
Atmospheric pressure chemical ionization tandem mass spectrometry of carotenoids

0014-2980/01/0505-1485$17.50+.50/0© WILEY-VCH Verlag GmbH, D-69451 Weinheim, 2001
In vivo MHC class II presentation of cytosolicproteins revealed by rapid automated tandem massspectrometry and functional analyses
Ashok R. Dongre1,2, Susan Kovats2,5, Paul deRoos1,2, Ashley L. McCormack3,Terry Nakagawa1,2, Vladislava Paharkova-Vatchkova5, Jimmy Eng3, Harlan Caldwell4,John R. Yates III3 and Alexander Y. Rudensky1,2
1 Howard Hughes Medical Institute, University of Washington School of Medicine, Seattle, USA2 Department of Immunology, University of Washington School of Medicine, Seattle, USA3 Department of Molecular Biotechnology, University of Washington School of Medicine, Seattle,
USA4 NIAID Rocky Mountain Laboratories, The Laboratory of Intracellular Parasites, Hamilton, USA5 Division of Immunology, Beckman Research Institute of the City of Hope, Duarte, USA
We report a strategy for high through-put sequence analyses of large MHC class II-boundpeptide repertoires which combines automated electrospray ionization tandem mass-spectrometry with computer-assisted interpretation of the tandem mass spectra using thealgorithm SEQUEST. This powerful approach discerned 128 peptide sequences displayedby the murine MHC class II molecule I-Ab in activated B cells and macrophages, including asurprisingly large number of peptides derived from self cytosolic proteins. Mice lacking thechaperone molecule H-2M were used to generate T cells specific for selected self peptides.Functional T cell analyses of ex vivo antigen-presenting cells indicated that peptides origi-nating from cytosolic proteins are efficiently presented by splenic and thymic dendritic cells,but less so by resting B cells or thymic cortical epithelial cells. These results suggest thatcentral tolerance to at least some MHC class II-bound self peptides derived from cytosolicproteins exists in vivo.
Key words: MHC class II presentation / Cytosolic peptide / Tandem mass-spectrometry
Received 10/1/01Accepted 19/2/01
[I 21643]
The first three authors equally contributed to this study.
Abbreviations: ESI-MS/MS: Electrospray ionization tan-dem mass spectrometry GAPDH: Glyceraldehyde-3-phosphate dehydrogenase AAT: Aspartate aminotransfer-ase cTEC: Thymic cortical epithelial cells SC: Spleno-cytes DC: Dendritic cells
1 Introduction
T cell recognition of foreign and self antigens in the formof short antigenic peptides bound to class I or class IIproteins encoded in the major histocompatibility genelocus (MHC) establishes a central paradigm for immuneresponses. Display of self peptides by MHC moleculeson APC figures importantly in both normal and abnormalfunctioning of the immune system. MHC-bound selfpeptides shape the developing TCR repertoire duringtolerance induction in the thymus and periphery, andthey may become targets of autoreactive T cells, leadingto initiation or progression of autoimmune disease.MHC-bound peptides derived from polymorphic regions
of donor proteins may also promote graft rejection byfunctioning as minor histocompatibility antigens [1].However, with regard to many of these immune pro-cesses, little is known about the precise nature and ori-gin of these self peptides, or the cell types on which theyare displayed. Thus, sequence information regarding therepertoire of self peptides bound to MHC molecules innormal and diseased tissue will provide insight into T celltolerance, autoimmunity, tumor immunity and transplan-tation biology.
These goals have led to development of methods forsequence analyses of MHC-bound peptides. Edmanmicrosequencing of HPLC fractionated peptide mixtureshas been used to characterize predominant peptidesbound to MHC class II molecules [2–6]. However, thepeptide complexity of these fractions poses serious limi-tations for sequence determination by this method. Huntet al. [7, 8] pioneered the use of electrospray ionizationtandem mass spectrometry (ESI-MS/MS) for analyses ofpeptides bound to both MHC class I and class II mole-cules. Using ESI-MS/MS, the murine class II allele, I-Ad,was found to be bound to 650–2,000 different peptides[8]; however, only 10–20 sequences of class II-bound
Eur. J. Immunol. 2001. 31: 1485–1494 In vivo MHC class II presentation of cytosolic proteins 1485

peptides have been identified previously by thisapproach, due to a slow tandem mass spectral dataacquisition and laborious manual interpretation of pep-tide sequences. Manual derivation of long class II-boundpeptide sequences from tandem mass spectra is furthercomplicated by highly protonated peptides produced bythe electrospray ionization process. The increase in pep-tide size frequently leads to an increase in observedcharge states which, upon ion activation, results inredundant fragmentation patterns with different chargestates.
To analyze the complex mixtures of MHC-bound pep-tides efficiently, we have applied a strategy for rapid,high through-put sequence analyses of MHC class II-bound peptides, which combines improved samplepreparation with automated tandem mass-spectral dataacquisition and computer-assisted interpretation of tan-dem mass spectra. Peptide sequences were discernedwith the computer algorithm SEQUEST, which uses theprotein and nucleotide sequence databases to performcross-correlation analyses of experimental and theoreti-cally generated tandem mass spectra [9, 10]. Using thisstrategy, we characterized the peptide repertoires dis-played by the murine MHC class II molecule I-Ab
expressed in macrophages and B cells. In addition topeptides derived from secretory and transmembraneproteins that traffic through the endocytic pathway, alarge proportion of peptides identified was derived fromcytosolic proteins. Functional analyses using peptide-specific T cells, developed in H-2M-deficient mice,showed that the peptides originating from select cyto-solic proteins were presented by ex vivo splenic and thy-mic APC. Our findings suggest that the routine display ofclass II-bound peptides originating from self cytosolicproteins induces tolerance to these peptides in vivo.
2 Results and discussion
2.1 Use of automated tandem massspectrometry and SEQUEST for sequenceanalyses of peptide repertoires displayed byMHC class II molecules
MHC class II I-Ab molecules were purified from cellsderived from C57BL/6 (B6) mice: the B lymphoma 1153,LPS-induced splenic B cell blasts, and the IFN- + -induced macrophage cell line BMC2.3. NP40 cell lysates(from 5×109 cells) were subjected to affinity chromatog-raphy using the I-Ab-specific mAb Y3P. Since polymericdetergents such as NP40 ionize well in the electrosprayionization process, trace detergent contamination in-creases the irrelevant signal and effectively competeswith the ionization of peptides. Thus, any NP40 associ-
ated with class II molecules was removed by exchangewith 1.5% sodium deoxycholate and washes with highand low salt-containing buffers. Peptides were releasedfrom ˚ 200 ? g of I-Ab by incubation in 2.5 M acetic acid,and collected by ultrafiltration through a 10-kDa cut offmembrane. The peptide mixture was fractionated on aC18 reverse phase-HPLC column and ˚ 30 fractionswere subjected to reverse phase-HPLC over a Poros10R2 microcapillary column interfaced with the elec-trospray ionization source of a Finnigan TSQ 7000 triplequadrupole mass spectrometer.
The peptide eluent from the microcapillary column wasintroduced directly into the electrospray ionizationsource. Two experiments were performed in series: thefirst recorded molecular mass information and the sec-ond obtained fragment ion masses for a selected proton-ated peptide. To automate this process, we developedsub-routines using Instrument Control Language (ICL)software, which perform mass spectral (MS) and tandemmass spectral (MS/MS) data acquisition by switching theinstrument parameters between MS and MS/MS modeevery 4–6 s. This allows 50–100 tandem mass spectra tobe acquired per HPLC fraction. Each tandem massspectrum was analyzed by the computer algorithmSEQUEST [9, 10] using the non-redundant protein (OWLand NCI) and EST databases. Peptide sequences wereidentified when the experimental tandem mass spectrummatched the database-derived theoretical tandem massspectrum with a cross correlational score greater than3.0×104. These tandem mass spectra were confirmedsubsequently by manual interpretation.
The 128 distinct peptide sequences (Table 1) are a com-pilation of two such analyses for each of the three cellu-lar sources of class II molecules. Of the peptides identi-fied during the first analysis, 95% were detected in sub-sequent runs, demonstrating the reproducibility of thistechnique. The origins of an additional ˚ 100 peptideions, with unique mass:charge ratios that producedstrong tandem mass spectra, have not been identifieddue to incomplete murine DNA and protein databases. Asignificant number of class II-bound peptide sequenceswere shared by macrophages and B cells.
As in previous reports, the majority of identified peptideswere derived from transmembrane or secreted proteinsthat intersect with, or reside in, the endocytic com-partment: e.g. IgM, apolipoproteins B and E, MHCclass II, CD22, LDL receptor, § 2-macroglobulin receptor,integrin- g 1, H-2M, g -galactosidase, invariant chain, g 2-microglobulin ( g 2m), and fetuin (Table 1 and 2). In addi-tion, 21% of peptides were derived from cytosolic pro-teins. The origins of these peptides include cytoskeletalproteins (actin, tubulin, F-actin capping protein), consti-
1486 A. R. Dongre et al. Eur. J. Immunol. 2001. 31: 1485–1494

Eur. J. Immunol. 2001. 31: 1485–1494 In vivo MHC class II presentation of cytosolic proteins 1487

tutive proteins [glyceraldehyde-3-phosphate dehydroge-nase (GAPDH), aspartate aminotransferase (AAT)], andproteins involved in vesicular trafficking (the GTP exchangeprotein Rab5A) (Table 3). These peptides were identifiedusing the non-redundant OWL protein database. Theremaining unidentified experimental tandem mass spectrawere searched against the murine nucleotide and ESTdatabases, leading to the identification of a peptide derivedfrom Clp36, a cytosolic LIM domain protein [11], and sev-eral peptides of unknown origin (Table 3 and 4).
To assess the I-Ab-binding capability of identified pep-tides, the corresponding synthetic peptides were testedfor their ability to inhibit binding of biotinylated E § 52–68peptide, a proven high affinity I-Ab binder, to purified I-Ab
(Table 5). All tested peptides were able to inhibit thebinding of the indicator peptide to I-Ab. The tandemmass spectra generated for all synthesized peptidesmatched precisely with the naturally occurring ones(data not shown).
2.2 Display of peptides derived from cytosolicproteins by MHC class II moleculesexpressed on ex vivo-derived APC
The large number of class II-bound peptides originatingfrom cytosolic proteins identified in our study, combinedwith previous detection of naturally processed actin,
GAPDH and Hsp70 epitopes [5, 6, 12, 13], and observa-tions of class II-mediated presentation of cytosolic viralepitopes [14–19], suggests that presentation of thesepeptides may play a significant role in immune re-sponses. Thus, we developed a strategy to determinewhether peptides naturally processed from self cytosolicproteins are normally presented by MHC class II mole-cules in vivo, and if so, what types of APC display thesepeptides.
To facilitate assessment of self peptide:class II complexexpression in vivo, a novel approach was used to gener-ate a panel of T cell hybrids specific for I-Ab-bound selfpeptides of cytosolic origin. Mice lacking the MHC classII-like molecule H-2M (M–/–) have a severely contractedclass II-bound endogenous peptide repertoire whichapparently skews the T cell repertoire such that M–/– Tcells respond vigorously to the class II-bound self pep-tides in wild-type B6 splenocytes (SC) [20–22]. We rea-soned that the M–/– mice may not be tolerant to themajority of self peptides, or that their tolerance would beoverridden by peptide immunization. We successfullygenerated I-Ab-restricted T cell hybrids specific for thenaturally processed actin, AAT, Clp36 and Rab5A epi-topes upon peptide immunization of M–/– mice. Each ofthese T cell hybrids responds to M–/– or Ii–/– SC in thepresence of the cognate peptide, but not to any of 19irrelevant self or foreign I-Ab binding peptides tested(Fig. 1A and data not shown). The irrelevant I-Ab binding
1488 A. R. Dongre et al. Eur. J. Immunol. 2001. 31: 1485–1494

peptides that were tested are included in Table 2. Thus, H-2M-deficient mice can be used generally as a source of Tcells specific for class II-bound self peptides, providingtools to assess presentation of self antigens in vivo.
A conventional approach was used to generate T cellhybrids specific for g 2m, IgM and CD22 epitopes.Mutant mice containing targeted disruptions of the g 2m,Ig ? chain, and CD22 genes were immunized with thecorresponding peptides to produce T cell hybrids. Eachof these T cell hybrids was specific for the cognate pep-tide presented by SC isolated from the g 2m–/–, IgM–/–,CD22–/– or M–/– mice [22].
Each self peptide-specific T cell hybrid recognizedendogenously formed peptide:class II complexes on
wild-type SC and the original cellular sources of the pep-tides (Fig. 1B, C; and data not shown). The ability ofB6 SC to form these cytosolic and transmembranepeptide:I-Ab complexes was determined by incubation ofvariable numbers of ex vivo isolated SC with the T cellhybrids in the absence of exogenously added antigen.Peptides derived from cell surface proteins were dis-played efficiently by ex vivo isolated SC at low APC num-bers. However, T cell responses to the cytosolicAAT, actin and Rab5A epitopes diminished rapidlywith decreasing numbers of APC. Approximately5×104–5×105 SC/well were required to detect a minimalT cell response to these cytosolic antigens, as com-pared to 5×102–5×103/well for the IgM, CD22 and g 2mantigens.
Eur. J. Immunol. 2001. 31: 1485–1494 In vivo MHC class II presentation of cytosolic proteins 1489

These data suggest that a minor subpopulation ofsplenic APC is capable of presenting these cytosolicantigens, or that these peptides are displayed at low lev-els on most APC. Thus, we investigated discrete class II+
APC populations. Little is known about the routine dis-play of class II-bound self peptides by different types ofresting APC in vivo. Using the T cell hybrids, we com-pared the display of cytosolic and non-cytosolic epi-topes by freshly isolated peripheral (splenic B cells andDC) and thymic [DC (tDC) and cortical epithelial cells(cTEC)] APC.
The majority of APC in the resting spleen are B cells,while DC and macrophages comprise about 2% of sple-nocytes. B220+ B cells and CD11c+ DC were isolated byFACS sorting, and plated directly with T cells withoutadditional activation (Fig. 2). Compared to the nearlyequivalent abilities of B cells and DC to present the g 2mepitope, these cytosolic epitopes were presented muchmore efficiently by DC; 50–100 fewer DC than B cellswere required to achieve the same level of T cell activa-tion. Both B cells and DC expressed negligible amountsof CD80 and CD86 and abundant cell surface MHC classII; DC expressed about 1.6-fold more surface class IIthan B cells (mean fluorescence intensity 1205 vs. 750).Since expression of costimulatory molecules on both celltypes was negligible, these data suggest that mecha-nisms for presentation of cytosolic antigens operatemore efficiently in DC, although the differential ability ofDC and B cells to present cytosolic epitopes may resultpartially from differences in surface MHC class II expres-sion. LPS activation of splenic B cells for 48 h did notincrease significantly the presentation of cytosolic epi-topes, suggesting that pathways for cytosolic antigen
1490 A. R. Dongre et al. Eur. J. Immunol. 2001. 31: 1485–1494
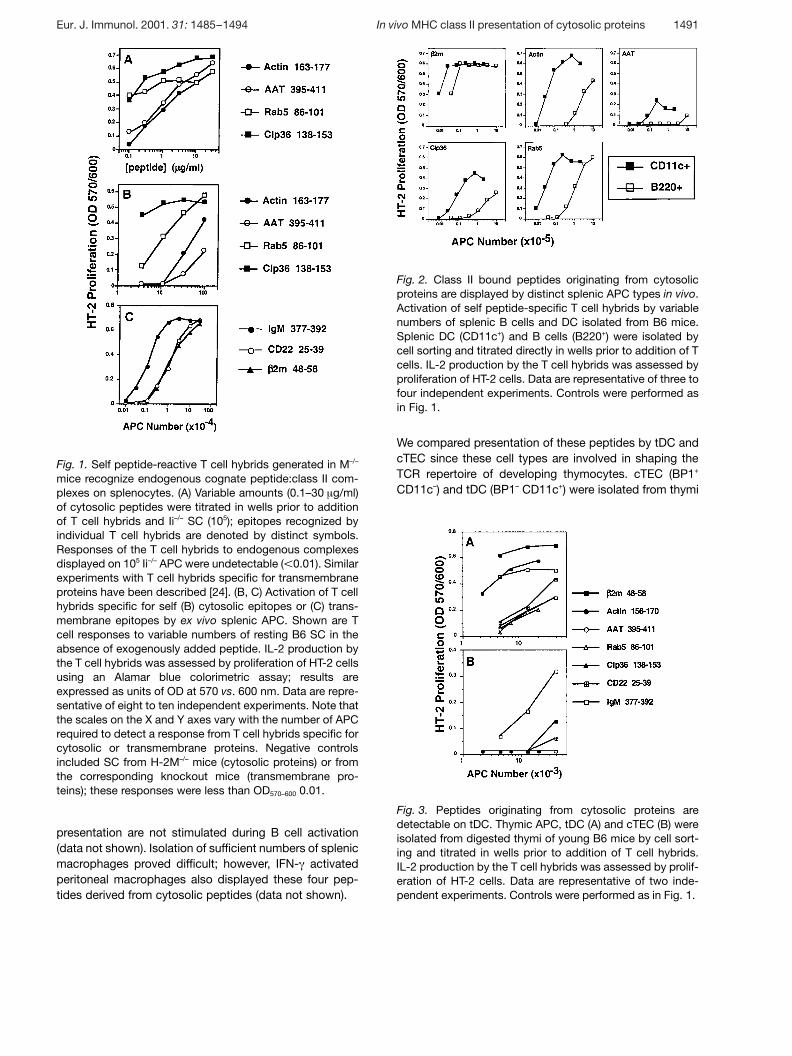
Fig. 1. Self peptide-reactive T cell hybrids generated in M–/–
mice recognize endogenous cognate peptide:class II com-plexes on splenocytes. (A) Variable amounts (0.1–30 ? g/ml)of cytosolic peptides were titrated in wells prior to additionof T cell hybrids and Ii–/– SC (105); epitopes recognized byindividual T cell hybrids are denoted by distinct symbols.Responses of the T cell hybrids to endogenous complexesdisplayed on 105 Ii–/– APC were undetectable ( X 0.01). Similarexperiments with T cell hybrids specific for transmembraneproteins have been described [24]. (B, C) Activation of T cellhybrids specific for self (B) cytosolic epitopes or (C) trans-membrane epitopes by ex vivo splenic APC. Shown are Tcell responses to variable numbers of resting B6 SC in theabsence of exogenously added peptide. IL-2 production bythe T cell hybrids was assessed by proliferation of HT-2 cellsusing an Alamar blue colorimetric assay; results areexpressed as units of OD at 570 vs. 600 nm. Data are repre-sentative of eight to ten independent experiments. Note thatthe scales on the X and Y axes vary with the number of APCrequired to detect a response from T cell hybrids specific forcytosolic or transmembrane proteins. Negative controlsincluded SC from H-2M–/– mice (cytosolic proteins) or fromthe corresponding knockout mice (transmembrane pro-teins); these responses were less than OD570–600 0.01.
Fig. 2. Class II bound peptides originating from cytosolicproteins are displayed by distinct splenic APC types in vivo.Activation of self peptide-specific T cell hybrids by variablenumbers of splenic B cells and DC isolated from B6 mice.Splenic DC (CD11c+) and B cells (B220+) were isolated bycell sorting and titrated directly in wells prior to addition of Tcells. IL-2 production by the T cell hybrids was assessed byproliferation of HT-2 cells. Data are representative of three tofour independent experiments. Controls were performed asin Fig. 1.
Fig. 3. Peptides originating from cytosolic proteins aredetectable on tDC. Thymic APC, tDC (A) and cTEC (B) wereisolated from digested thymi of young B6 mice by cell sort-ing and titrated in wells prior to addition of T cell hybrids.IL-2 production by the T cell hybrids was assessed by prolif-eration of HT-2 cells. Data are representative of two inde-pendent experiments. Controls were performed as in Fig. 1.
presentation are not stimulated during B cell activation(data not shown). Isolation of sufficient numbers of splenicmacrophages proved difficult; however, IFN- + activatedperitoneal macrophages also displayed these four pep-tides derived from cytosolic peptides (data not shown).
We compared presentation of these peptides by tDC andcTEC since these cell types are involved in shaping theTCR repertoire of developing thymocytes. cTEC (BP1+
CD11c–) and tDC (BP1– CD11c+) were isolated from thymi
Eur. J. Immunol. 2001. 31: 1485–1494 In vivo MHC class II presentation of cytosolic proteins 1491

of young mice by FACS sorting. tDC efficiently presentedall peptides tested, whereas cTEC only stimulateddetectable responses from those T cells specific for pep-tides derived from cell surface proteins (IgM, g 2m, CD22)(Fig. 3). It is interesting to note that thymic APC displaypeptides derived from proteins (IgM and CD22) not nor-mally endogenously expressed by DC and TEC.
Several pieces of evidence suggest that our T cellhybrids may be used to detect quantitative differences incognate peptide:class II complexes on APC, and are notdifferentially dependent upon costimulatory, adhesion orCD4 molecules. (1) These T cells express similar levels ofCD4. (2) The response of the T cells specific for g 2m,AAT, actin, Clp36 and Rab5A epitopes did not signifi-cantly change upon LPS activation of splenic B cells,although the activated B cells had significantly up-regulated surface expression of CD80 and CD86. (3)Both splenic DC and B cells express low levels of CD80/86, yet DC presented cytosolic epitopes much more effi-ciently. (4) These T cells respond to plate bound pep-tide:class II complexes (data not shown), suggesting thattheir differential activation is most likely due to differ-ences in numbers of cognate peptide:class II complexes.In some APC types, these differences in cognate com-plexes may reflect differences in overall surface class IIexpression.
Our experiments demonstrate that compared to splenicB cells, thymic and splenic DC are able to present pep-tides originating from cytosolic proteins extremely effi-ciently without prior activation. The constitutive displayof these cytosolic epitopes by DC argues that the T cellsin the periphery of wild-type mice have been tolerized tothese epitopes by mechanisms of either central orperipheral tolerance.
Class II-mediated presentation of these cytosolic epi-topes does not require the TAP-mediated mechanism ofpeptide transport from the cytosol to the endoplasmicreticulum, as judged using SC from TAP1-deficient mice(S.K. and A.Y.R., data not shown). Potential mechanismsby which cytosolic proteins or peptides could enter theendocytic pathway include: (1) uptake of dying cells [23];(2) micro- and macroautophagy [23, 24]; and (3) stress-induced selective transport of cytosolic proteins intolysosomes mediated by heat shock protein Hsp73 andthe lysosomal protein lgp96 [25].
3 Concluding remarks
Using a powerful approach for sequence analyses ofcomplex repertoires of peptides bound to MHC class IImolecules, we have identified 128 peptides bound to
I-Ab molecule in B cells and macrophages. We also showthat T cells specific for selected self peptides can begenerated in H-2M-deficient mice and used to comple-ment sequence analyses through functional studies ofthe in vivo expression of self peptide:class II complexesin different cell types. This approach may be useful foridentification of MHC class II-associated pathogen-derived peptides, or for comparison of class II-boundpeptides present in quiescent and inflamed tissues.
Our experiments revealed that MHC class II presentationin vivo is not limited to secreted or transmembrane pro-teins, but also includes a significant pool of peptidesderived from cytosolic proteins. MHC class II-mediatedpresentation of self peptides, derived from proteins thatnormally do not intersect the endocytic compartment, byDC may be important for establishing tolerance to selfproteins which may gain access to endosomal/lyso-somal compartments under pathological conditions.Recent studies in macrophages and DC have demon-strated efficient MHC class I-mediated presentation ofexogenous protein antigens internalized by macropino-cytosis or phagocytosis [26–28]. Hence, there is anextensive antigenic “cross-talk” between the classicalMHC class I and class II presentation pathways in pro-fessional APC such as DC and macrophages. Furtherdelineation of mechanisms regulating class II-mediatedpresentation of cytosolic proteins, and assessment oftheir potential role in development of autoimmunity are ofconsiderable interest.
4 Materials and methods
4.1 Purification of MHC class II-bound peptides
Cells (5×109) were lysed at 1×108/ml in PBS, 1% NP40,25 mM iodoacetamide, PMSF, TLCK, pH 7.4. I-Ab was iso-lated by affinity chromatography on an mAb Y3P-sepharosecolumn as described [3, 29]. The purified class II moleculeswere washed sequentially with (a) 1.5% deoxycholate in20 mM TRIS at pH 7.4, (b) 500 mM NaCl in 20 mM TRISpH 7.4, (c) 150 mM NaCl in 20 mM TRIS pH 7.4, (d) 20 mMTRIS pH 7.4. The peptides were eluted with 2.5 M aceticacid and separated from high molecular weight materialby ultrafiltration through a 10-kDa cut-off Amicon filter. Thepeptides were fractionated on a Vydac C18 column(2.1×250 mm) with a 90-min gradient of solvent B (80:20 v/vacetonitrile: 0.1%TFA) from 0% to 70% in solvent A (5:95 v/vacetonitrile: 0.1%TFA) at a flow rate of 250 ? l/min. Fractions( ˚ 50–65) were collected throughout the gradient and storedat –70°C.
1492 A. R. Dongre et al. Eur. J. Immunol. 2001. 31: 1485–1494

4.2 Microcapillary HPLC and automated tandem massspectrometry (ESI-MS/MS) on Finnigan TSQ 7000
Microcapillary columns were made from 100×200- ? m fusedsilica capillary tubing (Polymicro Technologies, Phoenix, AZ).The column frits were made by fusing a plug of LiChrosorbSi 60 beads (EM Separations Technologies, Gibbstown, NJ)to fused silica capillary walls. Columns, 10–12 cm, werepacked with Poros 10R2 reverse-phase packing material(PerSeptive Biosystems, Framingham, MA). Samples wereinjected on to the column using a helium pressure bomb at500 psi. Microcapillary HPLC was performed using ABI140B HPLC system (Applied Biosystems, Foster City, CA) atthe flow rate of 1 ? l/min with 30-min 0–60% gradient elutionconsisting of solvent B (80:20 v/v acetonitrile: 0.5% aceticacid) in solvent A (0.5% acetic acid). The column wasinserted directly into the Finnigan electrospray needle andthe post column flow rate was adjusted by flowing sheathliquid (methanol/water 70:30 containing 0.01% acetic acid)to a total of 2–3 ? l/min. Automated tandem mass spectraldata acquisition was performed on a Finnigan TSQ 700 tan-dem mass spectrometer (San Jose, CA) using ICL software.ICL subroutines are available upon request. The collisionenergy and mass range for tandem mass spectral scanswere selected by assuming that the precursor ion selected isdoubly charged. Four tandem mass spectra for the mostabundant peak were collected and the instrument was resetto scan in the normal mass spectral mode. The peak widthon the first mass analyzer (Q1) was set to 3.0u and thirdmass analyzer (Q2) was set to 2.0u. Typically, 60–80 loopswere performed per fraction subjected to microcapillaryHPLC.
4.3 Synthetic peptides and peptide binding studies
Peptides were synthesized on a Synergy 432 automatedpeptide synthesizer (Applied Biosystems, Foster City, CA)using Fmoc chemistry; purity of peptides was G 90%. Pep-tide binding assays were performed as described [20].
4.4 Generation and use of T cell hybridomas andisolation of APC
Mice were immunized in the hind footpads with syntheticpeptides (50 ? g) in CFA. After 7–10 days, T cells from drain-ing lymph nodes were restimulated in vitro with peptide for3 days and fused with BW5147. T cell hybrids were testedfor MHC restriction and peptide specificity by incubationwith variable amounts of either cognate or irrelevant pep-tides and M–/– or Ii–/– SC. T cell hybrids specific for actin163–177 (15.10), Clp36 138–153 (3.5), AAT 395–411 (15.15),and Rab5A 86–101 (2.3) were generated in M–/– mice [21],and T cell hybrids specific for g 2m 48–58 (4.1), IgM 377–392(77.1) and CD22 25–39 (7.6) were generated in mice lackingg 2m, IgM or CD22 [30]. T cell hybrids (2×104–10×104/well)
were incubated with variable numbers of ex vivo APC for
20 h. IL-2 production by the T cell hybrids was assessed byproliferation of HT-2 cells using an Alamar blue colorimetricassay; results are expressed as arbitrary units of OD at 570vs. 600 nm, average of duplicate wells.
For isolation of splenic B cells and dendritic cells (DC),splenic fragments from B6 mice were sequentially digestedwith several batches of 1 mg/ml collagenase/dispase (Boeh-ringer Mannheim), 50 ? g/ml DNase I at 37°C. The remainingcells were centrifuged over a discontinuous Percoll gradient(1.01, 1.03, 1.05, 1.07, 1.10 g/ml), and cells at the 1.01/1.03,1.03/1.05 and 1.05/1.07 interfaces collected. These DC-enriched cells were stained with mAb N418-FITC (hamsteranti-mouse CD11c; PharMingen) and sorted into CD11c+
and CD11c– populations using a FACStar Plus (Becton Dick-inson) flow cytometer. Immediately after sorting, cells weretitrated in wells for T cell assays. Flow cytometric analysesof sorted populations also were performed immediately aftersorting to determine the purity of the sort and the surfaceexpression levels of MHC class II, CD80 and CD86. TheCD11c+ DC were CDllb+, MHC class II+ (mean fluorescencechannel 1205) and CD80lo, CD86–. The CD11c– populationwas composed of 70% B220+, 20% CD3+, and 4% CDllb+
cells. The CD11c–, B220+ B cells were MHC class II+ (meanfluorescence channel 750) and CD80/86–. The CDllc–,CD11b+ cells were MHC class II–. A portion of the CD11c–
cells was activated with bacterial LPS for 48 h prior to incu-bation with T cells; LPS activated cells were 95% B220+,MHC class II+ (mean fluorescence channel 800) andexpressed high levels of CD80 and CD86.
Thymic APC, tDC and cTEC, were isolated from 3–4-week-old B6 mice by digestion of thymic fragments with collage-nase/dispase/DNase I followed by a Percoll gradient asdescribed above; the thymocyte-depleted cell suspensionwas stained with FITC-conjugated N418 and PE-conjugatedBP-1 mAb (PharMingen) and sorted into tDC (N418+, BP-1–)and cTEC (N418–, BP-1+) populations.
Acknowledgements: We thank Susan Eastman for experttechnical assistance and acknowledge the involvement ofCatherine Grubin at initial steps of these studies. We arethankful to Dr. L. Bikoff for Ii–/– mice, Dr. L. Van Kaer for M–/–
mice, Dr. E. Clark for CD22–/– mice and Dr. A. Farr for mAb.This work was supported by Howard Hughes Medical Insti-tute and a NIH grant (A.R.), NIH NRSA (S.K.), NSF STC grant(J.R.Y.).
References
1 Auchincloss, Jr., H. and Sultan, H., Antigen processing and pre-sentation in transplantation. Curr. Opin. Immunol. 1996. 8:681–687.
2 Jardetzky, T. S., Lane, W. S., Robinson, R. A., Madden, D. R.and Wiley, D. C., Identification of self peptides bound to purifiedHLA-B27. Nature 1991. 353: 326–329.
Eur. J. Immunol. 2001. 31: 1485–1494 In vivo MHC class II presentation of cytosolic proteins 1493

3 Rudensky, A. Y., Preston-Hurlburt, P., Hong, S. C., Barlow, A.,and Janeway, Jr., C. A., Sequence analysis of peptides bound toMHC class II molecules. Nature 1991. 353: 622–627.
4 Chicz, R. M., Urban, R. G., Gorga, J. C., Stern, L. J., Vignali, D.A. A. and Strominger, J. L., Predominant naturally processedpeptides bound to HLA-DR1 are derived from MHC-related mole-cules and are heterogeneous in size. Nature 1992. 358: 764–768.
5 Nelson, C., Roof, R. W., McCourt, D. W. and Unanue, E. R.,Identification of the naturally processed form of the hen egg whitelysozyme bound to the murine major histocompatibility complexclass II molecule I-Ak. Proc. Natl. Acad. Sci. USA 1992. 89:7380–7384.
6 Newcomb, J. R. and Cresswell, P., Characterization of endoge-nous peptides bound to purified HLS-DR molecules and theirabsence from invariant chain-associated § g dimers. J. Immunol.1993. 150: 499–506.
7 Hunt, D. F., Henderson, R. A., Shabanowitz, J., Sakaguchi, K.,Michel, H., Sevilir, N., Cox, A. L., Appella, E. and Engelhard,V. H., Characterization of peptides bound to class I MHC mole-cule HLA-A2. 1 by mass spectrometry. Science 1992. 255:1261–1264.
8 Hunt, D. F., Michel, H., Dickinson, T. A., Shabanowitz, J., Cox,A. L., Sakaguchi, K., Apella, E., Grey, H. M. and Sette, A., Pep-tides presented to the immune system by the murine class IImajor histocompatibility molecule I-Ad. Science 1992. 256:1817–1820.
9 Eng, J., McCormack, A. L. and Yates, III, J. R., An approach tocorrelate MS/MS data to amino acid sequences in a protein data-base. J. Am. Soc. Mass Spec. 1994. 5: 976–985.
10 Yates, III, J. R., Eng, J. K., McCormack, A. L. and Schieltz, D.,Method to correlate tandem mass spectra of modified peptidesto amino acid sequences in the protein database. Anal. Chem.1995. 67: 1426–1436.
11 Wang, H., Harrison-Shostak, D. C., Lemasters, J. J. and Her-man, B., Cloning of a rat cDNA encoding a novel LIM domainprotein with high homology to rat RIL. Gene 1995. 165: 267–271.
12 Chicz, R. M., Urban, R. G., Gorga, J. C., Vignali, D. A. A., Lane,W. S. and Strominger, J. L., Specificity and promiscuity amongnaturally processed peptides bound to HLA-DR alleles. J. Exp.Med. 1993. 178: 27–47.
13 Harris, P. E., Maffei, A., Colovai, A. I., Kinne, J., Tugulea, S.and Suciu-Foca, N., Predominant HLA-class II bound self-peptides of a hematopoietic progenitor cell line are derived fromintracellular proteins. Blood 1996. 87: 5104–5112.
14 Jin, Y., Shih, W. K. and Berkower, I., Human T cell response tothe surface antigen of hepatitis B virus (HBsAg): endosomal andnonendosomal processing pathways are accessible to bothendogenous and exogenous antigen. J. Exp. Med. 1988. 168:293–306.
15 Jacobson, S., Sekaly, R. P., Jacobson, C. L., McFarland, H. F.and Long, E. O., HLA class II-restricted presentation of cytoplas-mic measles virus antigens to cytotoxic T cells. J. Virol. 1989. 63:1756–1762.
16 Nuchtern, J. G., Biddison, W. and Klausner, R. D., Class II MHCmolecules can use the endogenous pathway of antigen presenta-tion. Nature 1990. 343: 74–77.
17 Malnati, M., Marti, M., LeVaute, T., Jaraquemada, D., Biddi-son, W., DeMars, R. and Long, E. O., Processing pathways forpresentation of cytosolic antigen to MHC class II-restricted cells.Nature 1992. 357: 702–705.
18 Oxenius, A., Bachmann, M. F., Ashton-Rickardt, P. G, Tone-gawa, S., Zinkernagel, R. M. and Hengartner, H., Presentationof endogenous viral proteins in association with major histocom-
patibility complex class II: on the role of intracellular compart-mentalization, invariant chain and the TAP transporter system.Eur. J. Immunol. 1995. 25: 3402–3411.
19 Oxenius, A., Bachmann, M. F., Mathis, D., Benoist, C., Zinker-nagel, R. M. and Hengartner, H., Functional in vivo MHC class IIloading by endogenously synthesized glycoprotein during viralinfection. J. Immunol. 1997. 158: 5717–5725.
20 Kovats, S., Grubin, C. E., Eastman, S. E., deRoos, P. C.,Dongre, A., Van Kaer, L. and Rudensky, A. Y., Invariant chain-independent function of H-2M in the formation of endogenouspeptide-major histocompatibility complex class II complexes invivo. J. Exp. Med. 1998. 187: 245–250.
21 Martin, W. D., Hicks, G. G., Mendiratta, S. K., Leva, H. I., Ruley,H. E. and Van Kaer, L., H-2M mutant mice are defective in thepeptide loading of class II molecules, antigen presentation, and Tcell repertoire selection. Cell 1996. 84: 543–551.
22 Grubin, C. E., Kovats, S., deRoos, P. C. and Rudensky, A. Y.,Deficient positive selection of CD4 T cells in mice displayingaltered repertoires of MHC class II-bound self-peptides. Immu-nity 1997. 7: 197–208.
23 Dunn, W. A., Studies on the mechanisms of autophagy: for-mation of the autophagic vacuole. J. Cell Biol. 1990. 110:1923–1931.
24 Ahlberg, J., Marzella L. and Glaumann, H., Uptake and degra-dation of proteins by isolated rat liver lysosomes: suggestion of amicroautophagic pathway of proteolysis. Lab. Invest. 1982. 47:523–532.
25 Cuervo, A. M. and Dice, J. F., A receptor for the selective uptakeand degradation of proteins by lysosomes. Science 1996. 273:501–504.
26 Kovacsovics-Bankowski, M. and Rock, K. L., A phagosome-to-cytosol pathway for exogenous antigens presented on MHCclass I molecules. Science 1995. 267: 243–246.
27 Reis e Sousa, C. and Germain, R. N., Major histocompatibilitycomplex class I presentation of peptides derived from solubleexogenous antigen by a subset of cells engaged in phagocytosis.J. Exp. Med 1995. 182: 841–851.
28 Norbury, C. C., Hewlett, L. J., Prescott, A. R., Shastri N., andWatts, C., Class I MHC presentation of exogenous soluble anti-gen via macropinocytosis in bone marrow macrophages. Immu-nity 1995. 3: 783–791.
29 Eastman, S., Deftos, M., deRoos, P. C., Hsu, D. H., Teyton, L.,Braunstein, N. S., Hackett, C. J. and Rudensky, A. Y., A studyof complexes of class II invariant chain peptide: major histocom-patibility complex class II molecules using a new complex-specific monoclonal antibody. Eur. J. Immunol. 1996. 26:385–393.
30 Otipoby, K. L., Andersson, K. B., Draves, K. E., Klaus, S. J.,Farr, A. G., Kerner, J. D., Perlmutter, R. M., Law, C. L. andClark, E. A., CD22 regulates thymus-independent responses andthe lifespan of B cells. Nature 1996. 384: 634–637.
Correspondence: Alexander Rudensky, HHMI, Box357370, University of Washington School of Medicine, HSBI-604I, Seattle, WA 98195, USAFax: +1-206-685-3612e-mail: aruden — u.washington.edu
A. R. Dongre’s present address: Department of AppliedGenomics, Bristol Myers-Squibb PRI, Pennington Rocky HillRoad, Pennington, NJ 08534, USA
1494 A. R. Dongre et al. Eur. J. Immunol. 2001. 31: 1485–1494
Copyright © 2022 FDOKUMEN




















