
Magnetic stimulation of axons in a nerve bundle: Effects of current redistribution in the bundle

Retinoic acid receptor b2 promotes functionalregeneration of sensory axons in the spinal cord
Liang-Fong Wong1,4, Ping K Yip2,4, Anna Battaglia2, John Grist2, Jonathan Corcoran3, Malcolm Maden3,Mimoun Azzouz1, Susan M Kingsman1, Alan J Kingsman1, Nicholas D Mazarakis1 & Stephen B McMahon2
The embryonic CNS readily undergoes regeneration, unlike the adult CNS, which has limited axonal repair after injury. Here
we tested the hypothesis that retinoic acid receptor b2 (RARb2), critical in development for neuronal growth, may enable adult
neurons to grow in an inhibitory environment. Overexpression of RARb2 in adult rat dorsal root ganglion cultures increased
intracellular levels of cyclic AMP and stimulated neurite outgrowth. Stable RARb2 expression in DRG neurons in vitro and in vivo
enabled their axons to regenerate across the inhibitory dorsal root entry zone and project into the gray matter of the spinal cord.
The regenerated neurons enhanced second-order neuronal activity in the spinal cord, and RARb2-treated rats showed highly
significant improvement in sensorimotor tasks. These findings show that RARb2 induces axonal regeneration programs within
injured neurons and may thus offer new therapeutic opportunities for CNS regeneration.
The survival and differentiation of developing vertebrate CNS neuronsis influenced by retinoic acid (reviewed in refs. 1 and 2). Retinoic acidcan stimulate both neurite number and length in a variety of embryonicneuronal types (reviewed in ref. 3); moreover, retinoic acid–responsiveelements are locally activated in the regenerating adult rat nerve afterperipheral nerve injury4. These findings suggest the involvement ofretinoid signaling systems in embryonic and peripheral axonal regen-eration. However, the role of retinoids in adult CNS repair, known to beseverely restricted by inhibitory factors, has not been explored. Inembryonic DRG neurons, RARb2 mediates neurite outgrowth inducedby retinoic acid5; further, in adult spinal cord explants, where RARb2 isabsent, exogenous RARb2 stimulates neurite outgrowth6. We thereforeasked whether the delivery of RARb2 to adult neurons could primethem to switch to a regenerative mode in response to nerve injury andcould encourage axonal growth in the inhibitory CNS.
Peripheral sensory information is conveyed to the spinal cord byprimary sensory neurons whose cell bodies reside in the dorsal rootganglia (DRG). Injury to the dorsal roots of these DRG neurons, forexample in brachial plexus trauma during traffic accidents or childbirthinjury, can result in major sensory deficits7. The centrally projectingaxons of the injured sensory neurons regenerate in the peripheraldivision of the dorsal root but are arrested at the transition zonebetween the peripheral nervous system and the CNS (referred to as thedorsal root entry zone (DREZ)) and do not re-enter the spinal dorsalhorn8,9. This defined barrier to axonal ingrowth, together with thepossibility of genetic manipulation in DRG neurons, makes the DREZ agood model for exploring the regeneration of injured axons in anonpermissive environment.
Here we report that a lentiviral vector engineered to express RARb2promoted the regeneration of injured sensory afferents across the inhi-bitory DREZ and stimulated appropriate second-order targets in thespinal cord, thereby improving functional recovery in sensorimotor be-havioral tasks. Our results show that RARb2 triggers axonal growth pro-grams in injured neurons so as to overcome the DREZ barrier and thusdemonstrate a role for retinoid signaling in adult neural regeneration.
RESULTS
RARb2 enhances neurite outgrowth in DRG neurons in vitro
To transduce DRG neurons with RARb2 at high efficiency, we con-structed a minimal equine infectious anemia virus (EIAV)–basedlentiviral vector10,11 expressing RARb2 (EIAV-RARb2). As a control,we also constructed a vector expressing b-galactosidase (b-gal) (EIAV-LacZ). We first confirmed that 80.6 ± 2.8% (mean ± s.e.m. of theneurons were transduced at 3 d in vitro (DIV) following the addition ofthe vectors to the adult rat DRG neurons (data not shown). In control(that is, EIAV-LacZ–transduced) cultures, mature DRG neurons whengrown sparsely (500 neurons per well) on a reduced laminin substrate(0.1 mg ml–1) extended either very short neurites or no neurites at all(Fig. 1a). In contrast, neurons in EIAV-RARb2 cultures projectedseveral long neurites on an identical substrate (Fig. 1b). We quantifiedthese fiber outgrowths and, to ensure that only transduced neuronswere assessed, we measured neurites from only those neurons that wereeither b-gal– and bIII tubulin–positive or RARb– and bIII tubulin–positive. At 3 DIV, expression of RARb2 improved fiber growthninefold compared to that in EIAV-LacZ–transduced neurons(Fig. 1c; P o 0.001; throughout the text, except when indicated
Received 24 October; accepted 30 November; Published online 25 December 2005; corrected 13 January 2006 (details online); doi:10.1038/nn1622
1Oxford BioMedica (UK) Ltd., Medawar Centre, Robert Robinson Avenue, Oxford Science Park, Oxford OX4 4GA, UK. 2Neurorestoration Group, Wolfson Centre for Age-Related Diseases, Wolfson Wing, Hodgkin Building, King’s College London, Guy’s Campus, London SE1 1UL, UK. 3MRC Centre for Developmental Neurobiology, King’sCollege London, New Hunts House, Guy’s Campus, London SE1 1UL, UK. 4These authors contributed equally to this work. Correspondence should be addressed to L.-F.W.([email protected]) or N.D.M. ([email protected]).
NATURE NEUROSCIENCE VOLUME 9 [ NUMBER 2 [ FEBRUARY 2006 243
ART ICLES©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

differently, P values represent results of Student’s two-tailed unpairedt-test) and increased the percentage of process-bearing neurons to42.7 ± 6.4% from the 12.1 ± 5.8% observed in EIAV-LacZ cultures(Fig. 1d; P o 0.05). These results demonstrate that RARb2 promotedoutgrowth in adult DRG neurons by increasing neurite extension andby stimulating neuritogenesis in a larger proportion of neurons. Wenext asked whether the increase in neurite outgrowth in RARb2-transduced neurons was due to the activation of cyclic AMP (cAMP)signaling pathways. In the presence of an adenylate cyclase inhibitor,2¢,5¢-dideoxyadenosine (DDA), or a cell-permeant inhibitor of cAMP-dependent protein kinase A (PKA), Rp-cAMP, EIAV-RARb2 neuronsshowed decreased neurite lengths (Fig. 1e–g). This blocking of RARb2-enhanced regeneration was dose dependent, and at higher DDAconcentrations (50 and 100 mM) neurite outgrowth was completelyabrogated. Furthermore, intracellular cAMP concentrations in EIAV-RARb2 neurons were 38 times higher than those in EIAV-LacZ neurons(Fig. 1h; P o 0.01).
RARb2 induces outgrowth on the nonpermissive spinal cord
We have shown that the transduction of DRG neurons in vitropromoted neurite outgrowth on a minimal substrate. However, theCNS environment contains inhibitory factors, such as myelin and itsderivatives, that can restrict regeneration. To investigate the ability ofRARb2 to overcome this inhibition and promote outgrowth in anonpermissive environment, we studied neurite outgrowth in a cocul-ture system. After adult rat spinal cord and DRG neurons weretransduced with either EIAV-LacZ or EIAV-RARb2 in vivo, the trans-duced DRG neurons were grown on spinal cord sections that providedan inhibitory substrate for neurite outgrowth. EIAV-LacZ–transduced
DRG neurons grown either on naive spinal cord or on EIAV-RARb2–transduced spinal cord did not extend many neurites to the spinal cord(Fig. 2a). In the occasional neuron that settled on the peripheral sectionof the dorsal root, neurite outgrowth was observed, but these projec-tions extended up to the DREZ and did not extend into the spinal cord(Fig. 2b). By contrast, EIAV-RARb2 neurons grown both on naive andon EIAV-LacZ spinal cord often extended neurite projections (Fig. 2c)up to and beyond the DREZ into the spinal cord (Fig. 2d). In the samecultures, neurons that were not transduced with EIAV-RARb2 did notextend neurites beyond the DREZ or project fibers onto the spinal cord(Fig. 2d). Of the EIAV-RARb2 neurons, 26.3 ± 2.9% projected neuritesonto the spinal cord, compared to 3.0 ± 1.2% of the EIAV-LacZneurons (Fig. 2e; P o 0.01), and the average length of the longestneurite in EIAV-RARb2 neurons was also significantly higher than thatof EIAV-LacZ neurons (Fig. 2f; P o 0.01). These data suggested thatexpression of RARb2 in the DRG neurons was sufficient to overcomethe inhibitory environment of the adult spinal cord in order to promoteaxonal extension.
In vivo transduction of DRG neurons by EIAV-RARb2Although the above evidence indicates that RARb2, through theinduction of cAMP, stimulated neurite outgrowth of DRG neurons,it does not address regeneration in vivo. Therefore, we sought todetermine if RARb2 expression was sufficient to promote the regene-ration of injured sensory axons from the dorsal root into the spinalcord to establish functional connectivity. Adult DRG neurons expresslow levels of RARb2 mRNA and protein, and to deliver RARb2 to theDRG neurons, we used lentiviral vectors. We obtained high genetransfer efficiency to DRG neurons in vivo at 3 weeks after the unilateral
80 60
50
40
30
20
10
0
70
60
5040
30
20
10
0
6045403530
Fol
d in
duct
ion
In c
AM
Ple
vels 25
201510
50
EIAV-LacZEIAV-RARβ250
40
30
20
10
DDA (µM)
0** **
**
***
**
*
* **
– – – –20 50 100Rp-cAMP (µM) – – – – – 20 50
EIAV-LacZ EIAV-LacZ
Leng
th o
f lon
gest
neu
rite
(µm
)Le
ngth
of l
onge
st n
eurit
e(µ
m)
Per
cent
age
proc
ess-
bear
ing
neur
ons
EIAV-RARβ2 EIAV-RARβ2
EIAV-LacZ EIAV-RARβ2
EIAV-LacZ EIAV-RARβ2
EIAV-RARβ2+ Rp-cAMP
EIAV-RARβ2+ DDA
a b
e f g h
c d
Figure 1 RARb2 enhances neurite outgrowth from DRG neurons via the activation of cAMP signaling pathways. (a,b) DRG neurons were plated on reduced
laminin substrate (0.1 mg ml–1) and transduced with lentiviral vectors expressing (a) the control—that is, b-galactosidase (EIAV-LacZ) or (b) RARb2 (EIAV-
RARb2) at a multiplicity of infection of 10. Green, vector expression of b-gal or RARb2; red, neurite outgrowth detected by bIII-tubulin. (c,d) The average
length of the longest neurite (c) and the number of neurons possessing neurites (d) in transduced cultures. (e,f) RARb2 cultures were also incubated in an
adenylate cyclase inhibitor, DDA (100 mM; e), or a PKA inhibitor, Rp-cAMP (50 mM; f). (g) The average length of the longest neurite was assessed in EIAV-
LacZ– and EIAV-RARb2–transduced neurons in normal medium and in the presence of DDA or Rp-cAMP at various concentrations. (h) cAMP levels intransduced DRG cultures were quantified using a competitive immunoassay. Results are the mean ± s.e.m of at least eight experiments, each carried out in
triplicate. n ¼ 4 in each group unless otherwise stated. For measurement of neurite outgrowth, 150–220 neurons were assessed in each group. Black bar;
EIAV-LacZ–transduced DRGs; red bar, EIAV-RARb2–transduced DRGs. ***P o 0.001; **P o 0.01; *P o 0.05. In g, *P o 0.05 from EIAV-RARb2, untreated
group. Scale bar, 100 mm.
244 VOLUME 9 [ NUMBER 2 [ FEBRUARY 2006 NATURE NEUROSCIENCE
ART ICLES©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

injection of between 3 � 106 and 6 � 106 transducing units of rabies-Gpseudotyped EIAV-LacZ into the spinal cord. We observed genetransfer in the cuneate fasciculus and the dorsal horn of the spinalcord, which receive afferent projections from DRG neurons (Fig. 3a).The vector was retrogradely transported to ipsilateral DRG cell bodies,where we detected extensive b-gal expression in about 35% of neurons(Fig. 3b). RARb2 transcripts and protein were more abundant in EIAV-RARb2–transduced neurons than in the control, nontransduced DRGs(Fig. 3c–f). Moreover, the amount of protein expression was compar-able to that of embryonic day 14 (E14) DRGs (Fig. 3g). In the adult rat,vector transduction was achieved in the following three DRG sub-classes, as detected by colocalization with the respective markers(Fig. 3h): the large-diameter myelinated neurons, which expressheavy neurofilament protein NF200 and subserve mechanoreceptivefunctions, and the nociceptive small-diameter peptidergic and non-peptidergic neurons that are identified by their expression of theneuropeptide calcitonin gene–related peptide (CGRP) and by plantisolectin B4 (IB4), respectively12,13. The transduction efficiency of DRGneurons was as follows: 46 ± 2% coexpressed NF200, 33 ± 5%coexpressed CGRP and 22 ± 7% expressed IB4. These data indicated
that EIAV vectors pseudotyped with the rabies-G glycoprotein couldmediate efficient gene transfer to all classes of DRG neurons afterinjection into the spinal cord. Injection of the EIAV vector into thespinal cord did not induce a significant inflammatory response, as wasevident by normal Nissl staining and the lack of increased markers formacrophages (OX42), microglia (ED1) and T cells (CD8); furthermore,we observed no increased vascular density in the injected spinal cord(Supplementary Fig. 1 online).
30** **
****
25
20
15
10
LacZDRG :
Per
cent
age
proc
ess-
bear
ing
neur
ons
Cord : Naive Naive
RARβ2RARβ2LacZ
LacZ
RARβ2 LacZDRG :
Leng
th o
f lon
gest
neur
ite (
µm)
Cord : Naive Naive
RARβ2RARβ2LacZ
LacZ
RARβ2
5
0
30
25
20
15
10
5
0
β-gal/GAP43/GFAPGFAP
RARβ2/GAP43/GFAPGFAP RARβ2/GAP43/GFAPGFAP
β-gal/GAP43/GFAPGFAP
SC
SC
DR
DR
a
c
e
d
f
b Figure 2 DRG neurite extension in spinal cord coculture. DRGs transduced
with EIAV-LacZ or EIAV-RARb2 were cocultured with transduced or naive
spinal cord. (a) Neurite extension as detected by GAP43 staining (red) was
absent in EIAV-LacZ DRG neurons (green) that were cultured on EIAV-
RARb2–transduced spinal cord. (b) EIAV-LacZ neurons that were grown on
the dorsal root (DR) projected fibers up to the DREZ but did not enter the
spinal cord (SC) (white arrow). The central and peripheral regions of the
DREZ were delineated by GFAP staining in blue (white dotted line). (c) EIAV-RARb2 transduced DRG neurons (green) extended several neurites onto EIAV-
LacZ–transduced spinal cord. (d) Neurons that were present in the dorsal root
projected extensions up to and beyond the central portion of the DREZ
(white arrows). Neurons that were not transduced with EIAV-RARb2 did not
project neurites onto the spinal cord or beyond the DREZ (indicated by
white arrowheads). (e) The percentage of transduced neurons bearing neurites
on naive or transduced cord. (f) The length of the longest neurite on the
spinal cord from EIAV-LacZ– or EIAV-RARb2–transduced neurons. All
experiments were performed in triplicate with n ¼ 4 in each group. For
measurement of neurite outgrowth, 50–60 neurons were assessed in each
group; data is represented as mean ± s.e.m. **P o 0.01. Scale bars,
100 mm (a,c); 400 mm (b,d).
a
c
e
h
f g
b
d
NF200/β-gal CGRP/β-gal IB4/β-gal
Figure 3 EIAV-mediated gene transfer in the spinal cord and DRG.
(a) b-galactosidase expression was observed in the cuneate fasciculus
and the dorsal horn of the spinal cord 3 weeks after the infusion of the EIAV-
LacZ vector into the spinal cord. (b) Retrograde transport of the vector via the
dorsal root resulted in b-galactosidase expression in the ipsilateral DRG.
(c,d) With the EIAV-RARb2 vector, RARb2 mRNA was detected in the
ipsilateral DRG (c) and not in the contralateral uninjected side (d).
(e–g) RARb2 protein was observed in the ipsilateral DRG (e), and not in the
contralateral DRG (f) or in E14 rat DRG (g). (h) To assess vector expression in
the various subclasses of DRG neurons, b-galactosidase expression (red) in
the DRG was colocalized immunohistochemically with NF200, CGRP or IB4,
all indicated in green. Overlapping expression is illustrated in yellow. Scale
bars, 200 mm (a–d); 50 mm (e–h).
NATURE NEUROSCIENCE VOLUME 9 [ NUMBER 2 [ FEBRUARY 2006 245
ART ICLES©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e
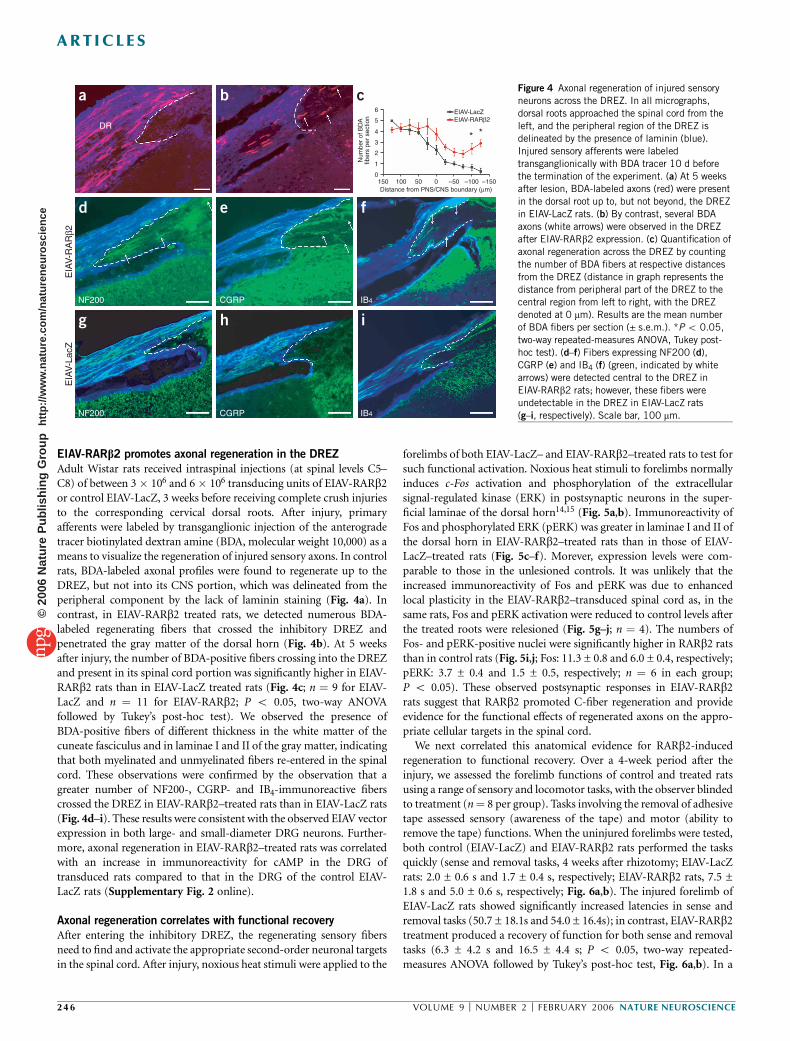
EIAV-RARb2 promotes axonal regeneration in the DREZ
Adult Wistar rats received intraspinal injections (at spinal levels C5–C8) of between 3 � 106 and 6 � 106 transducing units of EIAV-RARb2or control EIAV-LacZ, 3 weeks before receiving complete crush injuriesto the corresponding cervical dorsal roots. After injury, primaryafferents were labeled by transganglionic injection of the anterogradetracer biotinylated dextran amine (BDA, molecular weight 10,000) as ameans to visualize the regeneration of injured sensory axons. In controlrats, BDA-labeled axonal profiles were found to regenerate up to theDREZ, but not into its CNS portion, which was delineated from theperipheral component by the lack of laminin staining (Fig. 4a). Incontrast, in EIAV-RARb2 treated rats, we detected numerous BDA-labeled regenerating fibers that crossed the inhibitory DREZ andpenetrated the gray matter of the dorsal horn (Fig. 4b). At 5 weeksafter injury, the number of BDA-positive fibers crossing into the DREZand present in its spinal cord portion was significantly higher in EIAV-RARb2 rats than in EIAV-LacZ treated rats (Fig. 4c; n ¼ 9 for EIAV-LacZ and n ¼ 11 for EIAV-RARb2; P o 0.05, two-way ANOVAfollowed by Tukey’s post-hoc test). We observed the presence ofBDA-positive fibers of different thickness in the white matter of thecuneate fasciculus and in laminae I and II of the gray matter, indicatingthat both myelinated and unmyelinated fibers re-entered in the spinalcord. These observations were confirmed by the observation that agreater number of NF200-, CGRP- and IB4-immunoreactive fiberscrossed the DREZ in EIAV-RARb2–treated rats than in EIAV-LacZ rats(Fig. 4d–i). These results were consistent with the observed EIAV vectorexpression in both large- and small-diameter DRG neurons. Further-more, axonal regeneration in EIAV-RARb2–treated rats was correlatedwith an increase in immunoreactivity for cAMP in the DRG oftransduced rats compared to that in the DRG of the control EIAV-LacZ rats (Supplementary Fig. 2 online).
Axonal regeneration correlates with functional recovery
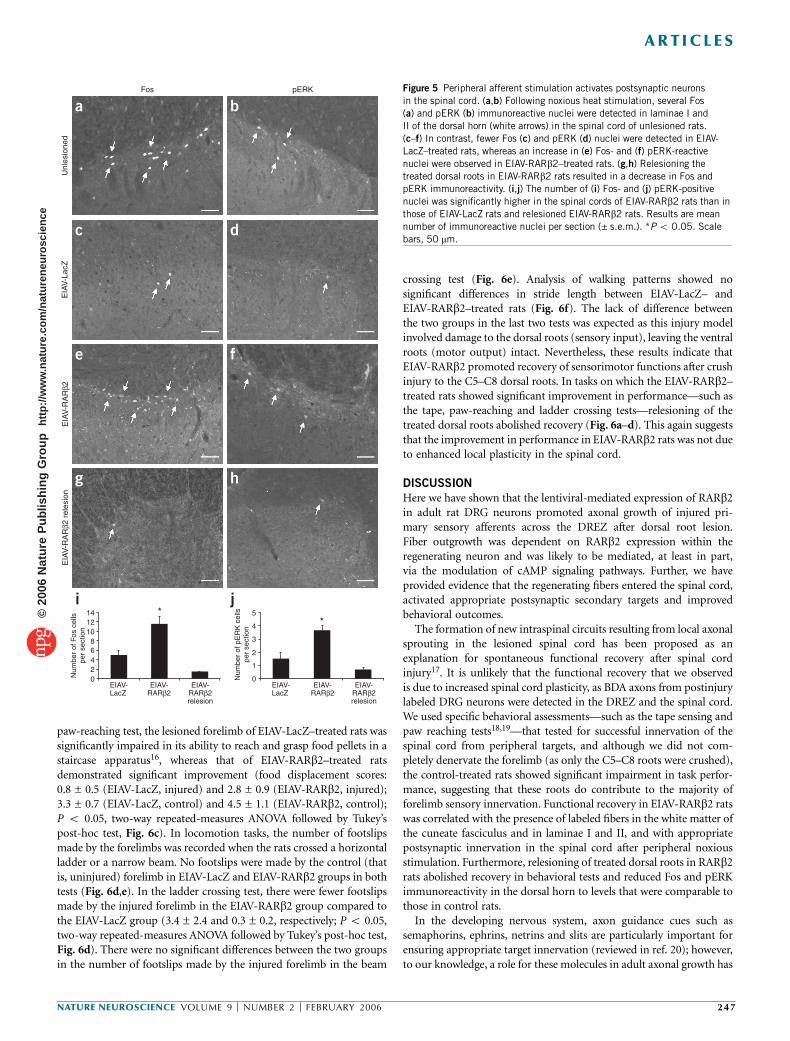
After entering the inhibitory DREZ, the regenerating sensory fibersneed to find and activate the appropriate second-order neuronal targetsin the spinal cord. After injury, noxious heat stimuli were applied to the
forelimbs of both EIAV-LacZ– and EIAV-RARb2–treated rats to test forsuch functional activation. Noxious heat stimuli to forelimbs normallyinduces c-Fos activation and phosphorylation of the extracellularsignal-regulated kinase (ERK) in postsynaptic neurons in the super-ficial laminae of the dorsal horn14,15 (Fig. 5a,b). Immunoreactivity ofFos and phosphorylated ERK (pERK) was greater in laminae I and II ofthe dorsal horn in EIAV-RARb2–treated rats than in those of EIAV-LacZ–treated rats (Fig. 5c–f). Morever, expression levels were com-parable to those in the unlesioned controls. It was unlikely that theincreased immunoreactivity of Fos and pERK was due to enhancedlocal plasticity in the EIAV-RARb2–transduced spinal cord as, in thesame rats, Fos and pERK activation were reduced to control levels afterthe treated roots were relesioned (Fig. 5g–j; n ¼ 4). The numbers ofFos- and pERK-positive nuclei were significantly higher in RARb2 ratsthan in control rats (Fig. 5i,j; Fos: 11.3 ± 0.8 and 6.0 ± 0.4, respectively;pERK: 3.7 ± 0.4 and 1.5 ± 0.5, respectively; n ¼ 6 in each group;P o 0.05). These observed postsynaptic responses in EIAV-RARb2rats suggest that RARb2 promoted C-fiber regeneration and provideevidence for the functional effects of regenerated axons on the appro-priate cellular targets in the spinal cord.
We next correlated this anatomical evidence for RARb2-inducedregeneration to functional recovery. Over a 4-week period after theinjury, we assessed the forelimb functions of control and treated ratsusing a range of sensory and locomotor tasks, with the observer blindedto treatment (n¼ 8 per group). Tasks involving the removal of adhesivetape assessed sensory (awareness of the tape) and motor (ability toremove the tape) functions. When the uninjured forelimbs were tested,both control (EIAV-LacZ) and EIAV-RARb2 rats performed the tasksquickly (sense and removal tasks, 4 weeks after rhizotomy; EIAV-LacZrats: 2.0 ± 0.6 s and 1.7 ± 0.4 s, respectively; EIAV-RARb2 rats, 7.5 ±1.8 s and 5.0 ± 0.6 s, respectively; Fig. 6a,b). The injured forelimb ofEIAV-LacZ rats showed significantly increased latencies in sense andremoval tasks (50.7 ± 18.1s and 54.0 ± 16.4s); in contrast, EIAV-RARb2treatment produced a recovery of function for both sense and removaltasks (6.3 ± 4.2 s and 16.5 ± 4.4 s; P o 0.05, two-way repeated-measures ANOVA followed by Tukey’s post-hoc test, Fig. 6a,b). In a
6
5
4 * *
Num
ber
of B
DA
fiber
s pe
r se
ctio
n
3
2
150 100Distance from PNS/CNS boundary (µm)
50 –50 –100
EIAV-LacZEIAV-RARβ2
–1500
1
0
DR
NF200
EIA
V-R
AR
β2E
IAV
-Lac
Z
NF200
CGRP
CGRP
IB4
IB4
a b
d e f
g h i
c Figure 4 Axonal regeneration of injured sensory
neurons across the DREZ. In all micrographs,
dorsal roots approached the spinal cord from the
left, and the peripheral region of the DREZ is
delineated by the presence of laminin (blue).
Injured sensory afferents were labeled
transganglionically with BDA tracer 10 d before
the termination of the experiment. (a) At 5 weeksafter lesion, BDA-labeled axons (red) were present
in the dorsal root up to, but not beyond, the DREZ
in EIAV-LacZ rats. (b) By contrast, several BDA
axons (white arrows) were observed in the DREZ
after EIAV-RARb2 expression. (c) Quantification of
axonal regeneration across the DREZ by counting
the number of BDA fibers at respective distances
from the DREZ (distance in graph represents the
distance from peripheral part of the DREZ to the
central region from left to right, with the DREZ
denoted at 0 mm). Results are the mean number
of BDA fibers per section (± s.e.m.). *P o 0.05,
two-way repeated-measures ANOVA, Tukey post-
hoc test). (d–f) Fibers expressing NF200 (d),
CGRP (e) and IB4 (f) (green, indicated by white
arrows) were detected central to the DREZ in
EIAV-RARb2 rats; however, these fibers were
undetectable in the DREZ in EIAV-LacZ rats
(g–i, respectively). Scale bar, 100 mm.
246 VOLUME 9 [ NUMBER 2 [ FEBRUARY 2006 NATURE NEUROSCIENCE
ART ICLES©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e
paw-reaching test, the lesioned forelimb of EIAV-LacZ–treated rats wassignificantly impaired in its ability to reach and grasp food pellets in astaircase apparatus16, whereas that of EIAV-RARb2–treated ratsdemonstrated significant improvement (food displacement scores:0.8 ± 0.5 (EIAV-LacZ, injured) and 2.8 ± 0.9 (EIAV-RARb2, injured);3.3 ± 0.7 (EIAV-LacZ, control) and 4.5 ± 1.1 (EIAV-RARb2, control);P o 0.05, two-way repeated-measures ANOVA followed by Tukey’spost-hoc test, Fig. 6c). In locomotion tasks, the number of footslipsmade by the forelimbs was recorded when the rats crossed a horizontalladder or a narrow beam. No footslips were made by the control (thatis, uninjured) forelimb in EIAV-LacZ and EIAV-RARb2 groups in bothtests (Fig. 6d,e). In the ladder crossing test, there were fewer footslipsmade by the injured forelimb in the EIAV-RARb2 group compared tothe EIAV-LacZ group (3.4 ± 2.4 and 0.3 ± 0.2, respectively; P o 0.05,two-way repeated-measures ANOVA followed by Tukey’s post-hoc test,Fig. 6d). There were no significant differences between the two groupsin the number of footslips made by the injured forelimb in the beam
crossing test (Fig. 6e). Analysis of walking patterns showed nosignificant differences in stride length between EIAV-LacZ– andEIAV-RARb2–treated rats (Fig. 6f). The lack of difference betweenthe two groups in the last two tests was expected as this injury modelinvolved damage to the dorsal roots (sensory input), leaving the ventralroots (motor output) intact. Nevertheless, these results indicate thatEIAV-RARb2 promoted recovery of sensorimotor functions after crushinjury to the C5–C8 dorsal roots. In tasks on which the EIAV-RARb2–treated rats showed significant improvement in performance—such asthe tape, paw-reaching and ladder crossing tests—relesioning of thetreated dorsal roots abolished recovery (Fig. 6a–d). This again suggeststhat the improvement in performance in EIAV-RARb2 rats was not dueto enhanced local plasticity in the spinal cord.
DISCUSSION
Here we have shown that the lentiviral-mediated expression of RARb2in adult rat DRG neurons promoted axonal growth of injured pri-mary sensory afferents across the DREZ after dorsal root lesion.Fiber outgrowth was dependent on RARb2 expression within theregenerating neuron and was likely to be mediated, at least in part,via the modulation of cAMP signaling pathways. Further, we haveprovided evidence that the regenerating fibers entered the spinal cord,activated appropriate postsynaptic secondary targets and improvedbehavioral outcomes.
The formation of new intraspinal circuits resulting from local axonalsprouting in the lesioned spinal cord has been proposed as anexplanation for spontaneous functional recovery after spinal cordinjury17. It is unlikely that the functional recovery that we observedis due to increased spinal cord plasticity, as BDA axons from postinjurylabeled DRG neurons were detected in the DREZ and the spinal cord.We used specific behavioral assessments—such as the tape sensing andpaw reaching tests18,19—that tested for successful innervation of thespinal cord from peripheral targets, and although we did not com-pletely denervate the forelimb (as only the C5–C8 roots were crushed),the control-treated rats showed significant impairment in task perfor-mance, suggesting that these roots do contribute to the majority offorelimb sensory innervation. Functional recovery in EIAV-RARb2 ratswas correlated with the presence of labeled fibers in the white matter ofthe cuneate fasciculus and in laminae I and II, and with appropriatepostsynaptic innervation in the spinal cord after peripheral noxiousstimulation. Furthermore, relesioning of treated dorsal roots in RARb2rats abolished recovery in behavioral tests and reduced Fos and pERKimmunoreactivity in the dorsal horn to levels that were comparable tothose in control rats.
In the developing nervous system, axon guidance cues such assemaphorins, ephrins, netrins and slits are particularly important forensuring appropriate target innervation (reviewed in ref. 20); however,to our knowledge, a role for these molecules in adult axonal growth has
14 5
4
3
2
1
0
**
Num
ber
of F
os c
ells
per
sect
ion
Num
ber
of p
ER
K c
ells
per
sect
ion
12
EIAV-LacZ
EIAV-RARβ2
EIAV-RARβ2relesion
EIAV-LacZ
EIAV-RARβ2
EIAV-RARβ2relesion
1086420
Fos
Unl
esio
ned
EIA
V-L
acZ
EIA
V-R
AR
β2E
IAV
-RA
Rβ2
rel
esio
n
pERK
a b
c d
e f
g h
i j
Figure 5 Peripheral afferent stimulation activates postsynaptic neurons
in the spinal cord. (a,b) Following noxious heat stimulation, several Fos
(a) and pERK (b) immunoreactive nuclei were detected in laminae I and
II of the dorsal horn (white arrows) in the spinal cord of unlesioned rats.
(c–f) In contrast, fewer Fos (c) and pERK (d) nuclei were detected in EIAV-
LacZ–treated rats, whereas an increase in (e) Fos- and (f) pERK-reactive
nuclei were observed in EIAV-RARb2–treated rats. (g,h) Relesioning the
treated dorsal roots in EIAV-RARb2 rats resulted in a decrease in Fos andpERK immunoreactivity. (i,j) The number of (i) Fos- and (j) pERK-positive
nuclei was significantly higher in the spinal cords of EIAV-RARb2 rats than in
those of EIAV-LacZ rats and relesioned EIAV-RARb2 rats. Results are mean
number of immunoreactive nuclei per section (± s.e.m.). *P o 0.05. Scale
bars, 50 mm.
NATURE NEUROSCIENCE VOLUME 9 [ NUMBER 2 [ FEBRUARY 2006 247
ART ICLES©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

not been demonstrated, suggesting that embryonic neural developmentmay be different from adult nerve regeneration. By contrast, we havedemonstrated that retinoid signaling, important in embryonic axongrowth, can be successfully manipulated to promote regeneration inthe adult CNS. Retinoic acid is important for the onset of neurogenesis,ventral spinal cord patterning and motor neuron development(reviewed in ref. 2 and references therein), and its receptor, RARb2,has a role in neuronal function. For example, rats deficient in RARbhave severe deficits in hippocampal long-term potentiation and long-term depression21. We have now further discovered a role for RARb2 instimulating fiber regeneration of injured neurons in an inhibitory CNSenvironment. We concluded that successful regeneration across theDREZ barrier was due to the expression of RARb2 in the DRG and nota consequence of the spinal cord being rendered more permissive byRARb2. We show that RARb levels are high in the DRG duringembryonic development, fall in the adult DRG and can be increasedto high levels again via lentiviral delivery. This exogenous RARb2expression may help prime embryonic-like regeneration programs topromote axonal extension. Indeed, embryonic DRG neurons, whenstimulated by retinoic acid to extend neurites, show a concordantincrease in RARb2 expression, presumably through the activation ofthe retinoic acid response element (RARE) present upstream of theRARb2 gene5. It is unlikely that the ligand for RARb2 is physiologicallylimiting, as retinoic acid is found throughout the adult rat brain(2.78 pmol per mg of tissue) and spinal cord (1.19 pmol per mg oftissue); these amounts comprise a proportionately higher level of theretinoid pool compared to those in other tissues22.
RARb2 expression in the adult DRG neurons was accompanied byan increase in intracellular cAMP, and inhibitors of adenylate cyclaseand PKA disrupted RARb2-induced neurite outgrowth, suggesting that
fiber outgrowth by RARb2 occurred, at least in part, via the modulationof cAMP-PKA pathways. Elevated cAMP concentrations23–25 and theactivation of cAMP response element binding protein (CREB) toovercome myelin inhibitors26 correlate with an enhanced regenerationcapacity in the adult CNS. Upon activation by its ligand, RARb2 acts asa transcription factor by binding as a heterodimer with retinoid Xreceptors (RXRs) to RAREs located in regulatory regions of targetgenes27. Retinoic acid increases mRNA levels of Gsa (the a-subunit ofthe stimulatory GTP-binding protein of adenylate cyclase) in mouseF9 teratocarcinoma cells28. Retinoic acid also induces calcium- andcalmodulin-stimulated adenylate cyclase activity, with an associatedspecific upregulation of adenylate cyclase isoform mRNAs, in P19cells29. It is likely that these increased transcriptional activities, leadingto increased cAMP levels, are mediated by ligand-activated RARspresent in these cell lines30. In an additional experiment, we foundthat the injection of dibutyryl cAMP (db-cAMP) into adult DRGs inthis DREZ model promoted functional recovery of injured sensoryafferents; however, recovery was not as robust as that of the afferents inEIAV-RARb2 rats (Supplementary Fig. 3 online). Enhanced RARb2-mediated regeneration may be due to differences in sustained cAMPlevels or peak physiological concentrations; alternatively, it is possiblethat other signaling pathways apart from cAMP contribute to RARb2-mediated regeneration.
Studies on dorsal root regeneration in the spinal cord have primarilyfocused on providing trophic support31–33 or increasing intracellularcAMP concentrations24,25. Here we provide evidence, using anatomicaland behavioral outcomes, that RARb2 acts as a molecular switchto stimulate the regeneration of injured sensory fibers across theinhibitory DREZ in the adult CNS. The use of EIAV vectors to expressa growth-promoting molecule such as RARb2 in situ may offer certain
80*
* *
**
* * * *
* * * * * * *80
60
40
20
0
Tape removal (sense) Tape removal (remove)
Tim
e (s
)F
ood
disp
lace
men
t sco
reN
umbe
r of
foot
slip
s
Tim
e (s
)
60
40
20
–1 0 1 2 3 4 RelesionWeeks post-injury
Paw reaching
Beam crossing Footprint analysis
Ladder crossing
Num
ber
of fo
otsl
ips
Str
ide
leng
th (
mm
)
–1 0 1 2 3 4 RelesionWeeks post-injury
–1 0 1 2 3 4Weeks post-injury
EIAV-LacZ injured forelimb EIAV-RARβ2 injured forelimb
EIAV-RARβ2 control forelimbEIAV-LacZ control forelimb
–1 0 1 2 3 4Weeks post-injury
–1 0 1 2 3 4 RelesionWeeks post-injury
–1 0 1 2 3 4 RelesionWeeks post-injury
0
6
9 200
150
100
50
0
7
5
3
1
15131197531
5
4
3
2
1
0
a
c d
e f
bFigure 6 RARb2 promoted functional recovery
after axonal regeneration. In all graphs, black
closed and open squares represent injured
and uninjured forelimbs of EIAV-LacZ rats,
respectively, whereas red closed and open
diamonds represent injured and uninjured
forelimbs of EIAV-RARb2 rats, respectively. Rats
were assessed in behavioral tasks before andfor 4 weeks after rhizotomy. (a,b) In a tape
removal task, the time taken to sense and
remove the tape by the lesioned and unlesioned
forelimbs was measured separately, and lower
latencies were observed with the injured
forelimbs of EIAV-RARb2 than those of
EIAV-LacZ rats. (c) Unilateral assessment
of food displacement in a staircase apparatus
showed improved scores with the injured forelimb
of EIAV-RARb2 rats compared to that of EIAV-
LacZ rats. (d–f) In locomotor tasks, the number
of footslips made by the injured forelimb of EIAV-
RARb2 rats was lower than that of EIAV-LacZ rats
(d), whereas no significant differences were
observed between the two groups during the beam
crossing test (e) and footprint analyses (f). After
the observed recovery in EIAV-RARb2 rats, treated
dorsal roots were relesioned; this resulted in task
performances that were comparable to those ofEIAV-LacZ rats in the tape, paw reaching and
ladder crossing tests (a–d). Data represent mean ±
s.e.m. *P o 0.05, two-way repeated-measures
ANOVA, Tukey’s post-hoc test).
248 VOLUME 9 [ NUMBER 2 [ FEBRUARY 2006 NATURE NEUROSCIENCE
ART ICLES©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

advantages over pharmacological approaches as it obviates the need forcontinuous delivery via intrathecal pumps to obtain long-term andefficacious therapy34,35, has target cell specificity and avoids undesirableside effects resulting from systemic manipulation of common intracel-lular signaling pathways. Uncovering other mechanisms of RARb2-induced neurite regeneration may also reveal new targets for treatingspinal cord injury, thereby offering new therapeutic opportunities innerve regeneration.
METHODSViral vector production. EIAV vector genomes were constructed from
pSMART2 lentiviral vectors as previously described35. The Rarb2 or LacZ gene
was inserted under the control of a minimal human cytomegalovirus (hCMV)
promoter in an EIAV transfer vector containing a 5¢ central polypurine tract
(cPPT) element and a 3¢ woodchuck post-transcriptional regulatory element
(WPRE) enhancer (vector map illustrated in Supplementary Fig. 1).
Viral vector stocks pseudotyped with the rabies-G envelope glycoprotein
were prepared by triple plasmid transient transfection of HEK293T cells as
previously described10,34–36. The titer of EIAV-LacZ was determined by
transduction of dog osteosarcoma D17 cells (4 � 108 TU ml–1), and the titer
of EIAV-RARb2 (8 � 108 TU ml–1) was calculated by determining the
normalized viral RNA genome copy number using quantitative reverse tran-
scriptase polymerase chain reaction (RT-PCR) analysis and comparing it to
EIAV-LacZ as previously described37.
DRG neuron culture. Neuronal dissociated DRG cultures were prepared as
previously described38. Adult male Wistar rats (200–250 g) were killed accord-
ing to institutional (King’s College, London) and UK Home Office regulations.
The DRGs were dissected and transferred to Ham’s F12 medium (Gibco).
DRGs were desheathed and trimmed, digested in 0.125% collagenase (Sigma)
at 37 1C for 2 h, and mechanically dissected by trituration with a P1000 Gilson
pipette in 1 ml of modified Bottenstein and Sato’s culture medium (BS) in
Ham’s F12. The cell suspension was centrifuged at 600g for 8 min through a
cushion of 15% bovine serum albumin (BSA, Sigma). The dissociated neurons
were resuspended in 100 ml of calcium- and magnesium-free Hank’s balanced
buffer solution (HBSS, Gibco) containing 50 mg ml–1 DNase (Type I, Sigma)
and 250 mg ml–1 soybean trypsin inhibitor (Type II, Sigma) and diluted in
modified BS culture medium to approximately 1,600 cells per ml. Cells (500 per
well) were plated in eight-well plates (Labtek) that were precoated with poly-
L-lysine (2 mg ml–1; Sigma), Engelbreth-Holm-Swarm (EHS) laminin (0.1 mg
ml–1 Sigma). Cells were incubated at 37 1C in a humidified atmosphere
containing 5% CO2. Neurite outgrowth from dissociated DRG neurons was
affected by low cell density and low laminin concentrations (data not shown).
The appropriate viral vectors were added to the cultures at a multiplicity of
infection (MOI) of 10 at DIV0. For cAMP inhibition experiments, cultures
were incubated from DIV1 in DDA (Sigma) or Rp-adenosine-3¢,5¢-cyclic
monophosphorothioate (sodium salt, Rp-cAMP, Sigma) at the concentrations
indicated. At DIV3, neurons were either harvested for measurement of cAMP
using a competitive immunoassay, according to the manufacturer’s instructions
(Amersham), or fixed for 30 min in 4% paraformaldehyde for assessment of
neurite outgrowth. Cells were permeabilized with methanol at –20 1C for
3 min, washed with phosphate-buffered saline (PBS) and incubated at room
temperature for 2 h with a combination of mouse b-III tubulin (1:1,000) with
either rabbit b-galactosidase (1:300, Europa Biolabs) or rabbit RARb (1:50,
Santa Cruz). Secondary antibody staining was performed with a mixture of
Alexa Fluor 488 (1:1,000, Molecular Probes) and Alexa Fluor 546 (1:1,000,
Molecular Probes) at room temperature for 1 h. After PBS washes, cells were
mounted with FluorSave reagent (Calbiochem) and observed under a Zeiss
microscope. The length of the longest neurite for the first 150–220 neurons
encountered when scanning the slide in a systematic manner was determined
using an image analysis program (SigmaScan Pro 4.01) and was expressed as
mean length ± s.e.m.38.
Preparation of coculture. Rats were administered either the EIAV-RARb2 or
the EIAV-LacZ viral vector as described below (n ¼ 3 in each group).
Transduced DRG neurons were prepared for culture 3 weeks after surgery, as
described above. Cryosections of the spinal cord were prepared according to a
previous study39. Longitudinal sections (10 mm) of the frozen cord were thaw-
mounted onto sterile 13-mm-diameter poly-L-lysine–coated glass coverslips,
and dissociated DRG neurons in modified BS culture medium were placed at
150 cells per well onto the cryosections. After 3–4 d of incubation, cocultures
were fixed and stained and neurite outgrowth was quantified as described
above. At least 50–60 transduced neurons in each experiment were analyzed.
Primary antibodies used were to mouse b-galactosidase (1:600, Promega),
mouse RARb (1:300, Chemicon), rabbit GAP43 (1:1,500, Chemicon) and
goat GFAP (1:300, Santa Cruz), and secondary antibodies used were to
Cy3 (1:600), 7-amino-4-methylcoumarin-3-acetic acid (AMCA, 1: 300) and
fluorescein-5-isothiocyanate (FITC, 1:300).
Viral vector delivery and nerve labeling. All rat experiments were approved by
the local veterinarian and ethical committees and carried out according to UK
Home Office regulations. In anesthetized adult Wistar rats (given ketamine and
medetomidine 60 mg kg–1 and 0.25 mg kg–1, respectively, intraperitoneally),
cervical spinal cords were exposed by laminectomy and EIAV vectors were
infused unilaterally into four sites in the spinal cord (C5–C8 spinal level) at a
rate of 0.2 ml min–1 via a finely drawn glass micropipette attached to an
infusion pump. We injected 3 � 106–6 � 106 transducing units of each vector.
The C5–C8 dorsal roots were crushed unilaterally (two times each, 10 s per
crush) midway between the DRG and DREZ by an operator blinded to the
treatment groups, 3 weeks after vector delivery; then the rats were tested
behaviorally for 4 weeks (n ¼ 8 per group). In the relesion experiments, the
C6–C8 dorsal roots of EIAV-RARb2–treated rats were transected at 4 weeks
after crushing and then tested again at 5 d after transection (n ¼ 4). For
transganglionic tracing, 1 ml of 10% BDA (molecular weight 10,000; Molecular
Probes) was injected directly into the DRG. Rats were killed at 10 d after
BDA labeling to assess axonal regeneration (n ¼ 9 EIAV-LacZ and n ¼ 11
EIAV-RARb2). For noxious stimulation experiments, rats were anesthetized
with urethane 2 h before the termination of the experiment, and the injured
forelimbs were dipped into a 52 1C water bath (3 � 20 s at 90-s intervals
and repeated at 2 h from the initial dip; n ¼ 6 each for EIAV-LacZ and
EIAV-RARb2).
In situ hybridization. Rats were killed and perfused transcardially with
heparinized 0.9% NaCl solution and 4% paraformaldehyde in 0.1 M phosphate
buffer. The cervical cords with attached DRGs were dissected, frozen and
sectioned (20 mm) on a cryostat. For detection of EIAV-RARb2 transcripts,
we carried out in situ hybridization using nonradioactive riboprobes specific
to RARb2 as previously described40,41; the development was performed after
2 h hybridization.
Immunohistochemistry. Sections were analyzed by 5-bromo-4-chloro-3-
indolyl-b-D-galactoside (X-gal) staining and immunohistochemistry. RARbwas also detected in 20-mm sections of E14 DRGs. Primary antibodies were
used as follows: mouse NF200 (1: 400, N52, Sigma), rabbit CGRP (1:8,000,
Sigma), Griffonia simplicifolia isolectin B4 (10 mg ml–1, Sigma), rabbit
b-galactosidase (1:300, Europa Labs), rabbit RARb (1:50, Santa Cruz), rabbit
laminin (1:200, Sigma), rabbit Fos (1:10,000, Oncogene Sciences) and rabbit
pERK (1:200, Sigma). For inflammatory response markers (Supplementary
Fig. 1), antibodies used were NeuroTrace fluorescent Nissl (1:100, Molecular
Probes), OX42 (1:100, Chemicon), ED1 (1:1,000, Chemicon), CD8 (1:100,
Serotec) and Glut1 (1:1,000, Serotec). cAMP staining (Supplementary Fig. 2)
was performed with rabbit antibody to cAMP (1:1,000, Chemicon). Extra-
avidin conjugated to FITC (1:200, Sigma) was used to detect BDA-labeled
axonal tracts. Secondary antibodies used were conjugated to one of the
following: fluorescein isothiocyanate, Texas Red (Jackson Labs), Alexa Fluor
488 and 546 (Molecular Probes), or AMCA (Jackson Labs). Quantitative
analysis was carried out by counting BDA-labeled fibers at measured intervals
within a 1-mm square grid graticule in the peripheral and central regions of
DREZ (delineated by laminin staining) by a blinded experimenter. At least 36
random sections were counted for each rat. For transduction efficiency,
immunopositive cells were counted in six sections per rat and expressed as a
percentage of b-galactosidase positive cells. For Fos and pERK quantification,
immunopositive cells were counted in 36 random sections per rat.
NATURE NEUROSCIENCE VOLUME 9 [ NUMBER 2 [ FEBRUARY 2006 249
ART ICLES©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

Behavioral analysis. Testing was carried out by experimenters blinded to the
treatment for 1 week before and for 4 weeks after crush injury (n ¼ 8 each in
EIAV-LacZ and EIAV-RARb2). In the tape removal test18, adhesive tape (1.5 cm
� 1 cm) was placed on each forepaw separately, and we scored the times taken
to sense the tape (indicated by paw shake) and remove the tape (cutoff
timepoint, 2 min). The paw reaching test assesses the rats’ ability to reach
and grasp food pellets in a staircase apparatus and measures side bias,
maximum forelimb extension and grasping skill16. Food displacement was
scored after 10 min in the staircase apparatus. For locomotor tasks (adapted
from ref. 19), rats were trained to cross a horizontal beam (2.5 cm � 100 cm)
or ladder (18 cm � 100 cm with each rung 5 cm apart) and the number of
forelimb footslips (off the beam or below the plane of the ladder) was recorded.
In footprint analysis (adapted from ref. 19), the stride lengths and widths were
calculated during continuous locomotion across a wooden runway. Behavioral
responses of treated and control groups were compared using two-way
repeated-measures ANOVA followed by Tukey post-hoc test.
Note: Supplementary information is available on the Nature Neuroscience website.
ACKNOWLEDGMENTSWe thank M. Agudo for help with the synthesis of the RARb2 riboprobe,E. Foster for help with DRG cultures and L. Walmsley for technical help. Thiswork was supported by Oxford BioMedica and the Medical Research Council.
COMPETING INTERESTS STATEMENTThe authors declare competing financial interests (see the Nature Neurosciencewebsite for details).
Published online at http://www.nature.com/natureneuroscience/
Reprints and permissions information is available online at http://npg.nature.com/
reprintsandpermissions/
1. Maden, M. Retinoid signalling in the development of the central nervous system. Nat.Rev. Neurosci. 3, 843–853 (2002).
2. Appel, B. & Eisen, J.S. Retinoids run rampant: multiple roles during spinal cord andmotor neuron development. Neuron 40, 461–464 (2003).
3. Maden, M. Role and distribution of retinoic acid during CNS development. Int. Rev.Cytol. 209, 1–77 (2001).
4. Zhelyaznik, N., Schrage, K., McCaffery, P. & Mey, J. Activation of retinoic acid signallingafter sciatic nerve injury: up-regulation of cellular retinoid binding proteins. Eur.J. Neurosci. 18, 1033–1040 (2003).
5. Corcoran, J., Shroot, B., Pizzey, J. & Maden, M. The role of retinoic acid receptors inneurite outgrowth from different populations of embryonic mouse dorsal root ganglia.J. Cell Sci. 113, 2567–2574 (2000).
6. Corcoran, J. et al. Retinoic acid receptor b2 and neurite outgrowth in the adult mousespinal cord in vitro. J. Cell Sci. 115, 3779–3786 (2002).
7. Dubuisson, D. Nerve root damage and arachnoiditis. in Textbook of Pain (eds. Wall, P.D.& Melzack, R.) Ch. 38, 544–565 (Churchill Livingstone, Edinburgh, 1989).
8. Ramon y Cajal, S. Degeneration and Regeneration of the Nervous System (Hafner,New York, 1928).
9. Carlstedt, T. Regenerating axons form nerve terminals at astrocytes. Brain Res. 347,188–191 (1985).
10. Mazarakis, N.D. et al. Rabies virus glycoprotein pseudotyping of lentiviral vectorsenables retrograde axonal transport and access to the nervous system after peripheraldelivery. Hum. Mol. Genet. 10, 2109–2121 (2001).
11. Wong, L.F. et al. Transduction patterns of pseudotyped lentiviral vectors in the nervoussystem. Mol. Ther. 9, 101–111 (2004).
12. Averill, S., McMahon, S.B., Clary, D.O., Reichardt, L.F. & Priestley, J.V. Immunocyto-chemical localization of trkA receptors in chemically identified subgroups of adult ratsensory neurons. Eur. J. Neurosci. 7, 1484–1494 (1995).
13. Bennett, D.L. et al. A distinct subgroup of small DRG cells express GDNF receptorcomponents and GDNF is protective for these neurons after nerve injury. J. Neurosci. 18,3059–3072 (1998).
14. Hunt, S.P., Pini, A. & Evan, G. Induction of c-fos-like protein in spinal cord neuronsfollowing sensory stimulation. Nature 328, 632–634 (1987).
15. Ji, R.R., Baba, H., Brenner, G.J. & Woolf, C.J. Nociceptive-specific activation of ERK inspinal neurons contributes to pain hypersensitivity. Nat. Neurosci. 2, 1114–1119(1999).
16. Montoya, C.P., Campbell-Hope, L.J., Pemberton, K.D. & Dunnett, S.B. The ‘‘staircasetest’’: a measure of independent forelimb reaching and grasping abilities in rats.J. Neurosci. Methods 36, 219–228 (1991).
17. Bareyre, F.M. et al. The injured spinal cord spontaneously forms a new intraspinal circuitin adult rats. Nat. Neurosci. 7, 269–277 (2004).
18. Thallmair, M. et al. Neurite growth inhibitors restrict plasticity and functional recoveryfollowing corticospinal tract lesions. Nat. Neurosci. 1, 124–131 (1998).
19. Kunkel-Bagden, E., Dai, H.N. & Bregman, B.S. Methods to assess the developmentand recovery of locomotor function after spinal cord injury in rats. Exp. Neurol. 119,153–164 (1993).
20. Fournier, A.E. & Strittmatter, S.M. Repulsive factors and axon regeneration in the CNS.Curr. Opin. Neurobiol. 11, 89–94 (2001).
21. Chiang, M.Y. et al. An essential role for retinoid receptors RARbeta and RXRgamma inlong-term potentiation and depression. Neuron 21, 1353–1361 (1998).
22. Werner, E.A. & Deluca, H.F. Retinoic acid is detected at relatively high levels in the CNSof adult rats. Am. J. Physiol. Endocrinol. Metab. 282, E672–E678 (2002).
23. Cai, D. et al. Neuronal cyclic AMP controls the developmental loss in ability of axons toregenerate. J. Neurosci. 21, 4731–4739 (2001).
24. Neumann, S., Bradke, F., Tessier-Lavigne, M. & Basbaum, A.I. Regeneration of sensoryaxons within the injured spinal cord induced by intraganglionic cAMP elevation. Neuron34, 885–893 (2002).
25. Qiu, J. et al. Spinal axon regeneration induced by elevation of cyclic AMP. Neuron 34,895–903 (2002).
26. Gao, Y. et al. Activated CREB is sufficient to overcome inhibitors in myelin and promotespinal axon regeneration in vivo. Neuron 44, 609–621 (2004).
27. Chambon, P. A decade of molecular biology of retinoic acid receptors. FASEB J. 10,940–954 (1996).
28. Chan, S.D., Strewler, G.J. & Nissenson, R.A. Transcriptional activation of Gs alphaexpression by retinoic acid and parathyroid hormone-related protein in F9 teratocarci-noma cells. J. Biol. Chem. 265, 20081–20084 (1990).
29. Lipskaia, L., Djiane, A., Defer, N. & Hanoune, J. Different expression of adenylyl cyclaseisoforms after retinoic acid induction of P19 teratocarcinoma cells. FEBS Lett. 415,275–280 (1997).
30. Hu, L. & Gudas, L.J. Cyclic AMP analogs and retinoic acid influence the expression ofretinoic acid receptor alpha, beta, and gamma mRNAs in F9 teratocarcinoma cells.Mol.Cell. Biol. 10, 391–396 (1990).
31. Romero, M.I., Rangappa, N., Garry, M.G. & Smith, G.M. Functional regeneration ofchronically injured sensory afferents into adult spinal cord after neurotrophin genetherapy. J. Neurosci. 21, 8408–8416 (2001).
32. Ramer, M.S., Priestley, J.V. & McMahon, S.B. Functional regeneration of sensory axonsinto the adult spinal cord. Nature 403, 312–316 (2000).
33. Zhang, Y., Dijkhuizen, P.A., Anderson, P.N., Lieberman, A.R. & Verhaagen, J. NT-3delivered by an adenoviral vector induces injured dorsal root axons to regenerate into thespinal cord of adult rats. J. Neurosci. Res. 54, 554–562 (1998).
34. Azzouz, M. et al. Multicistronic lentiviral vector-mediated striatal gene transfer ofaromatic L-amino acid decarboxylase, tyrosine hydroxylase, and GTP cyclohydrolase Iinduces sustained transgene expression, dopamine production, and functional improve-ment in a rat model of Parkinson’s disease. J. Neurosci. 22, 10302–10312 (2002).
35. Bienemann, A.S. et al. Long-term replacement of a mutated nonfunctional CNS gene:reversal of hypothalamic diabetes insipidus using an EIAV-based lentiviral vectorexpressing arginine vasopressin. Mol. Ther. 7, 588–596 (2003).
36. Mitrophanous, K. et al. Stable gene transfer to the nervous system using a non-primatelentiviral vector. Gene Ther. 6, 1808–1818 (1999).
37. Martin-Rendon, E., White, L.J., Olsen, A., Mitrophanous, K.A. & Mazarakis, N.D. Newmethods to titrate EIAV-based lentiviral vectors. Mol. Ther. 5, 566–570 (2002).
38. Gavazzi, I., Kumar, R.D., McMahon, S.B. & Cohen, J. Growth responses of differentsubpopulations of adult sensory neurons to neurotrophic factors in vitro. Eur.J. Neurosci. 11, 3405–3414 (1999).
39. Golding, J.P., Bird, C., McMahon, S. & Cohen, J. Behaviour of DRG sensory neuritesat the intact and injured adult rat dorsal root entry zone: postnatal neurites becomeparalysed, whilst injury improves the growth of embryonic neurites. Glia 26, 309–323(1999).
40. Zelent, A., Krust, A., Petkovich, M., Kastner, P. & Chambon, P. Cloning of murine alphaand beta retinoic acid receptors and a novel receptor gamma predominantly expressed inskin. Nature 339, 714–717 (1989).
41. Rattray, M. & Michael, G.J. Oligonucleotide probes for in situ hybridization. in In SituHybridization-A Practical Approach (ed. Wilkinson, D.G.) Ch. 2, 23–67 (Oxford Univ.Press, Oxford, 1998).
250 VOLUME 9 [ NUMBER 2 [ FEBRUARY 2006 NATURE NEUROSCIENCE
ART ICLES©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e
Copyright © 2022 FDOKUMEN




















