
In vitro optimization of retinoic acid–induced neuritogenesis and TH endogenous expression in...
10
In vitro optimization of retinoic acid–induced neuritogenesis and TH endogenous expression in human SH-SY5Y neuroblastoma cells by the antioxidant Trolox Mario Luiz Conte da Frota Junior • Andre ´ Simo ˜es Pires • Fares Zeida ´n-Chulia ´ • Ivi Juliana Bristot • Fernanda M. Lopes • Matheus Augusto de Bittencourt Pasquali • Alfeu Zanotto-Filho • Guilherme Anto ˆnio Behr • Fabio Klamt • Daniel Pens Gelain • Jose ´ Cla ´udio Fonseca Moreira Received: 26 April 2011 / Accepted: 29 June 2011 / Published online: 7 July 2011 Ó Springer Science+Business Media, LLC. 2011 Abstract Though, it is quite well-known how retinoic acid (RA) is able to induce neuritogenesis in different in vitro models, the putative role exerted by reactive oxygen species (ROS) during this process still need to be further studied. For such purpose, we used a neuronal-like cell line (SH-SY5Y cells) in order to investigate whether the anti- oxidant Trolox (a hydrophilic analog of alpha-tocopherol) could have any effect on the number of RA-induced neu- rites, and how significant changes in cellular redox homeostasis may affect the cellular endogenous expression of tyrosine hydroxylase (TH). Our results show a signifi- cant enhancement of RA (10 lM)-induced neuritogenesis and TH endogenous expression, when cells were co-treated with Trolox (100 lM) for 7 days. Moreover, this effect was associated with an improvement in cellular viability. The mechanism seems to mainly involve PI3 K/Akt rather than MEK signaling pathway. Therefore, our data dem- onstrate that concomitant decreases in basal reactive oxy- gen species (ROS) production could exert a positive effect on the neuritogenic process of RA-treated SH-SY5Y cells. Keywords SH-SY5Y cells Á Retinoic acid Á Trolox Á Neurite outgrowth Á Reactive oxygen species Á Parkinson’s disease Abbreviations RA Retinoic acid ROS Reactive oxygen species TH Tyrosine hydroxylase PD Parkinson’s disease CAT Catalase SOD Superoxide dismutase GPx Glutathione peroxidase hESCs Human embryonic stem cells iPS cells Induced pluripotent stem cells MSCs Mesenchymal stem cells Introduction Parkinson’s disease (PD) is a neurodegenerative disorder of the central nervous system (CNS), caused by chronic loss of dopamine-producing neurons in the substantia nigra pars compacta and subsequent depletion of dopamine in the striatum, the main projection area of the substantia nigra [1, 2]. Over the last decades, cell transplantation has become an alternative approach for the treatment of neurological diseases, including PD [3]. In fact, successful in vitro generation of dopaminergic neurons from human embry- onic stem cells (hESCs) [4], induced pluripotent stem cells (iPS cells) [5, 6], and some types of mesenchymal stem cells (MSCs) [7, 8] has raised even more expectations when it comes to the use of cell-based therapy for PD. Either the use of MSCs or reprogramming fibroblasts into iPS cells potentially solve the ethical issues associated with hESCs [9], but researchers in the field are still concerned about the potential risk of tumor generation that accom- panies the transplantation of these cells in clinical trials [10]. Nevertheless, the use of different in vitro models M. L. C. d. Frota Junior (&) Á A. S. Pires Á F. Zeida ´n-Chulia ´ Á I. J. Bristot Á F. M. Lopes Á M. A. de Bittencourt Pasquali Á A. Zanotto-Filho Á G. A. Behr Á F. Klamt Á D. P. Gelain Á J. C. F. Moreira Center of Oxidative Stress Research (CEEO), Department of Biochemistry, Institute of Basic Health Sciences (ICBS), Federal University of Rio Grande do Sul (UFRGS), Rio Grande do Sul, Rua Ramiro Barcelos 2600—ANEXO, Porto Alegre, RS 90035-003, Brazil e-mail: [email protected] 123 Mol Cell Biochem (2011) 358:325–334 DOI 10.1007/s11010-011-0983-2
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of In vitro optimization of retinoic acid–induced neuritogenesis and TH endogenous expression in...

In vitro optimization of retinoic acid–induced neuritogenesisand TH endogenous expression in human SH-SY5Yneuroblastoma cells by the antioxidant Trolox
Mario Luiz Conte da Frota Junior • Andre Simoes Pires • Fares Zeidan-Chulia •
Ivi Juliana Bristot • Fernanda M. Lopes • Matheus Augusto de Bittencourt Pasquali •
Alfeu Zanotto-Filho • Guilherme Antonio Behr • Fabio Klamt • Daniel Pens Gelain •
Jose Claudio Fonseca Moreira
Received: 26 April 2011 / Accepted: 29 June 2011 / Published online: 7 July 2011
� Springer Science+Business Media, LLC. 2011
Abstract Though, it is quite well-known how retinoic
acid (RA) is able to induce neuritogenesis in different in
vitro models, the putative role exerted by reactive oxygen
species (ROS) during this process still need to be further
studied. For such purpose, we used a neuronal-like cell line
(SH-SY5Y cells) in order to investigate whether the anti-
oxidant Trolox (a hydrophilic analog of alpha-tocopherol)
could have any effect on the number of RA-induced neu-
rites, and how significant changes in cellular redox
homeostasis may affect the cellular endogenous expression
of tyrosine hydroxylase (TH). Our results show a signifi-
cant enhancement of RA (10 lM)-induced neuritogenesis
and TH endogenous expression, when cells were co-treated
with Trolox (100 lM) for 7 days. Moreover, this effect
was associated with an improvement in cellular viability.
The mechanism seems to mainly involve PI3 K/Akt rather
than MEK signaling pathway. Therefore, our data dem-
onstrate that concomitant decreases in basal reactive oxy-
gen species (ROS) production could exert a positive effect
on the neuritogenic process of RA-treated SH-SY5Y cells.
Keywords SH-SY5Y cells � Retinoic acid � Trolox �Neurite outgrowth � Reactive oxygen species �Parkinson’s disease
Abbreviations
RA Retinoic acid
ROS Reactive oxygen species
TH Tyrosine hydroxylase
PD Parkinson’s disease
CAT Catalase
SOD Superoxide dismutase
GPx Glutathione peroxidase
hESCs Human embryonic stem cells
iPS cells Induced pluripotent stem cells
MSCs Mesenchymal stem cells
Introduction
Parkinson’s disease (PD) is a neurodegenerative disorder of
the central nervous system (CNS), caused by chronic loss
of dopamine-producing neurons in the substantia nigra pars
compacta and subsequent depletion of dopamine in the
striatum, the main projection area of the substantia nigra [1,
2]. Over the last decades, cell transplantation has become
an alternative approach for the treatment of neurological
diseases, including PD [3]. In fact, successful in vitro
generation of dopaminergic neurons from human embry-
onic stem cells (hESCs) [4], induced pluripotent stem cells
(iPS cells) [5, 6], and some types of mesenchymal stem
cells (MSCs) [7, 8] has raised even more expectations
when it comes to the use of cell-based therapy for PD.
Either the use of MSCs or reprogramming fibroblasts into
iPS cells potentially solve the ethical issues associated with
hESCs [9], but researchers in the field are still concerned
about the potential risk of tumor generation that accom-
panies the transplantation of these cells in clinical trials
[10]. Nevertheless, the use of different in vitro models
M. L. C. d. Frota Junior (&) � A. S. Pires � F. Zeidan-Chulia �I. J. Bristot � F. M. Lopes � M. A. de Bittencourt Pasquali �A. Zanotto-Filho � G. A. Behr � F. Klamt �D. P. Gelain � J. C. F. Moreira
Center of Oxidative Stress Research (CEEO), Department
of Biochemistry, Institute of Basic Health Sciences (ICBS),
Federal University of Rio Grande do Sul (UFRGS), Rio Grande
do Sul, Rua Ramiro Barcelos 2600—ANEXO, Porto Alegre,
RS 90035-003, Brazil
e-mail: [email protected]
123
Mol Cell Biochem (2011) 358:325–334
DOI 10.1007/s11010-011-0983-2

(e.g., human SH-SY5Y cells), for better comprehension of
the mechanisms by which dopaminergic differentiation is
promoted, might contribute to the design of novel thera-
peutic strategies, or at least, to improve the already existing
ones.
Human SH-SY5Y cells are a neuroblastic subclone of
the neuroblastoma cell line SK-N-SH [11], and despite
their tumoral origin, they are able to display a functional
and morphological neuronal phenotype upon in vitro
exposure to several agents [12–16], such as retinoic acid
(RA), an active metabolite of retinol [17]. The biological
effects of RA are mediated by specific nuclear receptors,
which function as transcriptional regulators and act by
modifying the transcriptional activity of specific genes [18,
19]. However, the wide spectrum of physiological and
pharmacological effects of retinoids is attributed to both
receptor-dependent and receptor-independent mechanisms
[20]. In this context, previous works have already dem-
onstrated that RA is able to generate reactive oxygen
species (ROS) in several cells types—an effect coupled to
differentiation [21–24].
Although, potential activation of signaling pathways
upon RA stimulation have extensively been studied in the
process of neuritogenesis, to the best of our knowledge, a
possible link between cellular redox status and the acqui-
sition of both morphological and functional neuronal
characteristics still remains unclear. Thus, we herein used
human neuroblastoma SH-SY5Y cells as a neuronal-like
model to study this possible relation. Specifically, we
treated the cells with 10 lM of RA for 7 days, either in the
presence or absence of the antioxidant Trolox (100 lM),
MEK inhibitor UO126 (10 lM), or PI3 K inhibitor wort-
mannin (500 nM). Our results suggest that concomitant
decreases in basal ROS production can potentiate RA-
induced effects on neurite development in SH-SY5Y cells,
associated with a higher viability and reduced cytotoxicity
of the culture. Furthermore, our in vitro results suggest that
a coordinated regulation of ROS production and activation
of PI3 K signaling pathway could have a significant posi-
tive effect on RA-derived neuritogenesis in this experi-
mental model.
Materials and methods
Cell culture and treatments
Human neuroblastoma SH-SY5Y cells were obtained from
the American Type Culture Collection (ATCC, Rockville,
MD) and were previously maintained in 75 cm2 flasks in
DMEN:F12 (1:1) supplemented with 10% heat-inactivated
fetal bovine serum, L-glutamine (2 mM), and 0.28 mg/ml
of gentamycin sulfate in a 5% CO2 humidified incubator at
37�C. For Western blot analysis, evaluation of neurito-
genesis and flow cytometer analysis, cells were grown in
6-well cluster dishes, whereas cell viability and ROS pro-
duction assays were performed by using 96-well plates.
Cells for drug treatment were initially seeded at 1 9 104
cells/well (96-well plates) or 2 9 105 cells/well (6-well
plates) and cultured for 24 h. Later, concentration of fetal
bovine serum (FBS) was reduced to 1% in the culture
medium, RA alone (10 lM) or RA plus Trolox 100 lM
were added, and the cells were left for 1 week in the
presence or absence of MEK inhibitor UO126 (10 lM) or
PI 3-K inhibitor wortmannin (500 nM). The culture med-
ium was replaced every 3 days. To reveal the effects of the
kinase inhibitors on RA-induced neurite formation, cul-
tures were pretreated for 45 min before addition of RA and
Trolox.
Cell morphology analysis and evaluation
of neuritogenesis
To evaluate the differences in cell morphology, cultures
were observed in an inverted microscope (Nikon Eclipse
TE300) and images were captured by using a digital
camera connected to the microscope. Cell bodies and
neurites present in 10 randomly selected fields were
counted. The ratio between cell bodies and neurites was
calculated yielding the average of neurites per neuron and
was expressed as the mean ± SEM value. Differences
were considered to be significant when P \ 0.05;
Immunoblotting
Proteins (20 lg) were separated by SDS–PAGE—10% (w/
v) acrylamide, 0.275% (w/v) bisacrylamide gels—and
electrotransferred onto nitrocellulose membranes. Mem-
branes were then incubated in Tris-buffered saline Tween-
20 [TBS-T; 20 mM Tris–HCl, pH 7.5, 137 mM NaCl,
0.05% (v/v) Tween 20] containing 1% (w/v) non-fat milk
powder for 1 h at room temperature. Subsequently, the
membranes were incubated for 12 h with polyclonal rabbit
anti-TH (1:10.000 dilution; Sigma). After washing in TBS-
T, blots were incubated with horseradish peroxidase-linked
anti-immunoglobulin G (IgG) antibodies (1:10.000 dilu-
tion) for 1.5 h at room temperature. Chemiluminescent
bands were detected, and densitometric analysis was per-
formed by Image-J� software;
ROS production
Intracellular ROS production was determined by the
DCFH-DA-based real-time assay using intact living cells
[25]. Briefly, SH-SY5Y cells were initially seeded onto
326 Mol Cell Biochem (2011) 358:325–334
123

96-well plates and incubated for 24 h. Later, concentration
of fetal bovine serum (FBS) was reduced to 1% in the
culture medium, and cells were incubated for 1 h with
DCFH-DA 100 lM (stock solution in DMSO, 10 mM) at
5% CO2 and 37�C. Then, cells were washed, and treat-
ments were carried out. During the treatment, changes in
fluorescence by the oxidation of DCFH into the fluorogen
DCF were monitored in a microplate fluorescence reader
(F2000, Hitachi Ltd., Tokyo, Japan) at 37�C. H2O2 1 mM
was used as positive control for ROS production (data not
shown). Excitation filter was set at 485 ± 10 nm, and the
emission filter was set at 530 ± 12.5 nm. Data were
recorded every 30 s and plotted in Excel software.
Total reactive antioxidant potential (TRAP assay)
The antioxidant potential of cells was estimated by the total
reactive antioxidant potential parameter (TRAP), which
determines the non-enzymatic antioxidant potential of the
cell. Briefly, the reaction was initiated by adding luminol
(4 mM)—as an external probe to monitoring radical pro-
duction—and AAPH (10 mM)—a free radical source that
produces peroxyl radical at a constant rate—in glycine
buffer (0.1 M) pH 8.6 at room temperature that resulted in
steady luminescence emission. Afterward, SH-SY5Y cell
samples were suspended in glycine buffer and mixed into
the reaction vial. The decrease in luminescence, which is
proportional to the non-enzymatic antioxidant potential,
was monitored in a liquid scintillation counter (Wallace
1409) as counts per minute (CPM). The luminescence
emission was followed for 30 min after the addition of
sample homogenates (150 lg of protein). Chemilumines-
cence values were standardized against protein content.
Flow cytometry analysis
SH-SY5Y cells were initially seeded onto 6-well plates and
incubated for 24 h. Later, concentration of fetal bovine
serum (FBS) was reduced to 1% in the culture medium, and
cells were treated for 7 days. At the end of treatment, cells
were washed twice in PBS (137 mM NaCl, 2.7 mM KCl,
4.3 mM Na2HPO4.2H2O, 1.4 mM KH2PO4, pH 7.4) con-
taining 1 mM EDTA (PBS-EDTA) and subsequently
trypsinized with 0.13 g/L trypsin in PBS-EDTA. Trypsin
was inhibited with bovine fetal serum 10%, and medium
washes, and cells were combined and centrifuged (5 min,
200g, 4�C). Cells were allowed to recover from trypsin-
ization in complete medium (30 min, 37�C). Propidium
iodide (2 lM) was added 10 min prior to analysis. PI is
excluded from healthy cells, but following loss of mem-
brane integrity, enters the cells, binds to DNA, and
becomes highly fluorescent. Red fluorescence of DNA-
bound PI in individual cells was measured using a
FACScalibur (BD PharMingen). Ten thousand cells were
analyzed per sample, and data were reported as the per-
centage of necrotic cells. H2O2 1 mM was used as positive
control of cell death (data not shown).
Thiobarbituric acid reactive species (TBARS)
As an index of lipid peroxidation, we used the formation of
TBARS during an acid-heating reaction as previously
described [26]. Briefly, cells were harvested at the end of
differentiation, the samples were mixed with 1 ml of tri-
chloroacetic acid 10% (TCA) and 1 ml of thiobarbituric
acid 0.67% (TBA), then heated in a boiling water bath for
15 min. TBARS were determined by the absorbance at
535 nm and were expressed as TBARS/mg protein.
Antioxidant enzyme activities
To determine CAT activity, cells were sonicated in 50 mM
phosphate buffer (pH 7.0) and the resulting suspension was
centrifuged at 3,000 g for 10 min. The supernatant was
used for enzyme assays. CAT activity was assayed by
measuring the rate of decrease in H2O2 absorbance at
240 nm [27]. SOD activity was assayed by measuring the
inhibition of adrenaline auto-oxidation [28]. For GPx
activity, NADPH oxidation was followed at 340 nm in the
presence of reduced glutathione, tert-butyl hydroperoxide,
and glutathione reductase [29].
Protein determination
For immunoblotting analysis, the protein contents were
measured by the Bradford assay [30]. For TBARS and
antioxidant enzyme assays, protein content of each sample
was measured by Lowry method [31].
Statistical analysis
Results were expressed as the mean ± SEM of at least
three independent experiments. Data were analyzed by a
one-way analysis of variance (ANOVA), using a Newman–
Keuls test to compare mean values across groups. Differ-
ences were considered to be significant when P \ 0.05.
Results
SH-SY5Y cells exposed to 10 lM RA assume a neuronal-
like phenotype by activating neurite production and the
expression of neuronal-specific markers [32]. First, we
evaluated the possible morphological changes in SH-SY5Y
cells when treated with 10 lM RA, in the presence or
absence of the antioxidant Trolox (100 lM). As expected,
Mol Cell Biochem (2011) 358:325–334 327
123

cells clearly underwent neurite-like development and
acquire neuronal-like morphology after 7 days of treatment
with RA (Fig. 1). Interestingly, in the presence of Trolox,
the number of viable cells in culture seems to be higher and
thus, positively affecting neuritogenesis in RA-treated cells
(Fig. 1).
In order to determine whether the presence of Trolox
could have some kind of effect on RA-induced neurite-like
development, we quantified the number of neurites per cell
body in each case. The results show a significant increase
in the number of neurites per cell, when cultures were
co-treated with both RA and Trolox in comparison with
those ones exposed to RA alone (Fig. 2a). Since tyrosine
hydroxylase (TH, EC1.14.16.2) is the rate-limiting enzyme
in the catecholamine synthesis pathway, and its regulation
controls the levels of dopamine, epinephrine, and norepi-
nephrine in tissues [33], we performed immunoblotting
experiments to quantify possible variations at the protein
level of the enzyme. We found that TH immunocontent,
a classical biochemical marker of catecholaminergic
differentiation, was significantly increased in cells treated
with RA plus Trolox when compared to RA-treated cells
and control cells (Fig. 2b).
These first results seemed to point out that ROS pro-
duction may have a possible role in the effectiveness of
RA-induced neurite production. So then, we decided to
investigate whether RA is able to induce ROS production
in our cell model. To achieve this goal, an in vitro real-time
DCFH-DA assay was performed showing that ROS pro-
duction in RA-treated cells was not increased in compari-
son with control cells. However, co-treatment with 100 lM
Trolox significantly decreased the basal levels of free
radical generation, observed under our assays conditions
(Fig. 3a). Next, we conducted parallel experiments for
evaluating possible changes in the cellular redox status
after each different treatment. Figure 3b shows that expo-
sure to RA for 1 week was able to increase the values
obtained by TRAP assay and measured by chemilumines-
cence-enhanced method, all together indicating that RA
treatment for 1 week was pro-oxidant, indeed. Again,
(200 X)(100 X)
Co
ntr
ol
RA
RA
+ T
rolo
x
Fig. 1 Phase-contrast
micrographs of SH-SY5Y cells.
Control, control cells; RA, cells
treated with RA 10 lM for
7 days; RA ? Trolox, cells
treated with RA plus Trolox
100 lM for 7 days. The
medium was replaced every
3 days. Scale bar, 100 lm
328 Mol Cell Biochem (2011) 358:325–334
123
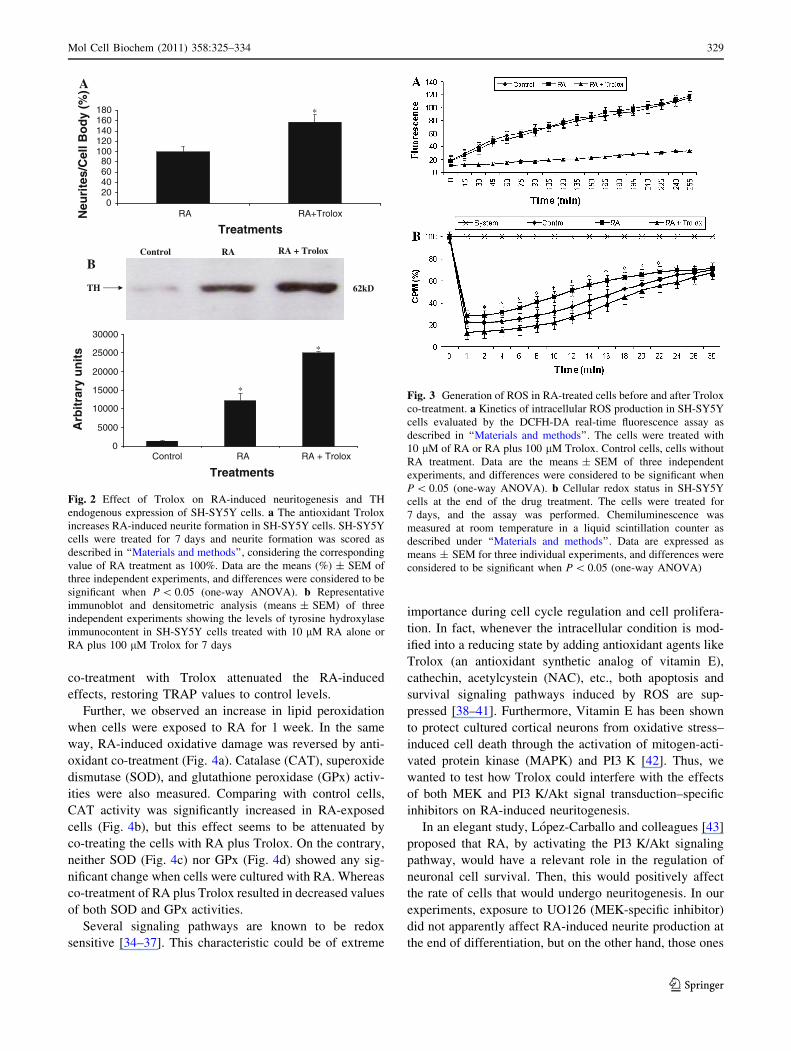
co-treatment with Trolox attenuated the RA-induced
effects, restoring TRAP values to control levels.
Further, we observed an increase in lipid peroxidation
when cells were exposed to RA for 1 week. In the same
way, RA-induced oxidative damage was reversed by anti-
oxidant co-treatment (Fig. 4a). Catalase (CAT), superoxide
dismutase (SOD), and glutathione peroxidase (GPx) activ-
ities were also measured. Comparing with control cells,
CAT activity was significantly increased in RA-exposed
cells (Fig. 4b), but this effect seems to be attenuated by
co-treating the cells with RA plus Trolox. On the contrary,
neither SOD (Fig. 4c) nor GPx (Fig. 4d) showed any sig-
nificant change when cells were cultured with RA. Whereas
co-treatment of RA plus Trolox resulted in decreased values
of both SOD and GPx activities.
Several signaling pathways are known to be redox
sensitive [34–37]. This characteristic could be of extreme
importance during cell cycle regulation and cell prolifera-
tion. In fact, whenever the intracellular condition is mod-
ified into a reducing state by adding antioxidant agents like
Trolox (an antioxidant synthetic analog of vitamin E),
cathechin, acetylcystein (NAC), etc., both apoptosis and
survival signaling pathways induced by ROS are sup-
pressed [38–41]. Furthermore, Vitamin E has been shown
to protect cultured cortical neurons from oxidative stress–
induced cell death through the activation of mitogen-acti-
vated protein kinase (MAPK) and PI3 K [42]. Thus, we
wanted to test how Trolox could interfere with the effects
of both MEK and PI3 K/Akt signal transduction–specific
inhibitors on RA-induced neuritogenesis.
In an elegant study, Lopez-Carballo and colleagues [43]
proposed that RA, by activating the PI3 K/Akt signaling
pathway, would have a relevant role in the regulation of
neuronal cell survival. Then, this would positively affect
the rate of cells that would undergo neuritogenesis. In our
experiments, exposure to UO126 (MEK-specific inhibitor)
did not apparently affect RA-induced neurite production at
the end of differentiation, but on the other hand, those ones
A
020406080
100120140160180
RA+TroloxRA
Treatments
Neu
rite
s/C
ell B
od
y (%
)
B
*
0
5000
10000
15000
20000
25000
30000
Control RA RA + Trolox
Treatments
Arb
itra
ry u
nit
s
TH
Control RA
62kD
*
*
RA + Trolox
Fig. 2 Effect of Trolox on RA-induced neuritogenesis and TH
endogenous expression of SH-SY5Y cells. a The antioxidant Trolox
increases RA-induced neurite formation in SH-SY5Y cells. SH-SY5Y
cells were treated for 7 days and neurite formation was scored as
described in ‘‘Materials and methods’’, considering the corresponding
value of RA treatment as 100%. Data are the means (%) ± SEM of
three independent experiments, and differences were considered to be
significant when P \ 0.05 (one-way ANOVA). b Representative
immunoblot and densitometric analysis (means ± SEM) of three
independent experiments showing the levels of tyrosine hydroxylase
immunocontent in SH-SY5Y cells treated with 10 lM RA alone or
RA plus 100 lM Trolox for 7 days
Fig. 3 Generation of ROS in RA-treated cells before and after Trolox
co-treatment. a Kinetics of intracellular ROS production in SH-SY5Y
cells evaluated by the DCFH-DA real-time fluorescence assay as
described in ‘‘Materials and methods’’. The cells were treated with
10 lM of RA or RA plus 100 lM Trolox. Control cells, cells without
RA treatment. Data are the means ± SEM of three independent
experiments, and differences were considered to be significant when
P \ 0.05 (one-way ANOVA). b Cellular redox status in SH-SY5Y
cells at the end of the drug treatment. The cells were treated for
7 days, and the assay was performed. Chemiluminescence was
measured at room temperature in a liquid scintillation counter as
described under ‘‘Materials and methods’’. Data are expressed as
means ± SEM for three individual experiments, and differences were
considered to be significant when P \ 0.05 (one-way ANOVA)
Mol Cell Biochem (2011) 358:325–334 329
123

treated with wortmannin (PI3 K-/Akt-specific inhibitor)
showed an apparent reduction in cell viability and there-
fore, impaired the process of RA-induced neuritogenesis,
as assessed by morphological criteria (Fig. 5a). On the
contrary, when cells were co-treated with RA, wortmannin
and the antioxidant Trolox, we noticed a clear rescue of the
previously observed wortmannin-derived inhibitory effects
on cell viability and thus, the number of cell undergoing
neurite-like development (morphological criteria)
(Fig. 5a). In order to support these observations, we per-
formed further analysis of cell-culture viability by flow
cytometry after 7 days, showing that cells treated with
RA and wortmannin have a significant reduction in cell
viability compared with cells treated with RA alone, as
expected (Fig. 5b). On the other hand, when cells were
co-treated with RA, wortmanin, and Trolox, cell viability
was rescued when comparing to RA/wortmannin-treated
cultures. Moreover, PI incorporation was higher in cells
treated with 10 lM RA than that in control cells, sug-
gesting that RA caused a loss of membrane integrity. Such
effect was abolished by the addition of Trolox.
Discussion
Parkinson’s disease (PD) is a neurodegenerative disease
with no definitive cure so far [44], which is pathologically
characterized by chronic loss of dopamine-producing
neurons in the substantia nigra pars compacta [1, 2]. Cell
transplantation for replacing the damaged neurons repre-
sents a promising approach to restore tissue functionality in
different neurodegenerative diseases, including PD. How-
ever, many are the obstacles to overcome, such as ethical
concerns, selection of the best viable model for cell
transplantation or how to induce and control the differen-
tiation of these cells into specific cellular phenotypes; in
the case of PD, dopamine-producing cells.
SH-SY5Y cells respond to RA and give rise to neuronal-
like cells, acquiring morphological and neurochemical
properties characteristic of neurons. In that sense, they may
represent an available and optimal model to determine
what kind of factors affect biochemical routes leading to
neuronal differentiation in vitro, and whether they might
potentially be beneficial in future clinical trials. In general,
promotion of cellular differentiation implies a complex
re-arrangement of intrinsic cellular programs, cell-to-cell
interaction and the effect of diverse growth factors, hor-
mones as well as other signaling molecules. As a matter of
fact, it is well-known the crucial importance RA during
early embryonic development and organogenesis [45–47];
but the possible influence of ROS production during these
functions still needs further studies.
In the present work, we examined the effect of
the antioxidant Trolox on RA-induced neuritogenesis of
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
RA+TroloxRAControl
Treatments
MD
A e
qu
ival
ents
0
0.5
1
1.5
2
2.5
RA+TroloxRAControl
Treatments
CA
T (
U/m
g p
rote
in)
02468
101214
Control RA RA + Trolox
TreatmentsSO
D (
U/m
g p
rote
in)
02468
101214161820
RA+TroloxRAControl
Treatments
GP
x (U
/mg
pro
tein
)
BA
C D
*
*
**
(nm
ol/m
g p
rote
in)
Fig. 4 Lipid peroxidation and oxidative stress enzymatic profile of
RA-treated cells before and after Trolox co-treatment. a Determina-
tion of TBARS in SH-SY5Y cells at the end of the drug treatment.
Data are the means ± SEM of three independent experiments, and
differences were considered to be significant when P \ 0.05 (one-
way ANOVA). b Catalase activity in SH-SY5Y cells at the end of the
drug treatment. Data are the means ± SEM of three independent
experiments, and differences were considered to be significant when
P \ 0.05 (one-way ANOVA). c Superoxide dismutase activity in SH-
SY5Y cells at the end of the drug treatment. Data are the
means ± SEM of three independent experiments, and differences
were considered to be significant when P \ 0.05 (one-way ANOVA).
d Glutathione peroxidase activity in SH-SY5Y cells at the end of the
drug treatment. Data are the means ± SEM of three independent
experiments, and differences were considered to be significant when
P \ 0.05 (one-way ANOVA)
330 Mol Cell Biochem (2011) 358:325–334
123

SH-SY5Y cells. Our results show a significant enhance-
ment of RA (10 lM)-induced neuritogenesis and TH
endogenous expression, when cells were co-treated with
Trolox (100 lM) for 7 days.
To search for a link between cellular redox status and
the induction of neurite-like processes in SH-SY5Y cells,
we performed a real-time DCFH-DA assay. As shown in
Fig. 3a, RA did not induce ROS production when com-
pared to control cells. However, co-treatment with 100 lM
Trolox was able to significantly decrease the basal levels of
free radical generation observed under this condition.
Moreover, evaluation of total reactive antioxidant potential
in RA-treated SH-SY5Y cells by TRAP assay suggested
that RA was leading to partial consumption of cellular non-
enzymatic antioxidants (Fig. 3b). In other words, whenever
RA is inducing neuritogenesis in our cell model, Trolox
optimize this process by increasing, at least in part, the
viability of the culture.
ROS are known to activate an array of intracellular
signaling cascades that are closely associated with both cell
death and cell survival pathways [48]. On one hand, pre-
vious studies have reported that H2O2 induces apoptosis in
SH-SY5Y cells and rat pheochromocytoma PC12 cells (a
model system for catecholamine-containing neurons), but
the mechanisms involved are not fully understood [49, 50].
On the other hand, an interesting work suggested that a
transient enhancement of the intracellular ROS level plays
an important role during docosahexaenoic acid–enhanced
neurite outgrowth [51], and more recently, Franco et al.
[52] demonstrated that SH-SY5Y cells transfected with
human TH isoform 1 were substantially more resistant to
H2O2 and 6-hydroxydopamine-induced cell death, when
A
B
0
5
10
15
20
25
30
35
40
Control RA RA + Trolox RA + UO126 RA + W RA + W +Trolox
Treatments
% P
I po
siti
ve c
ells
*
*
Control RA RA + Trolox
RA + W + TroloxRA + WRA + UO126
100
X
Fig. 5 Effects of MEK and PI3 K/Akt inhibitors on RA-induced
neuritogenesis and cell viability before and after Trolox treatment.
a Phase-contrast micrographs of SH-SY5Y cells. Control, control
cells; RA, RA-treated cells (10 lM) for 7 days; RA ? Trolox,
RA-treated cells (10 lM) plus Trolox (100 lM) for 7 days;
RA ? UO126, RA-treated cells (10 lM) plus MEK inhibitor
UO126 (10 lM) for 7 days; RA ? W, RA-treated cells (10 lM)
plus PI-3 K inhibitor wortmannin (500 nM) for 7 days; RA ?
W ? Trolox, RA-treated cells (10 lM) plus wortmannin (500 nM)
plus Trolox (100 lM) for 7 days. Culture medium was replaced every
3 days. b Effects of RA and specific inhibitors of MEK and PI-3 K/
Akt signal pathways on cell viability. Red fluorescence of DNA-
bound PI in individual SH-SY5Y cells was measured using a flow
cytometer as described under ‘‘Materials and methods’’. The cells
were treated for 7 days, and the assay was performed. Data are the
means (%) ± SEM of three independent experiments, and differences
were considered to be significant when P \ 0.05 (one-way ANOVA)
Mol Cell Biochem (2011) 358:325–334 331
123

compared to proliferative neuroblastoma cells, suggesting
that a preconditioning-like mechanism linked to higher
dopamine levels increased the resistance of cells against
oxidative insults.
That is why we decided to study the effects exerted by
our in vitro treatments on the oxidative damage and anti-
oxidant enzyme activities. When neuroblastoma cells were
exposed to RA for 1 week, an increase in lipid peroxidation
was observed (Fig. 4a). The regulation of the antioxidant
enzymes seems to be important for holding the balance
between generation and consumption of ROS. For this
reason, we additionally determined possible changes in
CAT, SOD, and GPx activities. In comparison with pro-
liferative cells, CAT activity was significantly increased in
RA-treated cells (Fig. 4b). In contrast, neither SOD nor
GPx activities significantly vary. However, co-treatment
with Trolox resulted in a significant decrease in SOD
(Fig. 4c) and GPx (Fig. 4d) activities, indicating a reduc-
tion in ROS production.
We further investigated the influence of two signaling
pathways, MEK and PI3 K/Akt pathways in RA-induced
neuritogenesis (Fig. 5a) and cell viability (Fig. 5b) and in
which way co-treatment with Trolox could affect it. Both
signaling pathways are known to be important in mediating
neuronal survival and differentiation [53]. Cells treated
with RA plus wortmannin showed a decreased viability
of the culture and therefore, less efficient neuritogenic
induction when compared to RA-treated cells. However,
when cells were co-treated with RA, wortmanin, and
Trolox, we observed that those negative effects were res-
cued. UO126 did not apparently affect RA-induced neurite
formation (assessed by morphological criteria).
Some authors have reported that RA actives ERK1/2,
PKC, and PI3 K in cells where retinoid RXR/RAR recep-
tors were silenced, suggesting a RA receptor-independent
mechanism [54, 55]. Others demonstrated that RA actives
PI-3 K/Akt pathway by RA-mediated interaction of RAR
with the catalytic subunit of PI-3 K-inducing Akt phos-
phorylation [56]. However, the exact mechanism underly-
ing non-genomic actions of RA and other retinoids still
requires further studies.
In contrast to the physiological effects of RA (10-9–
10-8 M), which are primarily mediated by specific nuclear
receptors, the mechanism underlying RA toxicity upon
pharmacological exposure (10-7–10-5 M) remains unde-
fined [46], and nowadays, it is unclear whether oxidative
stress is the main mechanism of RA toxicity. Our results
suggest that a concomitant decrease in basal ROS pro-
duction can exert a positive effect on RA-induced neurite
development. Moreover, a coordinated regulation of ROS
production and PI3-K activation may play an important
role in RA-induced neuritogenesis of human SH-SY5Y
neuroblastoma cells; it is possible that Trolox-induced
antioxidant effects may counter balance the basal
RA-induced cytotoxicity, and thus, survival effects of
RA-induced phosphorylation of Akt would take place.
Thus, we believe that an antioxidant-based strategy could
avoid a premature death of cells entering in differentiation
programs, allowing the survival of newly formed TH
positive cells. Nevertheless, additional studies are required
for better comprehension of the exact mechanism by which
Trolox affects the RA-induced neurite-like process for-
mation, as well as its possible significance in the modula-
tion of PI 3-K pathway. Since optimal neuritogenesis and
survival of the newly formed neurons is a critical step of
neuronal development, we expect that our results may be
relevant to the development of new strategies involving
cellular transplantation and replacement of dopamine-pro-
ducing cells.
Acknowledgments This work was supported by CNPq, CAPES,
FAPERGS, PROPESQ-UFRGS and ‘‘Rede Instituto Brasileiro deNeurociencia’’ (IBM-Net) # 01.06.0842.00.
Conflict of interest None.
References
1. Bernheimer H, Birkmayer W, Hornykiewiez O, Jellinger K,
Seitelberger F (1973) Brain dopamine and the syndromes of
Parkinson and Huntington. Clinical, morphological and neuro-
chemical correlations. J Neurol Sci 20(4):415–455
2. Hirsch E, Graybiel AM, Agid YA (1988) Melanized dopami-
nergic neurons are differentially susceptible to degeneration in
Parkinson’s disease. Nature 334:345–348
3. Bjorklund A, Lindvall O (2000) Cell replacement therapies for
central nervous system disorders. Nat Neurosci 3:537–544
4. Perrier AL, Taba V, Barberi T, Rubio ME, Bruses J, Topf N,
Harrison NL, Studer L (2004) Derivation of midbrain dopamine
neurons from human embryonic stem cells. Proc Natl Acad Sci
USA 34:12543–12548
5. Wernig M, Zhao JP, Pruszak J, Hedlund E, Fu D, Soldner F,
Broccoli V, Constantine-Paton M, Isacson O, Jaenisch R (2008)
Neurons derived from reprogrammed fibroblasts functionally
integrate into the fetal brain and improve symptoms of rats with
Parkinson’s disease. Proc Natl Acad Sci USA 15:5856–5861
6. Swistowski A, Peng J, Liu Q, Mali P, Rao MS, Cheng L, Zeng X
(2010) Efficient generation of functional dopaminergic neurons
from human induced pluripotent stem cells under defined con-
ditions. Stem Cells 10:1893–1904
7. Trzaska KA, Kuzhikandathil EV, Rameshwar P (2007) Specifi-
cation of a dopaminergic phenotype from adult human mesen-
chymal stem cells. Stem Cells 11:2797–2808
8. Chen S, Xianwen C, Dehua X, Zhenguo L, Lingfei X, Smith SW,
Zhongcheng Z (2003) Behavioral correction of Parkinsonian rats
following the transplantation of immortalized fibroblasts geneti-
cally modified with TH and GCH genes. Parkinsonism Relat
Disord 9(2):91–97
9. Zeidan-Chulia F, Noda M (2009) ‘‘Opening’’ the mesenchymal
stem cell tool box. Eur J Dent 3:240–249
10. Arenas E (2010) Towards stem cell replacement therapies for
Parkinson’s disease. Biochem Biophys Res Commun 396(1):
152–156
332 Mol Cell Biochem (2011) 358:325–334
123

11. Ross RA, Spengler BA, Biedler JL (1983) Coordinate morpho-
logical and biochemical interconversion of human neuroblastoma
cells. J Natl Cancer Inst 71:741–747
12. Nicolini G, Miloso M, Zoia C, Di Silvestro A, Cavaletti G,
Tredici G (1998) Retinoic acid differentiated SH-SY5Y human
neuroblastoma cells: an in vitro model to assess drug neurotox-
icity. Anticancer Res 8:2477–2481
13. Pahlman S, Hoehner JC, Nanberg E, Hedborg F, Fagerstrom S,
Gestblom C, Johansson I, Larsson U, Lavenius E, Ortoft E, Soder-
holm H (1995) Differentiation and survival influences of growth
factors in human neuroblastoma. Eur J Cancer 31A:453–458
14. Kim B, Leventhal PS, Saltiel AR, Feldman EL (1997) Insulin-
like growth factor immediate neurite outgrowth in vitro requires
mitogen-activated protein kinase activation. J Biol Chem
272:21268–21273
15. Encinas M, Iglesias M, Llecha N, Comella JX (1999) Extracel-
lular-regulated kinases and phosphatidylinositol 3-kinase are
involved in brain-derived neurotrophic factor mediated survival
and neuritogenesis of the neuroblastoma cell line SH-SY5Y.
J Neurochem 73:1409–1421
16. Olsson AK, Vadhammar K, Nanberg E (2000) Activation and
protein kinase C dependent nuclear accumulation of ERK in
differentiating human neuroblastoma cells. Exp Cell Res 256:
454–467
17. Miloso M, Villa D, Crimi M, Galbiati S, Donzelli E, Nicolini G,
Tredici G (2004) Retinoic acid-induced neuritogenesis of human
neuroblastoma SH-SY5Y cells is ERK independent and PKC
dependent. J Neurosci Res 75:241–252
18. Glass CK, Rosenfeld MG (2000) The coregulator exchange in
transcriptional functions of nuclear receptors. Genes Dev 14:
121–141
19. Aranda A, Pascual A (2001) Nuclear hormone receptors and gene
expression. Physiol Rev 81:1269–1304
20. Radominska-Pandya A, Chen G, Czernik PJ, Little JM, Samo-
kyszyn VM, Carter CA, Nowak G (2000) Direct interaction of
All-trans-retinoic acid with protein kinase C (PKC): implications
for PKC signaling and cancer therapy. J Biol Chem 275:
22324–22330
21. Frota MLC, Silva EG, Behr GA, Oliveira MR, Dal-Pizzol F,
Klamt F, Moreira JCF (2006) All-trans retinoic acid induces free
radical generation and modulate antioxidant enzyme activities in
rat sertoli cells. Mol Cell Biochem 285:173–179
22. Castro-Obregon S, Covarrubias L (1996) Role of retinoic acid
and oxidative stress in embryonic stem cell death and neuronal
differentiation. FEBS Lett 381:93–97
23. Delia D, Aiello A, Meroni L, Nicolini M, Reed JC, Pierotti MA
(1997) Role of antioxidants and intracellular free radicals in re-
tinamide-induced cell death. Carcinogenesis 18:943–948
24. Furuke K, Sasada T, Ueda-Taniguchi Y, Yamauchi A, Inamoto T,
Yamaoka Y, Masutani H, Yodoi J (1997) Role of intracellular
redox status in apoptosis induction of human T-cell leukemia
virus type I- infected lymphocytes by 13 cis-retinoic acid. Cancer
Res 57:4916–4923
25. Wang H, Joseph JA (1999) Quantifying cellular oxidative stress
by dichlorofluorescein assay using microplate reader. Free Radic
Biol Med 27:612–616
26. Draper HH, Hadley M (1990) Malondialdehyde determination as
index of lipid peroxidation. Methods Enzymol 186:421–431
27. Aebi H (1984) Catalase in vitro. Methods Enzymol 105:121–126
28. Bannister JV, Calabrese L (1987) Assays for SOD. Methods
Biochem Anal 32:279–312
29. Flohe L, Gunzler WA (1984) Assays of glutathione peroxidase.
Methods Enzymol 105:114–121
30. Bradford MM (1976) A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principle of protein-dye binding. Anal Biochem 72:248–254
31. Lowry OH, Rosebrough AL, Farr AL, Randal RJ (1951) Protein
measurement with the Folin phenol reagent. J Biol Chem 193:
265–275
32. Encinas M, Iglesias M, Liu Y, Wang H, Muhaisen A, Cena V,
Gallego C, Comella JX (2000) Sequential treatment of SH-SY5Y
cells with retinoic acid and brain-derived neurotrophic factor
gives rise to fully differentiated, neurotrophic factor-dependent,
human neuron-like cells. J Neurochem 75:991–1003
33. Dunkley PR, Bobrovskaya L, Graham ME, von Nagy-Felsobuki
EI, Dickson PW (2004) Tyrosine hydroxylase phosphorylation:
regulation and consequences. J Neurochem 91:1025–1043
34. Fernandez-Checa JC (2003) Redox regulation and signaling lip-
ids in mitochondrial apoptosis. Biochem Biophys Res Commun
3:471–479
35. Pantano C, Reynaert NL, van der Vliet A, Janssen-Heininger YM
(2006) Redox-sensitive kinases of the nuclear factor-kappaB
signaling pathway. Antioxid Redox Signal 9–10:1791–1806
36. Hansen JM (2006) Oxidative stress as a mechanism of terato-
genesis. Birth Defects Res C Embryo Today 4:293–307
37. Anilkumar N, Sirker A, Shah AM (2009) Redox sensitive sig-
naling pathways in cardiac remodeling, hypertrophy and failure.
Front Biosci 14:3168–3187
38. Inanami O, Watanabe Y, Syuto B, Nakano M, Tsuji M, Kuwa-
bara M (1998) Oral administration of (-)catechin protects against
ischemia-reperfusion-induced neuronal death in the gerbil. Free
Radic Res 4:359–365
39. Inanami O, Takahashi K, Kuwabara M (1999) Attenuation of
caspase-3-dependent apoptosis by Trolox post-treatment of
X-irradiated MOLT-4 cells. Int J Radiat Biol 2:155–163
40. Niwa K, Inanami O, Yamamori T, Ohta T, Hamasu T, Kubawara
M (2003) Redox regulation of PI3K/Akt and p53 in bovine aortic
endothelial cells exposed to hydrogen peroxide. Antioxid Redox
Signal 6:713–722
41. Kuwabara M, Asanuma T, Niwa K, Inanami O (2008) Regulation
of cell survival and death signals induced by oxidative stress.
J Clin Biochem Nutr 2:51–57
42. Numakawa Y, Numakawa T, Matsumoto T, Yagasaki Y, Kuma-
maru E, Kunugi H, Taguchi T, Niki E (2006) Vitamin E protected
cultured cortical neurons from oxidative stress-induced cell death
through the activation of mitogen-activated protein kinase and
phosphatidylinositol 3-kinase. J Neurochem 4:1191–1202
43. Lopez-Carballo G, Moreno L, Masia S, Perez P, Barettino D(2002) Activation of the phosphatidylinositol 3-kinase/Akt sig-
naling pathway by retinoic acid is required for neural differen-
tiation of SH-SY5Y human neuroblastoma cells. J Biol Chem
28:25297–25304
44. Kedar NP (2003) Can we prevent Parkinson’s and Alzheimer’s
disease? J Postgrad Med 49:236–245
45. Bain G, Kitchens D, Yao M, Huettner JE, Gottlieb DI (1995)
Embryonic stem cells express neuronal properties in vitro. Dev
Biol 2:342–357
46. Ross SA, McCaffery PJ, Drager UC, De Luca LM (2000) Reti-
noids in embryonal development. Physiol Rev 80:1021–1054
47. Duester G (2008) Retinoic acid synthesis and signaling during
early organogenesis Gregg Duester. Cell 6:921–931
48. Kamata H, Hirata H (1999) Redox regulation of cellular signal-
ling. Cell Signal 11:1–14
49. Zhang L, Zhao B, Yew DT, Kusiak JW, Roth GS (1997) Pro-
cessing of Alzheimer’s amyloid precursor protein during H2O2-
induced apoptosis in human neuronal cells. Biochem Biophys
Res Commun 235:845–848
50. Yamakawa H, Ito Y, Naganawa T, Banno Y, Nakashima S,
Yoshimu S, Sawada M, Nishimura Y, Nozawa Y, Sakai N (2000)
Activation of caspase-9 and 3 during H2O2-induced apoptosis of
PC12 cells independent of ceramide formation. Neurol Res 22:
556–564
Mol Cell Biochem (2011) 358:325–334 333
123

51. Wu H, Ichikawa S, Tani C, Zhu B, Tada M, Shimoishi Y, Murata
Y, Nakamura Y (2009) Docosahexaenoic acid induces ERK1/2
activation and neuritogenesis via intracellular reactive oxygen
species production in human neuroblastoma SH-SY5Y cells.
Biochim Biophys Acta 1791:8–16
52. Franco JL, Posser T, Gordon SL, Bobrovskaya L, Schneider JJ,
Farina M, Dafre AL, Dickson PW, Dunkley PR (2010) Expres-
sion of tyrosine hydroxylase increases the resistance of human
neuroblastoma cells to oxidative insults. Toxicol Sci 113(1):
150–157
53. Kaplan DR, Stephens RM (1994) Neurotrophin signal transduc-
tion by the Trk receptor. J Neurobiol 25:1404–1417
54. Hoyos B, Imam A, Chua R, Swenson C, Tong GX, Levi E, Noy
N, Hammerling U (2000) The cysteine-rich regions of the regu-
latory domains of Raf and protein kinase C as retinoid receptors.
J Exp Med 192:835–845
55. Aggarwal S, Kim SW, Cheon K, Tabassam F, Joon JH, Koo JS
(2006) Nonclassical action of retinoic acid on the activation of
the camp response element-binding protein in normal human
bronchial epithelial cells. Mol Biol Cell 17:566–575
56. Masia M, Alvarez S, de Lera AR, Barettino D (2007) Rapid,
nongenomic actions of retinoic acid on phosphatidylinositol-3-
kinase signaling pathway mediated by the retinoic acid receptor.
Mol Endocrinol 21(10):2391–2402
334 Mol Cell Biochem (2011) 358:325–334
123




















