
Examine the Relationship between Blood Groups and Intercity ...

ORIGINAL ARTICLE
Validation of the dorsal air pouch model to predict andexamine immunostimulatory responses in the gutA. Kourelis1, I. Zinonos1, M. Kakagianni1, A. Christidou1, N. Christoglou1, E. Yiannaki2, T. Testa1,C. Kotzamanidis1, E. Litopoulou-Tzanetaki3, N. Tzanetakis3 and M. Yiangou1
1 Department of Genetics, Development and Molecular Biology, Biology School, Aristotle University of Thessaloniki, Thessaloniki, Greece
2 Haematological Laboratory, Anticancer Hospital ‘‘Theagenio’’, Thessaloniki, Greece
3 Laboratory of Food Microbiology and Hygiene, Faculty of Agriculture, Aristotle University of Thessaloniki, Thessaloniki, Greece
Introduction
Several nonpathogenic bacteria are given as probiotics
or as live bacterial vaccines or as gene delivery vectors
(Steidler et al. 1998; Steidler 2003; Braat et al. 2006).
Given the recent concerns on probiotic treatment, it is
important to investigate their local and systemic
immune activities (Vogel 2008). Probiotics are defined
as ‘live micro-organisms that when being administered
in adequate amounts confer a health benefit on the
host’ (FAO ⁄ WHO 2002). The health benefits of probio-
tics are achieved by bacterial antagonism and immuno-
modulation, which help the healthy host to maintain a
‘physiological state of inflammation’ in the intestine
(Galdeano et al. 2007) or to control several infectious,
inflammatory and immunologic reactions. The pene-
tration of probiotics or their antigens in the intestine
activates the innate immunity through interaction with
the Toll-like receptors (TLRs) present on immune cells
(Akira et al. 2001) and subsequently the production of
cytokines and chemokines (Vinderola et al. 2005).
Under physiological conditions, probiotic bacteria can
act also as mucosal and systemic adjuvants (Galdeano
et al. 2007).
Keywords
air pouch, cytokines, gut mucosa,
Lactobacillus paracasei, probiotic, toll-like
receptors.
Correspondence
Minas Yiangou, Department of Genetics,
Development and Molecular Biology, Biology
School, Aristotle University of Thessaloniki,
54124 Thessaloniki, Greece.
E-mail: [email protected]
2009 ⁄ 0023: received 6 January 2009, revised
7 May 2009 and accepted 21 May 2009
doi:10.1111/j.1365-2672.2009.04421.x
Abstract
Aims: To validate the use of the air pouch system to predict and examine early
immune responses induced by the presumptive probiotics Lactobacillus paraca-
sei subsp. paracasei B112, DC205, DC215 and DC412 strains in the gut
mucosa.
Methods and Results: Only the DC412 strain interacted strongly with the cells
forming the air pouch lining tissue and induced early innate immune responses
such as polymorphonuclear (PMN) cell recruitment, phagocytosis and tumour
necrosis factor alpha (TNF-a) production that equal the respective responses
induced by the probiotic Lactobacillus acidophilus NCFB 1748. The strains
exhibiting strong immunoregulatory activity in the air pouch also interacted
strongly with the gut-associated lymphoid tissue (GALT). The strain DC412
exerts its effect on the intestine through stimulation of Toll-like receptor
(TLR)2 ⁄ TLR4-mediated signalling events leading to secretion of a certain pro-
file of cytokines in which gamma interferon (IFN-c), TNF-a, interleukin (IL)-6
and IL-10 are included. The probiotic Lact. acidophilus NCFB 1748 induces the
same cytokine profile in addition to IL-12B, and this response is potentially
mediated by the synergy of TLR2 and TLR9.
Conclusion: The strain DC412 possesses the in vitro and in vivo characteristics
of a probiotic micro-organism.
Significance and Impact of the Study: The dorsal mouse or rat air pouch may
be used as an alternative and rapid method for the initial discrimination and
selection of potential probiotic Lactobacillus strains.
Journal of Applied Microbiology ISSN 1364-5072
274 Journal compilation ª 2009 The Society for Applied Microbiology, Journal of Applied Microbiology 108 (2010) 274–284
ª 2009 The Authors

Mucosal and systemic administration of probiotics
promotes IgA antibody production (Vitini et al. 2000;
Alvarez-Olmos and Oberhelman 2001), and the induction
of a certain cytokine pattern profile that in turn regulates
the helper T cell 1 (Th1) ⁄ helper T cell 2 (Th2) balance
(Galdeano et al. 2007). The positive influence of many
probiotics on the phagocytic capacity of monocytes and
polymorphonuclear (PMN) cells has been recently
reviewed (Delcenserie et al. 2008). Macrophages and
PMN represent the first line of host defense against
pathogens, and probiotic-mediated local increase in PMN
recruitment successfully treated bovine mastitis (Crispie
et al. 2008). The route of probiotic delivery and the sub-
sequent PMN recruitment and phagocytic activity as well
as chemokine and cytokine production at the local site of
administration are early events and important parameters
for the initiation of non-well-defined immune responses.
In previous studies, we isolated several Lactobacillus
paracasei subsp. paracasei strains from infant’s gastroin-
testinal tract (faeces) or feta cheese prine (Tzanetakis and
Litopoulou-Tzanetaki 1992; Xanthopoulos et al. 2000).
These strains possessed biotechnologically important
characteristics (Litopoulou-Tzanetaki and Tzanetakis,
1992) as well as in vitro probiotic properties such as
viability at low pH and in human gastric juice, bile salt
tolerance, antibacterial activity and cholesterol reduction
(Xanthopoulos et al. 1998, 2000). These observations
provide us with the biological system to study the mech-
anism of the early local action of Lactobacillus strains
possessing in vitro probiotic properties on immune
responses. The 6-day-old epithelium-enclosed air pouch
is developed by epithelial, phagocytic and fibroblast-like
lining cells and comprises the intima (epithelial and poly-
morphonuclear cells) and the subintima (macrophages
and fibroblasts) layers, with an abundance of cells fol-
lowed by an organized vasculature that acts as a mechan-
ical barrier (Sedgwick et al. 1985; Coates and McColl
2001). Activation of the air pouch by antigen causes the
rapid recruitment of PMN cells, while the immune
response products such as cytokines and chemokines are
retained in the air pouch, enhancing early inflammatory
responses not easily detectable in systemic immunity.
Bacterial administration in the air pouch involves their
interaction with the barrier-forming resident macrophag-
es and epithelial cells (Yam et al. 2008). Importantly,
probiotics or their antigens interact with the intestine
barrier through the epithelia of gut mucosa and dendritic
cells and macrophages of villi and Peyer’s patches (Perdi-
gon et al. 2001; Dogi and Perdigon 2006). The similarity
of bacterial activation between the intestine and the air
pouch barriers indicates that the dorsal air pouch may
provide an excellent model to discriminate presump-
tive probiotic strains for their immunostimulatory activ-
ity by examination of their early local effects on immune
responses such as cell-to-cell interactions, chemotaxis and
phagocytic activity. We present data showing that the
Lact. paracasei DC412 strain as well as the Lactobacillus
acidophilus NCFB 1748 probiotic strain exhibit immuno-
regulatory activity in both the dorsal air pouch and the
gut mucosa.
Materials and methods
Bacterial strains and growth conditions
The Lact. paracasei subsp. paracasei B112, DC205, DC215
(Tzanetakis and Litopoulou-Tzanetaki 1992) and DC412
(Xanthopoulos et al. 2000) strains were obtained from the
collection of the Laboratory of Food Microbiology and
Hygiene, Aristotle University of Thessaloniki. The strains
DC205 and DC215 may be considered as possible probiot-
ics because of their ability to withstand low pH values
(pH 3Æ0) and high bile concentration (N. Tzanetakis and
E. Litopoulou-Tzanetaki; unpublished data). The probiotic
strain Lact. acidophilus NCFB 1748 was isolated from
human intestine (Efthymiou and Hansen 1962) and is
commonly used by dairy industry as probiotic (Sullivan
and Nord 2006). NCFB 1748 was included in this study
on the basis that exhibited in vitro probiotic properties
similar to those described for Lact. paracasei strains
(Xanthopoulos et al. 1998, 2000) as well as immunomod-
ulatory activity (Nerstedt et al. 2007). The Lactobacillus
helveticus ATCC 15009 strain was commercially available
and exhibited no proven in vitro or in vivo probiotic prop-
erties. All lactobacilli were stored at )80�C in MRS broth
with 25% glycerol. Lactobacilli were subcultured at least
three times prior to experimental use. NCFB 1748 and
ATCC 15009 strains were grown for 18 h at 37�C, while
all Lact. paracasei subsp. paracasei strains were grown for
18 h at 30�C.
Preparation of strains
Bacterial cells from an overnight culture were washed
twice with 0Æ9% NaCl and finally resuspended at the
appropriate number (CFU ml)1) according to McFarland
standard. Labelling of Lactobacillus strains or heat-killed
baker’s yeast (Saccharomyces cerevisiae) with fluorosceine
isothiocyanate (FITC) was performed as described previ-
ously (Dogi and Perdigon 2006). Briefly, cells were washed
twice with phosphate-buffered saline (PBS) and then incu-
bated for 1 h at 37�C in 0Æ05 mol l)1 Na2CO3, pH 9Æ0,
containing 100 lg ml)1 FITC. After repeated washings,
the Lactobacillus-FITC or yeast-FITC pellets were resus-
pended in saline at 5 · 108 CFU ml)1 or 107 cells ml)1,
respectively, and stored at 4�C until used.
A. Kourelis et al. Probiotic injection in mouse air pouch
ª 2009 The Authors
Journal compilation ª 2009 The Society for Applied Microbiology, Journal of Applied Microbiology 108 (2010) 274–284 275

Animals
Balb ⁄ c (20–30 g) inbred mice or Fisher-344 inbred rats
(130–180 g) were housed under standard laboratory con-
ditions (12 h light ⁄ 12 h dark cycle) and received a diet of
commercial food pellets and tap water. Animals were
sacrificed by light ether anaesthesia followed by cervical
dislocation. All experiments were performed in an accred-
ited animal facility (number EL 54 BIO 02, School of
Biology, Aristotle University of Thessaloniki) and com-
plied with the current ethical regulations on animal
research of our university, which is according to both
Greek National and EU legislation. All groups included in
the present study consisted of five animals, and each
experiment was repeated at least two times.
Formation of the air pouch
Air pouches were created on the back of mice or rats as
described previously (Sedgwick et al. 1985) by subcutaneous
injection of 3 or 20 ml sterile air, respectively. The forma-
tion of equal size of air pouch in each animal was achieved
by refilling the 3- and 5-day-old air pouches with the
appropriate volume of air and confirmed using a vernier.
Subsequently, Lactobacillus strains were administered in the
6-day-old mouse or rat air pouches. Mice were injected with
0Æ2 ml pyrogenic-free saline containing 108 CFU of bacteria,
while rat air pouches received 5 · 108 CFU of bacteria in
1 ml of pyrogenic-free saline. Control animals in the air
pouch received only pyrogenic-free saline.
Determination of PMN accumulation in Lactobacillus
treated air pouches
The PMN cells accumulated in Lactobacillus-treated rat or
mouse air pouches were harvested 3 h postbacteria adminis-
tration after injection in the air pouch of 10 or 2 ml of saline,
respectively, followed by air pouch lavage. This time point
was selected because time-course kinetic experiments
revealed that in air pouches injected with the NCFB 1748
strain, the maximum accumulation of PMN cells and comp-
lete bacterial clearance occurred 3–6 h postbacteria admini-
stration (data not shown). After centrifugation, the
supernatant was collected, filtered through 0Æ22 lm filter
and hence referred to as the ‘air pouch exudate’. The cell
pellet containing the air pouch exudate PMN cells was
washed twice in saline and counted using a haemocytometer,
while cell viability was determined by trypan blue exclusion.
Phagocytic activity of air pouch PMN cells
Phagocytic activity of the rat air pouch PMN cells was
determined as described previously (Loose et al. 1978).
Heat-killed baker’s yeasts (106 cells) were opsonized by
incubation with rat serum for 30 min at 37�C. Then, the
106 opsonized heat-killed baker’s yeasts were mixed with
2Æ5 · 106 air pouch PMN cells and incubated at 37�C for
additional 30 min. Phagocytosis was stopped by the addi-
tion of FBS. Samples were then mixed with trypan blue,
and PMN cells containing phagocytosed yeasts were
counted using a haemocytometer under light microscope.
Phagocytic activity of air pouch PMN cells was also deter-
mined by flow cytometric-based assay using FITC-labelled
baker’s yeast (Nuutila and Lilius 2005). Samples were
then analysed on an EPICS XL (Coulter, Miami, FL,
USA) cytometer. At least 3 · 106 events were measured
for each sample at a low flow rate. To exclude yeast
adhesion from phagocytosis, parallel experiments were
performed on ice (0–4�C) with or without the addition
of trypan blue to quench external fluorescence. The
percentage of fluorescence of air pouch PMN cell popu-
lation on ice was excluded from the percentage estimated
for the same population at 37�C.
Cytokine detection in air pouch exudates
Air pouch exudates isolated from control or Lactobacillus
treated air pouches were subjected to ELISA to detect the
levels of gamma interferon (IFN-c), tumour necrosis
factor alpha (TNF-a) and interleukin (IL)-10 using the
ELISA kit (eBioscience, Inc., San Diego, CA, USA),
according to manufacturer’s instructions. As positive sam-
ples for the assay, we used purified cytokines included in
the kit as well as air pouch exudates isolated from bacte-
rial lipopolysaccharide or Freund’s complete adjuvant-
treated air pouches.
Preparation of paraffin blocks
Mouse air pouch membrane or intestine was removed,
dissected into small pieces, fixed in 4% paraformaldehyde
in PBS and finally embedded in paraffin blocks (Sainte-
Marie 1962). Sections of 4 lm thickness were either
stained with haematoxylin–eosin or further processed for
immunohistochemistry (Avramidis et al. 2002).
Interaction of Lactobacillus strains with air pouch lining
tissue or gut-associated lymphoid tissue (GALT)
Mice injected in the air pouch with 200 ll of each FITC-
labelled Lactobacillus strain (108 CFU) or orally received
100 ll of each FITC-labelled bacterial strain at doses of
108 CFU ml)1 were sacrificed 3 h or 20 min later, respec-
tively. To investigate whether the administered strains
interact with the air pouch lining tissue or GALT, air
pouch membranes and small or large intestine were
Probiotic injection in mouse air pouch A. Kourelis et al.
276 Journal compilation ª 2009 The Society for Applied Microbiology, Journal of Applied Microbiology 108 (2010) 274–284
ª 2009 The Authors

removed and embedded in paraffin blocks as described
above.
Immunohistochemical detection of cells producing IgA,
cytokine and TLR in the small and large intestines
Mice housed in separate cages orally received 50 ll saline
containing 109 CFU of each of the bacterial strains for
ten consecutive days, while control mice received only
50 ll of pyrogenic-free saline. One day after the last dose,
the intestine was removed and processed for paraffin
embedding.
The number of cells producing IgA or TLR2 was deter-
mined on samples by direct immunofluorescence using
monospecific anti-IgA (Sigma, St Louis, MO, USA) anti-
body or TLR2 (Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) conjugated with FITC. Indirect immuno-
fluorescence was used to determine the cells producing
TLRs 4, 6 and 9, cytokines and inflammatory mediators
by using specific antibodies directed against mouse TLRs
4, 9 (Abcam plc, Cambridge, UK), TLR6, COX-1, COX-2,
IL-12B (Santa Cruz Biotechnology, Inc), IFN-c, TNF-aand IL-10 (R&D systems, Inc., Minneapolis, MN, USA).
Antibodies were applied for 2 h at room temperature,
and after four washes in PBS, the slices were incubated
for 1 h with the FITC-conjugated anti-goat IgG (Sigma)
as secondary antibody. After four washes in PBS, the sam-
ples were examined using fluorescence light microscope.
The number of positive cells was counted in ten fields at
400· magnification. Samples incubated only with the
secondary antibody were also included to confirm
specificity of the immunodetection, and the number of
positive cells was excluded from the number of cells esti-
mated for testing samples. For the final scoring, two
investigators achieved consensus.
Statistical analysis
The results are reported as the mean ± SEM. Multiple
comparisons were performed by one-way anova followed
by Tukey’s test, and statistical significance was accepted at
values of P < 0Æ05.
Results
Recruitment of PMN in Lactobacillus-treated air pouches
To determine the in vivo probiotic characteristics of the
four Lact. paracasei subsp. paracasei strains included in
this study, we examined their ability to activate immune
responses in the dorsal air pouch of mice. The strains
Lact. acidophilus NCFB 1748 and Lact. helveticus ATCC
15009 were used as positive and negative controls, respec-
tively. Administration of bacterial strains in the mouse air
pouch resulted in rapid accumulation of significant num-
ber of PMN cells in the air pouch exudates (Table 1).
Recruitment of PMN cells in the air pouch exudates was
increased by 10- to 12-fold after administration of the
strains DC412 or NCFB 1748 and by three to fivefold
after administration of the strains DC215, DC205, B112
and ATCC 15009 (Table 1). Similar data were observed
after administration of the above strains in rat air
pouches (Table 1). The above data indicate that the
DC412 strain exhibits immunostimulatory activity in both
Table 1 Determination of the immunostimulatory activity of Lactobacillus paracasei subsp. paracasei strains in the mouse and rat dorsal
air pouch
PMN recruitment in air pouch
(cells · 10)6) Cytokine levels in mouse air pouch exudates (pg ml)1)
Mouse Rat IFN-c TNF-a IL-10
Control 0Æ6 ± 0Æ1 8Æ5 ± 1Æ9 89Æ4 ± 19Æ5 29Æ9 ± 1Æ9 43Æ6 ± 5Æ1
Lactobacillus acidophilus
NCFB 1748 7Æ2 ± 1Æ1* 73Æ8 ± 8Æ7* 68Æ4 ± 14Æ8 198Æ2 ± 18Æ1* 54Æ6 ± 10Æ9
Lact. paracasei subsp. paracasei
B112 1Æ5 ± 0Æ6*� ND 58Æ3 ± 20Æ3 231Æ1 ± 40Æ3* 35Æ4 ± 12Æ6
DC205 3Æ2 ± 0Æ3*� ND 74Æ3 ± 10Æ7 94Æ91 ± 19Æ6* 41Æ2 ± 14Æ8
DC215 1Æ7 ± 0Æ5*� 33Æ7 ± 8Æ2*� 81Æ4 ± 9Æ8 135Æ7 ± 26Æ1* 44Æ3 ± 18Æ6
DC412 6Æ6 ± 0Æ8* 75Æ4 ± 7Æ6* 68Æ4 ± 14Æ8 284Æ1 ± 45Æ4* 48Æ1 ± 5Æ6
Lactobacillus helveticus
ATCC 15009 1Æ8 ± 0Æ9*� 41Æ8 ± 7Æ1*� 64Æ4 ± 12Æ7 39Æ5 ± 6Æ6 61Æ7 ± 21Æ2
Values are means ± SEM from three independent experiments (n = 5).
ND, not determined.
*Statistically significant differences in comparison with the control group, P < 0Æ05.
�Statistically significant differences in comparison with the probiotic NCFB 1748-treated group, P < 0Æ05.
A. Kourelis et al. Probiotic injection in mouse air pouch
ª 2009 The Authors
Journal compilation ª 2009 The Society for Applied Microbiology, Journal of Applied Microbiology 108 (2010) 274–284 277
mice and rat air pouches, which equals the respective one
observed with the probiotic NCFB 1748 strain.
Effect of Lactobacillus on air pouch PMN
phagocytic activity
The total number of PMN cells accumulated in the air
pouches of normal treated mice (Table 1) is below the
number of cells (2Æ5 · 106) required to perform the phago-
cytic activity assay. To avoid mixing air pouch PMN cells
from a high number of normal mice, phagocytic activity
was performed using rat air pouch PMN cells. Kinetic
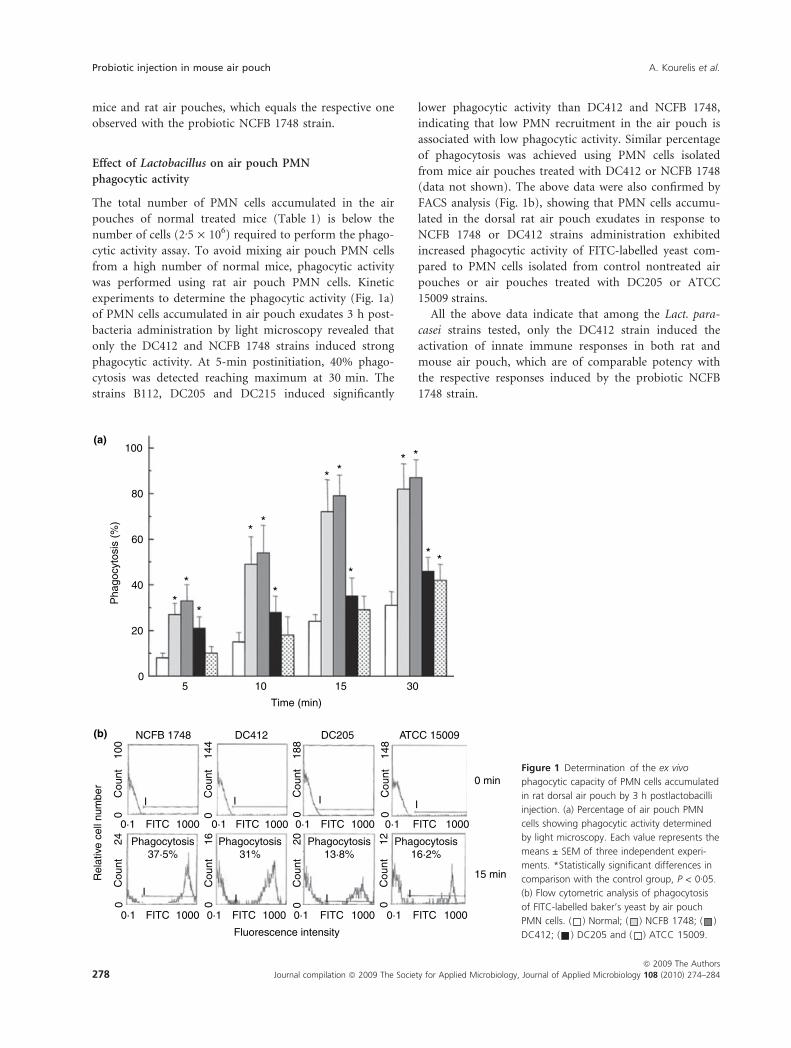
experiments to determine the phagocytic activity (Fig. 1a)
of PMN cells accumulated in air pouch exudates 3 h post-
bacteria administration by light microscopy revealed that
only the DC412 and NCFB 1748 strains induced strong
phagocytic activity. At 5-min postinitiation, 40% phago-
cytosis was detected reaching maximum at 30 min. The
strains B112, DC205 and DC215 induced significantly
lower phagocytic activity than DC412 and NCFB 1748,
indicating that low PMN recruitment in the air pouch is
associated with low phagocytic activity. Similar percentage
of phagocytosis was achieved using PMN cells isolated
from mice air pouches treated with DC412 or NCFB 1748
(data not shown). The above data were also confirmed by
FACS analysis (Fig. 1b), showing that PMN cells accumu-
lated in the dorsal rat air pouch exudates in response to
NCFB 1748 or DC412 strains administration exhibited
increased phagocytic activity of FITC-labelled yeast com-
pared to PMN cells isolated from control nontreated air
pouches or air pouches treated with DC205 or ATCC
15009 strains.
All the above data indicate that among the Lact. para-
casei strains tested, only the DC412 strain induced the
activation of innate immune responses in both rat and
mouse air pouch, which are of comparable potency with
the respective responses induced by the probiotic NCFB
1748 strain.
100(a)
(b)
80
60
40
Pha
gocy
tosi
s (%
)
20
05
**
**
* ** *
***
**
10 15 30
Time (min)
NCFB 1748
I
100
Cou
nt0
24C
ount
0
16C
ount
0
20C
ount
0
12C
ount
014
8C
ount
0
188
Cou
nt0
144
Cou
nt0
II I
II
I
I
0·1 FITC
Phagocytosis37·5%
Phagocytosis31%
Phagocytosis13·8%
Phagocytosis16·2%
1000 0·1 FITC
Fluorescence intensity
Rel
ativ
e ce
ll nu
mbe
r
1000 0·1 FITC 1000 0·1 FITC
0 min
15 min
1000
0·1 FITC 10000·1 FITC 10000·1 FITC 10000·1 FITC 1000
DC412 DC205 ATCC 15009
Figure 1 Determination of the ex vivo
phagocytic capacity of PMN cells accumulated
in rat dorsal air pouch by 3 h postlactobacilli
injection. (a) Percentage of air pouch PMN
cells showing phagocytic activity determined
by light microscopy. Each value represents the
means ± SEM of three independent experi-
ments. *Statistically significant differences in
comparison with the control group, P < 0Æ05.
(b) Flow cytometric analysis of phagocytosis
of FITC-labelled baker’s yeast by air pouch
PMN cells. ( ) Normal; ( ) NCFB 1748; ( )
DC412; ( ) DC205 and ( ) ATCC 15009.
Probiotic injection in mouse air pouch A. Kourelis et al.
278 Journal compilation ª 2009 The Society for Applied Microbiology, Journal of Applied Microbiology 108 (2010) 274–284
ª 2009 The Authors

Determination of TNF-a, IFN-c and IL-10 in air pouch
exudates
To examine whether the above Lactobacillus strains
induce differential production of cytokines in the air
pouch, we performed ELISA immunosorbent assay in the
air pouch exudates. The data in Table 1 show that among
the cytokines tested in Lactobacillus-treated mouse air
pouch exudates, only TNF-a levels were increased signifi-
cantly by 3 h postbacteria administration. However,
DC412 as well as B112 strains induced the higher TNF-alevels followed by NCFB 1748, DC205 and ATCC 15009.
The IFN-c and IL-10 levels were not increased by any of
the lactobacilli investigated. Similar data were obtained in
Lactobacillus-treated rat air pouch exudates (data not
shown). Taken together, all the above data indicate that
the strain DC412 exhibits immunoregulatory activity that
equals the respective one observed with the probiotic
NCFB 1748.
Interaction of FITC-labelled Lactobacillus strains with
the air pouch lining tissue and the GALT
The strains DC412, DC205, NCFB 1748 and ATCC 15009
were further studied for their ability to interact with the
cells forming the intima or subintima layer of the air
pouch (Fig. 2a). By fluorescence microscopy, we observed
differential interaction of the strains NCFB 1748, DC412,
DC205 and ATCC 15009 with the air pouch lining tissue
(Fig. 2a). The FITC-labelled DC412 cells or their antigens
were associated with both the intima and subintima layers
of the air pouch (Fig. 2b), while FITC-labelled NCFB
1748 cells or antigens were detected mainly in cells form-
ing the air pouch subintima layer (Fig. 2c). The strains
DC205 and ATCC 15009 showed low interaction with the
air pouch lining tissue. The above data indicate that the
potency of Lactobacillus to interact with the air pouch is
strain specific, while the differential interaction of DC412
and NCFB 1748 strains with the cells forming the air
pouch is suggestive of possible differential activation of
immune responses by these Lactobacillus strains.
To further investigate the capacity of DC412, NCFB
1748 and DC205 strains to interact with the gut, we used
FITC-labelled bacteria. DC412 and NCFB 1748 FITC-
labelled strains and their antigens exhibited increased
capacity to interact with the GALT (Fig. 2d–f). We
observed antigens of both fluorescent bacteria in immune
cells associated with the lamina propria of the small intes-
tine, in Peyer’s patches in macrophage and DC harbour-
ing areas and in nodules and crypts of the large intestine.
The strain DC205 showed low interaction with the GALT.
The above data show that the strain-specific interaction
observed in the intestine barrier is similar with the
respective one in the air pouch. In addition, the DC412
strain interacts with the intestine cells, suggesting poten-
tial colonization ability and capacity to activate immune
responses in the gut.
Detection of cells producing IgA, COX-1 and COX-2 in
the intestine
To confirm that the DC412 strain induces immune
responses in the small and large intestines, in vivo studies
were performed by feeding mice for 10 days with the
DC412 or NCFB 1748 strains. DC205 strain was also
included as a low responder positive control. Oral admin-
istration of Lactobacillus strains in mice and subsequent
haematoxylin–eosin-stained sections of the small intestine
(a) (b) (c)
(d) (e) (f)
Figure 2 Histological analysis of FITC-labelled
lactobacilli interaction with the cells forming
the air pouch lining tissue or the gut-associ-
ated lymphoid tissue. (a) Eosin–hematoxylin
staining of normal air pouch lining tissue
showing the intima (arrowheads) and
subintima (arrows) regions. (b–c) Fluorescence
microscopy of air pouch membrane 3 h post
(b) Lactobacillus paracasei subsp. paracasei or
(c) Lactobacillus acidophilus NCFB 1748
strains injection. (d–f) Fluorescence micro-
photography showing the interaction of
FITC-labelled DC412 bacteria with (d) the
lamina propria of the small intestine, (e) the
Peyer’s patches and (f) the crypts of the large
intestine.
A. Kourelis et al. Probiotic injection in mouse air pouch
ª 2009 The Authors
Journal compilation ª 2009 The Society for Applied Microbiology, Journal of Applied Microbiology 108 (2010) 274–284 279
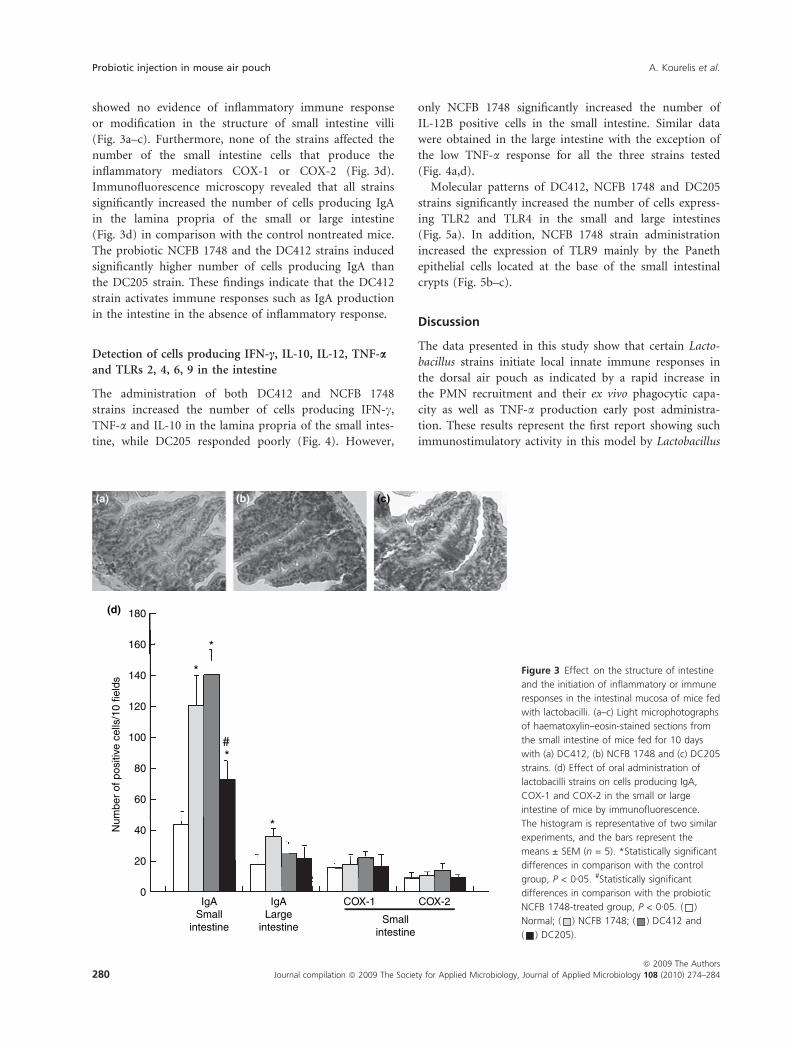
showed no evidence of inflammatory immune response
or modification in the structure of small intestine villi
(Fig. 3a–c). Furthermore, none of the strains affected the
number of the small intestine cells that produce the
inflammatory mediators COX-1 or COX-2 (Fig. 3d).
Immunofluorescence microscopy revealed that all strains
significantly increased the number of cells producing IgA
in the lamina propria of the small or large intestine
(Fig. 3d) in comparison with the control nontreated mice.
The probiotic NCFB 1748 and the DC412 strains induced
significantly higher number of cells producing IgA than
the DC205 strain. These findings indicate that the DC412
strain activates immune responses such as IgA production
in the intestine in the absence of inflammatory response.
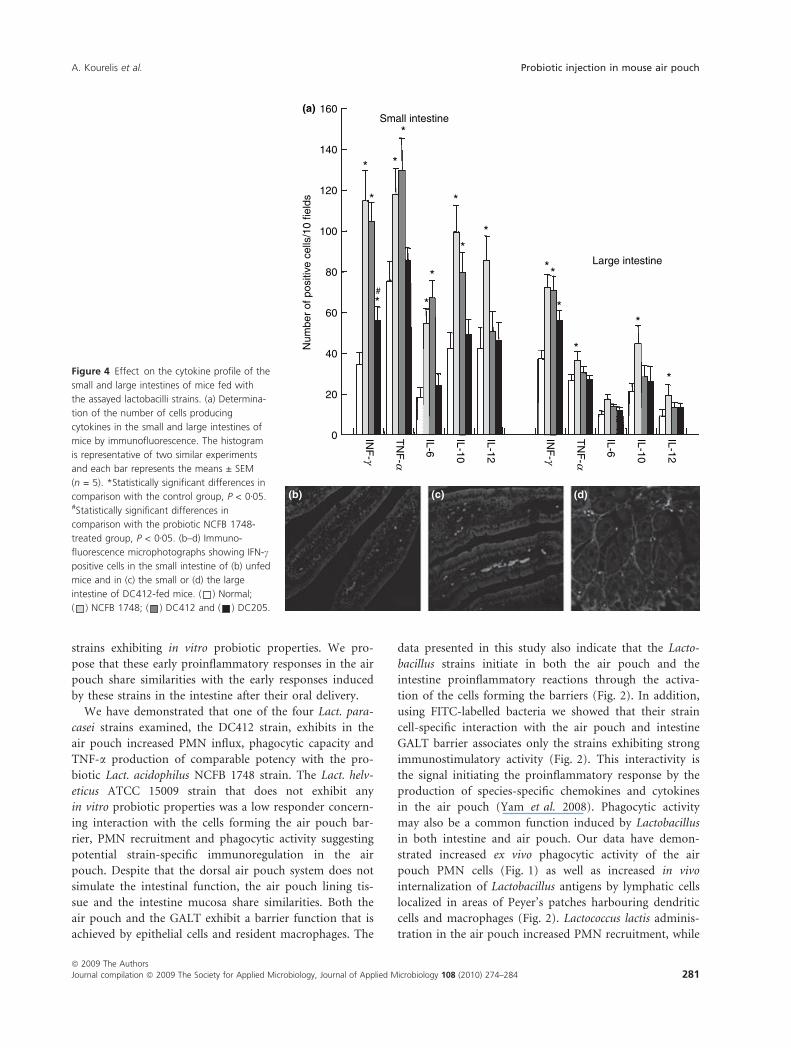
Detection of cells producing IFN-c, IL-10, IL-12, TNF-a
and TLRs 2, 4, 6, 9 in the intestine
The administration of both DC412 and NCFB 1748
strains increased the number of cells producing IFN-c,
TNF-a and IL-10 in the lamina propria of the small intes-
tine, while DC205 responded poorly (Fig. 4). However,
only NCFB 1748 significantly increased the number of
IL-12B positive cells in the small intestine. Similar data
were obtained in the large intestine with the exception of
the low TNF-a response for all the three strains tested
(Fig. 4a,d).
Molecular patterns of DC412, NCFB 1748 and DC205
strains significantly increased the number of cells express-
ing TLR2 and TLR4 in the small and large intestines
(Fig. 5a). In addition, NCFB 1748 strain administration
increased the expression of TLR9 mainly by the Paneth
epithelial cells located at the base of the small intestinal
crypts (Fig. 5b–c).
Discussion
The data presented in this study show that certain Lacto-
bacillus strains initiate local innate immune responses in
the dorsal air pouch as indicated by a rapid increase in
the PMN recruitment and their ex vivo phagocytic capa-
city as well as TNF-a production early post administra-
tion. These results represent the first report showing such
immunostimulatory activity in this model by Lactobacillus
180
160
140
120
100
80
60
Num
ber
of p
ositi
ve c
ells
/10
field
s
40
20
0IgA
Smallintestine
IgALarge
intestineSmall
intestine
COX-2COX-1
*
*
*
#
*
(a) (b)
(d)
(c)
Figure 3 Effect on the structure of intestine
and the initiation of inflammatory or immune
responses in the intestinal mucosa of mice fed
with lactobacilli. (a–c) Light microphotographs
of haematoxylin–eosin-stained sections from
the small intestine of mice fed for 10 days
with (a) DC412, (b) NCFB 1748 and (c) DC205
strains. (d) Effect of oral administration of
lactobacilli strains on cells producing IgA,
COX-1 and COX-2 in the small or large
intestine of mice by immunofluorescence.
The histogram is representative of two similar
experiments, and the bars represent the
means ± SEM (n = 5). *Statistically significant
differences in comparison with the control
group, P < 0Æ05. #Statistically significant
differences in comparison with the probiotic
NCFB 1748-treated group, P < 0Æ05. ( )
Normal; ( ) NCFB 1748; ( ) DC412 and
( ) DC205).
Probiotic injection in mouse air pouch A. Kourelis et al.
280 Journal compilation ª 2009 The Society for Applied Microbiology, Journal of Applied Microbiology 108 (2010) 274–284
ª 2009 The Authors
strains exhibiting in vitro probiotic properties. We pro-
pose that these early proinflammatory responses in the air
pouch share similarities with the early responses induced
by these strains in the intestine after their oral delivery.
We have demonstrated that one of the four Lact. para-
casei strains examined, the DC412 strain, exhibits in the
air pouch increased PMN influx, phagocytic capacity and
TNF-a production of comparable potency with the pro-
biotic Lact. acidophilus NCFB 1748 strain. The Lact. helv-
eticus ATCC 15009 strain that does not exhibit any
in vitro probiotic properties was a low responder concern-
ing interaction with the cells forming the air pouch bar-
rier, PMN recruitment and phagocytic activity suggesting
potential strain-specific immunoregulation in the air
pouch. Despite that the dorsal air pouch system does not
simulate the intestinal function, the air pouch lining tis-
sue and the intestine mucosa share similarities. Both the
air pouch and the GALT exhibit a barrier function that is
achieved by epithelial cells and resident macrophages. The
data presented in this study also indicate that the Lacto-
bacillus strains initiate in both the air pouch and the
intestine proinflammatory reactions through the activa-
tion of the cells forming the barriers (Fig. 2). In addition,
using FITC-labelled bacteria we showed that their strain
cell-specific interaction with the air pouch and intestine
GALT barrier associates only the strains exhibiting strong
immunostimulatory activity (Fig. 2). This interactivity is
the signal initiating the proinflammatory response by the
production of species-specific chemokines and cytokines
in the air pouch (Yam et al. 2008). Phagocytic activity
may also be a common function induced by Lactobacillus
in both intestine and air pouch. Our data have demon-
strated increased ex vivo phagocytic activity of the air
pouch PMN cells (Fig. 1) as well as increased in vivo
internalization of Lactobacillus antigens by lymphatic cells
localized in areas of Peyer’s patches harbouring dendritic
cells and macrophages (Fig. 2). Lactococcus lactis adminis-
tration in the air pouch increased PMN recruitment, while
Large intestine
Small intestine160
140
120
100
80
60N
umbe
r of
pos
itive
cel
ls/1
0 fie
lds
40
20
0 INF
-g
TN
F-a
IL-6
IL-10
IL-12
INF
-g
TN
F-a
IL-6
IL-10
IL-12
*
**
*
**#
**
*
*
*
*
*
*
*
*
(a)
(b) (c) (d)
Figure 4 Effect on the cytokine profile of the
small and large intestines of mice fed with
the assayed lactobacilli strains. (a) Determina-
tion of the number of cells producing
cytokines in the small and large intestines of
mice by immunofluorescence. The histogram
is representative of two similar experiments
and each bar represents the means ± SEM
(n = 5). *Statistically significant differences in
comparison with the control group, P < 0Æ05.#Statistically significant differences in
comparison with the probiotic NCFB 1748-
treated group, P < 0Æ05. (b–d) Immuno-
fluorescence microphotographs showing IFN-c
positive cells in the small intestine of (b) unfed
mice and in (c) the small or (d) the large
intestine of DC412-fed mice. ( ) Normal;
( ) NCFB 1748; ( ) DC412 and ( ) DC205.
A. Kourelis et al. Probiotic injection in mouse air pouch
ª 2009 The Authors
Journal compilation ª 2009 The Society for Applied Microbiology, Journal of Applied Microbiology 108 (2010) 274–284 281

PMN recruitment in the udder because of L. lactis
administration successfully treated bovine mastitis (Crispie
et al. 2008), suggesting that DC412 and NCFB 1748
strains may also exhibit pathogenic bacterial clearance
activity. Another similarity between the air pouch and the
intestine barriers is the increase in TNF-a production in
both the air pouch and the small intestine in response to
DC412 and NCFB 1748 administration, suggesting that
this proinflammatory cytokine may be considered among
the major initial signals required for the activation of
innate immune response by these strains. The increase in
TNF-a level may be required to initiate the interaction
between the intima and subintima cells of the air pouch
as this mechanism is also suggested for intestine epithelial
cells (Galdeano et al. 2007). The precise mechanism by
which DC412 and NCFB 1748 administration in the air
pouch resulted in increased PMN recruitment, phagocytic
activity and cytokine production remains unclear in this
study. Possible mechanism could include the activation of
chemokines such as MIP-2 (Walley et al. 1997; Yam et al.
2008), TLRs (Doyle et al. 2004) and ⁄ or adjuvant activity
(Vitini et al. 2000).
Based on the above, we propose the eligibility of the
air pouch model to discriminate presumptive probiotic
Lactobacillus strains exhibiting immunostimulatory acti-
vity. The efficiency of the air pouch model to discrimi-
nate micro-organisms possessing probiotic properties
with respect to their immunomodulatory activity has
been also successfully shown for other Lactobacillus spe-
cies as well as for yeast strains in the unpublished studies
of our group (C. Kotzamanidis 2009, A. Kourelis 2009).
Our studies also showed differences concerning the
inflammatory response in the air pouch versus the intes-
tine. On the basis of PMN recruitment, air pouch
administration of Lactobacillus strains activates pro-
inflammatory reaction of higher potency than the
respective response observed in the intestine after oral
administration, suggesting enhancing capacity of the air
pouch on the innate immune responses. This may be
because of higher number of resident immune cells as
well as bacterial antigen availability in the air pouch
than in the intestine. However, in the intestine, the
Lactobacillus internalization is not followed by activation
of a general inflammatory response because oral DC412
Large intestine
Small intestine
140
120
100
80
60
Num
ber
of p
ositi
ve c
ells
/10
field
s
40
20
0 TLR
2
TLR
4
TLR
6
TLR
9
TLR
2
TLR
4
TLR
6
TLR
9
*
**
**
*
*
**
**
*#
*
*
(a)
(b) (c)
Figure 5 Effect of NCFB 1748, DC412 or
DC205 strains oral administration on the
number of Toll-like receptor (TLR) positive
cells in small and large intestines. (a) Determi-
nation of the number of TLR positive cells in
the small and large intestines of mice by
immunofluorescence. The histogram is
representative of two similar experiments and
each bar represents the means ± SEM (n = 5).
*Statistically significant differences in
comparison with the control group, P < 0Æ05.#Statistically significant differences in
comparison with the probiotic NCFB 1748-
treated group, P < 0Æ05. (b–c) Immuno-
fluorescence microphotographs showing TLR9
positive cells in the small intestine lamina
propria crypts of (b) unfed mice or (c) NCFB
1748 fed mice, arrows show the TLR9 positive
Paneth cells on the base of crypts. ( )
Normal; ( ) NCFB 1748; ( ) DC412 and
( ) DC205.
Probiotic injection in mouse air pouch A. Kourelis et al.
282 Journal compilation ª 2009 The Society for Applied Microbiology, Journal of Applied Microbiology 108 (2010) 274–284
ª 2009 The Authors

and NCFB 1748 delivery does not affect the structure of
the villi and the massive influx of PMN (Fig. 3a–c) as
well as the number of cells producing the inflammatory
mediators COX-1 and COX-2 (Fig. 3d). Thus, the signi-
ficance of the route of the DC412 and NCFB 1748
delivery may reflect their potential immunoadjuvant or
immunoregulatory activity.
Oral administration of DC412 strain resulted in
increased immune responses concerning IgA (Fig. 3) and
cytokine production (Fig. 4) confirming that the data
obtained in the air pouch show immunoregulatory capa-
city of this strain. The DC205 strain that was a low
responder when administered in the air pouch also
responded poorly after its oral administration. Activation
of the innate immunity and IgA production are thought
to be the most important mechanisms involved in gut
mucosa immune stimulation by probiotics (Galdeano
et al. 2007). The involvement of TLRs in the mechanism
that activates the innate immune response in the gut
mucosa is well documented (Takeda et al. 2003). Our
findings demonstrated that DC412 induced the produc-
tion of IgA, TNF-a, IFN-c, IL-6 and IL-10 by small
intestine cells, and our studies indicate that this profile
could be mediated through TLR2 and TLR4 (Fig. 5). The
cytokine profile of IFN-c, TNF-a, IL-6, IL-10 and IL-12B
that is induced in the mouse intestine after oral adminis-
tration of the probiotic NCFB 1748 strain could be medi-
ated through TLR2 and TLR9. CpG sequences activate a
TLR9-mediated signalling cascade with subsequent release
of pro-inflammatory cytokines such as TNF-a, IL-6 and
IL-12 (Krieg 2002). Thus, the increased IL-12B observed
in NCFB 1748-treated mice could be because of the acti-
vation through intestinal epithelial cell surface TLR9
(Ewaschuk et al. 2007) or through macrophage ⁄ dendritic
cell endoplasmic TLR9 (Latz et al. 2004). These data are
in agreement with previous data concerning the cytokine
profile activated by LAFTI L10 (Lact. acidophilus) and
LAFTI L26 (Lact. paracasei) strains in mouse small
intestine (Paturi et al. 2007). Thus, the cytokine and TLR
pattern profiles observed in DC412 and NCFB 1748
strains are suggestive of potential differential activity in
which both strains promote balanced Th1 and Th2
immune responses. In summary, the DC412 as well as the
NCFB 1748 strains exhibit similar in vitro probiotic prop-
erties and immunostimulatory activity in the air pouch
and the gut mucosa with the exception of TLR9 expres-
sion, suggesting potential differential activity. Thus, the
DC412 strain may be considered as presumptive probiotic
candidate that differs from NCFB 1748.
In conclusion, the Lact. paracasei subsp. paracasei
DC412 strain exhibited regulatory activity of early
immune responses in both air pouch and intestine, which
is characterized by stimulation of PMN chemotaxis, phago-
cytic activity, TLR2 ⁄ TLR4 signalling and secretion of a cer-
tain cytokine profile. The dorsal air pouch system provides
the environment that permits the examination and predic-
tion of innate immune reactions in the gut and thus may
be used as a model to rapidly and efficiently discriminate
potential probiotic Lactobacillus strains.
References
Akira, S., Takeda, K. and Kaisho, T. (2001) Toll-like receptors:
critical proteins linking innate and acquired immunity.
Nat Immunol 2, 675–680.
Alvarez-Olmos, M.I. and Oberhelman, R.A. (2001) Probiotic
agents and infectious diseases: a modern perspective on a
traditional therapy. Clin Infect Dis 32, 1567–1576.
Avramidis, N., Victoratos, P., Yiangou, M. and Hadjipetrou-
Kourounakis, L. (2002) Adjuvant regulation of cytokine
profile and antibody isotype of immune responses to
Mycoplasma agalactiae in mice. Vet Microbiol 88, 325–338.
Braat, H., Rottiers, P., Hommes, D.W., Huyghebaert, N.,
Remaut, E., Remon, J.P., van Deventer, S.J., Neiryncks, S.
et al. (2006) A phase I trial with transgenic bacteria
expressing interleukin-10 in Crohn’s disease. Clin Gastro-
enterol Hepatol 4, 754–759.
Coates, N.J. and McColl, S.R. (2001) Production of chemokin-
es in vivo in response to microbial stimulation. J Immunol
166, 5176–5182.
Crispie, F., Alonso-Gomez, M., O’Loughlin, C., Klostermann,
K., Flynn, J., Arkins, S., Meaney, W., Ross, R.P. et al.
(2008) Intramammary infusion of a live culture for treat-
ment of bovine mastitis: effect of live lactococci on the
mammary immune response. J Dairy Res 75, 374–384.
Delcenserie, V., Martel, D., Lamoureux, M., Amiot, J., Boutin,
Y. and Roy, D. (2008) Immunomodulatory effects of
probiotics in the intestinal tract. Curr Issues Mol Biol 10,
37–54.
Dogi, C.A. and Perdigon, G. (2006) Importance of the host
specificity in the selection of probiotic bacteria. J Dairy Res
73, 357–366.
Doyle, S.E., O’Connell, R.M., Miranda, G.A., Vaidya, S.A.,
Chow, E.K., Liu, P.T., Suzuki, S., Suzuki, N. et al. (2004)
Toll-like receptors induce a phagocytic gene program
through p38. J Exp Med 199, 81–90.
Efthymiou, C. and Hansen, P.A. (1962) An antigenic analysis
of Lactobacillus acidophilus. J Infect Dis 110, 258–267.
Ewaschuk, J.B., Backer, J.L., Churchill, T.A., Obermeier, F.,
Krause, D.O. and Madsen, K.L. (2007) Surface expression
of Toll-like receptor 9 is upregulated on intestinal epithe-
lial cells in response to pathogenic bacterial DNA. Infect
Immun 75, 2572–2579.
FAO ⁄ WHO (2002) Guidelines for the Evaluation of Probiotics
in Food. London Ontario, Canada: Food and Agriculture
Organization of the United Nations and World Health
Organization Working Group Report.
A. Kourelis et al. Probiotic injection in mouse air pouch
ª 2009 The Authors
Journal compilation ª 2009 The Society for Applied Microbiology, Journal of Applied Microbiology 108 (2010) 274–284 283

Galdeano, C.M., de Moreno de LeBlanc, A., Vinderola, G.,
Bonet, M.E. and Perdigon, G. (2007) Proposed model:
mechanisms of immunomodulation induced by probiotic
bacteria. Clin Vaccine Immunol 14, 485–492.
Krieg, A.M. (2002) From A to Z on CpG. Trends Immunol 23,
64–65.
Latz, E., Visintin, A., Espevik, T. and Golenbock, D.T. (2004)
Mechanisms of TLR9 activation. J Endotoxin Res 10,
406–412.
Litopoulou-Tzanetaki, E. and Tzanetakis, N. (1992) Microbio-
logical study of white-brined cheese made from raw goat
milk. Food Microbiol 9, 13–19.
Loose, L.D., Silkworth, J.B. and Simpson, D.W. (1978) Influ-
ence of cadmium on the phagocytic and microbicidal
activity of murine peritoneal macrophages, pulmonary
alveolar macrophages, and polymorphonuclear neutrophils.
Infect Immun 22, 378–381.
Nerstedt, A.N., Nilsson, E.C., Ohlson, K., Hakansson, J., Svens-
son, L.T., Lowenadler, B., Svensson, U.K. and Mahlapuu,
M. (2007) Administration of Lactobacillus evokes coordi-
nated changes in the intestinal expression profile of genes
regulating energy homeostasis and immune phenotype in
mice. Br J Nutr 97, 1117–1127.
Nuutila, J. and Lilius, E.M. (2005) Flow cytometric quantita-
tive determination of ingestion by phagocytes needs the
distinguishing of overlapping populations of binding and
ingesting cells. Cytometry A 65, 93–102.
Paturi, G., Phillips, M., Jones, M. and Kailasapathy, K. (2007)
Immune enhancing effects of Lactobacillus acidophilus
LAFTI L10 and Lactobacillus paracasei LAFTI L26 in mice.
Int J Food Microbiol 115, 115–118.
Perdigon, G., Fuller, R. and Raya, R. (2001) Lactic acid bacte-
ria and their effect on the immune system. Curr Issues
Intest Microbiol 2, 27–42.
Sainte-Marie, G. (1962) A paraffin embedding technique for
studies employing immunofluorescence. J Histochem Cyto-
chem 10, 250.
Sedgwick, A.D., Moore, A.R., Al-Duaij, A.Y., Edwards, J.C. and
Willoughby, D.A. (1985) The immune response to pertus-
sis in the 6-day air pouch: a model of chronic synovitis. Br
J Exp Pathol 66, 455–464.
Steidler, L. (2003) Genetically engineered probiotics. Best Pract
Res Clin Gastroenterol 17, 861–876.
Steidler, L., Robinson, K., Chamberlain, L., Schofield, K.M.,
Remaut, E., Le Page, R.W. and Wells, J.M. (1998) Mucosal
delivery of murine interleukin-2 (IL-2) and IL-6 by recom-
binant strains of Lactococcus lactis coexpressing antigen
and cytokine. Infect Immun 66, 3183–3189.
Sullivan, A. and Nord, C.E. (2006) Probiotic lactobacilli and
bacteraemia in Stockholm. Scand J Infect Dis 38, 327–331.
Takeda, K., Kaisho, T. and Akira, S. (2003) Toll-like receptors.
Annu Rev Immunol 21, 335–376.
Tzanetakis, N. and Litopoulou-Tzanetaki, E. (1992) Changes
in numbers and kinds of lactic acid bacteria in Feta and
Teleme, two Greek cheeses from ewes’ milk. J Dairy Sci 75,
1389–1393.
Vinderola, G., Matar, C. and Perdigon, G. (2005) Role of
intestinal epithelial cells in immune effects mediated by
gram-positive probiotic bacteria: involvement of toll-like
receptors. Clin Diagn Lab Immunol 12, 1075–1084.
Vitini, E., Alvarez, S., Medina, M., Medici, M., de Budeguer,
M.V. and Perdigon, G. (2000) Gut mucosal immunosti-
mulation by lactic acid bacteria. Biocell 24, 223–232.
Vogel, G. (2008) Clinical trials: deaths prompt a review of
experimental probiotic therapy. Science 319, 557.
Walley, K.R., Lukacs, N.W., Standiford, T.J., Strieter, R.M.
and Kunkel, S.L. (1997) Elevated levels of macrophage
inflammatory protein 2 in severe murine peritonitis
increase neutrophil recruitment and mortality. Infect
Immun 65, 3847–3851.
Xanthopoulos, V., Tzanetakis, N. and Litopoulou-Tzanetaki, E.
(1998) In vitro effect of lactobacilli and pediococci on
cholesterol. Microbiol Aliments Nutr 16, 199–203.
Xanthopoulos, V., Hatzikamari, M., Adamidis, T., Tsakalidou,
E., Tzanetakis, N. and Litopoulou-Tzanetaki, E. (2000)
Heterogeneity of Lactobacillus plantarum isolates from feta
cheese throughout ripening. J Appl Microbiol 88,
1056–1064.
Yam, K.K., Pouliot, P., N’diaye, M.M., Fournier, S., Olivier,
M. and Cousineau, B. (2008) Innate inflammatory
responses to the Gram-positive bacterium Lactococcus
lactis. Vaccine 26, 2689–2699.
Probiotic injection in mouse air pouch A. Kourelis et al.
284 Journal compilation ª 2009 The Society for Applied Microbiology, Journal of Applied Microbiology 108 (2010) 274–284
ª 2009 The Authors
Copyright © 2022 FDOKUMEN




















