
The immunostimulatory activity of phosphorothioate CpG oligonucleotides is affected by distal...
8
Molecular Immunology 48 (2011) 1027–1034 Contents lists available at ScienceDirect Molecular Immunology journal homepage: www.elsevier.com/locate/molimm The immunostimulatory activity of phosphorothioate CpG oligonucleotides is affected by distal sequence changes Tara L. Roberts a,b,∗ , Jasmyn A. Dunn a,c , Matthew J. Sweet a , David A. Hume d , Katryn J. Stacey a,c a The University of Queensland, Institute for Molecular Bioscience, Qld 4072, Australia b Radiation Biology and Oncology Laboratory, Queensland Institute of Medical Research, 300 Herston Road, Brisbane 4006, Australia c The University of Queensland, School of Chemistry and Molecular Biosciences, Qld 4072, Australia d The Roslin Institute, University of Edinburgh, Roslin EH259PS, Scotland, United Kingdom article info Article history: Received 26 November 2010 Received in revised form 14 January 2011 Accepted 17 January 2011 Available online 16 February 2011 Keywords: CpG TLR9 Innate immunity Phosphorothioate oligonucleotides abstract CpG motifs in bacterial DNA activate innate immune cells via toll like receptor 9 (TLR9). Short synthetic oligodeoxynucleotides (ODN) containing a six base CpG motif can mimic the immunostimulatory activity of bacterial DNA. Phosphorothioate (PS) modification of the backbone of ODN makes them more resistant to nuclease degradation and consequently preferable for therapeutic use. Previous studies have shown that the sequence requirements for PS-ODN to have maximal stimulatory activity are more stringent than for normal phosphodiester (PO) ODN. Here we show small sequence changes distal to the CpG motif can affect the activity of PS-ODN whilst having no effect on the activity of PO-ODN. The addition of terminal dG residues and other minor changes to the potently immunostimulatory PS-ODN 1668S caused delayed signalling. The reduction in immunostimulatory activity of PS-ODN was associated with a delay in the activation of MAP kinases. Crown Copyright © 2011 Published by Elsevier Ltd. All rights reserved. 1. Introduction Bacterial and viral DNAs activate the innate immune system due to recognition of “CpG motifs” which are unmethylated CG dinucleotides within particular sequence contexts (Krieg et al., 1995). CpG-containing ODN potently activate the innate immune response and induce a Th1-biased immune response against co- administered antigens when delivered in vivo (Davis et al., 1998). The immunostimulatory activity of CpG ODN has led to clinical tri- als for their use as vaccine adjuvants, and as cancer and allergy therapy (Jurk and Vollmer, 2007). Normal phosphodiester (PO) ODN are rapidly degraded in vivo, so nuclease-resistant phospho- rothioate (PS) ODN, where one of the non-bridging oxygens in the phosphate backbone is replaced by sulfur, are being used in clinical trials. Cellular uptake is an essential step in the response to CpG-containing DNA (Hartmann and Krieg, 2000; Manzel and Macfarlane, 1999). Uptake of DNA initiates TLR9 relocation from the endoplasmic reticulum to the endosome (de Jong et al., 2010; Latz et al., 2004; Sanjuan et al., 2006) where the presence of CpG ∗ Corresponding author at: Radiation Biology and Oncology Laboratory, Queens- land Institute of Medical Research, 300 Herston Road, Brisbane 4006, Australia. Tel.: +61 7 3362 0352; fax: +61 7 3362 0106. E-mail address: [email protected] (T.L. Roberts). motifs can be detected by TLR9 (Hemmi et al., 2000; Latz et al., 2004; Rutz et al., 2004). Evidence suggests that TLR9 binds directly to immunostimulatory DNA (Latz et al., 2007; Rutz et al., 2004). TLR9 then signals via MyD88 to activate MAP kinases, PI-3K and IRFs (Hacker et al., 2000; Park et al., 2002; Sester et al., 2006; Tsujimura et al., 2004; Yi and Krieg, 1998). Induction of these signalling cas- cades results in production of pro-inflammatory cytokines and a Th1 type immune response. PS-modified ODN have some effects not seen with normal PO-ODN. PS-ODN without CpG motifs can cause some TLR9-dependent and -independent cellular activation (Roberts et al., 2005b; Sester et al., 2006; Vollmer et al., 2004b). Long non-CpG containing PS-ODN can also inhibit some TLR9-dependent responses (Trieu et al., 2006). In terms of CpG-dependent responses, PS-ODN have higher uptake than PO-ODN and are more potent in many assays of macrophage responses to CpG DNA (Sester et al., 2000). However we previously showed that a CpG PS-ODN was poorly active in short-term assays of macrophage activation when compared to its PO version. This was attributed to delayed signalling by the PS- ODN, as measured by phosphorylation of ERK and p38 MAP kinases (Sester et al., 2000; Stacey et al., 2000). This data lead to a hypoth- esis that cellular responses to PS-ODN are delayed compared to PO-ODN. Here we show that delayed signalling caused by the PS modification is not a general phenomenon. Delayed signalling by PS-ODN is a sequence-dependent phenomenon, and can be caused by subtle sequence changes distant from the active CpG motif. Our 0161-5890/$ – see front matter. Crown Copyright © 2011 Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.molimm.2011.01.011
-
Upload
westernsydney -
Category
Documents
-
view
0 -
download
0
Transcript of The immunostimulatory activity of phosphorothioate CpG oligonucleotides is affected by distal...

Ta
Ta
b
c
d
a
ARRAA
KCTIP
1
dd1raTatOrpt
CMtL
lT
0d
Molecular Immunology 48 (2011) 1027–1034
Contents lists available at ScienceDirect
Molecular Immunology
journa l homepage: www.e lsev ier .com/ locate /mol imm
he immunostimulatory activity of phosphorothioate CpG oligonucleotides isffected by distal sequence changes
ara L. Robertsa,b,∗, Jasmyn A. Dunna,c, Matthew J. Sweeta, David A. Humed, Katryn J. Staceya,c
The University of Queensland, Institute for Molecular Bioscience, Qld 4072, AustraliaRadiation Biology and Oncology Laboratory, Queensland Institute of Medical Research, 300 Herston Road, Brisbane 4006, AustraliaThe University of Queensland, School of Chemistry and Molecular Biosciences, Qld 4072, AustraliaThe Roslin Institute, University of Edinburgh, Roslin EH259PS, Scotland, United Kingdom
r t i c l e i n f o
rticle history:eceived 26 November 2010eceived in revised form 14 January 2011ccepted 17 January 2011
a b s t r a c t
CpG motifs in bacterial DNA activate innate immune cells via toll like receptor 9 (TLR9). Short syntheticoligodeoxynucleotides (ODN) containing a six base CpG motif can mimic the immunostimulatory activityof bacterial DNA. Phosphorothioate (PS) modification of the backbone of ODN makes them more resistant
vailable online 16 February 2011
eywords:pGLR9nnate immunityhosphorothioate oligonucleotides
to nuclease degradation and consequently preferable for therapeutic use. Previous studies have shownthat the sequence requirements for PS-ODN to have maximal stimulatory activity are more stringent thanfor normal phosphodiester (PO) ODN. Here we show small sequence changes distal to the CpG motif canaffect the activity of PS-ODN whilst having no effect on the activity of PO-ODN. The addition of terminaldG residues and other minor changes to the potently immunostimulatory PS-ODN 1668S caused delayedsignalling. The reduction in immunostimulatory activity of PS-ODN was associated with a delay in theactivation of MAP kinases.
. Introduction
Bacterial and viral DNAs activate the innate immune systemue to recognition of “CpG motifs” which are unmethylated CGinucleotides within particular sequence contexts (Krieg et al.,995). CpG-containing ODN potently activate the innate immuneesponse and induce a Th1-biased immune response against co-dministered antigens when delivered in vivo (Davis et al., 1998).he immunostimulatory activity of CpG ODN has led to clinical tri-ls for their use as vaccine adjuvants, and as cancer and allergyherapy (Jurk and Vollmer, 2007). Normal phosphodiester (PO)DN are rapidly degraded in vivo, so nuclease-resistant phospho-
othioate (PS) ODN, where one of the non-bridging oxygens in thehosphate backbone is replaced by sulfur, are being used in clinicalrials.
Cellular uptake is an essential step in the response to
pG-containing DNA (Hartmann and Krieg, 2000; Manzel andacfarlane, 1999). Uptake of DNA initiates TLR9 relocation fromhe endoplasmic reticulum to the endosome (de Jong et al., 2010;atz et al., 2004; Sanjuan et al., 2006) where the presence of CpG
∗ Corresponding author at: Radiation Biology and Oncology Laboratory, Queens-and Institute of Medical Research, 300 Herston Road, Brisbane 4006, Australia.el.: +61 7 3362 0352; fax: +61 7 3362 0106.
E-mail address: [email protected] (T.L. Roberts).
161-5890/$ – see front matter. Crown Copyright © 2011 Published by Elsevier Ltd. All rioi:10.1016/j.molimm.2011.01.011
Crown Copyright © 2011 Published by Elsevier Ltd. All rights reserved.
motifs can be detected by TLR9 (Hemmi et al., 2000; Latz et al.,2004; Rutz et al., 2004). Evidence suggests that TLR9 binds directlyto immunostimulatory DNA (Latz et al., 2007; Rutz et al., 2004).TLR9 then signals via MyD88 to activate MAP kinases, PI-3K and IRFs(Hacker et al., 2000; Park et al., 2002; Sester et al., 2006; Tsujimuraet al., 2004; Yi and Krieg, 1998). Induction of these signalling cas-cades results in production of pro-inflammatory cytokines and aTh1 type immune response. PS-modified ODN have some effectsnot seen with normal PO-ODN. PS-ODN without CpG motifs cancause some TLR9-dependent and -independent cellular activation(Roberts et al., 2005b; Sester et al., 2006; Vollmer et al., 2004b). Longnon-CpG containing PS-ODN can also inhibit some TLR9-dependentresponses (Trieu et al., 2006).
In terms of CpG-dependent responses, PS-ODN have higheruptake than PO-ODN and are more potent in many assays ofmacrophage responses to CpG DNA (Sester et al., 2000). Howeverwe previously showed that a CpG PS-ODN was poorly active inshort-term assays of macrophage activation when compared to itsPO version. This was attributed to delayed signalling by the PS-ODN, as measured by phosphorylation of ERK and p38 MAP kinases(Sester et al., 2000; Stacey et al., 2000). This data lead to a hypoth-
esis that cellular responses to PS-ODN are delayed compared toPO-ODN. Here we show that delayed signalling caused by the PSmodification is not a general phenomenon. Delayed signalling byPS-ODN is a sequence-dependent phenomenon, and can be causedby subtle sequence changes distant from the active CpG motif. Ourghts reserved.

1028 T.L. Roberts et al. / Molecular Immu
Table 1ODN sequences. The active CG is underlined. The names of PS-modified ODN areindicated in bold and PO-ODN in regular type.
ODN name Sequence
1668S/1668 TCCATGACGTTCCTGATGCTLong1668S/Long1668 GTCCATGACGTTCCTGATGCTGAOS/AO GCTCATGACGTTCCTGATGCTGLAOS/LAO CTCATGACGTTCCTGATGCTGTT GTTCATGACGTTCCTGATGCTGGCC GCCCATGACGTTCCTGATGCTGG1466G GTCAACGTTGAG
dP
2
2
tBtms(gf
2
A
2
treEba
2
wsiedpH
2
(f
assay (Fig. 1E). Long1668S with a 5′ terminal GTC was superior
1466 TCAACGTTGA1555L GCTAGACGTTAGCGTG1555S CTAGACGTTAGCGT
ata emphasise that sequence changes that have no effect in aO-ODN have quite marked effect on PS-ODN activity.
. Methods
.1. Cells and mice
The mouse macrophage cell line RAW264.7 was purchased fromhe American Type Tissue Culture Collection (Manassas, VA, USA).one marrow-derived macrophages (BMMs) were obtained by cul-uring femoral bone marrow cells from adult BALB/c or C57BL/6
ice in the presence of 10,000 U/ml recombinant human colonytimulating factor 1 (hCSF-1) (Chiron, Emeryville, USA) for 7 daysCostelloe et al., 1999). RPMI 1640 medium with 10% FCS, 2 mM l-lutamine, 20 U/ml penicillin, and 20 �g/ml streptomycin was usedor the culture of all cells.
.2. Oligonucleotides
PS- and PO-ODN were obtained from Geneworks (Adelaide,ustralia). Sequences are shown in Table 1.
.3. ELAM promoter assay
The mouse macrophage cell line ELAM9, developed by stableransfection of the RAW264.7 cell line with the human NF-�B-esponsive ELAM promoter driving expression of destabilisednhanced GFP has been described previously (Stacey et al., 2003).LAM9 cells were treated with indicated amount of DNA for 5 hefore harvesting for analysis of GFP expression by flow cytometrys described previously (Stacey et al., 2003).
.4. Western blotting
Cells were lysed in boiling 66 mM Tris–HCl (pH 7.4), 2% SDSith 1 mM sodium pyrophosphate, 10 mM sodium fluoride, 1 mM
odium molybdate, and 1 mM sodium vanadate as phosphatasenhibitors. Western blotting was performed as described (Sestert al., 2000), or using Invitrogen 4–12% pre-cast Bis–Tris gels asescribed previously (Roberts et al., 2009). Antibodies detecting38, phospho-p38, ERK (p42/44) and phospho-ERK and anti-rabbitRP were obtained from Cell Signalling Technology (Beverly, USA).
.5. Oligonucleotide uptake
The ODNs AOS and 1668S were synthesized with a 5′ Cy3 labelGeneworks, Adelaide, Australia). ODN uptake assays were per-ormed as described previously (Roberts et al., 2005a).
nology 48 (2011) 1027–1034
2.6. Down-regulation of the CSF-1 receptor (CSF-1R)
BMMs were starved of CSF-1 overnight to increase surfaceexpression of CSF-1R. Cells were treated with or without ODN for1 h. Cells were then stained for the expression of CSF-1R using theAFS98 monoclonal antibody and analysed by flow cytometry asdescribed previously (Sester et al., 1999).
2.7. Nitrite assay
100,000 RAW264.7 cells/well were plated in 96-well plates in200 �l of complete media and cultured overnight. Cells were thenpretreated with 375 pg/ml of IFN� (R&D Systems, Minneapolis,MI) for 2 h. Cells were then treated as indicated for 24 h. Nitritemeasurement using Griess reagent was performed as describedpreviously (Sweet et al., 1998).
3. Results
3.1. Terminal dG residues reduce the activity of PS-ODN 1668S
In our laboratory two very similar CpG PS-ODN, 1668S and AOShave been routinely used to activate mouse macrophage-like cells.AOS was derived from 1668S (Yi et al., 1996) by the addition ofterminal dG residues and a minor alteration in two bases at the5′ end (Sester et al., 1999). The activities of these two ODN werecompared using the NF-�B dependent reporter cell line ELAM9.Despite minimal sequence difference, there was a significant andunexpected difference in the ability to induce ELAM promoter activ-ity (Fig. 1A). Fig. 1A also shows that the difference in activity wasonly manifested by ODN with a PS backbone and not by PO-ODN.As ODN uptake is a requirement for TLR9-dependent responses wedetermined whether differences in uptake could explain the dif-ferences in ODN activity. Fig. 1B shows no difference in the uptakeof the two PS-ODN that would account for the higher activity of1668S. Uptake of Cy3-AOS was, in fact, reproducibly higher thanuptake of Cy3-1668S. As Cy3-AOS is a longer ODN, enhanced uptakemay be due to increased non-specific binding, a known character-istic of PS-ODN (Brown et al., 1994). As expected, the PO-ODN AOand 1668, which have identical activity, showed no difference inuptake.
The possibility that terminal dG residues on AOS compromisedactivity of the ODN was investigated. Addition of the terminal dGresidues to 1668S to create the ODN Long1668S reduced activityand removal of dG residues from AOS to create the ODN LAOSincreased activity (Fig. 1C) but the same sequence changes to PO-ODN had no effect on activity (Fig. 1D). Thus the addition of terminaldG residues to the PS-ODN 1668S is unfavourable. However, theremoval of terminal dG residues to create LAOS did not generate anODN with the full activity of 1668S, and the small changes at the 5′
end must play some role.
3.2. Small sequence changes can affect PS-ODN activity
In order to try and identify which bases, apart from terminalG, are limiting the activity of AOS, stepwise changes were madeto the sequence of AOS. Changes in bases 2 and 3 from the 5′ endof AOS were made to create the PS-ODN GCC and GTT (Table 1).The activity of these PS-ODN were then determined by ELAM
in activity to GTT, GCC and AOS with a 5′ terminal GCT. In theabsence of terminal dG residues, this sequence preference was pre-served, i.e. 5′ terminal TC was favoured over CT (Fig. 1C, 1668S vsLAOS).

T.L. Roberts et al. / Molecular Immunology 48 (2011) 1027–1034 1029
Fig. 1. Small sequence changes to ODN distal to the CpG motif affect the activity of PS but not PO-ODN. For parts A, C, D and E ELAM9 cells were treated with the indicatedconcentrations of ODN for 5 h and then assayed for ELAM-promoter driven GFP expression by flow cytometry. Experiments were performed two or three times, with a typicalresult shown. In all panels, data points show the average of duplicates within an experiment, and error bars, which generally fall within the size of the symbol, show therange of the values. (A) Immunostimulatory activity of two very similar PS-ODN, 1668S and AOS and the PO-ODN of the same sequence. (B) ELAM9 cells were treated witht ke mee activ( e LAOS le base
3
emctv
he indicated concentration of Cy3-labelled ODN for 1 h and oligonucleotide uptafficiencies of labelling of the ODN. (C) and (D) Terminal dG residues decreased theC) and PO-ODN (D) 1668/S to create Long1668/S, and removed from AO/S to creattepwise changes were made to convert Long1668S to AOS to see the effect of sing
.3. Delayed signalling in response to PS-ODN is sequence specific
Our group has previously reported that the PS-ODN, AOS, gen-
rated delayed ERK and p38 MAP kinase phosphorylation in mouseacrophages compared to a PO-ODN of the same sequence, AO, andonsequently AOS gave poor activation of normal early responseso CpG DNA (Sester et al., 2000; Stacey et al., 2000). These obser-ations led to the hypothesis that PS-ODN in general give delayed
asured by flow cytometry. Results were adjusted to account for slightly differentity of PS but not PO-ODN. dG residues were added to the 5′ and 3′ ends of PS-ODN/S. (E) Small sequence changes distal to the CpG motif can affect PS-ODN activity.changes.
signalling and subsequently a qualitatively different response thanPO-ODN (Sester et al., 2000). Given the superior activity of 1668Sit was investigated whether signalling was induced more rapidly
by it than AOS. Fig. 2 shows the kinetics of responses to AOS and1668S are very different. 1668S triggered maximal p38 phospho-rylation by 20 min but the response to AOS was slight at 20 minand gradually increased over 2 h. The timecourse of signalling inresponse to 1668S was similar to the PO-ODNs AO and 1668 (Fig. 2),
1030 T.L. Roberts et al. / Molecular Immunology 48 (2011) 1027–1034
F Ns AOe o-p38b
aoits
3M
tmiMpoattbew
FSs
ig. 2. Signalling in response to AOS is delayed compared to 1668S and the PO-ODxtracts were made at the times indicated. Samples were probed with anti-phospheen removed from the lower panel which shows results from a single membrane.
lthough 1668S was a more potent stimulus. Thus the previouslybserved delayed signalling in response to AOS (Sester et al., 2000)s not a general property of PS-ODN. We then investigated whetherhe terminal dG residues on AOS were responsible for the delay inignalling.
.4. Terminal dG residues on 1668S or AOS delays induction ofAPK signalling
ERK MAPK and p38 are similarly phosphorylated after 20 min ofreatment with 1668S, although thereafter p38 phosphorylation is
aintained whilst ERK phosphorylation declines (Figs. 2 and 3). Tonvestigate the effect of terminal dG residues, the kinetics of ERK
APK phosphorylation in response to Long1668S, LAOS and theirarent ODNs was determined by Western blotting. With both setsf ODN the presence of terminal dG residues on the ODN resulted inslower timecourse of ERK phosphorylation (Fig. 3). 1668S induced
he most rapid induction of ERK phosphorylation, and addition oferminal dG residues to create Long1668S delayed the signallingy 20 min and substantially reduced the maximal response. How-ver Long1668S was still a markedly better stimulus than AOS,hich gave a lower and further delayed response. Removal of ter-
ig. 3. Addition of terminal dG residues to PS-ODN can cause delayed signalling. BMMs wamples were probed with anti-phospho ERK antibodies and reprobed for total ERK ashows results from a single membrane.
and 1668. BMMs were treated with 1 �M PS-ODN or 3 �M PO-ODN and Westernantibodies, and reprobed for total p38 as a loading control. An irrelevant lane has
minal dG residues from AOS to create LAOS gave more rapid andpotent signalling, but still did not generate an ODN like 1668S,which signalled strongly at 20 min. Together this data shows thatwhilst terminal dG residues can cause delayed signalling, other basechanges 6 or more bases 5′ to the active CpG dinucleotide can alsobe important.
3.5. Other sub-optimal CpG PS-ODN show a delay in signalling
During the early development of CpG ODN, a large range ofdifferent sequences was tested, and we chose two of these previ-ously reported PS-ODN (Yamada et al., 2002) to compare in activityand signalling strength to 1668S. These PS-ODN were synthesisedwith and without terminal G residues (G1466G/1466 and 1555L/S).Both PS-ODN were dramatically less stimulatory for ELAM pro-moter activation than 1668S (Fig. 4) and the addition of terminal dGresidues to the ODN had little effect on their already low stimula-
tory potential. The kinetics of phosphorylation of p38 in response tothe 1466 and 1555 PS-ODN was also compared to phosphorylationinduced by 1668S. 1668S induced phosphorylation of p38 within20 min of addition to cells, whereas signalling for the sub-optimalPS-ODN was not detected until 40 min or longer after additionere treated with 1 �M ODN and Western extracts were made at the times indicated.a loading control. An irrelevant lane has been removed from the top panel which
T.L. Roberts et al. / Molecular Immu
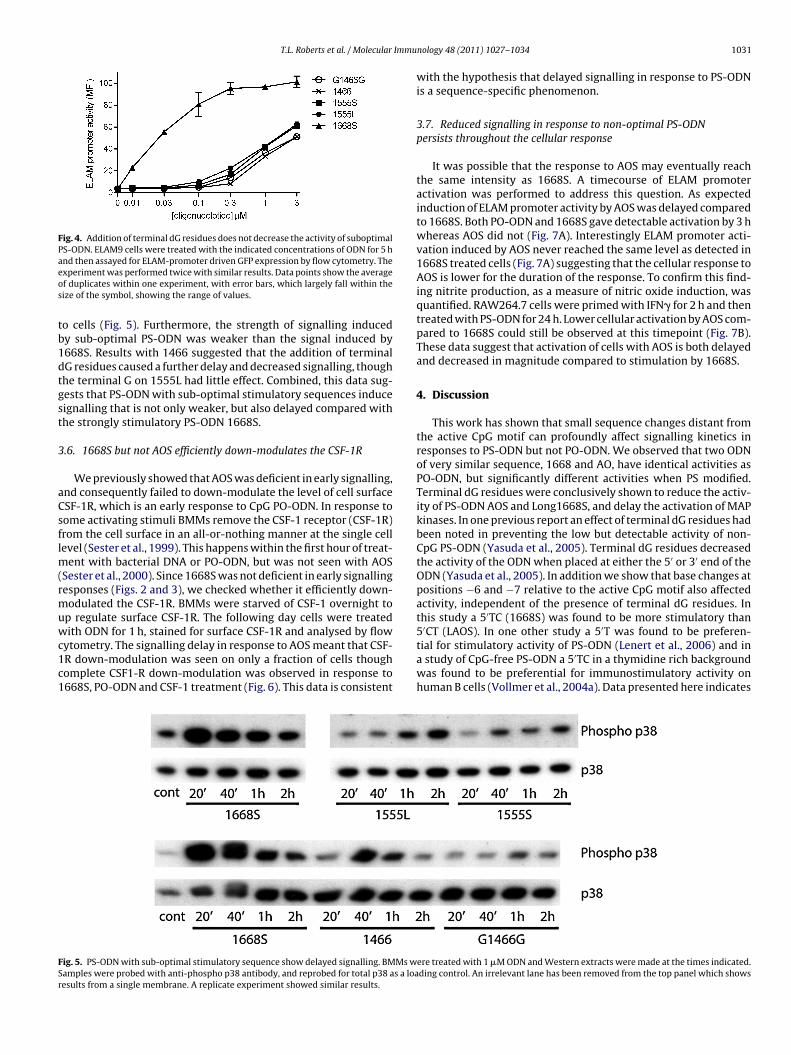
Fig. 4. Addition of terminal dG residues does not decrease the activity of suboptimalPS-ODN. ELAM9 cells were treated with the indicated concentrations of ODN for 5 haeos
tb1dtgst
3
aCsflm(rmuwc1c1
FSr
nd then assayed for ELAM-promoter driven GFP expression by flow cytometry. Thexperiment was performed twice with similar results. Data points show the averagef duplicates within one experiment, with error bars, which largely fall within theize of the symbol, showing the range of values.
o cells (Fig. 5). Furthermore, the strength of signalling inducedy sub-optimal PS-ODN was weaker than the signal induced by668S. Results with 1466 suggested that the addition of terminalG residues caused a further delay and decreased signalling, thoughhe terminal G on 1555L had little effect. Combined, this data sug-ests that PS-ODN with sub-optimal stimulatory sequences induceignalling that is not only weaker, but also delayed compared withhe strongly stimulatory PS-ODN 1668S.
.6. 1668S but not AOS efficiently down-modulates the CSF-1R
We previously showed that AOS was deficient in early signalling,nd consequently failed to down-modulate the level of cell surfaceSF-1R, which is an early response to CpG PO-ODN. In response toome activating stimuli BMMs remove the CSF-1 receptor (CSF-1R)rom the cell surface in an all-or-nothing manner at the single cellevel (Sester et al., 1999). This happens within the first hour of treat-
ent with bacterial DNA or PO-ODN, but was not seen with AOSSester et al., 2000). Since 1668S was not deficient in early signallingesponses (Figs. 2 and 3), we checked whether it efficiently down-odulated the CSF-1R. BMMs were starved of CSF-1 overnight to
p regulate surface CSF-1R. The following day cells were treated
ith ODN for 1 h, stained for surface CSF-1R and analysed by flowytometry. The signalling delay in response to AOS meant that CSF-R down-modulation was seen on only a fraction of cells thoughomplete CSF1-R down-modulation was observed in response to668S, PO-ODN and CSF-1 treatment (Fig. 6). This data is consistent
ig. 5. PS-ODN with sub-optimal stimulatory sequence show delayed signalling. BMMs wamples were probed with anti-phospho p38 antibody, and reprobed for total p38 as a loaesults from a single membrane. A replicate experiment showed similar results.
nology 48 (2011) 1027–1034 1031
with the hypothesis that delayed signalling in response to PS-ODNis a sequence-specific phenomenon.
3.7. Reduced signalling in response to non-optimal PS-ODNpersists throughout the cellular response
It was possible that the response to AOS may eventually reachthe same intensity as 1668S. A timecourse of ELAM promoteractivation was performed to address this question. As expectedinduction of ELAM promoter activity by AOS was delayed comparedto 1668S. Both PO-ODN and 1668S gave detectable activation by 3 hwhereas AOS did not (Fig. 7A). Interestingly ELAM promoter acti-vation induced by AOS never reached the same level as detected in1668S treated cells (Fig. 7A) suggesting that the cellular response toAOS is lower for the duration of the response. To confirm this find-ing nitrite production, as a measure of nitric oxide induction, wasquantified. RAW264.7 cells were primed with IFN� for 2 h and thentreated with PS-ODN for 24 h. Lower cellular activation by AOS com-pared to 1668S could still be observed at this timepoint (Fig. 7B).These data suggest that activation of cells with AOS is both delayedand decreased in magnitude compared to stimulation by 1668S.
4. Discussion
This work has shown that small sequence changes distant fromthe active CpG motif can profoundly affect signalling kinetics inresponses to PS-ODN but not PO-ODN. We observed that two ODNof very similar sequence, 1668 and AO, have identical activities asPO-ODN, but significantly different activities when PS modified.Terminal dG residues were conclusively shown to reduce the activ-ity of PS-ODN AOS and Long1668S, and delay the activation of MAPkinases. In one previous report an effect of terminal dG residues hadbeen noted in preventing the low but detectable activity of non-CpG PS-ODN (Yasuda et al., 2005). Terminal dG residues decreasedthe activity of the ODN when placed at either the 5′ or 3′ end of theODN (Yasuda et al., 2005). In addition we show that base changes atpositions −6 and −7 relative to the active CpG motif also affectedactivity, independent of the presence of terminal dG residues. Inthis study a 5′TC (1668S) was found to be more stimulatory than
5′CT (LAOS). In one other study a 5′T was found to be preferen-tial for stimulatory activity of PS-ODN (Lenert et al., 2006) and ina study of CpG-free PS-ODN a 5′TC in a thymidine rich backgroundwas found to be preferential for immunostimulatory activity onhuman B cells (Vollmer et al., 2004a). Data presented here indicatesere treated with 1 �M ODN and Western extracts were made at the times indicated.ding control. An irrelevant lane has been removed from the top panel which shows

1032 T.L. Roberts et al. / Molecular Immunology 48 (2011) 1027–1034
Fig. 6. AOS is less efficient at inducing down-regulation of surface CSF-1R than 1668S. BMM were starved of CSF-1 overnight and either left untreated (filled purple histogramboth panels) or treated with 10,000 U/ml CSF-1 as a positive control (green line, both pa(light blue, right panel) or 1 �M 1668 (orange, right panel) for 1 h before being harvestedremaining on the cell surface. Data is a representative sample from 4 replicates.
Fig. 7. Delayed signalling in response to AOS results in a long-term reducedresponse. (A) A timecourse of ODN activity was performed using the ELAM pro-moter activity. Cells were treated with ODN and harvested at the indicated times.Points show the average of duplicates and the error bars, which may fall within thesize of the symbol, show the range of the values obtained. Data is representative oftwo experiments. (B) IFN� primed RAW264.7 cells were treated with the indicatedconcentrations of AOS or 1668S for 24 h. Nitrite production was measured usingGriess reagent. In both panels, data points are the average of triplicates and errorbars show the standard deviation. Experiment is representative of two independentassays.
nels), 0.3 �M AOS (dark blue, left panel), 0.3 �M 1668S (red, left panel), 1 �M AOand stained for surface CSF-1R expression. The x-axis shows the amount of CSF-1R
that a 5′TC motif may also increase immunostimulatory activityof a CpG-containing PS-ODN that does not have a thymidine richsequence.
5′ T residues are also present in a number of commonly usedinhibitory oligonucleotides (Trieu et al., 2006). The importance ofindividual residues for inhibitory activity of ODN has also beenexamined by Lenert et al. (Ashman and Goeken, 2005); in thisstudy they found that at the 5′ end the T residue could be changedto any other residue or removed without affecting the inhibitorypotency of the oligonucleotide and that CC was preferable in thenext two positions. It is currently unclear whether inhibitory ODNand stimulatory ODN bind to the same region of TLR9 and furtherwork will be required to determine exact binding sites of each ODNclass.
Sequence requirements for efficient TLR9 activation by PS-ODNhave been previously observed to be different than those for PO-ODN (Krieg, 2002). We have observed that the published speciesspecificity for recognition of different CpG motifs (Bauer et al., 2001;Hartmann and Krieg, 2000) is a PS-ODN-specific phenomenon notseen with natural DNA (Roberts et al., 2005b). In that context,differences in activity of different sequences may be due to PS-ODN-specific conformation at the 6 base pair CpG motif. However, workpresented here and elsewhere (Lenert et al., 2006) suggests thatPS modification may also have more subtle effects, with sequencechanges distant from the CpG motif having an impact. PS modifica-tion of the backbone has known effects on the physical propertiesof DNA. NMR and CD spectroscopy data suggested that the PS mod-ification can slightly increase the flexibility of the ODN backbone(Hartmann et al., 1999). Whether such an effect could alter inter-action of distal bases with TLR9 remains to be established.
Previous work in this laboratory showed that the PS-ODN AOShad delayed MAPK signalling compared to its PO-ODN counter-part AO (Sester et al., 2000; Sweet et al., 2002). Here we showedthat 1668S signals with similar kinetics to its PO version 1668. AOSwas subsequently a poorer stimulus than 1668S in both short andlong term assays of activation (Figs. 1 and 5). Delayed signalling
is therefore not a feature of all PS-ODN, but occurs with PS-ODNthat have sub-optimal stimulatory sequences (Fig. 5). Interestingly,differences in signalling kinetics were also involved in the species-specific responses to “human” and “mouse” CpG motifs in PS-ODN(Roberts et al., 2005b).
Immu
iiw1rfobt(eien
mnsrdcwmasi
C
A
t(tt
R
A
B
B
C
D
d
H
H
H
T.L. Roberts et al. / Molecular
Delayed signalling could result from decreased uptake or lim-ted ability to activate TLR9. We found no detectable differencen uptake of fluorescently labelled 1668S and AOS (Fig. 1B). Thus,
e suggest that the small sequence differences between AOS and668S affect TLR9-mediated recognition and signalling. Sequencesemote from the CpG motif could affect activation of TLR9 in theollowing ways: (i) structure- or sequence-related interaction withther proteins inhibiting binding to TLR9. PS-ODN are known toe “sticky”; interacting with a range of proteins which have lit-le affinity for native DNA (Brown et al., 1994; Zhang et al., 2005);ii) Alteration in the recruitment of TLR9 to late endosomes. Forxample, non-stimulatory methylated CpG ODN did not efficientlynduce TLR9 relocalisation to the endosomal compartment (de Jongt al., 2010) or (iii) Altered affinity for TLR9 in a PS-specific man-er.
In summary small changes to sequences distal from the CpGotif can affect the immunostimulatory activity of PS-ODN but
ot PO-ODN. Particularly detrimental to activity of the stronglytimulatory PS-ODN 1668S was the addition of terminal dGesidues. The observed decrease in activity was associated withelayed MAPK signalling. PS-ODN are in clinical trials as vac-ine adjuvants and cancer therapies (Jurk and Vollmer, 2007)here optimal immunostimulatory activity is required and ter-inal dG residues should be avoided. In other applications such
s antisense therapy, immunostimulation may be undesirable andequence alterations may be used to decrease stimulatory activ-ty.
onflict of interest
No competing financial interests exist.
cknowledgements
This work was supported by Grants 301211 and 455882 fromhe National Health and Medical Research Council of AustraliaNHMRC). TLR is supported by a Peter Doherty Fellowship fromhe NHMRC. KJS is supported by a Future Fellowship from the Aus-ralian Research Council.
eferences
shman, R.F., Goeken, J.A., Drahos, J., Lenert, P., 2005. Sequence requirements foroligodeoxyribonucleotide inhibitory activity. Int. Immunol. 17 (4), 411–420.
auer, S., Kirschning, C.J., Hacker, H., Redecke, V., Hausmann, S., Akira, S., Wagner,H., Lipford, G.B., 2001. Human TLR9 confers responsiveness to bacterial DNAvia species-specific CpG motif recognition. Proc. Natl. Acad. Sci. U.S.A. 98 (16),9237–9242.
rown, D.A., Kang, S.H., Gryaznov, S.M., DeDionisio, L., Heidenreich, O., Sullivan,S., Xu, X., Nerenberg, M.I., 1994. Effect of phosphorothioate modification ofoligodeoxynucleotides on specific protein binding. J. Biol. Chem. 269 (43),26801–26805.
ostelloe, E.O., Stacey, K.J., Antalis, T.M., Hume, D.A., 1999. Regulation of theplasminogen activator inhibitor-2 (PAI-2) gene in murine macrophages. Demon-stration of a novel pattern of responsiveness to bacterial endotoxin. J. LeukocyteBiol. 66 (1), 172–182.
avis, H.L., Weeratna, R., Waldschmidt, T.J., Tygrett, L., Schorr, J., Krieg, A.M., 1998.CpG DNA is a potent enhancer of specific immunity in mice immunized withrecombinant hepatitis B surface antigen. J. Immunol. 160 (2), 870–876.
e Jong, S.D., Basha, G., Wilson, K.D., Kazem, M., Cullis, P., Jefferies, W., Tam, Y.,2010. The immunostimulatory activity of unmethylated and methylated CpGoligodeoxynucleotide is dependent on their ability to colocalize with TLR9 inlate endosomes. J. Immunol. 184 (11), 6092–6102.
acker, H., Vabulas, R.M., Takeuchi, O., Hoshino, K., Akira, S., Wagner, H., 2000.Immune cell activation by bacterial CpG-DNA through myeloid differentiationmarker 88 and tumor necrosis factor receptor-associated factor (TRAF)6. J. Exp.
Med. 192 (4), 595–600.artmann, B., Bertrand, H., Fermandjian, S., 1999. Sequence effects on energetic andstructural properties of phosphorothioate DNA: a molecular modelling study.Nucleic Acids Res. 27 (16), 3342–3347.
artmann, G., Krieg, A.M., 2000. Mechanism and function of a newly identified CpGDNA motif in human primary B cells. J. Immunol. 164 (2), 944–953.
nology 48 (2011) 1027–1034 1033
Hemmi, H., Takeuchi, O., Kawai, T., Kaisho, T., Sato, S., Sanjo, H., Matsumoto, M.,Hoshino, K., Wagner, H., Takeda, K., Akira, S., 2000. A Toll-like receptor recognizesbacterial DNA. Nature 408 (6813), 740–745.
Jurk, M., Vollmer, J., 2007. Therapeutic applications of synthetic CpG oligodeoxynu-cleotides as TLR9 agonists for immune modulation. Biodrugs 21 (6), 387–401.
Krieg, A.M., 2002. CpG motifs in bacterial DNA and their immune effects. Annu. Rev.Immunol. 20, 709–760.
Krieg, A.M., Yi, A.K., Matson, S., Waldschmidt, T.J., Bishop, G.A., Teasdale, R., Koret-zky, G.A., Klinman, D.M., 1995. CpG motifs in bacterial DNA trigger direct B-cellactivation. Nature 374 (6522), 546–549.
Latz, E., Schoenemeyer, A., Visintin, A., Fitzgerald, K.A., Monks, B.G., Knetter, C.F., Lien,E., Nilsen, N.J., Espevik, T., Golenbock, D.T., 2004. TLR9 signals after translocatingfrom the ER to CpG DNA in the lysosome. Nat. Immunol. 5 (2), 190–198.
Latz, E., Verma, A., Visintin, A., Gong, M., Sirois, C.M., Klein, D.C., Monks, B.G., McK-night, C.J., Lamphier, M.S., Duprex, W.P., Espevik, T., Golenbock, D.T., 2007.Ligand-induced conformational changes allosterically activate Toll-like receptor9. Nat. Immunol. 8 (7), 772–779.
Lenert, P., Goeken, A.J., Ashman, R.F., 2006. Extended sequence preferences foroligodeoxyribonucleotide activity. Immunology 117 (4), 474–481.
Manzel, L., Macfarlane, D.E., 1999. Lack of immune stimulation by immobilized CpG-oligodeoxynucleotide. Antisense Nucleic Acid Drug Dev. 9 (5), 459–464.
Park, Y., Lee, S.W., Sung, Y.C., 2002. Cutting Edge: CpG DNA inhibits dendritic cellapoptosis by up-regulating cellular inhibitor of apoptosis proteins through thephosphatidylinositide-3′-OH kinase pathway. J. Immunol. 168 (1), 5–8.
Roberts, T.L., Dunn, J.A., Terry, T.D., Jennings, M.P., Hume, D.A., Sweet, M.J., Stacey,K.J., 2005a. Differences in macrophage activation by bacterial DNA and CpG-containing oligonucleotides. J. Immunol. 175 (6), 3569–3576.
Roberts, T.L., Idris, A., Dunn, J.A., Kelly, G.M., Burnton, C.M., Hodgson, S., Hardy, L.L.,Garceau, V., Sweet, M.J., Ross, I.L., Hume, D.A., Stacey, K.J., 2009. HIN-200 proteinsregulate caspase activation in response to foreign cytoplasmic DNA. Science 323(5917), 1057–1060.
Roberts, T.L., Sweet, M.J., Hume, D.A., Stacey, K.J., 2005b. Cutting edge: species-specific TLR9-mediated recognition of CpG and non-CpG phosphorothioate-modified oligonucleotides. J. Immunol. 174 (2), 605–608.
Rutz, M., Metzger, J., Gellert, T., Luppa, P., Lipford, G.B., Wagner, H., Bauer, S., 2004.Toll-like receptor 9 binds single-stranded CpG-DNA in a sequence- and pH-dependent manner. Eur. J. Immunol. 34 (9), 2541–2550.
Sanjuan, M.A., Rao, N., Lai, K.T., Gu, Y., Sun, S., Fuchs, A., Fung-Leung, W.P., Colonna,M., Karlsson, L., 2006. CpG-induced tyrosine phosphorylation occurs via a TLR9-independent mechanism and is required for cytokine secretion. J. Cell. Biol. 172(7), 1057–1068.
Sester, D.P., Beasley, S.J., Sweet, M.J., Fowles, L.F., Cronau, S.L., Stacey, K.J., Hume, D.A.,1999. Bacterial/CpG DNA down-modulates colony stimulating factor-1 receptorsurface expression on murine bone marrow-derived macrophages with con-comitant growth arrest and factor-independent survival. J. Immunol. 163 (12),6541–6550.
Sester, D.P., Brion, K., Trieu, A., Goodridge, H.S., Roberts, T.L., Dunn, J., Hume,D.A., Stacey, K.J., Sweet, M.J., 2006. CpG DNA activates survival in murinemacrophages through TLR9 and the phosphatidylinositol 3-kinase-Akt pathway.J. Immunol. 177 (7), 4473–4480.
Sester, D.P., Naik, S., Beasley, S.J., Hume, D.A., Stacey, K.J., 2000. Phosphoroth-ioate backbone modification modulates macrophage activation by CpG DNA.J. Immunol. 165 (8), 4165–4173.
Stacey, K.J., Sester, D.P., Sweet, M.J., Hume, D.A., 2000. Macrophage activation byimmunostimulatory DNA. Curr. Top. Microbiol. Immunol. 247, 41–58.
Stacey, K.J., Young, G.R., Clark, F., Sester, D.P., Roberts, T.L., Naik, S., Sweet, M.J., Hume,D.A., 2003. The molecular basis for the lack of immunostimulatory activity ofvertebrate DNA. J. Immunol. 170 (7), 3614–3620.
Sweet, M.J., Campbell, C.C., Sester, D.P., Xu, D., McDonald, R.C., Stacey, K.J., Hume, D.A.,Liew, F.Y., 2002. Colony-stimulating factor-1 suppresses responses to CpG DNAand expression of toll-like receptor 9 but enhances responses to lipopolysac-charide in murine macrophages. J. Immunol. 168 (1), 392–399.
Sweet, M.J., Stacey, K.J., Kakuda, D.K., Markovich, D., Hume, D.A., 1998. IFN-gammaprimes macrophage responses to bacterial DNA. J. Interferon Cytokine Res. 18(4), 263–271.
Trieu, A., Roberts, T.L., Dunn, J.A., Sweet, M.J., Stacey, K.J., 2006. DNA motifs suppress-ing TLR9 responses. Crit. Rev. Immunol. 26 (6), 527–544.
Tsujimura, H., Tamura, T., Kong, H.J., Nishiyama, A., Ishii, K.J., Klinman, D.M., Ozato, K.,2004. Toll-like receptor 9 signaling activates NF-kappaB through IFN regulatoryfactor-8/IFN consensus sequence binding protein in dendritic cells. J. Immunol.172 (11), 6820–6827.
Vollmer, J., Weeratna, R., Payette, P., Jurk, M., Schetter, C., Laucht, M., Wader, T.,Tluk, S., Liu, M., Davis, H.L., Krieg, A.M., 2004a. Characterization of three CpGoligodeoxynucleotide classes with distinct immunostimulatory activities. Eur.J. Immunol. 34 (1), 251–262.
Vollmer, J., Weeratna, R.D., Jurk, M., Samulowitz, U., McCluskie, M.J., Payette, P.,Davis, H.L., Schetter, C., Krieg, A.M., 2004b. Oligodeoxynucleotides lacking CpGdinucleotides mediate Toll-like receptor 9 dependent T helper type 2 biasedimmune stimulation. Immunology 113 (2), 212–223.
Yamada, H., Gursel, I., Takeshita, F., Conover, J., Ishii, K.J., Gursel, M., Takeshita, S.,
Klinman, D.M., 2002. Effect of suppressive DNA on CpG-induced immune acti-vation. J. Immunol. 169 (10), 5590–5594.Yasuda, K., Yu, P., Kirschning, C.J., Schlatter, B., Schmitz, F., Heit, A., Bauer, S.,Hochrein, H., Wagner, H., 2005. Endosomal translocation of vertebrate DNAactivates dendritic cells via TLR9-dependent and -independent pathways. J.Immunol. 174 (10), 6129–6136.

1 Immu
Y
Y
034 T.L. Roberts et al. / Molecular
i, A.K., Chace, J.H., Cowdery, J.S., Krieg, A.M., 1996. IFN-gamma promotes IL-6 andIgM secretion in response to CpG motifs in bacterial DNA and oligodeoxynu-cleotides. J. Immunol. 156 (2), 558–564.
i, A.K., Krieg, A.M., 1998. CpG DNA rescue from anti-IgM-induced WEHI-231 Blymphoma apoptosis via modulation of I kappa B alpha and I kappa B beta
nology 48 (2011) 1027–1034
and sustained activation of nuclear factor-kappa B/c-Rel. J. Immunol. 160 (3),1240–1245.
Zhang, Z., Weinschenk, T., Schluesener, H.J., 2005. Uptake, cellular distributionand novel cellular binding proteins of immunostimulatory CpG oligodeoxynu-cleotides in glioblastoma cells. Mol. Cell. Biochem. 272 (1–2), 35–46.




















