
Calcium-activated proteolysis of neurofilament proteins in goldfish Mauthner axons
9
Joririitrl oJ' Neiwoclieinistiy Raven Press, New York 'C 1986 International Society for Neurochemistry Calcium-Activated Proteolysis of Neurofilament Proteins in the Squid Giant Neuron *?Paul E. Gallant, *tHarish C. Pant, $Rebecca M. Pruss, and tPHarold Gainer *Laboratory of Preclinicul Studies, Nutionul Institirte on Alcohol Abirse and Alcoholism, Rockville, §Laboratory of Neurochemistry nnd Neur-oimrnirnology, Nationnl Institute oj Child Health and Human Development, Bethesdu, $Laboratory of Cell Biology, Nutionul Institutr of Mental Health, Bethesda, Maryland; und fMarine Biological Laboratory, Woods Hole, Massachim?tts, U.S.A. Abstract: The phosphorylation and proteolysis of squid neurofilament proteins by endogenous kinase and cal- cium-activated protease activities, respectively, were studied. When axoplasm was incubated in the presence of [y-'2P]ATP, most of the phosphate was incorporated into two neurofilament proteins: a 220-kilodalton (NF-220) and a high-molecular-weight (HMW) protein. When this phosphorylated axoplasm was subjected to en- dogenous calcium-activated proteolysis, two significant phosphorylated fragments were generated, ix., a soluble 110K fragment and a pelletable IOOK fragment. Irnrnuno- chemical and other analyses suggest that the pelletable IOOK fragment contains the common helical neurofila- ment rod region and that the soluble 110K protein is the putative side arm of the NF-220. In contrast, neither the HMW or the NF-220 was detected in the region of the stellate ganglion which contains the cell bodies of the giant axon. However, this region did contain a number of Neurofilaments along with microtubules and mi- crofilaments are the major components of the cyto- plasmic (or neuroplasmic) network in the squid giant axon (Hodge and Adelman, 1979). The neuro- filaments appear to act as a three-dimensional structural lattice that provides tensile strength to the axon (Rubinson and Baker, 1979; Lazarides, 1980) and also may be determinants of axonal size (Hoffman et al., 1984). After being synthesized in the cell body, neurofilaments are subject to post- translational modifications by protein kinase (Pant et al., 1978; Eagles et al., 1981; Julien and Mu- shynski, 1982) and by calcium-activated protease (CAP) (Gilbert et al., 1975; Schlaepfer and Micko, Received May 21, 1985; accepted December 3, 1985. Address correspondence and reprint requests to Dr. P. E. Gallant at Laboratory of Preclinical Studies, National Institute on Alcohol Abuse and Alcoholism, 12501 Washington Avenue, Rockville, MD 20852, U.S.A. Ahhreviatiom used: BME, p-mercaptoethanol; CAP, calcium- proteins that were sensitive to calcium-activated proteol- ysis and reacted with a monoclonal intermediate filament antibody. This intermediate filament antibody reacts with most of the axoplssmic proteins that copurify with neuro- filaments, i.e., in the order of their intermediate filament antibody staining intensity, a 60K, 65K, 220K, and 74K protein. In the cell body preparation, the intermediate fil- ament antibody labeled, in order of their staining inten- sity, a 65K, 60K, 74K, and 180K protein. In both the axo- plasmic and cell body preparations, endogenous calcium- activated proteolysis generated characteristic fragments that could be labeled with the anti-intermediate filament antibody. Key Words: Neurofilament -Phosphorylation - Calcium-activated proteol y si s - Squid - Giant axon - Intermediate filament. Gallant P. E. et al. Calcium-acti- vated proteolysis of neurofilament proteins in the squid giant neuron. J. Neurochem. 46, 1573- 1581 (1986). 1978; Pant et al., 19790; Pant and Gainer, 1980) ac- tivities. Phosphorylated neurofilament proteins are present in mammalian brain (Julien and Mushynski, 1981) and squid (Pant et al., 1979b,c) axons in vivo, but are not found in a phosphorylated form in many neuronal perikarya (Goldstein et al., 1983; Stern- berger and Sternberger, 1983). However, it is not clear what function neurofilament phosphorylation serves. It has been suggested that phosphorylation may control the assembly of neurofilament proteins into formed neurofilaments (Wong et al., 1984), or that it might stabilize the neurofilaments into com- pact structures in the axon (Sternberger and Stern- berger, 1983). activated protease; HEPES, N-2-hydroxyethylpiperazine-N'-2- ethanesulfonic acid; HMW, high molecular weight; IFA, inter- mediate filament antibody; NF-60 and NF-220, neurofilament polypeptides with molecular sizes of 60 and 220 kilodaltons, re- spectively; PAGE, polyacrylamide gel electrophoresis; SDS, so- dium dodecyl sulfate. 1573
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Calcium-activated proteolysis of neurofilament proteins in goldfish Mauthner axons

Joririitrl oJ' Neiwoclieinistiy Raven Press, New York 'C 1986 International Society for Neurochemistry
Calcium- Activated Proteolysis of Neurofilament Proteins in the Squid Giant Neuron
*?Paul E. Gallant, *tHarish C. Pant, $Rebecca M. Pruss, and tPHarold Gainer
*Laboratory of Preclinicul Studies, Nutionul Institirte on Alcohol Abirse and Alcoholism, Rockville, §Laboratory of Neurochemistry nnd Neur-oimrnirnology, Nationnl Institute o j Child Health and Human Development,
Bethesdu, $Laboratory of Cell Biology, Nutionul Institutr of Mental Health, Bethesda, Maryland; und fMarine Biological Laboratory, Woods Hole, Massachim?tts, U.S.A.
Abstract: The phosphorylation and proteolysis of squid neurofilament proteins by endogenous kinase and cal- cium-activated protease activities, respectively, were studied. When axoplasm was incubated in the presence of [y-'2P]ATP, most of the phosphate was incorporated into t w o neurofilament proteins: a 220-kilodalton (NF-220) and a high-molecular-weight (HMW) protein. When this phosphorylated axoplasm was subjected to en- dogenous calcium-activated proteolysis, two significant phosphorylated fragments were generated, ix., a soluble 110K fragment and a pelletable IOOK fragment. Irnrnuno- chemical and other analyses suggest that the pelletable IOOK fragment contains the common helical neurofila- ment rod region and that the soluble 110K protein is the putative side arm of the NF-220. In contrast, neither the HMW or the NF-220 was detected in the region of the stellate ganglion which contains the cell bodies of the giant axon. However, this region did contain a number of
Neurofilaments along with microtubules and mi- crofilaments are the major components of the cyto- plasmic (or neuroplasmic) network in the squid giant axon (Hodge and Adelman, 1979). The neuro- filaments appear to act as a three-dimensional structural lattice that provides tensile strength to the axon (Rubinson and Baker, 1979; Lazarides, 1980) and also may be determinants of axonal size (Hoffman et al., 1984). After being synthesized in the cell body, neurofilaments are subject to post- translational modifications by protein kinase (Pant et al., 1978; Eagles et al., 1981; Julien and Mu- shynski, 1982) and by calcium-activated protease (CAP) (Gilbert et al., 1975; Schlaepfer and Micko,
Received May 21, 1985; accepted December 3 , 1985. Address correspondence and reprint requests to Dr. P. E .
Gallant at Laboratory of Preclinical Studies, National Institute on Alcohol Abuse and Alcoholism, 12501 Washington Avenue, Rockville, MD 20852, U.S.A.
Ahhreviatiom used: BME, p-mercaptoethanol; CAP, calcium-
proteins that were sensitive to calcium-activated proteol- ysis and reacted with a monoclonal intermediate filament antibody. This intermediate filament antibody reacts with most of the axoplssmic proteins that copurify with neuro- filaments, i.e., in the order of their intermediate filament antibody staining intensity, a 60K, 65K, 220K, and 74K protein. In the cell body preparation, the intermediate fil- ament antibody labeled, in order of their staining inten- sity, a 65K, 60K, 74K, and 180K protein. In both the axo- plasmic and cell body preparations, endogenous calcium- activated proteolysis generated characteristic fragments that could be labeled with the anti-intermediate filament antibody. Key Words: Neurofilament -Phosphorylation - Calcium-activated proteol y si s - Squid - Giant axon - Intermediate filament. Gallant P. E. et al. Calcium-acti- vated proteolysis of neurofilament proteins in the squid giant neuron. J . Neurochem. 46, 1573- 1581 (1986).
1978; Pant et al., 19790; Pant and Gainer, 1980) ac- tivities. Phosphorylated neurofilament proteins are present in mammalian brain (Julien and Mushynski, 1981) and squid (Pant et al., 1979b,c) axons in vivo, but are not found in a phosphorylated form in many neuronal perikarya (Goldstein et al., 1983; Stern- berger and Sternberger, 1983). However, it is not clear what function neurofilament phosphorylation serves. It has been suggested that phosphorylation may control the assembly of neurofilament proteins into formed neurofilaments (Wong et al., 1984), or that it might stabilize the neurofilaments into com- pact structures in the axon (Sternberger and Stern- berger, 1983).
activated protease; HEPES, N-2-hydroxyethylpiperazine-N'-2- ethanesulfonic acid; HMW, high molecular weight; IFA, inter- mediate filament antibody; NF-60 and NF-220, neurofilament polypeptides with molecular sizes of 60 and 220 kilodaltons, re- spectively; PAGE, polyacrylamide gel electrophoresis; SDS, so- dium dodecyl sulfate.
1573

1574 P. E. GALLANT ET AL.
One of the proposed functions of calcium-acti- vated proteolysis is that limited proteolysis of some neurofilament proteins during axonal transport leads to an altered structure and perhaps function of the neurofilaments along the length of the axon (Nixon et al., 1982). Another proposal is that CAP activity is involved in the turnover of neurofilament proteins in nerve terminals (Lasek and Hoffman, 1976), and is thus one mechanism of regulating ax- onal growth (Lasek and Black, 1977). A third pro- posal is that CAP catalyzes the hydrolysis of neu- rofilament proteins during neuronal degeneration (Schlaepfer, 1974; Schlaepfer and Micko, 1978).
The present experiments were undertaken to de- termine the ability of endogenous calcium-activated proteolysis and phosphorylation to modify squid neurofilament proteins in vitro and to identify the products of calcium-activated proteolysis. The ad- vantages of using the squid giant neuron are that the enzymes and substrates can be extruded from the giant axon, thus avoiding contamination of the preparation with nonaxonal enzymes, which may become nonspecifically attached during the ho- mogenization of whole tissue. Second, enzyme ac- tivities and substrates present in axoplasm and axoplasmic preparations can be compared with the activities and substrates present in the region of the stellate ganglion, which contains the cell bodies of the giant axon.
In this study, we show that kinase and CAP ac- tivities remain attached to a neurofilament-enriched cytoskeleton preparation prepared from axoplasm. The kinase in this preparation catalyzes the incor- poration of 32P into specific regions on the neuro- filament proteins, and activation of the CAP results in the cleavage of the neurofilament proteins and the release of characteristic soluble and pelletable fragments. These proteolytic fragments were ana- lyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), protein staining, 32P- autoradiography, and immunochemical procedures, and the results are considered in relation to the neurofilament structures in the squid giant axon.
MATERIALS AND METHODS Live squid were obtained from the Marine Resources
Department of the Marine Biological Laboratory (Woods Hole, MA, U.S.A.). They were kept in flowing seawater and used within 24 h of capture. The area in the stellate ganglion that contains the cell bodies that give rise to the giant axon was dissected free f rom the ganglion in flowing seawater and then rinsed in EGTA-containing sa- line before homogenization in 2 volumes of experimental buffer (buffer E)/volume of tissue. Buffer E consisted of 0.1 - 1 .0 mM EGTA, 10 mM MgCI,, and 200 mM N-2-hy- droxyethylpiperazine-N’-2-ethanesulfonic a c i d (HEPES), p H 7.3. Buffer E was chosen because i t could support phosphorylation and control the p H shift that would otherwise result from the release of H’ caused by addition of CaClz to EGTA. Dissection of the giant axon
J . Nrrr,oclzrin.. V o l . 46, No. 5 , 1986
and extrusion of the axoplasm were performed as de- scribed by Lasek (1974), except that the intact axon was prerinsed in a buffered saline solution containing 5 mM EGTA to prevent calcium contamination during the ex- trusion process. Extruded axoplasm was then placed into ice-cold experimental buffer E . After 0.5-2.0 h of incu- bation on ice, the axoplasm was homogenized with 4-8 strokes in a ground glass microhomogenizer. To separate supernatant from pellet, the homogenate was centrifuged at 100,000 g for 1 h.
To prepare the neurofilament-enriched cytoskeletal ex- tract, axoplasm was extruded into ice-cold extraction buffer ( 1 mM EGTA, 450 mM NaF, 100 mM HEPES, and 1 % Triton X-100, p H 7 . 5 ) , homogenized, and then centri- fuged at 100,000 g for 30 min. The pellet was rinsed briefly and resuspended in 2 volumes of buffer E (without or with EGTA)/volume of original axoplasm. Morpholog- ical analysis of this preparation by negative staining showed an enrichment of neurofilaments (Pant et al., 1986).
Phosphorylation was initiated in buffer E by addition of 10-20 pCi of [Y-~’P]ATP (27 Ciimmol in 5-10 kl of water), obtained from New England Nuclear, to 50- 100 p1 of homogenate. The mixture was incubated for various intervals from 3 to 30 min, and the reaction was stopped by addition of an equal volume of SDS-BME buffer [2% SDS, 5% P-mercaptoethanol (BME), and 100 mM ‘Iris- sulfate, pH 7.01. In some experiments, the phosphory- lated proteins in the homogenate were subjected to cal- cium-activated proteolysis before SDS buffer was added. Calcium-activated proteolysis was initiated by addition of 6 mM CaCI, to the homogenate in buffer E. The protease was then allowed to act for varying intervals, and the re- action was stopped by addition of an equal volumc of SDS-BME buffer. When phosphorylation or centrifuga- tion was performed after calcium-activated proteolysis, EGTA was added in excess of the calcium to stop proteol- ysis.
SDS-PAGE was performed on polyacrylamide gels ac- cording to the method described by Neville (1971). Fol- lowing electrophoresis, the gels were stained with Coo- massie Brilliant Blue or with silver (Merril et al., 1980). Apparent molecular weights were determined using Biorad molecular weight standards. Autoradiography was performed by placing a dried gel against a Kodak X-omat film in a n X-omat casset te with intensifying screens and exposing the film for from 0.5 h to 4 days. Density under each band was digitally integrated after scanning with an Optronics scanner. Long and short au- toradiographic exposures of each gel were analyzed to quantify both the lightly and the heavily phosphorylated polypeptides.
Immunostaining was performed after proteins were transferred from a 7.5% acrylamide gel onto nitrocellu- lose paper. Transfer was carried out in a Hoefer Trans- phore unit for 15- 18 h at 1 5 T , 60 Volts, and 1,000-2,000 mA. The paper was then removed, the reaction was blocked with fetal calf serum for 18 h, and then a 1:500 dilution of anti-intermediate filament mouse monoclonal antibody (anti-IFA; Pruss et al., 1981) was applied (in 10% serum in phosphate-buffered saline) overnight. After 10- 12 washes in phosphate-buffered saline, the second antibody (peroxidase-labeled goat anti-mouse antibody; Boehringer Mannheim) was applied for 2 h. The paper was then washed in Tris-buffered saline (20 mM Tris and 0.9% NaC1, p H 7.2) 10-12 times, and the bound peroxi-
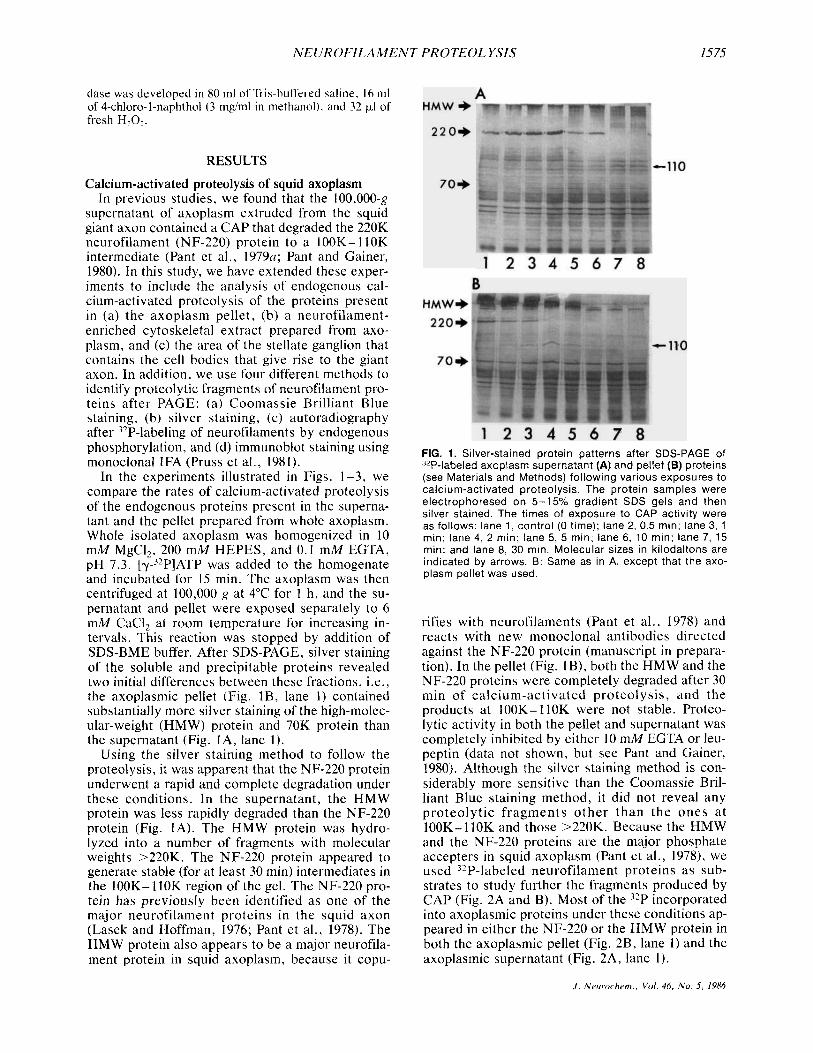
NEUROFILAMENT PROTEOL YSIS 1575
dase was developed in 80 in1 ofTris-buffered saline, 16 ml of 4-chloro-1-naphthol (3 mgiml in methanol), and 32 pI of fresh H,02.
RESULTS
Calcium-activated proteolysis of squid axoplasm In previous studies, we found that the 100,000-g
supernatant of axoplasm extruded from the squid giant axon contained a CAP that degraded the 220K neurofilament (NF-220) protein to a IOOK- 11OK intermediate (Pant et al., 1979~ ; Pant and Gainer, 1980). In this study, we have extended these exper- iments to include the analysis of endogenous cal- cium-activated proteolysis of the proteins present in (a) the axoplasm pellet, (b) a neurofilament- enriched cytoskeletal extract prepared from axo- plasm, and (c) the area of the stellate ganglion that contains the cell bodies that give rise to the giant axon. In addition, we use four different methods to identify proteol ytic fragments of neurofilament pro- teins after PAGE: (a) Coomassie Brilliant Blue staining, (b) silver staining, (c) autoradiography after 3’P-labeling of neurofilaments by endogenous phosphorylation, and (d) immunoblot staining using monoclonal IFA (Pruss et al., 1981).
In the experiments illustrated in Figs. 1-3, we compare the rates of calcium-activated proteolysis of the endogenous proteins present in the superna- tant and the pellet prepared from whole axoplasm. Whole isolated axoplasm was homogenized in 10 mM MgCI2, 200 mM HEPES, and 0.1 mM EGTA, pH 7.3. [Y-~’P]ATP was added to the homogenate and incubated for 15 min. The axoplasm was then centrifuged at 100,000 g at 4°C for 1 h , and the su- pernatant and pellet were exposed separately to 6 mM CaC12 at room temperature for increasing in- tervals. This reaction was stopped by addition of SDS-BME buffer. After SDS-PAGE, silver staining of the soluble and precipitable proteins revealed two initial differences between these fractions, i.e., the axoplasmic pellet (Fig. lB , lane 1) contained substantially more silver staining of the high-molec- ular-weight (HMW) protein and 70K protein than the supernatant (Fig. IA, lane 1).
Using the silver staining method to follow the proteolysis, it was apparent that the NF-220 protein underwent a rapid and complete degradation under these conditions. In the supernatant, the HMW protein was less rapidly degraded than the NF-220 protein (Fig. IA). The HMW protein was hydro- lyzed into a number of fragments with molecular weights >220K. The NF-220 protein appeared to generate stable (for at least 30 min) intermediates in the 100K- 110K region of the gel. The NF-220 pro- tein has previously been identified as one of the major neurofilament proteins in the squid axon (Lasek and Hoffman, 1976; Pant et al., 1978). The HMW protein also appears to be a major neurofila- ment protein in squid axoplasm, because it copu-
FIG. 1. Silver-stained protein patterns after SDS-PAGE of 32P-labeled axoplasm supernatant (A) and pellet (B) proteins (see Materials and Methods) following various exposures to calcium-activated proteolysis The protein samples were electrophoresed on 5-15% gradient SDS gels and then silver stained The times of exposure to CAP activity were as follows lane 1, control (0 time); lane 2, 0 5 min, lane 3, 1 min, lane 4, 2 mm, lane 5, 5 min, lane 6, 10 min, lane 7, 15 min, and lane 8, 30 min Molecular sizes in kilodaltons are indicated by arrows B Same as in A, except that the axo- plasm pellet was used
rifies with neurofilaments (Pant et al., 1978) and reacts with new monoclonal antibodies directed against the NF-220 protein (manuscript in prepara- tion). In the pellet (Fig. IB), both the HMW and the NF-220 proteins were completely degraded after 30 min of calcium-activated proteolysis, and the products at 100K-110K were not stable. Proteo- lytic activity in both the pellet and supernatant was completely inhibited by either 10 mM EGTA or leu- peptin (data not shown, but see Pant and Gainer, 1980). Although the silver staining method is con- siderably more sensitive than the Coomassie Bril- liant Blue staining method, it did not reveal any proteolytic f ragments o ther than the ones a t 100K- 110K and those >220K. Because the HMW and the NF-220 proteins are the major phosphate accepters in squid axoplasm (Pant et al., 1978), we used 32P-labeled neurofilament proteins as sub- strates to study further the fragments produced by CAP (Fig. 2A and B). Most of the 32P incorporated into axoplasmic proteins under these conditions ap- peared in either the NF-220 or the HMW protein in both the axoplasmic pellet (Fig. 2B, lane 1) and the axoplasmic supernatant (Fig. 2A, lane 1).
J . Nrurochem., Vol. 46, No. 5 , 1986

1576 P . E. GALLANT E T A L .
FIG. 2. Phosphoprotein pattern following calcium-activated proteolysis of axoplasm. A: Autoradiograph of the superna- tant (corresponding to the gel in Fig. 1A). B: Autoradiograph of the pellet fraction (corresponding to the gel in Fig. 1B).
The data illustrated in Fig. 2A and B are autora- diographs of the same silver-stained gels illustrated in Fig. 1A and B, respectively. The '2P-autoradio- graphs provide a more complete and sensitive assay of the intermediates of neurofilament protein degradation produced by the CAP. As was found by the silver staining method (Fig. IA and B), the phosphorylated HMW and NF-220 proteins were more rapidly degraded in the pellet (Fig. 2B) than in the supernatant (Fig. 2A) fractions. Despite the dif- ferent rates of proteolysis, both preparations re- vealed a similar pattern of intermediate product formation. In the supernatant (Fig. 2A), these in- cluded phosphorylated intermediates at 160K, llOK, IOOK, 90K, 80K, and 35K. The liOK inter- mediate was the most intensely phosphorylated in- termediate. The silver-stained HMW intermediates (>220K) in the supernatant corresponded to the phosphorylated intermediates in this molecular weight range. Although the pellet contained consid- erably more silver staining in the HMW band than the supernatant, the 32P-labeling in the supernatant was comparable to that found in the pellet (Fig. 2).
A relationship between the degradation of the
32P-labeled NF-220 and the phosphorylated inter- mediates is suggested by the quantitative densi- tometry shown in Fig. 3. For both the supernatant (Fig. 3A) and pellet (Fig. 3B) preparations, the rapid breakdown of the NF-220 protein is accompa- nied by the simultaneous appearance of the minor 160K and major 100K-l lOK proteins (early products), suggesting that these are formed imme- diately by cleavage of the NF-220 protein. In con- trast, the minor 90K, 80K, and 35K proteins appear later (late products), with time courses that suggest they may be formed by subsequent proteolysis of the 160K and llOK intermediates. Some of these latter products may also be derived from the HMW protein. Although the rates of proteolysis varied from preparation to preparation and could be in- creased by addition of higher concentrations of CaCI2, the sequential qualitative pattern of frag- ment production was the same in pellet and super- natant preparations in four separate experiments conducted during two different summers at the Ma- rine Biological Laboratory.
Calcium-activated proteolysis of neurofilarnent-enriched cytoskeletal proteins
To evaluate the solubility of the proteolytic frag- ments produced by calcium-activated proteolysis of the neurofilament proteins, experiments were per- formed on a neurofilament-enriched cytoskeletal preparation, which was made by homogenizing axoplasm in 450 mM NaF, 1% Triton X-100. and 50 mM HEPES (pH 7.51, centrifuging the homogenate at 100,000 g, and resuspending the pellet in buffer E. The use of NaF (as opposed to NaCl or KCI) in the buffer has been shown to stabilize neurofila- ment proteins under these conditions (Baumgold el al., 1981). This preparation has also been examined by negative staining and has been found to be enriched in long 10-nm neurofilaments that contain short projecting side arms (Pant et al., 1986). This preparation contained both protein kinase and CAP activity. This is illustrated in Figs. 4 and 5 .
The Coomassie Brilliant Blue staining pattern of the gel of the neurofilament-enriched preparation (Fig. 4A, lane I ) shows a relative enrichment of the NF-220 and neurofilament 60K (NF-60) proteins compared with whole axoplasm (Fig. 4A, lane 3), largely due to the loss of low-molecular-weight pro- teins. Stimulation of the CAP by addition of CaCI, (5 mM) to the cytoskeletal preparation produced a loss of Coomassie Brilliant Blue staining density at 220K and an increased staining density at IOOK (Fig. 4A, lane 2). This lOOK proteolytic fragment comigrated with the IOOK fragment produced by addition of 5 mM CaCI, to whole axoplasm (Fig. 4A, lane 4). Therefore, the cytoskeletal preparation retained the same CAP activity with the same sub- strate preferences present in whole axoplasm. The
.I. Neur.o<.lrcrii., Vo l . 46. N o . 5 , 1986

NEUROFILAMENT PROTEOL YSIS I577
A 8
NF breakdown I00
NF-220 N F - 2 2 0
100
early products early products
50
110 160 160
50 late products late products
5 ' 10 1.5 5 10 15
TIME IMin) TIME (Min)
0 N PI Y z
i f I- -
i-
W Z U (L W
.a.
resolution of the 100K- 110K region of the gel was better in Fig. 4 than in Figs. 1 and 2 because of the use here of a nongradient 5% gel (see legend to Fig. 4).
This cytoskeletal preparation aIso retained the protein kinase activity, and, as illustrated in Fig. 4B, in the cytoskeletal preparation (Fig. 4B, lane l), as in whole axoplasm (Fig. 4B, lane 3), the HMW and NF-220 proteins were the major phosphate ac- ceptors. When phosphorylation was performed after the calcium-activated proteolysis step in ei- ther the axoplasmic (Fig. 4B, lane 4) or the cyto- skeletal preparation (Fig. 4B, lane 2), the kinase re- mained very active and phosphorylated a number of breakdown products, including products at llOK, IOOK, 160K, and some HMW intermediates >220K. These phosphorylated proteolytic products
FIG. 4. Comparison of the SDS- PAGE Coomassie Brilliant Blue- stained pattern (A) and autora- diographs of the phosphorylation pattern (6) in a neurofilament- enriched cytoskeletal prepara- tion (lanes 1 and 2) and in whole axoplasm (lanes 3 and 4). The samples in lanes 2 and 4 were ex- posed for 30 min to 5 mM CaCI,, and phosphorylation was per- formed after proteolysis had been stopped by EGTA. In this experiment proteins, were sepa- rated on an SDS-5% polyacryl- amide gel to separate better the 110K from the 100K protein, and then the gels were stained with Coomassie Brilliant Blue. The gel was then dried and exposed for autoradiography.
FIG. 3. Quantitative analysis of the time courses of neurofilament (NF) degrada- tion and product formation following cal- cium-activated proteolysis of 32P-labeled axoplasm. Long- and short-exposure au- toradiographs of the gels shown in Fig. 1 were density scanned with an Optronics scanner and Gould Image array pro- cessor. The density under each band was digitally integrated and expressed as a percentage of the initial (time 0) inte- grated density of the NF-220 protein. A: Supernatant (see Fig. 2A). 6: Axoplasmic pellet (see Fig. 28). The molecular sizes of the original protein or products (in ki- lodaltons) are shown on the right for each curve.
were virtually identical in the neurofilament- enriched (Fig. 4B, lane 2) and the unfractionated axoplasm (Fig. 4B, lane 4) preparations. The only difference was that when phosphorylation was per- formed in axoplasm (Fig. 4B, lane 4) , the lOOK fragment was much less phosphorylated than the 110K fragment, whereas when phosphorylation was performed in the cytoskeletal preparation (Fig. 4B, lane 2), the relative incorporation of 32P into the lOOK fragment increased dramatically.
The data illustrated in Fig. 5 evaluate the solu- bility of the neurofilament proteins and the proteo- lytically generated intermediates seen in Fig. 4. For this purpose, the neurofilament-enriched prepara- tion was used. In this preparation, the neurofila- ment proteins were found primarily in the pelleted fraction (HMW, 220K, and'60K in Fig. 5A and B,
J . Neurochem., Voi. 46, No. 5, 1986
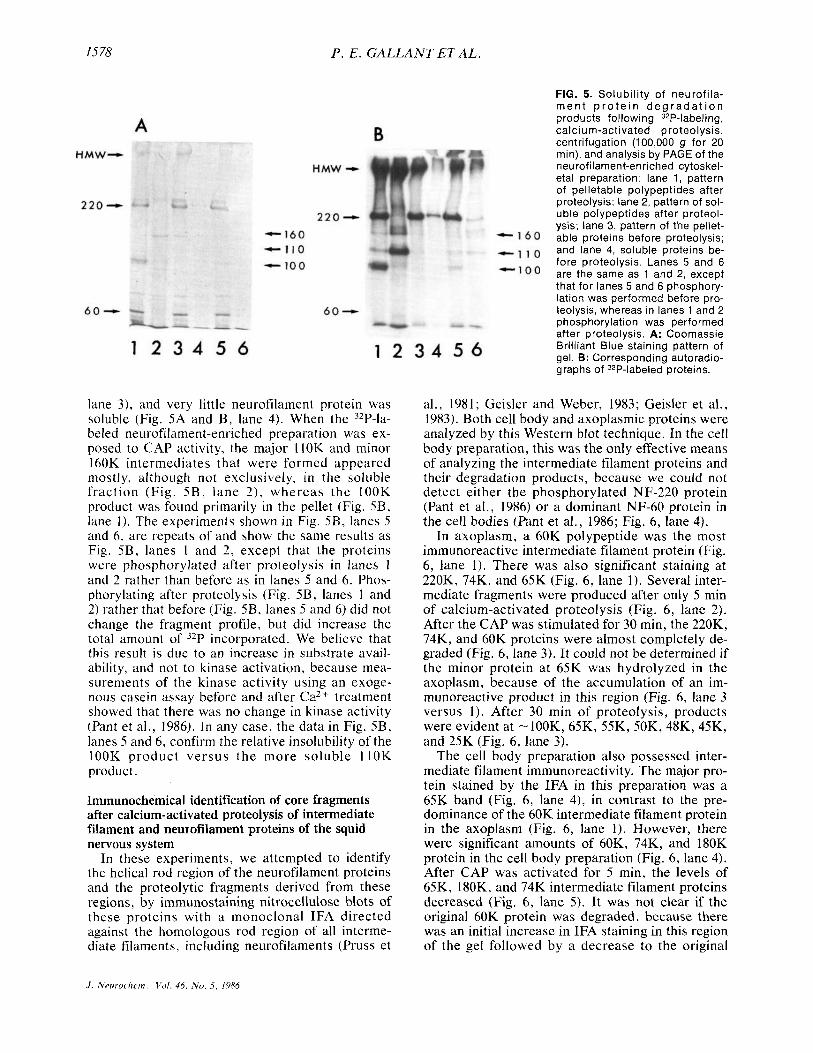
1578 P . E. GALLANT ET AL.
lane 3 ) , and very little neurofilament protein was soluble (Fig. 5A and B , lane 4). When the 32P-la- beled neurofilament-enriched preparation was ex- posed to CAP activity, the major llOK and minor 160K intermediates that were formed appeared mostly, although not exclusively, in the soluble fraction (Fig. 5 B , lane 2) , whereas the IOOK product was found primarily in the pellet (Fig. 5B, lane 1). The experiments shown in Fig. SB, lanes 5 and 6, are repeats of and show the same results as Fig. SB, lanes 1 and 2, except that the proteins were phosphorylated after proteolysis in lanes I and 2 rather than before as in lanes S and 6. Phos- phorylating after proteolysis (Fig. 5B, lanes 1 and 2) rather that before (Fig. 5B, lanes S and 6) did not change the fragment profile, but did increase the total amount of 32P incorporated. We believe that this result is due to an increase in substrate avail- ability, and not to kinase activation, because mea- surements of the kinase activity using an exoge- nous casein assay before and after Ca2+ treatment showed that there was no change in kinase activity (Pant et al., 1986). In any case, the data in Fig. 5B, lanes 5 and 6, confirm the relative insolubility of the lOOK product versus the more soluble l lOK product.
Immunochemical identification of core fragments after calcium- activated proteolysis of intermediate filament and neurofilament proteins of the squid nervous system
In these experiments, we attempted to identify the helical rod region of the neurofilament proteins and the proteolytic fragments derived from these regions, by immunostaining nitrocellulose blots of these proteins with a monoclonal IFA directed against the homologous rod region of all interme- diate filaments, including neurofilaments (Pruss et
FIG. 5. Solubil ity of neurofi la- rnen t p r o t e i n d e g r a d a t i o n products following 32P-labeling, calcium-activated proteolysis, centrifugation (100,000 g for 20 rnin), and analysis by PAGE of the neurofilarnent-enriched cytoskel- eta1 preparation: lane 1, pattern of pelletable polypeptides after proteolysis; lane 2, pattern of sol- uble polypeptides after proteol- ysis; lane 3, pattern of the pellet- able proteins before proteolysis; and lane 4, soluble proteins be- fore proteolysis. Lanes 5 and 6 are the same as 1 and 2, except that for lanes 5 and 6 phosphory- lation was performed before pro- teolysis, whereas in lanes 1 and 2 phosphorylation was performed after proteolysis. A: Coomassie Brilliant Blue staining pattern of gel. 8: Corresponding autoradio- graphs of 32P-labeled proteins.
al., 1981; Geisler and Weber, 1983; Geisler et al., 1983). Both cell body and axoplasmic proteins were analyzed by this Western blot technique. In the cell body preparation, this was the only effective means of analyzing the intermediate filament proteins and their degradation products, because we could not detect either the phosphorylated NF-220 protein (Pant et al., 1986) or a dominant NF-60 protein in the cell bodies (Pant et al., 1986; Fig. 6, lane 4).
In axoplasm, a 60K polypeptide was the most immunoreactive intermediate filament protein (Fig. 6, lane 1). There was also significant staining at 220K, 74K, and 65K (Fig. 6, lane 1). Several inter- mediate fragments were produced after only 5 min of calcium-activated proteolysis (Fig. 6, lane 2). After the CAP was stimulated for 30 min, the 220K, 74K, and 60K proteins were almost completely de- graded (Fig. 6, lane 3). It could not be determined if the minor protein at 65K was hydrolyzed in the axoplasm, because of the accumulation of an im- munoreactive product in this region (Fig. 6, lane 3 versus 1). After 30 min of proteolysis, products were evident at -100K, 65K, 55K, 50K, 48K, 45K, and 25K (Fig. 6, lane 3).
The cell body preparation also possessed inter- mediate filament immunoreactivity. The major pro- tein stained by the IFA in this preparation was a 65K band (Fig. 6, lane 4), in contrast to the pre- dominance of the 60K intermediate filament protein in the axoplasm (Fig. 6, lane 1). However, there were significant amounts of 60K, 74K, and 180K protein in the cell body preparation (Fig. 6, lane 4). After CAP was activated for 5 min, the levels of 65K, 180K, and 74K intermediate filament proteins decreased (Fig. 6, lane 5). It was not clear if the original 60K protein was degraded, because there was an initial increase in IFA staining in this region of the gel followed by a decrease to the original
J . Nmruclwm. , V o l . 44, N u . 5 , 1984

NEUROFILAMENT PROTEOL YSIS 1579
FIG. 6. Immunoblot, using the anti-IFA, of whole axoplasm (lanes 1-3) and the cell body region of the stellate ganglion (lanes 4-6) before and after proteolysis. Proteins were transferred to nitrocellulose paper for immunoblotting after SDS-PAGE on 7.5% gels: lane 1, axoplasm before proteol- ysis; lane 2, axoplasm after 5 min of treatment with CAP; lane 3, axoplasrn after 30 min of treatment with CAP: lane 4, cell bodies before proteolysis; lane 5, cell bodies after 5 min of treatment with CAP; and lane 6, cell bodies after 30 rnin of treatment with CAP. The faintly staining bands between lanes 3 and 4 are of blue dye-coupled protein markers (BRL Inc.) used to align the two nitrocellulose strips. Because axoplasm was much more immunoreactive than the cell body extract, the nitrocellulose paper was cut, and the two halves were developed separately.
(Fig. 6, lane 1) levels. After 30 min, with the excep- tion of the 60K immunoreactivity, most of the in- termediate filament proteins were reduced to low- molecular-weight immunoreactive products (Fig. 6, lane 6) that migrated at -25K.
DISCUSSION
Calcium-activated proteol ysis of whole squid axoplasm and of a neurofilament-enriched cyto- skeletal preparation derived from axoplasm pro- duces characteristic proteolytic polypeptide products. In this article, we have used four dif- ferent analytical methods to examine these products.
One of the major neurofilament proteins in squid axoplasm, the NF-220 protein, is particularly vul- nerable to calcium-activated proteolysis. Two of the products derived from the proteolysis of the NF-220 protein are a lOOK protein and a 110K pro- tein. Both of these protein products appear to be phosphorylated (Figs. 4 and 5). Further analysis of these proteolytic fragments revealed that the IOOK protein coprecipitates with the intact NF-220 pro- tein, whereas the llOK protein is soluble in par- tially proteolyzed neurofilament-enriched cytoskel- eta1 preparations (Fig. 5 , lanes I and 2). The ability of the lOOK fragment to coprecipitate with the un- degraded neurofilaments suggests that this frag- ment maintains its association (via interpeptide coiled-coils; Geisler et al., 1983) with the assem-
bled neurofilaments. In contrast, the I IOK protein is more soluble, suggesting that it might be pe- ripherally located on the NF-220 protein. Another indication that the 110K fragment is peripherally located is that the llOK fragment is effectively phosphorylated by the endogenous kinase in whole axoplasm (Fig. 4, lane 4), whereas the IOOK frag- ment, like core regions present in keratin (Steinert et al., 1982) and mammalian (Julien and Mu- shynski, 1982) or myxicola (Eagles et al., 1981) neurofilaments, is less accessible for phosphoryla- tion (Fig. 4, lane 4).
The above information about the NF-220 and the CAP products at lOOK and llOK provides some new insights into the structure of the intact NF-220 protein. The NF-220 protein is believed to provide the side arms found in the 10-nm neurofilaments in axoplasm (Zackroff et a]., 1984), and we suggest that the IOOK product is from the N-terminal region of the NF-220 protein, which is involved in neuro- filament assembly (Geisler et al., 1983), whereas the llOK product represents the side arm projec- tion of this molecule. Rapid calcium-activated pro- teolysis of a highly phosphorylated peripherally lo- cated neurofilament polypeptide (NF-220 protein) also occurs in the mammalian nervous system (Zimmerman and Schlaepfer, 1982; Paggi and Lasek, 1984; Schlaepfer et al., 1984, 1985). This proteolysis might be expected to disturb the cyto- skeletal structure (Schlaepfer and Micko, 1978), because these phosphorylated neurofilament side arms are thought to participate in complex forma- tion between neurofilaments and other cytoskeletal elements (Runge et al., 1981).
The other two major neurofilament proteins that copurify with neurofilaments are the NF-60 protein and the HMW protein. The NF-60 protein present in the squid axoplasm is believed to be the back- bone of neurofilaments in the squid giant axon (Zackroff and Goldman, 1980; Zackroff et al., 1984), whereas the exact structural relationship of the HMW protein to the neurofilament is unclear at present. What is clear is that the HMW protein (Pant et al., 1978) is phosphorylated by the neuro- filament kinase activity in axoplasm (Pant et al., 1978, 1986) and is proteolyzed by axoplasmic CAP (Fig. 1). In addition, we have recently prepared several monoclonal antibodies that are directed against the NF-220 protein and that cross-react with the HMW protein but no other axoplasmic proteins, including the NF-60 protein (manuscript in preparation). Understanding the role of the HMW protein in squid neurofilaments will require further study.
We also studied the effect of CAP activity on the intermediate filament proteins in a stellate ganglion preparation, which is enriched in the cell bodies that give rise to the giant axon. However, because the neurofilament and other intermediate filament
J . Neurochem., Vol. 46, N o . 3. 1986

1580 P. E . G A L L A N T ET AL .
proteins have not yet been characterized in this preparation, we used the anti-IFA (monoclonal an- tibody) to study the intermediate filament proteins present in this preparation (Fig. 6, lanes 4-6). This antibody succeeded in staining a 65K protein in- tensely and several other proteins (60K, 74K, and 180K) less intensely. The greater immunostaining of the 65K versus the 60K in the ganglion prepara- tion corresponds to the greater abundance of the 65K protein found in this preparation on stained gels (Tytell et al., I984), but differs from the situa- tion in axoplasm, where the staining of the 60K protein dominates (Fig. 6, lanes 1 and 4). Calcium- activated proteolysis in the cell body preparation leads to degradation of the 180K, 74K, and 65K im- munoreactive proteins, leaving behind significant reactivity in the 60K and 25K proteins (Fig. 6, lanes 4-6). The ganglion data are more difficult to inter- pret than the axoplasm data, because the ganglion preparation contains, in addition to the neuronal cell bodies, other cellular elements (e.g., glia and axons) that also contain intermediate filaments. However, the only intermediate filaments that are known to be >100K are neurofilaments. Therefore, it is likely that the 180K immunoreactive protein found in the cell body preparation (Fig. 6, lane 4) is the unphosphorylated form in the cell bodies of the NF-220 protein found in the axoplasm, which is highly phosphorylated. In the mammalian nervous system, dephosphorylation of the 200K neurofila- ment protein reduces its apparent molecular weight to 150K (Carden et al., 1985). It is also interesting to note that the 180K protein is rapidly proteolyzed by CAP, as is the 220K protein (Fig. 6).
With respect to the intermediate filament pro- teins at 60K and 65K, we find that the total immu- noreactivity is greater in axoplasm than in the cell body preparation. This observation is consistent with the finding that there is more total protein at 60K and 65K in the axon than in the cell body prep- aration in another molluscan nervous system, i.e., in Aplysiu (Drake and Lasek, 1985). However, in contrast to the Aplysia studies, where the 60K/65K ratio in the cell body was greater than in the axons (Drake and Lasek, 1984), in the squid system the reverse was true (Fig. 6, lanes 1 and 4). This differ- ence may be either due to intrinsic differences be- tween the two molluscan species or due to the dif- ferent contamination by glial elements in the squid ganglion, Aplysia cell body, and Aplysia axonal preparations. Only the squid axoplasm represents a pure neuronal component. Resolution of these issues in the cell bodies, in both species, will re- quire the development of antibody reagents that are specific for the various identified forms of the neu- rofilaments (e.g., Carden et al., 1985; Schlaepfer et al., 1984, 1985) and the use of these characterized antibodies in immunocytochemical studies.
Acknowledgment: We thank Dr. lchiji Tasaki for his in- terest and support.
REFERENCES Baumgold J . , Terakawa S . , Iwasa K. , and Gainer H. (1981)
Membrane-associated cytoskeletal proteins in squid giant axons. J . Neurochem. 36, 759-764.
Carden M. J., Schlaepfer W. W., and Lee V. M. Y. (1985) The structure, biochemical properties, and immunogenicity of neurofilament peripheral regions are determined by phos- phorylation state. J . B i d . Chem. 260, 9805-9817.
Drake P. E. and Lasek R. J. (1984) Regional differences in the neuronal cytoskeleton. J . Neurmci . 4, 1173-1 186.
Eagles P. A. M., Gilbert D. S. , and Maggs A. (1981) The loca- tion of phosphorylation sites and Ca’+-dependent cleavage sites on the major neurofilament polypeptides from myxi- cola infundibulum. Biachem. J . 199, 101 - I I I.
Geisler N. and Weber K. (1983) Amino acid sequence data on glial fibrillary acidic protein (GFA) implications for the sub- division of intermediate filaments into epithelial and non-ep- ithelial members. EMBO J . 2, 2059-2063.
Geisler N., Kaufmann E., Fisher S., Plessmann U., and Weber K. (1983) Neurofilament architecture combines structural principles of intermediate filaments with carboxy-terminal extensions increasing in size between triplet proteins.
Gilbert D. S. , Newby B. J., and Anderton B. H. (1975) Neuro- filament disguise, destruction and discipline. Nature 265, 586-589.
Goldstein M. E . , Sternberger L . A , , and Sternberger N. H. (1983) Microheterogeneity (“neurotypy”) of neurofilament proteins. Proc. Nu//. Acud. Sci. USA 80, 3101-3105.
Hodge A. J. and Adelman W. J. (1979) The neuroplasmic net- work in loligo and hermassenda neurons. J . Ultrustruct. Res. 70, 220-241.
Hoffman P. N., Griffin J. W., and Price D. L. (1984) Control of axonal caliber by neurofilament transport. J . Cell Biol. 99, 704 - 7 14.
Julien J. P. and Mushynski W. E. (1981) A comparison of in vitro and in vivo-phosphorylated neurofilament polypeptides. J . Neurochem. 37, 1579-1585.
Julien J. P. and Mushynski W. E. (1982) Multiple phosphoryla- tion sites in mammalian neurofilament polypeptides. J . Bid. Chem. 257, 10467- 10470.
Lasek R. J. (1974) Biochemistry of the squid giant axon, in A Guide to Laborutoiy Use of the Squid, Loligo peulii (Arnold J. M., ed), pp. 69-74. Marine Biological Laboratory, Woods Hole, Massachusetts.
Lasek R . J. and Black M. M. (1977) How do axons stop growing? Some clues from the metabolism of the proteins in the slow component of axonal transport, in Mechanisms, Regulations and Special Functions of Protein Synthesis in the Bruin (Roberts S. , Lajtha A , , and Gispen W. H. , eds), pp. 161- 169. Elsevier-North Holland, New York.
Lasek R . J. and Hoffman P. N . (1976) The neuronal cyto- skeleton, axonal transport and axonal growth, in Cell Mn- tility (Goldman R., Pollard T., and Rosenbaum E., eds), pp. 1021 - 1049. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
Lazarides E. (1980) Intermediate filaments as mechanical inte- grators of cellular space. Nature 283, 249-256.
Merril C. R., Goldman D., Sedman S. A., and Ebert M. H. (1980) Ultrasensitive stain for proteins in polyacrylamide gels shows regional variation in cerebrospinal fluid proteins. Science 211, 1437- 1438.
Neville D. (197 I ) Molecular weight determination of protein-do- decyl sulfate complexes by gel electrophoresis in a discon- tinuous buffer system. J . Biol. Chem. 246, 6328-6334.
Nixon R. A., Brown B. A , , and Marotta C. A. (1982) Posttrans-
EMBO J . 2, 1295-1302.
J . Neurochem., Vol. 46, No. 5 , 1986

NEUROFILAMENT PROTEOL YSIS 1581
lational modification of a neurofilament protein during axo- plasmic transport: implications for regional specialization of CNS axons. J . Cell B i d . 94, 150-158.
Paggi P. and Lasek R. J. (1984) Degradation of purified neuro- filament subunits by calcium-activated neutral protease: characterization of the cleavage products. Neurochem. Inf. 6, 589-597.
Pant H. C. and Gainer H. (1980) Properties of a calcium-acti- vated protease in squid axoplasm which selectively de- grades neurofilament proteins. J . Neurobiol. l l , 1 - 12.
Pant H. C., Shecket G., Gainer H., and Lasek R. J. (1978) Neurofilament is phosphorylated in the squid giant axon. J . Cell Biol. 78, R23-R27.
Pant H. C., Terakawa S . , and Gainer H. (1979~) A calcium-acti- vated protease in squid axoplasm. J . Neurochem. 32,
Pant H. C., Yoshioka T., Tasaki I., and Gainer H. (1979b) Diva- lent cation dependent phosphorylation of proteins in the squid giant axon. Bruin Res. 162, 303-313.
Pant, H. C., Terakawa S. , Yoshioka T, Tasaki I., and Gainer H. (1979~) Evidence for the utilization of extracellular (y3’) ATP for the phosphorylation of intracellular proteins in the squid giant axon. Biochim. Biophys. Acta 582, 107-1 14.
Pant H. C., Gallant P. E., and Gainer H. (1986) Characterization of a cyclic-nucleotide and calcium independent-neurofila- ment protein kinase activity in axoplasm from the squid giant axon. J. B i d . Chem. (in press).
Pruss R. M., Mirsky R., Raff M. C., Thorp R.. Dowding A. J., and Anderton B. H. (1981) All classes of intermediate fila- ments share a common antigenic determinant defined by a monoclonal antibody. Cell 27, 419-428.
Rubinson K. A. and Baker P. F. (1979) The flow properties of axoplasm in a defined chemical environment: influences of anions and calcium. Proc. R . SOC. Lond. 205, 323-345.
Runge M. S. , Laue T. M. , Yphantis D. A., Lifsics M. R., Saito A, , Akin M., Reinke K., and Williams R. C. (1981) ATP-in- duced formation of an associated complex between micro- tubules and neurofilaments. Proc. Nut/. Acad. Sci. U . S . A . 78, 1431-1435.
99-102.
Schlaepfer W. W. (1974) Calcium induced degeneration of axo- plasm in iaolated segments of rat peripheral nerve. Bruin Res. 136, 1-9.
Schlaepfer W. W., Lee C. , Trojanowski J. Q., and Lee V. M. Y. (1984) Persistence of immunoreactive neurofilament protein breakdown products in transected rat sciatic nerve. J . Neurochem. 43, 857-864.
Schlaepfer W. W. and Micko S . (1978) Chemical and structural changes of neurofilaments in transected rat sciatic nerve. J . Cell Biol. 78, 369-378.
Schlaepfer W. W., Lee C., Lee V. M.. and Zimmerman U. J. (1985) An immunoblot study of neurofilament degradation in situ and during calcium-activated proteolysis. J . Nertro- chem. 44, 502-509.
Steinert P. M., Wantz M. L., and Idler W. W. (1982) O-Phospho- serine content of intermediate subunits. Biochpmisrry 21,
Sternberger L. A. and Sternberger N. H . (1983) Monoclonal an- tibodies distinguish phosphorylated and nonphosphorylated forms of neurofilaments in situ. Proc. Nurl. Ac,ud. Sci. USA
Tytell M., Hill W. D., Zackroff R. V., and Goldman R. D. (1984) Regional and biochemical characterization of squid neuro- filaments. (Abstr) Truns. A m . Suc. Neirroclirm. 15 , 260.
Wong J . , Hutchison S. B., and Liem R. K. H. (1984) An iso- electric variant of the 150,000-dalton neurofilament poly- peptide: evidence that phosphorylation state affects its as- sociation with the filament. J . B i d . Chem. 259, 10867- 10874.
Zackroff R. V. and Goldman R. D. (1980) In vitro reassembly of squid brain intermediate filaments (neurofilaments): purifi- cation by assembly and disassembly. Science 208, 1152- 1155.
Zackroff R. V., Hill W. D., and Tytell M. (1984) Assembly and organization of neurofilaments in the squid giant axon. J . Cell Biol. 99, 323a.
Zimmerman U. P. and Schlaepfer W. W. (1982) Characterization of a brain calcium-activated protease that degrades neurc filament proteins. Biochemisfry 21, 3977-3983.
177- 183.
80, 6126-6130.
J . Neurorhem., Vol. 46, No. 5 , 1986




















