technology (DST), Government of India Delhi,India and Ministry of Education, Science and Technology, Govern-ment of Bulgaria. Author is highly thankful to the Defence Researchand Development Organization (DRDO), Ministry of Defence, Govt.of India and Trakia University, Stara Zagora, Bulgaria to providefinancial support and the basic facilities to undertake the presentwork.
Please cite this article in press as: R. Kumar et al., Induction of immunostimulglucoside derivative (SQGD) isolated from a Bacillus sp. INM-1, Cell. Immunol. (
References
[1] K. Takeda, S. Akira, Toll-like receptors in innate immunity, Int. Immunol. 17(2005) 1–14.
[2] S. Akira, K. Takeda, Toll-like receptor signaling, Nat. Rev. Immunol. 4 (2004)499–511.
[3] N. Tokumasa, A. Suto, S. Kagami, S. Furuta, K. Hirose, N. Watanabe, Y. Saito, K.Shimoda, I. Iwamoto, H. Nakajima, Expression of Tyk2 in dendritic cells isrequired for IL-12, IL-23, and IFN-gamma production and the induction of Th1cell differentiation, Blood 110 (2007) 553–560.
[4] S.A. Stanilova, Z.T. Karakolev, G.S. Dimov, Z.G. Dobreva, L.D. Miteva, E.S. Slavov,C.S. Stefanov, N.S. Stanilov, High interleukin 12 and low interleukin 10production after in vitro stimulation detected in sepsis survivors, IntensiveCare Med. 31 (2005) 401–407.
[5] K.G. Hogg, S. Kumkate, A.P. Mountford, IL-10 regulates early IL-12-mediatedimmune responses induced by the radiation-attenuated schistosome vaccine,Int. Immunol. 15 (2003) 1451–1459.
[6] K.G. Hogg, S. Kumkate, S. Anderson, A.P. Mountford, Interleukin-12 p40secretion by cutaneous CD11c+ and F4/80+ cells is a major feature of the innateimmune response in mice that develop Th1-mediated protective immunity toSchistosoma mansoni, Infect. Immun. 71 (2003) 3563–3571.
[7] T. Chen, K.A. Burke, Y. Zhan, X. Wang, D. Shibata, Y. Zhao, IL-12 facilitates boththe recovery of endogenous hematopoiesis and the engraftment of stem cellsafter ionizing radiation, Exp. Hematol. 35 (2007) 203–213.
[8] G. Gri, C. Chiodoni, E. Gallo, A. Stoppacciaro, F.Y. Liew, M.P. Colombo,Antitumor effect of interleukin (IL)-12 in the absence of endogenous IFN-gamma: A role for intrinsic tumor immunogenicity and IL-15, Cancer Res. 62(2002) 4390–4397.
[9] E.A. Lafleur, S.F. Jia, L.L. Worth, Z. Zhou, L.B. Owen-Schaub, E.S. Kleinerman,Interleukin (IL)-12 and IL-12 gene transfer up-regulate Fas expression inhuman osteosarcoma and breast cancer cells, Cancer Res. 61 (2001) 4066–4071.
[10] C. Holscher, The power of combinatorial immunology: IL-12 and IL-12-relateddimeric cytokines in infectious diseases, Med. Microbiol. Immunol. 193 (2004)1–17.
[11] A.M. Cooper, A.D. Roberts, E.R. Rhoades, J.E. Callahan, D.M. Getzy, I.M. Orme,The role of interleukin-12 in acquired immunity to Mycobacterium tuberculosisinfection, Immunology 84 (1995) 423–432.
[12] F.P. Heinzel, D.S. Schoenhaut, R.M. Rerko, L.E. Rosser, M.K. Gately, Recombinantinterleukin 12 cures mice infected with Leishmania major, J. Exp. Med. 177(1993) 1505–1509.
[13] C.S. Hsieh, S.E. Macatonia, C.S. Tripp, S.F. Wolf, A. O’Garra, K.M. Murphy,Development of TH1 CD4+ T cells through IL-12 produced by Listeria-inducedmacrophages, Science 260 (1993) 547–549.
[14] C.M. Coughlin, M. Wysocka, G. Trinchieri, W.M. Lee, The effect of interleukin 12desensitization on the antitumor efficacy of recombinant interleukin 12,Cancer Res. 57 (1997) 2460–2467.
[15] C.S. Lankford, D.M. Frucht, A unique role for IL-23 in promoting cellularimmunity, J. Leukoc. Biol. 73 (2003) 49–56.
[16] P.R. Lowe, H.F. Galley, A. Abdel-Fattah, N.R. Webster, Influence of interleukin-10 polymorphisms on interleukin-10 expression and survival in critically illpatients, Crit. Care Med. 31 (2003) 34–38.
[17] S.A. Stanilova, Z.G. Dobreva, E.S. Slavov, L.D. Miteva, C3 binding glycoproteinfrom Cuscuta europea induced different cytokine profiles from human PBMCcompared to other plant and bacterial immunomodulators, Int.Immunopharmacol. 5 (2005) 723–734.
[18] K. Kawamura, R. Bahar, W. Natsume, S. Sakiyama, M. Tagawa, Secretion ofinterleukin-10 from murine colon carcinoma cells suppresses systemicantitumor immunity and impairs protective immunity induced against thetumors, Cancer Gene Ther. 9 (2002) 109–115.
[19] S. Mocellin, F.M. Marincola, H.A. Young, Interleukin-10 and the immuneresponse against cancer: A counterpoint, J. Leukoc. Biol. 78 (2005)1043–1051.
[20] M. Aste-Amezaga, X. Ma, A. Sartori, G. Trinchieri, Molecular mechanisms of theinduction of IL-12 and its inhibition by IL-10, J. Immunol. 160 (1998) 5936–5944.
[21] B. Oppmann, R. Lesley, B. Blom, J.C. Timans, Y. Xu, B. Hunte, F. Vega, N. Yu, J.Wang, K. Singh, F. Zonin, E. Vaisberg, T. Churakova, M. Liu, D. Gorman, J.Wagner, S. Zurawski, Y. Liu, J.S. Abrams, K.W. Moore, D. Rennick, R. de Waal-Malefyt, C. Hannum, J.F. Bazan, R.A. Kastelein, Novel p19 protein engages IL-12p40 to form a cytokine, IL-23, with biological activities similar as well asdistinct from IL-12, Immunity 13 (2000) 715–725.
[22] L. Miteva, S. Stanilova, The combined effect of interleukin (IL)-10 and IL-12polymorphisms on induced cytokine production, Hum. Immunol. 69 (2008)562–566.
[23] Z.G. Dobreva, S.A. Stanilova, The immunomodulatory activity of C3 bindingglycoprotein (C3bgp) is mediated by the complement receptor type III andmitogen-activated protein kinase signal transduction pathways,Immunopharmacol. Immunotoxicol. 29 (2007) 549–562.
[24] M. Noorizadeh, J. Hadjati, A. Khabiri, M. Vodjgani, H. Khadem-Shariat, Effectsof gamma irradiation on proliferation and IL-5 production of peripheral bloodlymphocytes, Iran. Biomed. J. 8 (2004) 211–214.
[25] R. Schwandner, R. Dziarski, H. Wesche, M. Rothe, C.J. Kirschning,Peptidoglycan- and lipoteichoic acid-induced cell activation is mediated bytoll-like receptor 2, J. Biol. Chem. 274 (1999) 17406–17409.
atory cytokine genes expression in human PBMCs by a novel semiquinone2010), doi:10.1016/j.cellimm.2010.11.005

505506507508509510511512513514515516517518519520521522523524525
526527528529530531532533534535536537538539540541542543544545546
R. Kumar et al. / Cellular Immunology xxx (2010) xxx–xxx 9
YCIMM 2814 No. of Pages 10, Model 5G
29 November 2010
[26] Z.D. Zhelev, S.A. Stanilova, B.G. Carpenter, Isolation, partial characterisationand complement inhibiting activity of a new glycoprotein from Cuscutaeuropea, Biochem. Biophys. Res. Commun. 202 (1994) 186–194.
[27] A. Schwarz, A. Maeda, K. Kernebeck, H. van Steeg, S. Beissert, T. Schwarz,Prevention of UV radiation-induced immunosuppression by IL-12 isdependent on DNA repair, J. Exp. Med. 201 (2005) 173–179.
[28] M.M. Bradford, A rapid and sensitive method for the quantification ofmicrogram quantities of protein utilizing the principle of protein-dyebinding, Anal. Biochem. 72 (1976) 248–254.
[29] P.K. Singh, R. Kumar, A. Sharma, R. Arora, R. Chawla, S.K. Jain, R.K. Sharma,Podophyllum hexandrum fraction (REC-2006) shows higher radioprotectiveefficacy in the p53-carrying hepatoma cell line: A role of cell cycle regulatoryproteins, Integr. Cancer Ther. 8 (2009) 261–272.
[30] M. Neta, S.M. Stiefe, F. Finkelman, S. Herrmann, N. Ali, IL-12 protects bonemarrow from and sensitizes intestinal tract to ionizing radiation, J. Immunol.153 (1994) 4230–4237.
[31] J.S. Hao, B.E. Shan, Immune enhancement and anti-tumour activity of IL-23,Cancer Immunol. Immunother. 55 (2006) 1426–1431.
[32] A. Billiau, H. Heremans, K. Vermeire, P. Matthys, Immunomodulatoryproperties of interferon-gamma. An update, Ann. N. Y. Acad. Sci. 856 (1998)22–32.
547
Please cite this article in press as: R. Kumar et al., Induction of immunostimulglucoside derivative (SQGD) isolated from a Bacillus sp. INM-1, Cell. Immunol. (
[33] R.G. Hawley, L.C. Berger, Growth control mechanisms in multiple myeloma,Leuk. Lymphoma 29 (1998) 465–475.
[34] A.E. Medvedev, T. Flo, R.R. Ingalls, D.T. Golenbock, G. Teti, S.N. Vogel, T.Espevik, Involvement of CD14 and complement receptors CR3 and CR4 innuclear factor-kappaB activation and TNF production induced by lipopoly-saccharide and group B streptococcal cell walls, J. Immunol. 160 (1998)4535–4542.
[35] K. De Bosscher, W. Vanden Berghe, G. Haegeman, The interplay between theglucocorticoid receptor and nuclear factor-kappaB or activator protein-1:Molecular mechanisms for gene repression, Endocr. Rev. 24 (2003)488–522.
[36] K.W. Moore, R. de Waal Malefyt, R.L. Coffman, A. O’Garra, Interleukin-10 andthe interleukin-10 receptor, Annu. Rev. Immunol. 19 (2001) 683–765.
[37] K.F. Hoffmann, S.L. James, A.W. Cheever, T.A. Wynn, Studies with doublecytokine-deficient mice reveal that highly polarized Th1- and Th2-typecytokine and antibody responses contribute equally to vaccine-inducedimmunity to Schistosoma mansoni, J. Immunol. 163 (1999) 927–938.
[38] S.A. Stanilova, L.D. Miteva, S.G. Tanchev, Immunomodulatory effects of C3bgpon the antibody response to hemocyanin in outbred rabbits and the F1generation of breeding with siblings, Vet. Immunol. Immunopathol. 106(2005) 15–21.
atory cytokine genes expression in human PBMCs by a novel semiquinone2010), doi:10.1016/j.cellimm.2010.11.005
Copyright © 2022 FDOKUMEN




















