
Glucocorticoids induce mitochondrial gene transcription in HepG2 cells

Cell Biology International 2001, Vol. 25, No. 8, 835–840doi:10.1006/cbir.2001.0765, available online at http://www.idealibrary.com on
GP130-SPECIFIC ANTISENSE OLIGONUCLEOTIDES INHIBIT IL-6 SIGNALINDUCING JUNB MRNA TRANSCRIPTION IN THE HUMAN HEPATOMA CELL
LINE, HEPG2
VALEuRIA LIA VARGA1, ANDRAu S KRISTOu F FU} LO} P1, MARIANNA CSILLA HOLUB1, SAu RA TOu TH1,CSABA SZALAI2 and ANDRAu S FALUS1,2*
1Department of Genetics, Cell and Immunobiology, Semmelweis University, Budapest2Molecular Immunology Research Group, National Academy of Sciences, Budapest, Hungary
Received 6 December 2000; accepted 29 March 2001
The biosynthesis of interleukin-6 receptor (IL-6R) and gp130 in vitro was blocked using specificantisense oligonucleotides (ASO) in HepG2 liver cells and the efficacy of various ASOs wastested on the generation of IL-6-induced junB mRNA. We used three ASOs specific for the IL-6receptor, three specific for gp130 and a control (nonsense) oligonucleotide specific for �-chain ofIgE (not expressing in HepG2 cells). Our data indicate that a gp130-specific ASO, g2, was themost effective blocker of IL-6-induced junB mRNA, whilst the IL-6 receptor ASOs alone wereineffective. The mechanism of gene inactivation by ASO treatment was partially elucidated bydemonstration of the loss of gp130 mRNA from cells treated with ASOs showing functionalefficacy. Our data may help to design antisense oligonucleotides that are effective in therapy(e.g. as anti-inflammatory agents) in the future. � 2001 Academic Press
K: antisense; IL-6; IL-6 receptor; gp130; junB.
*To whom correspondence should be addressed: Professor AndrasFalus, Department of Genetics, Cell and Immunology, SemmelweisUniversity, Budapest, Nagyvarad ter 4, H-1089, Budapest, Hungary.Tel.: (36) 1 210 2929; Fax: (36) 1 303 6968; E-mail: [email protected]
INTRODUCTION
Pleiotropy and redundancy are characteristicfeatures of most cytokines, including IL-6, IL-6promotes the growth of myeloma/plasmacytomacells, T cells, and renal mesangial cells. It inhibitsthe growth of myeloid leukemic cell lines andcertain carcinoma cell lines, and induces differen-tiation or tissue-specific gene expression, includingneural differentiation of P12 cells, megakaryocytematuration, immunoglobulin production in B cells,and acute-phase protein synthesis (Taga, 1997).One of the first effects of IL-6 in hepatocytes is theinduction of junB transcription factor gene expres-sion (Igaz et al., 1998). The IL-6 signal is trans-duced through a homodimer of membraneglycoprotein gp130, which upon its interaction withIL-6, associates with a ligand specific receptor,IL-6R (Taga et al., 1989; Hibi et al., 1990).
1065–6995/01/080835+06 $35.00/0
ASOs are short, traditionally 15–25 baseslong, single-stranded DNA fragments, which aretargeted against a specific mRNA. To be effectiveantisense agents, ASOs, in addition to their selec-tivity, must fulfil other requirements, includingresistance to nucleases (before binding to targetmRNA), low toxicity and sufficient penetration(Oberbauer, 1997; Wagner, 1995; Varga et al.,1999). The oligonucleotides have to be modified inorder to increase their stability. Several modifiedversions of oligonucleotide have been synthesised.Phosphorothioate oligonucleotides (S-oligos) arethe most widely used analogues for antisense appli-cation. S-oligos have a sulphur for oxygen substi-tution of one of the non-bridged oxygen atoms ofinternucleotide linkages. S-oligonucleotides arenegatively charged, taken up by endocytosis, acti-vate RNaseH and also have a strong interactionwith plasma proteins (Varga et al., 1999). RNaseH,a ubiquitous enzyme, recognizes the RNA:DNAheteroduplex and cleaves the RNA at that site bydestroying the target gene transcript (Cerritelliet al., 1998). By this exogenous antisense DNA
� 2001 Academic Press

836 Cell Biology International, Vol. 25, No. 8, 2001
treatment and targeted blocking of translation, wehave a tool to transiently ‘knock out’ a specificgene and to study the influence or potential func-tion of certain genes in the cell. In our studies, weused three ASOs against the gp130 (g1, g2, g3)cDNA and three ASOs against the IL-6R (r1, r2,r3) cDNA. For controlling non-specific effects(e.g. negative charge) we used a ‘nonsense’ ASOof similar size specific to the heavy chain of IgE.All the antisense oligonucleotides were 18-merS-oligos. We tested the vulnerability of a sensitiveand immediate function of IL-6, induction of junBmRNA. Moreover, the mechanism of the action ofspecific ASOs was followed by directly measuringthe amount of target mRNA (gp130). Our dataindicate that gp130 specific ASOs (particularly theone which is complementary to nucleotide bases173–190 on gp130 mRNA) significantly suppressIL-6 function in HepG2 cells, in vitro.
MATERIALS AND METHODS
Cell culture and treatment
HepG2, a human hepatoma cell line, was culturedin RPMI 1640 medium containing 10% FCS(Gibco), supplemented with 2 m glutamine and160 �g/ml gentamycin in six-well plates (2�106
cells/ml). During the treatment, cells were kept inserum-free medium for 48 h, at 37�C in an atmos-phere containing 5% CO2. Cells were incubatedfirst with ASOs (r1, r2, r3, g1, g2, g3, nonsense IgE)for 24 h and then induced with IL-6 for a further24 h. ASO concentration was 20 �g/ml in each case,and 10 U/ml of IL-6 was used as this had beenfound to be optimal in previous studies (notshown). The cells were harvested for RNA prep-aration. They were certified at source, used at lowpassage number and tested free of mycoplasma.
Antisense sequences
Phosphorothioated 18-mer antisense oligodeoxy-nucleotides were used, three of them specific forIL-6R mRNA (r1 3� hca ca tgc gct cct cch 5�, r2 3�hhg ata agt acg atg gcc 5�, r3 3� hgt aag gac caa cgacch 5�) corresponding to IL-6R cDNA sequences31–48, 328–345 and 1147–1164, respectively (Baueret al., 1989). Three ASOs were specific for gp130mRNA (g1 3� hcc ggc tgc cca gac cgg 5�, g2 3� hactgt aaa tct tca tch 5�, g3 3� hha cgg gaa cac ttc caa5�) corresponding to gp130 cDNA sequences 71–88, 173–190 and 859–876, respectively (Hibi et al.,1990). ASO r3 corresponds to the transmembrane
region of the IL-6 receptor, whilst all others arecomplementary to extracellular parts of IL-6R andgp130. For controlling the specificity of ASOs a18-mer phosphorothioated control antisense oligo-nucleotide (‘nonsense’) was used which is specificfor the immunoglobulin � chain gene (Fekete et al.,1997). Uptake and catabolism of ASOs in HepG2cells used in this experiment were controlled inpreliminary experiments (not shown).
Oligonucleotide synthesis
These antisense oligonucleotides were synthesisedby automated DNA synthesis using standardphosphoramidite chemistry in a type 8700 DNAsynthesiser (Millipore, U.S.A.) on 1 �mol scales.Purification was carried out by a reversed-phaseHPLC (Jasco) method using a Shandon HypersilODS column (5 �m, 8�250 mm). The main peakwas dialysed in a Spectra/Por-3 dialysis bag (mol-ecular weight cut-off 3500), freeze-dried and storedat �25�C, as described elsewhere (Wagner, 1997).All oligonucleotides were synthesised as phospho-rothiotates containing base modifications. In orderto obtain biologically highly stable oligonucleo-tides, the thymines were replaced by 5-(n-1-hexynil)uracils. Purification and desalting was car-ried out as described above (Wagner, 1997;Denhardt, 1992). Purity was checked by HPLC andconcentrations were calculated on the basis of 1OD260 unit=40 �g oligonucleotide potassium salt.
RNA isolation
Total cellular RNA was isolated according to themethod of Chomczynski and Sacchi (1987) usingguanidium thiocyanate-containing lysis buffer. TheRNA pellet was dissolved in sterile DEPC water.RNA concentration was measured by a BeckmanDU-6 photometer. RNAs were stored at �80�Cuntil use.
Reverse transcription (RT) and PCR for junB,glycerol-3-phosphate-dehydrogenase (G3PDH) andgp130
RT was carried out using a Perkin Elmer RT-PCRkit on a Pharmacia Ataq Gene Controller. MuLVreverse transcriptase and oligo-dT primers wereused in the reaction. The reaction mixture (total20 �l) was made up of the following: Mg2+ (25 m)4 �l, 10�buffer 2 �l, dNTPs (4�10 m) 2 �l, RN-ase inhibitor 1 �l, oligo-dT primer 1 �l MuLVreverse transcriptase 0.5 �l, DEPC-H2O 2.5 �l.RNA (1 �g) was added to each reaction mixture.

Cell Biology International, Vol. 25, No. 8, 2001 837
The protocol used was the one given by themanufacturer (samples were incubated at 42�C for20 min and 99�C for 5 min). cDNAs were stored at�80�C until use.
The PCR reaction of junB (Igaz et al., 1998b)was carried out using the following programme onPharmacia Ataq Gene Controller: 94�C for 4 min,95�C for 30 s, 57�C for 45 s, 73�C for 45 s�35,72�C for 5 min. The reaction mixture (total 50 �l)contained 10�buffer (Promega) 5 �l, Mg2+
(25 m) (Promega) 3 �l, dNTPs (4�10 m)(Perkin Elmer) 4 �l, primers (50 pmol/�lsense+antisense) 1 �l, H2O 35 �l, Taq polymerase(Promega) 0.2 �l, sample (cDNA) 2 �l.
The junB primers were synthesised by Pharma-cia. They were junB-specific primers (Smetsers etal., 1996) located in the 3� non-coding region of themRNA. The PCR product was 257 bp long. Thesequences of the primers was:
sense: CCAGTCCTTCCACCTCGACGTTTACAAG;
antisense: GACTAAGTGCGTGTTTTCTTTTCCACAGTAC.
The PCR reaction of G3PDH and gp130 wascarried out using the following programme onPharmacia Ataq Gene Controller: 94�C for 4 min,95�C for 30 s, 55�C for 1 min, 72�C for 1 min�32,72�C for 5 min. The reaction mixture (total 50 �l)contained 10�buffer (Promega) 5 �l, Mg2+
(25 m) (Promega) 3 �l, dNTPs (4�10 m) (Per-kin Elmer) 4 �l, primers (50 pmol/�l sense+antisense) 1 �l, H2O 35 �l, Taq polymerase(Promega) 0.2 �l, sample (cDNA) 2 �l.
The G3PDH primers were synthesised by Phar-macia. The PCR product was 452 bp long. Thesequences of the primers were:
sense: ACCACAGTCCATGCCCATCAC;
antisense: TCCACCACCCTGTTGCTGTA.
The gp130 (Hibi et al., 1990) primers were synthe-sised by Pharmacia. The PCR product was 579 bplong. The sequences of the primers were:
sense: CTC GTG TGG AAG ACA TTG CCTCCT T;
antisense: TTG GTC CCA CTC TAA GAC AGCTTC G.
PCR products were run on 1% agarose gelsin 1�TBE buffer containing 0.05% ethidiumbromide. Samples were run at 100 V for 45 min.
Quantification using ImageQuant
G3PDH was used as an internal control. Data wereexpressed as a proportion of junB/G3PDH ana-lysed by the MD ImageQuant Software Version3.3.
Statistical analysis was performed usingStudent’s t-test.
RESULTS AND DISCUSSION
In order to test the efficacy of IL-6R- and gp130-specific ASOs, we tested junB oncogene expressionin HepG2 human hepatoma cells induced by IL-6.Figure 1A shows that in this system optimal upregu-lation of junB transcription was found with 10 U/ml IL-6, therefore we used this concentration forfurther experiments with the ASOs. As a nonsenseASO control, an �-chain (IgE) specific ASO wasused, but had only a slight effect. We tested IL-6R(r1–r3) and gp130 (g1–g3) specific ASOs both aloneand in various combinations. By demonstratingonly some of the combinations in Figure 1A, thejunB RT-PCR amplicons are shown, while inFigure 1B the normalised junB cDNA intensityvalues (divided by the intensity of the correspond-ing glycerol-3-phosphate dehydrogenase values)are presented. These results suggest that the gp130-related g2 ASO strongly inhibits IL-6-induced junBproduction. However, g3 alone is less effective,whilst g1 is totally without effect. Neither g1 nor g3can further increase the blocking effect of g2. TheIL-6 receptor specific ASOs (r1, r2 and r3), how-ever, were all ineffective alone (not shown) andwhen combined (r1+r2+r3), the inhibitory poten-tial does not reach that of g2 or any mixturescontaining g2. The strongest inhibition was foundwhen g2+r1 ASOs were added together, but theeffects did not significantly exceed that of g2 alone.If the inhibition values are deducted from thatobserved in the presence of �-specific ASO, thespecific inhibitory potentials can be calculated(Table 1). The relatively high ‘non-specific’ effect ofIgE specific ASO is probably due to the fact that inthis study (to increase resistance of ASOs toRNases during the 48 h incubation) we used ASOsmodified not only at the 3� ends, but along thewhole sequence, resulting in highly negativelycharged ASO molecules. The specific values (de-ducted from nonsense ASO) ranged from 13–46%.The mechanism of inactivation of translation byASOs is still unclear, however the role of RNaseHin rapid degradation of ASO-mRNA heterodimers(heteroduplexes) is strongly suspected. To obtain
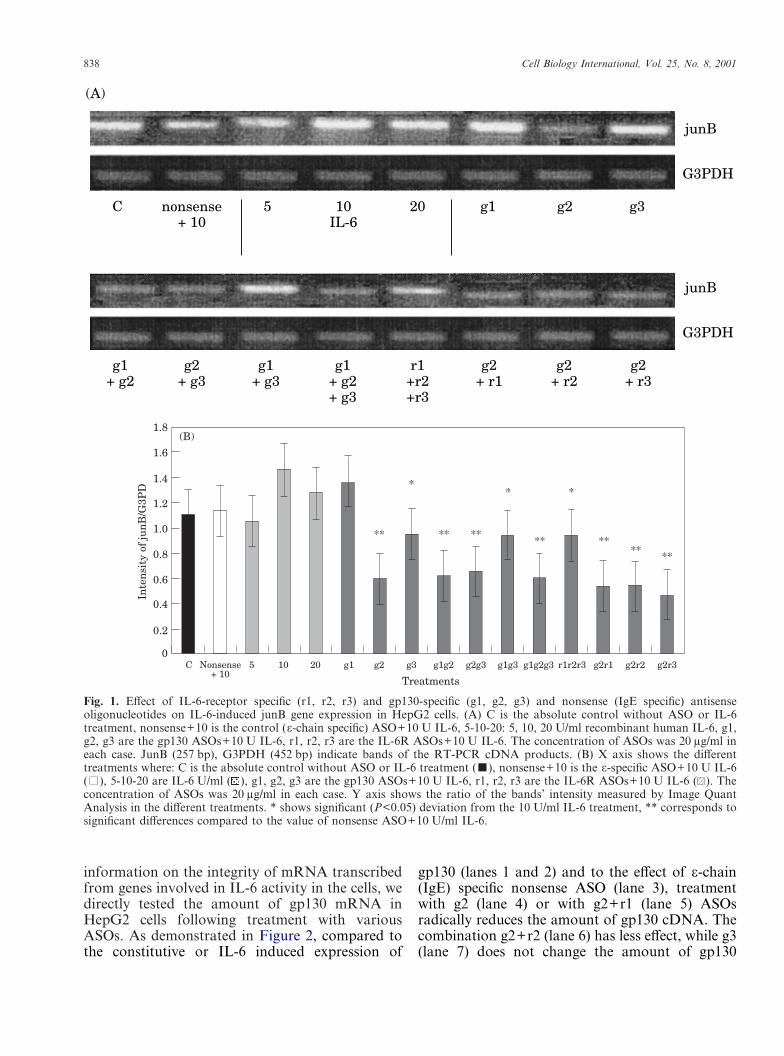
838 Cell Biology International, Vol. 25, No. 8, 2001
0
1.8
Nonsense+ 10
(B)
Inte
nsi
ty o
f ju
nB
/G3P
D
C
1.6
1.4
1.2
1.0
0.8
0.6
0.4
0.2
5 10 20 g1 g2 g3 g1g2 g2g3 g1g3 g1g2g3 r1r2r3 g2r1 g2r2 g2r3
Treatments
** ** **** **
** **
***
Fig. 1. Effect of IL-6-receptor specific (r1, r2, r3) and gp130-specific (g1, g2, g3) and nonsense (IgE specific) antisenseoligonucleotides on IL-6-induced junB gene expression in HepG2 cells. (A) C is the absolute control without ASO or IL-6treatment, nonsense+10 is the control (�-chain specific) ASO+10 U IL-6, 5-10-20: 5, 10, 20 U/ml recombinant human IL-6, g1,g2, g3 are the gp130 ASOs+10 U IL-6, r1, r2, r3 are the IL-6R ASOs+10 U IL-6. The concentration of ASOs was 20 �g/ml ineach case. JunB (257 bp), G3PDH (452 bp) indicate bands of the RT-PCR cDNA products. (B) X axis shows the differenttreatments where: C is the absolute control without ASO or IL-6 treatment (�), nonsense+10 is the �-specific ASO+10 U IL-6(�), 5-10-20 are IL-6 U/ml ( ), g1, g2, g3 are the gp130 ASOs+10 U IL-6, r1, r2, r3 are the IL-6R ASOs+10 U IL-6 ( ). Theconcentration of ASOs was 20 �g/ml in each case. Y axis shows the ratio of the bands’ intensity measured by Image QuantAnalysis in the different treatments. * shows significant (P<0.05) deviation from the 10 U/ml IL-6 treatment, ** corresponds tosignificant differences compared to the value of nonsense ASO+10 U/ml IL-6.
information on the integrity of mRNA transcribedfrom genes involved in IL-6 activity in the cells, wedirectly tested the amount of gp130 mRNA inHepG2 cells following treatment with variousASOs. As demonstrated in Figure 2, compared tothe constitutive or IL-6 induced expression of
gp130 (lanes 1 and 2) and to the effect of �-chain(IgE) specific nonsense ASO (lane 3), treatmentwith g2 (lane 4) or with g2+r1 (lane 5) ASOsradically reduces the amount of gp130 cDNA. Thecombination g2+r2 (lane 6) has less effect, while g3(lane 7) does not change the amount of gp130

Cell Biology International, Vol. 25, No. 8, 2001 839
Table 1.Inhibition of IL-6 induced junB mRNA by IL-6 receptor
and gp130 specific antisense oligonucleotides
Cells treated with10 U/ml IL-6+
Totalinhibition
%*
Specificinhibition
%**
0 — —Nonsense (�) 21.8�1.4 —g1 6.1�1.2 —g2 58.8�5.5 37.0g3 34.3�4.0 12.5g1+g2 57�6.1 35.2g2+g3 54.7�5.9 32.9g1+g3 34.7�4.8 12.9g1+g2+g3 58.4�2.1 36.6r1+r2+r3 34.9�3.9 13.1g2+r1 63.3�3.9 41.5g2+r2 63.3�5.7 41.5g2+r3 67.7�5.5 45.9
Nonsense, � chain (IgE) specific antisense oligonucleotide;n=4; *mean�S.E.M.g1, g2, g3: gp130-specific antisense oligonucleotides (20 �g/ml).r1, r2, r3: IL-6 receptor-specific antisense oligonucleotides.**Reduced by value of nonsense oligonucleotide.
�
��������
� � � �Fig. 2. Gp130 cDNA after ASO treatments in HepG2 cells tested by RT-PCR. Lanes 1: control, 2: 10 U/ml IL-6, 3: 10 U/mlIL-6+nonsense (�) ASO, 4: g2, 5: g2+r1, 6: g2+r2, 7: g3. The concentration of ASOs was 20 �g/ml in each case.
cDNA. Most data showed that the ability of dif-ferent ASOs to enhance RNaseH-related degra-dation of gp130 mRNA corresponded to theirfunctional capability to impair the function of IL-6.However in some experiments (e.g. g2+r1 andg2+r2 resulted in similar inhibitions, see Fig. 1A)the functional effectiveness of ASOs did not corre-late proportionally with their ability to decreasethe gp130 mRNA. These findings suggest that theinactivation of ASO-mRNA heteroduplexes byRNaseH is not the only mechanism for thefunctional effects of ASOs.
Our results indicate that the different antisenseoligonucleotides have entirely different abilities todecrease the availability of IL-6 receptor/gp130 com-plex, that is, to interfere with the effect of IL-6. Theoptimal treatment protocol was found to be 24 h inthe presence of antisense oligonucleotides followed
by 24 h of IL-6 induction. We showed that theantisense oligonucleotides acted differently inHepG2, some (e.g. g2) were more effective thanothers (e.g. g1). The inhibition of IL-6 induction wasfound to be more effective using gp130 ASOs thanusing IL-6R ASOs, which is why we used the IL-6RASOs only in combination with gp130 ASOs. Theparticularly high effectiveness of gp130-specific ASOg2 deserves special attention. The ASO g2 binds toposition 173–190 on gp130 cDNA, which corre-sponds to exon 3. Location of complementary se-quences on gp130 mRNA for ASO g1 is also ratherclose (position 71–88), however this ASO is func-tionally completely inactive. ASO g3, correspondingto nucleotides 859–876 located in the central part ofgp130 mRNA, provides a positive, but smaller ef-fect. The effects of individual gp130 ASOs are notadditive. This is opposite to the results obtainedfrom r1, r2 and r3 IL-6R ASOs, where inactivationwas found only when IL-6R mRNA bound all 3ASOs simultaneously.
The reason why the generation of heteroduplex(ASO-mRNA) at this location on gp130 mRNAmakes the gp130 message so vulnerable (g2) or soresistant (g1) requires further stereochemical andmRNA half-life studies. Contribution of sequenceand phosphorothioate content may both influencethe actual effectiveness of ASOs (Smetsers et al.,1996; Chavany et al., 1995) and even small nucleo-tide changes may completely abolish the functionaleffect of specific ASOs (Fukuda et al., 1997;Branch, 1996; Broaddus et al., 1997).
In our earlier studies, when IL-6 receptor-relatedASOs were designed as overlapping sequencesaround the translational start codon ATG, as littleas one nucleotide shift resulted in more than 50%difference in effect on proliferation of BAF cells(e.g. GGCTACCACTCCATGGTC was inactive,while CGGCTACCACTCCATGGT caused al-most 60% inhibition) (Varga et al., 2000). In thefuture we plan to make stable antisense oligo-nucleotide transfectants (McCall, 1998; de Langeet al., 1993) of HepG2 cells which cannot expressIL-6R or gp130.

840 Cell Biology International, Vol. 25, No. 8, 2001
These studies should help to establish successfulstrategies for prediction and design of antisenseoligonucleotides in human gene therapy.
ACKNOWLEDGEMENTS
This work was supported by grants OTKA T022065, T 003022, T 021175, FKFP090, ETT05123.
REFERENCES
B J, B TM, K T, T T, L G, H T,K T, A G, M L, G W, 1989. Regu-lation of interleukin 6 receptor expression in human mono-cytes and monocyte-derived macrophages. Comparison withthe expression in human hepatocytes. J Exp Med 170:1537–1549.
B AD, 1996. A hitchhiker’s guide to antisense andnonantisense biochemical pathways. Hepatology 24: 1517–1529.
B WC, C ZJ, P SS, L WG, GGT, P LL, F H, 1997. Antiproliferative effectof c-myc antisense phosphorothioate oligodeoxynucleotidesin malignant glioma cells. Neurosurgery 41: 908–915.
C SM, C RJ, 1998. Cloning, expression, andmapping of ribonucleases H of human and mouse related tobacterial RNase HI. Genomics 53: 300–307.
C C, C Y, N L, 1995. Contribution ofsequence and phosphorothioate content to inhibition of cellgrowth and adhesion caused by c-myc antisense oligomers.Mol Pharmacol 48: 738–746.
C P, S N, 1987. Single step method of RNAisolation by acid guanidium thiocyanate-phenol-chloroformextraction. Anal Biochem 162: 156–163.
L P, B GJ, M JN, K JM, 1993.Conditional inhibition of beta-glucuronidase expression byantisense gene fragments in petunia protoplasts. Plant MolBiol 23: 45–55.
D DT, 1992. Mechanism of action of antisense RNA.Sometime inhibition of transcription, processing, transportor translation. In: Bfaserga R, Denhardt DT, eds. AntisenseStrategies. Annals of the New York Academy of Sciences,660: 70–76.
F B, S J, S A, K L, P K, VVL, T K, F A, 1997. Inhibition of IgE produc-
tion by epsilon (e) chain-specific antisense oligonucleotides(aos) studied on human myeloma cell line U266 and periph-eral blood mononuclear cells of a patient with hypereo-sinophilia. Immunol Let 58: 181–190.
F N, K A, W Y, N T, S M,I Y, K K, 1997. Antisense oligodeoxynucle-otide complementary to platelet-derived growth factorA-chain messenger RNA inhibits the arterial proliferation inspontaneously hypertensive rats without altering their bloodpressures. J Hypertens 15: 1123–1136.
H M, M M, S M, H T, T T,K T, 1990. Molecular cloning and expression of anIL-6 signal transducer, gp130. Cell 63: 1149–1157.
I P, T S, R-J S, M I, F G, SC, F A, 1998a. Soluble interleukin-6 (IL-6) receptorinfluences the expression of the protooncogene junB andthe production of fibrinogen in the HepG2 humanhepatoma cell line and primary rat hepatocytes. Cytokine 10:620–626.
I P, F G, S C, T S, F A, 1998b.Development of competitive mRNA PCR for the quantifi-cation of interleukin-6 responsive junB oncogene expression.Biotechniques 24: 854–860.
O R, 1997. Not nonsense but antisense—applicationsof antisense oligonucleotides in different fields of medicine.Wien Klin Wochenschr 109: 40–46.
T T, H M, H Y, Y K, Y K,M T, H T, K T, 1989. Interleukin-6triggers the association of its receptor with a possible signaltransducer, gp130. Cell 58: 573–581.
T T, 1997a. gp130 and the interleukin-6 family ofcytokines. Ann Rev Immun 15: 797–819.
S TF, B JB, M EJ, 1996. Bias innucleotide composition of antisense oligonucleotides. Anti-sense Nucleic Acid Drug Dev 6: 63–67.
V VL, F A, P E, F AK, T S,R-J S, in press. A novel and rapid assay for predic-tion of IL-6 receptor antisense oligonucleotides. Cell BiolIntern 2000.
MC MN, 1998. Repressible antisense inhibition in Blymphocytes. Biochim Biophys Acta 1397: 65–72.
V VL, T S, N I, F A, 1999. Antisensestrategies: functions and application in immunology, areview. Immun Let 69: 217–224.
W RW, 1995. The state of the art in antisense research.Nature Medicine 1: 1116–1118.
W RW, 1997. Antisense technology and prospects fortherapy of viral infections and cancer. Mol Med Today 3:31–38.
Copyright © 2022 FDOKUMEN




















