
Alteration of the hormonal bioactivity of parathyroid hormone-related protein (PTHrP) as a result of...
9
ELSEVIER ALTERATION OF THE HORMONAL BIOACTIVITY OF PARATHYROID HORMONE-RELATED PROTEIN (PTHrP) AS A RESULT OF LIMITED PROTEOLYSIS BY PROSTATE-SPECIFIC ANTIGEN * MASATSUGU IWAMURA, JUKKA HELLMAN, ABRAHAM T. K. COCKETT, HANS LILJA, AND STEN GERSHAGEN ABSTRACT Objectives. To discover whether the proteolytic activity of prostate-specific antigen (PSA) affects the struc- ture and function of parathyroid hormone-related protein (PTHrP), as both are abundant components of human seminal plasma. Methods. The ability of PTHrP to act as a substrate was studied by incubating a synthetic polypeptide, consisting of 34 amino acid residues of the amino-terminal domain of PTHrP, with purified PSA. The incubate was then analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, high-pressure liquid chro- matography separation, amino-terminal peptide sequencing, and mass spectrometry. The physiologic effect of the proteolytic activity of PSA on PTHrP was studied by measuring any alteration in PTHrP (1-34)-induced elevation of cyclic adenosine monophosphate (CAMP) production by UMR-106 rat osteosarcoma cells in culture. All cell culture experiments were performed with PSA and PTHrP (l-34) at physiologic concentra- tions. Results. Our data show that PSA proteolytically cleaves PTHrP (1-34) after either residue 22 or 23, gen- erating three peptide fragments. Both cleavages occur carboxy terminally of a phenylalanine residue. The CAMP production in rat osteosarcoma cells, induced by the amino-terminal portion of PTHrP (l-34), as a result of its structural similarity with parathyroid hormone (PTH), was abated by PSA in a dose- and time- dependent fashion. In contrast, heat-inactivated PSA had no effect on CAMP production. Conclusions. Our study demonstrates that PTHrP is a substrate for PSA. The cleavage of the amino-terminal portion of PTHrP completely disrupts its ability to interact with the PTH/PTHrP receptor and thus inhibits its PTH-like activity. The proteolytic processing of PTHrP by PSA may play an important role in the post-trans- lational/post-secretional regulation of prostatic PTHrP activities, which are believed to include regulation of prostate growth and differentiation. UROLOGY 48: 3 17-325, 1996. P arathyroid hormone-related protein ( PTHrP) was originally isolated from a human lung can- * This work was supported by grants from the Swedish Medical Research Council (project # 7903), the Swedish Cancer Society (projects # 3555 and 3078-B95-OZXBC), the Faculty ofMedicine at Lund University, the Research Fund and the Cancer Research Fund at the University Hospital, Malmo, the Crafoord Founda- tion, and Fundacion Federico S.A. From the Department of Urology, University of Rochester Medical Center, Rochester, New York; Department of Biotech- nology, University of Turku, Turku, Finland; and Departments of Clinical Chemistry and Urology, University of Lund, Malmb University Hospital MAS, Malmo, Sweden Reprint requests: Sten Gershagen, M.D., Department of Urol- ogy, University oflund, Malmii University Hospital MAS, S-205 02 Malmo, Sweden Submitted (Rapid Communication): April 24, 1996, accepted: April 24, 1996 COPYRIGHT 1996 BY ELSEVIER SCIENCE INC. AIL RIGHTS RESERVED cer cell line and has been considered a major hu- moral factor responsible for hypercalcemia of ma- 1ignancy.l Although the exact structure of the secreted form of PTHrP is unclear, the PTHrP gene encodes at least three isoforms of the protein, PTHrP ( l-139)) PTHrP (l-141)) and PTHrP (l- 173)) in which 8 of the first 13 amino acid resi- dues are common to parathyroid hormone (PTH) .lm3 The three PTHrP isoforms are probably precursors that are processed into several smaller peptides with distinct, region-specific activities4” The amino-terminal region of PTHrP, which is common to all three isoforms, has been rather ex- tensively characterized owing to its structural and functional similarity to PTH. Studies have shown that the PTHrP fragment ( l-34) binds to the PTH receptor and that the amino-terminal region plays 0090-4295/96/$15.00 PII SOO90-4295(96)00182-3 317
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Alteration of the hormonal bioactivity of parathyroid hormone-related protein (PTHrP) as a result of...

ELSEVIER
ALTERATION OF THE HORMONAL BIOACTIVITY OF PARATHYROID HORMONE-RELATED PROTEIN (PTHrP)
AS A RESULT OF LIMITED PROTEOLYSIS BY PROSTATE-SPECIFIC ANTIGEN *
MASATSUGU IWAMURA, JUKKA HELLMAN, ABRAHAM T. K. COCKETT, HANS LILJA, AND STEN GERSHAGEN
ABSTRACT Objectives. To discover whether the proteolytic activity of prostate-specific antigen (PSA) affects the struc- ture and function of parathyroid hormone-related protein (PTHrP), as both are abundant components of human seminal plasma. Methods. The ability of PTHrP to act as a substrate was studied by incubating a synthetic polypeptide, consisting of 34 amino acid residues of the amino-terminal domain of PTHrP, with purified PSA. The incubate was then analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, high-pressure liquid chro- matography separation, amino-terminal peptide sequencing, and mass spectrometry. The physiologic effect of the proteolytic activity of PSA on PTHrP was studied by measuring any alteration in PTHrP (1-34)-induced elevation of cyclic adenosine monophosphate (CAMP) production by UMR-106 rat osteosarcoma cells in culture. All cell culture experiments were performed with PSA and PTHrP (l-34) at physiologic concentra- tions. Results. Our data show that PSA proteolytically cleaves PTHrP (1-34) after either residue 22 or 23, gen- erating three peptide fragments. Both cleavages occur carboxy terminally of a phenylalanine residue. The CAMP production in rat osteosarcoma cells, induced by the amino-terminal portion of PTHrP (l-34), as a result of its structural similarity with parathyroid hormone (PTH), was abated by PSA in a dose- and time- dependent fashion. In contrast, heat-inactivated PSA had no effect on CAMP production. Conclusions. Our study demonstrates that PTHrP is a substrate for PSA. The cleavage of the amino-terminal portion of PTHrP completely disrupts its ability to interact with the PTH/PTHrP receptor and thus inhibits its PTH-like activity. The proteolytic processing of PTHrP by PSA may play an important role in the post-trans- lational/post-secretional regulation of prostatic PTHrP activities, which are believed to include regulation of prostate growth and differentiation. UROLOGY 48: 3 17-325, 1996.
P arathyroid hormone-related protein ( PTHrP) was originally isolated from a human lung can-
* This work was supported by grants from the Swedish Medical Research Council (project # 7903), the Swedish Cancer Society (projects # 3555 and 3078-B95-OZXBC), the Faculty ofMedicine at Lund University, the Research Fund and the Cancer Research Fund at the University Hospital, Malmo, the Crafoord Founda- tion, and Fundacion Federico S.A.
From the Department of Urology, University of Rochester Medical Center, Rochester, New York; Department of Biotech- nology, University of Turku, Turku, Finland; and Departments of Clinical Chemistry and Urology, University of Lund, Malmb University Hospital MAS, Malmo, Sweden
Reprint requests: Sten Gershagen, M.D., Department of Urol- ogy, University oflund, Malmii University Hospital MAS, S-205 02 Malmo, Sweden
Submitted (Rapid Communication): April 24, 1996, accepted: April 24, 1996
COPYRIGHT 1996 BY ELSEVIER SCIENCE INC. AIL RIGHTS RESERVED
cer cell line and has been considered a major hu- moral factor responsible for hypercalcemia of ma- 1ignancy.l Although the exact structure of the secreted form of PTHrP is unclear, the PTHrP gene encodes at least three isoforms of the protein, PTHrP ( l-139)) PTHrP (l-141)) and PTHrP (l- 173)) in which 8 of the first 13 amino acid resi- dues are common to parathyroid hormone (PTH) .lm3 The three PTHrP isoforms are probably precursors that are processed into several smaller peptides with distinct, region-specific activities4” The amino-terminal region of PTHrP, which is common to all three isoforms, has been rather ex- tensively characterized owing to its structural and functional similarity to PTH. Studies have shown that the PTHrP fragment ( l-34) binds to the PTH receptor and that the amino-terminal region plays
0090-4295/96/$15.00 PII SOO90-4295(96)00182-3 317
60 -
PTH(lnM) - +
PSA (nM) - - &l A $5 Al
PTHrP (1nM) - +
PSA(nM) - - &I
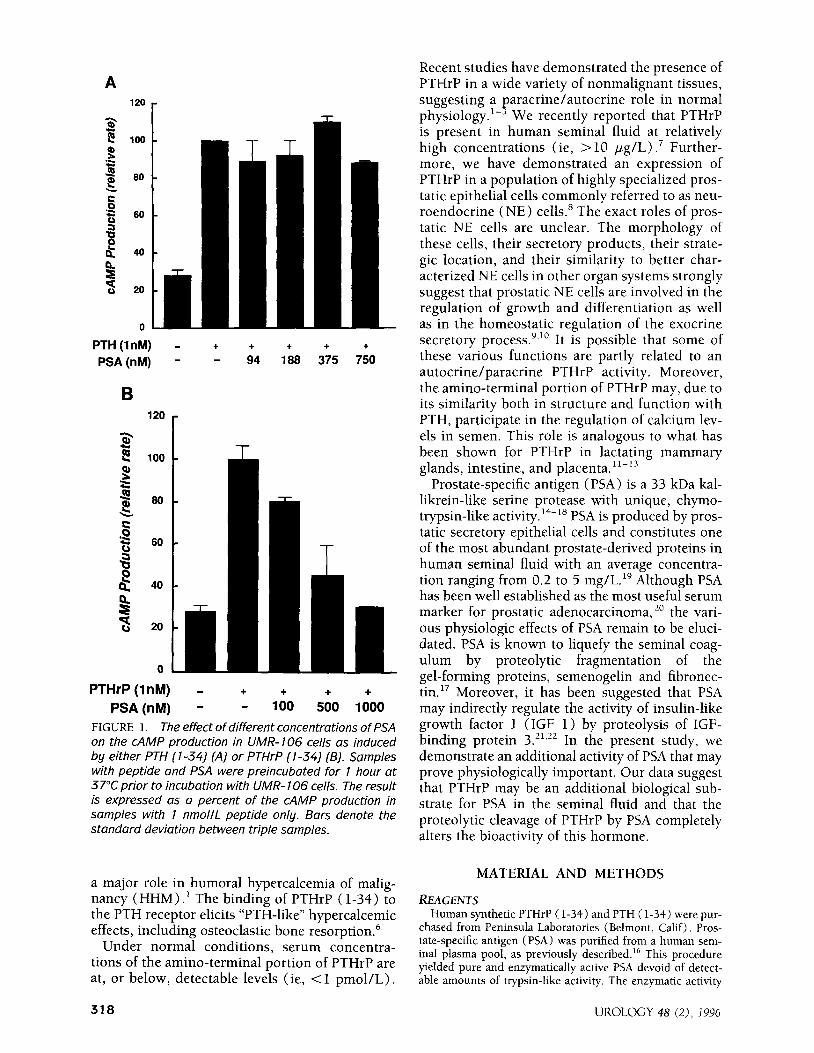
FIGURE 1. The effect of different concentrations of PSA on the CAMP production in UMR- 106 cells as induced by either PTH (l-34) (A) or PTHrP (l-34) (B). Samples with peptide and PSA were preincubated for 1 hour at 37°C prior to incubation with UMR- 106 cells. The result is expressed as a percent of the CAMP production in samples with 1 nmollL peptide only. Bars denote the standard deviation between triple samples.
a major role in humoral hypercalcemia of malig- nancy (HHM > .l The binding of PTHrP ( l-34) to the PTH receptor elicits “PTH-like” hypercalcemic effects, including osteoclastic bone resorption.6
Under normal conditions, serum concentra- tions of the amino-terminal portion of PTHrP are at, or below, detectable levels (ie, < 1 pmol/L).
Recent studies have demonstrated the presence of PTHrP in a wide variety of nonmalignant tissues, suggesting a paracrine/autocrine role in normal physiology.‘-3 We recently reported that PTHrP is present in human seminal fluid at relatively high concentrations (ie, >lO PgIL) .’ Further- more, we have demonstrated an expression of PTHrP in a population of highly specialized pros- tatic epithelial cells commonly referred to as neu- roendocrine (NE) cells.’ The exact roles of pros- tatic NE cells are unclear. The morphology of these cells, their secretory products, their strate- gic location, and their similarity to better char- acterized NE cells in other organ systems strongly suggest that prostatic NE cells are involved in the regulation of growth and differentiation as well as in the homeostatic regulation of the exocrine secretory process.9~10 It is possible that some of these various functions are partly related to an autocrine/paracrine PTHrP activity. Moreover, the amino-terminal portion of PTHrP may, due to its similarity both in structure and function with PTH, participate in the regulation of calcium lev- els in semen. This role is analogous to what has been shown for PTHrP in lactating mammary glands, intestine, and placenta.l’-l3
Prostate-specific antigen (PSA > is a 33 kDa kal- likrein-like serine protease with unique, chymo- trypsin-like activity.14-l8 PSA is produced by pros- tatic secretory epithelial cells and constitutes one of the most abundant prostate-derived proteins in human seminal fluid with an average concentra- tion ranging from 0.2 to 5 mg/L.” Although PSA has been well established as the most useful serum marker for prostatic adenocarcinoma, 2o the vari- ous physiologic effects of PSA remain to be eluci- dated. PSA is known to liquefy the seminal coag- ulum by proteolytic fragmentation of the gel-forming proteins, semenogelin and fibronec- tin.” Moreover, it has been suggested that PSA may indirectly regulate the activity of insulin-like growth factor 1 (IGF 1) by proteolysis of IGF- binding protein 3.21,22 In the present study, we demonstrate an additional activity of PSA that may prove physiologically important. Our data suggest that PTHrP may be an additional biological sub- strate for PSA in the seminal fluid and that the proteolytic cleavage of PTHrP by PSA completely alters the bioactivity of this hormone.
MATERIAL AND METHODS
REAGENTS Human synthetic PTHrP ( l-34) and PTH (l-34) were pur-
chased from Peninsula Laboratories (Belmont, Calif). Pros- tate-specific antigen (PSA) was purified from a human sem- inal plasma pool, as previously described.16 This procedure yielded pure and enzymatically active PSA devoid of detect- able amounts of trypsin-like activity. The enzymatic activity
318 UROLOGY 48 (2)) 1996

and specificity were determined by measuring the hydrolysis of synthetic substrates by PSA. Judging by PSA’s ability to form a complex with alphai-antichymotrypsin (ACT), about 65% was in the single-chain form and enzymatically active. The rest of the PSA was enzymatically inactive because of internal peptide bond cleavage. Unless described otherwise, all cell culture reagents and standard chemicals were obtained from Sigma (St. Louis, MO).
CELL CULTURE A rat osteosarcoma cell line, UMR-106, was obtained from
the American Type Tissue Collection (Rockville, Md) and cultured in Dulbecco’s modified Eagle’s medium (DMEM) with 10% heat-inactivated fetal bovine serum (FBS) The BEN cell line (kindly provided by Dr. Leonard Deftos, University of California, San Diego VA Medical Center, La Jolla, Calif) was incubated in RPM1 1640 medium with 10% FBS. This cell line was originally established from a human squamous-cell lung cancer associated with severe hypercalcemia23 and has been well characterized for its secretion of PTHrP.24 BEN con- ditioned medium was obtained by incubating cells for 48 hours in a serum-free medium consisting of a 1:l mixture of DMEM and Ham’s F-12 medium supplemented with 0.1% bo- vine serum albumin (BSA; fraction V), 5 PgImL insulin, 5 pg/mL transferrin, and 5 ng/mL selenious acid (ITS; Becton Dickinson Labware, Bedford, Mass)
CYCLK AMP ASSAY The biological activity of PTHrP (l-34) was determined by
measuring its effect on the production of CAMP in UMR-106 cells as has been described.25 In brief, 3 X lo5 cells were plated into each well of a 12-well plate (Falcon; Becton Dick- inson Labware, Lincoln Park, NJ ) and incubated to 90% con- fluency. Wells were rinsed twice with serum-free RPM1 1640 medium and preincubated for 10 minutes at 37°C with pre- warmed assay medium consisting of RPM1 1640 supple- mented with 0.2% BSA, 15 mmol/L Hepes, and 1 pmol/L isobutyl-methylxanthine. Samples diluted in the same media were then added and incubated for 10 minutes at 37°C. The supernatant was discarded, and the reaction was terminated by the addition of 0.5 mL ice-cold 65% ethanol. Cellular CAMP was extracted by repeating the freeze-thaw cycle three times. Cells and supematants were transferred to 5-mL poly- propylene tubes. Wells were then rinsed with an additional 0.5 mL ethanol, transferred to the same tubes, and centrifuged at 10,OOOg for 15 minutes at room temperature. The super- natants were lyophilized and resuspended in 0.5 mL distilled water. Finally, CAMP was measured using the BIOTRAK CAMP enzyme-immunoassay (EIA) system (Amersham, Ar- lington Heights, Ill).
SDS-POLYACRYLAMIDE GEL ELECTROPHORESIS The effect of PSA on PTHrP was studied by SDS-polyacryl-
amide gel electrophoresis (SDS-PAGE) essentially according to standard protocols. Samples were prepared by incubating PTHrP (l-34) (20 pg, 5 nmol) with PSA (0 to 1000 nmol) in phosphate-buffered saline (PBS), pH 7.2, at 37°C for 1 hour. The reaction was terminated by adding one volume of SDS sample buffer (2% SDS, 10% glycerol, and 5% beta-mer- captoethanol) After electrophoresis, the gel was stained with Coomassie blue according to standard procedures.
AMINO-TERMINAL SEQUENCE ANALYSIS PSA and PTHrP were incubated in PBS, pH 7.2, at 37°C for
4 hours. After the incubation, the sample was applied directly onto a C4 column (2.1 mm X 30 mm Aquapore Butyl 7 pm, Brownlee Labs, Santa Clara, Calif), equilibrated with 50 mmol/L phosphate buffer, pH 6.8. The material was eluted
with a linear acetonitrile gradient in the same buffer from 0% to 50% (vol/vol) for 60 minutes at a flow rate of 0.1 mllmin. Fractions were collected manually. The HPLC system con- sisted of a Beckman System Gold PSM 126 (Palo Alto, Calif) connected to a Waters 990 Photodiode Assay Detector (Mil- ford, Mass). The collected fractions were subjected to auto- mated amino-terminal sequence determination using a gas- phase sequencer (Applied Biosystems 470A, Foster City, Calif). Phenylthiohydantoin derivatives were identified in an on-line HPLC equipped with a 12OA analyzer.
MASS SPECTROMETRY Sample aliquots of 0.5 PL were mixed with an equal volume
of alpha-cyano-4-hydroxy cinnamic acid (Aldrich, Milwau- kee, Wise) (10 mg/mL) in 60% acetonitrile, 0.05% trifluo- roacetic acid. The samples were analyzed by Matrix Assisted Laser Desorption Ionisation Time-of-flight Mass Spectrometry (Lasermat; Finnigan MAT Ltd) External calibration was used.
RESULTS
EFFECT OF PTHrP (I-34) AND PTH (I-34) ON CAMP PRODUCTION rN UMR- 106 CELL LINE
As PTHrP (l-34) has been shown to share a common receptor with PTH on UMR-106 cells by affecting the production of CAMP, this system was utilized to determine the effect of enzymatically active PSA on the biological activity of PTHrP. In the absence of PSA both PTH ( l-34) and PTHrP ( l-34) stimulated the production of CAMP, and in line with earlier findings, as shown in Figures 1A and 1B.25
INH~B~TORYEFFECTOF PSAONPTH~P (I-34) ACTIVITY
Figure 1B illustrates the effect of PSA on the bio- logical activity of PTHrP (l-34). Enzymatically active, PSA-inhibited PTHrP (l-34) (1 nmol/L) induced CAMP production in a dose-dependent manner. Preincubation of PSA with aprotinin (20 PglmL) , an inhibitor of the trypsin-like protease pancreatic glandular kallikrein but not an inhibi- tor of the chymotrypsin-like PSA activity,“j did not alter the PSA-dependent inhibition of the PTHrP (l-34) induction of CAMP production in UMR- 106 cells. However, the inhibitory effect of PSA was abolished through inactivation of the enzyme activity by preheating PSA at 100°C for 5 minutes prior to incubation with PTHrP (data not shown). In contrast, enzymatically active PSA did not have any effect on the stimulation of CAMP produc- tion in UMR-106 cells induced by PTH (l-34) (Fig. 1A).
The inhibition of PTHrP (l-34) activity by PSA was time dependent (Fig. 2). A significant effect was seen at 10 minutes of incubation. This effect increased when the incubation was extended to 1 hour, after which a 70% decrease in PTHrP (l- 34) -dependent stimulation of CAMP production was recorded.
UROLOGY 48 (Z), 1996 319
01 I I I I I
0 30 60 90 120 150
Incubation Time (min) FIGURE 2. Time-course for PSA-induced inhibition of the effect of PTHrP on CAMP production in UMR- 106 cells. Samples containing 1 nmollL PTHrP (l-34) and 1000 nmollL PSA were preincubated for different time
periods (ranging from 0 to 120 minutes) at 37°C prior to incubation with UMR- 106 cells. The result is ex- pressed as percent decrease in CAMP production by UMR-106 cells compared with a sample containing 1 nmollL PTHrP (l-34) only.
120 r
A B C D E F
Treatments FIGURE 3. The effect of PSA on the induction of the CAMP production in UMR- 106 cells by a BEN cell culture medium (CCM). Samples were preincubated for 1 hour at 37°C prior to incubation with UMR- 106 cells. (A) Con- trol (serum free medium); (B) serum free medium, 0.5 nmollL PTHrP (l-34); (C) BEN CCM; (0) BEN CCM, 125 nmollL PSA; (E) BEN CCM, 250 nmollL PSA; (F) BEN CCM, 1000 nmollL PSA. The result is expressed as a percent of CAMP production in samples with 1 nmollL peptide only (B). Bars denote the standard deviation be- tween triple samples.
320 UROLOGY 48 (21, 1996
-PTHrP (l-34)
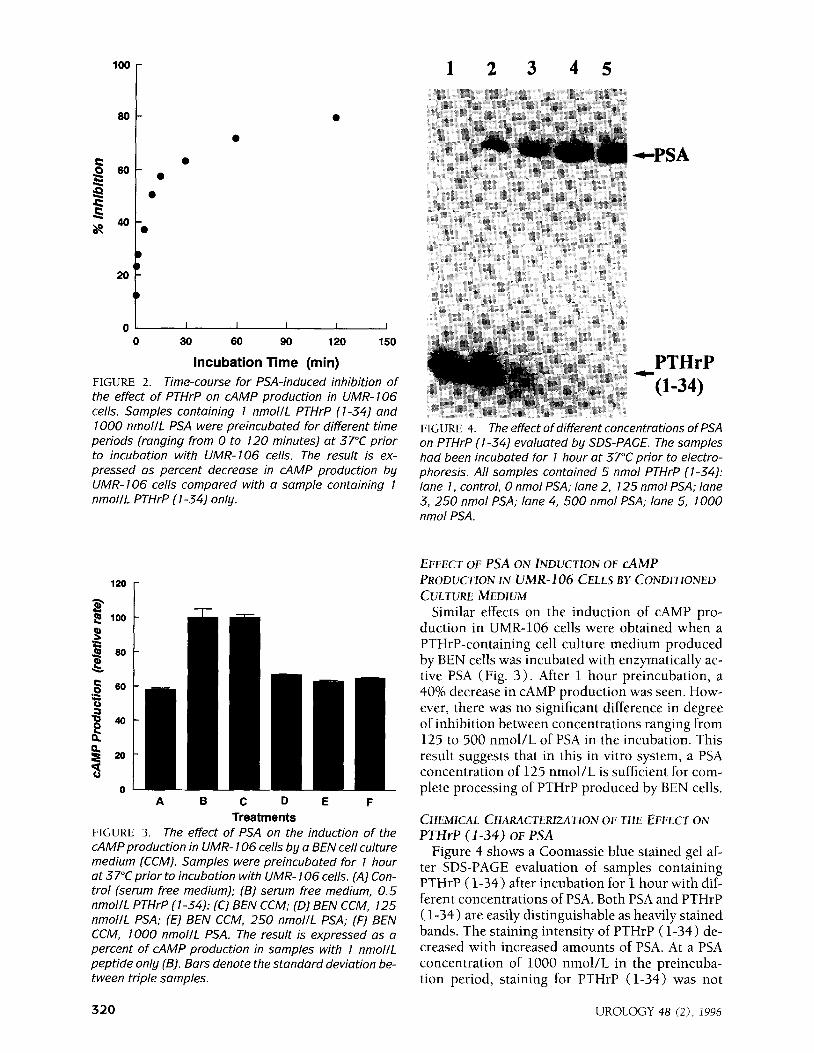
FIGURE 4. The effect of different concentrations of PSA on PTHrP (l-34) evaluated by SDS-PACE. The samples had been incubated for 1 hour at 37°C prior to electro- phoresis. All samples contained 5 nmol PTHrP (l-34): lane 1, control, 0 nmol PSA; lane 2, 125 nmol PSA; lane 3, 250 nmol PSA; lane 4, 500 nmol PSA; lane 5, 1000 nmol PSA.
EFFECT OF PSA ON INDUCTION OF CAMP PRODUCTIONIN UMR-106 CELLSBY CONDITIONED CULTURE MEDIUM
Similar effects on the induction of CAMP pro- duction in UMR-106 cells were obtained when a PTHrP-containing cell culture medium produced by BEN cells was incubated with enzymatically ac- tive PSA (Fig. 3). After 1 hour preincubation, a 40% decrease in CAMP production was seen. How- ever, there was no significant difference in degree of inhibition between concentrations ranging from 125 to 500 nmol/L of PSA in the incubation. This result suggests that in this in vitro system, a PSA concentration of 125 nmol/L is sufficient for com- plete processing of PTHrP produced by BEN cells.
CHEMICAL CHARACTERIZATIONOFTHE EFFECTON PTHrP (l-34) OFPSA
Figure 4 shows a Coomassie blue stained gel af- ter SDS-PAGE evaluation of samples containing PTHrP ( l-34) after incubation for 1 hour with dif- ferent concentrations of PSA. Both PSA and PTHrP ( 1-34) are easily distinguishable as heavily stained bands. The staining intensity of PTHrP (l-34) de- creased with increased amounts of PSA. At a PSA concentration of 1000 nmol/L in the preincuba- tion period, staining for PTHrP (l-34) was not
20 40
minutes 60
B ’ -
.8
8 .6
s 2 8 9 .4
1
/
4
k
0 20 40 minutes
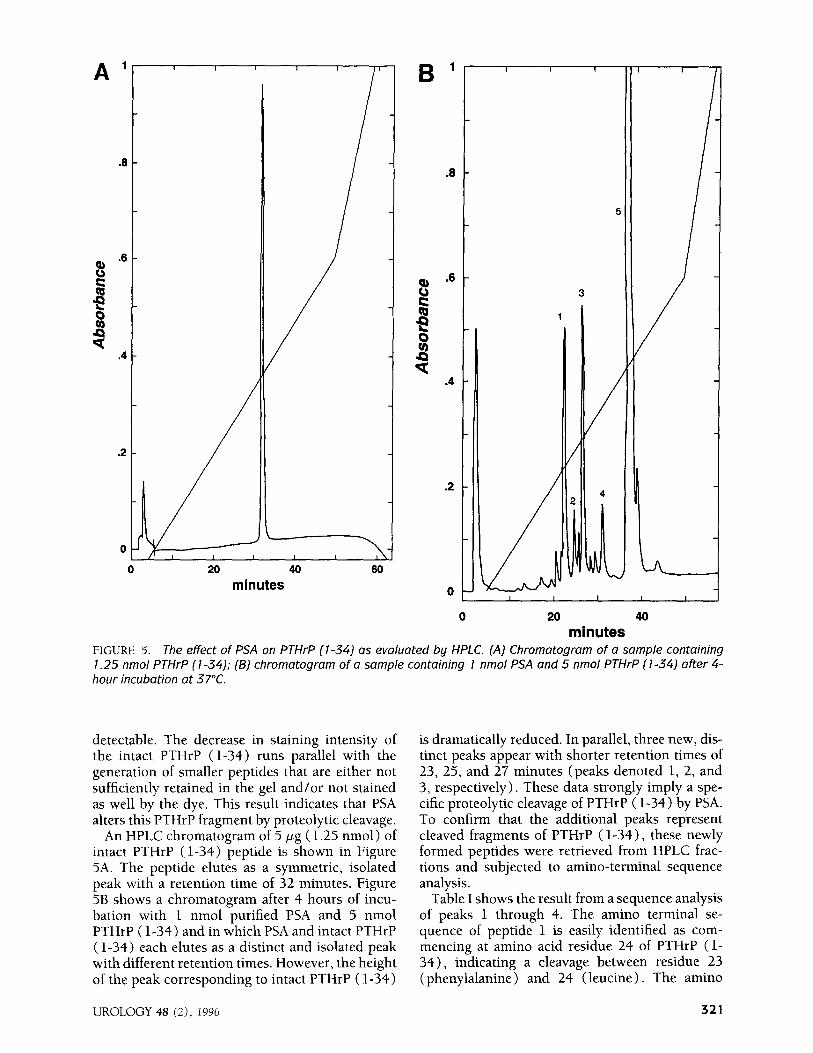
FIGURE 5. The effect of PSA on PTHrP (l-34) as evaluated by HPLC. (A) Chromatogram of a sample containing 1.25 nmol PTHrP (l-34); (B) chromatogram of a sample containing 1 nmol PSA and 5 nmol PTHrP (l-34) after 4-
hour incubation at 37°C.
detectable. The decrease in staining intensity of the intact PTHrP (l-34) runs parallel with the generation of smaller peptides that are either not sufficiently retained in the gel and/or not stained as well by the dye. This result indicates that PSA alters this PTHrP fragment by proteolytic cleavage.
An HPLC chromatogram of 5 pg ( 1.25 nmol) of intact PTHrP (l-34) peptide is shown in Figure 5A. The peptide elutes as a symmetric, isolated peak with a retention time of 32 minutes. Figure 5B shows a chromatogram after 4 hours of incu- bation with 1 nmol purified PSA and 5 nmol PTHrP ( l-34) and in which PSA and intact PTHrP ( l-34) each elutes as a distinct and isolated peak with different retention times. However, the height of the peak corresponding to intact PTHrP ( l-34)
UROLOGY 48 (21, 1996 321
is dramatically reduced. In parallel, three new, dis- tinct peaks appear with shorter retention times of 23, 25, and 27 minutes (peaks denoted 1, 2, and 3, respectively) . These data strongly imply a spe- cific proteolytic cleavage of PTHrP ( l-34) by PSA. To confirm that the additional peaks represent cleaved fragments of PTHrP (l-34)) these newly formed peptides were retrieved from HPLC frac- tions and subjected to amino-terminal sequence analysis.
Table I shows the result from a sequence analysis of peaks 1 through 4. The amino terminal se- quence of peptide 1 is easily identified as com- mencing at amino acid residue 24 of PTHrP (l- 34)) indicating a cleavage between residue 23 (phenylalanine) and 24 (leucine) . The amino
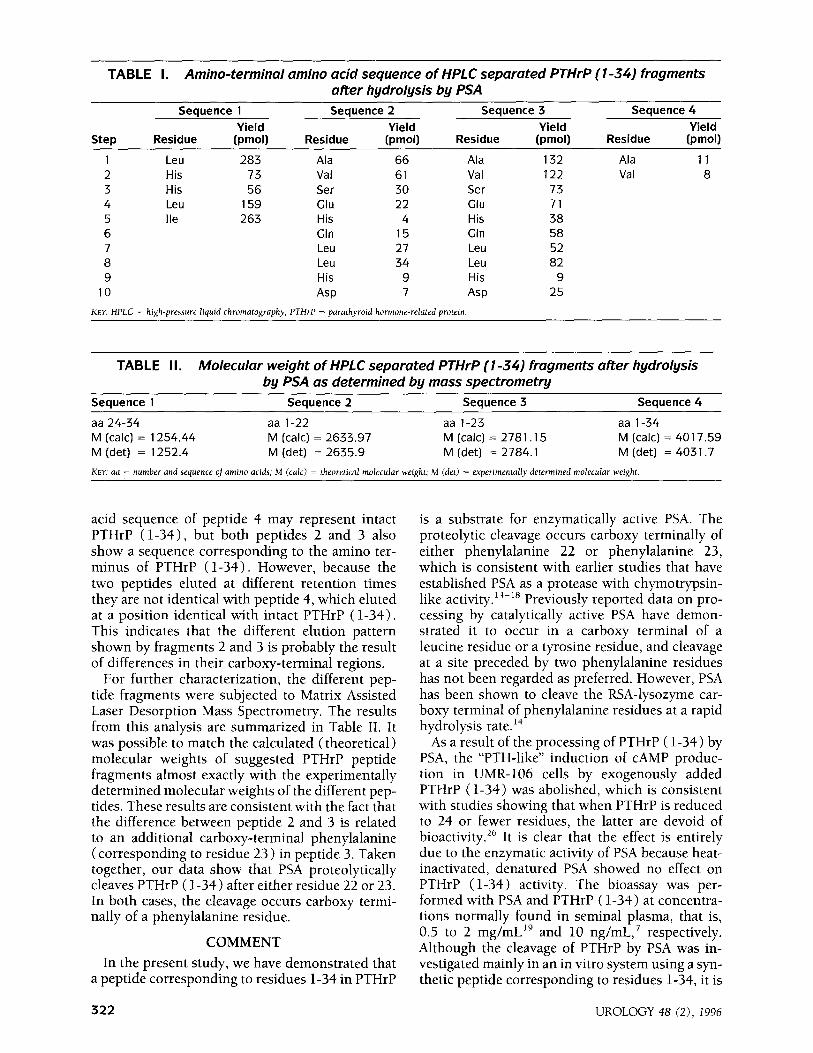
TABLE I. Amino-terminal amino acid sequence of HPLC separated PTHrP (l-34) fragments after hydrolysis by PSA
Sequence 1 Sequence 2 Sequence 3 Sequence 4
Yield Yield Yield Yield Step Residue (pm011 Residue (pmoll Residue (pm011 Residue (pmoll
1 Leu 283 Ala 66 Ala 132 Ala 11 2 His 73 Val 61 Val 122 Val 8 3 His 56 Ser 30 Ser 73 4 Leu 159 Gill 22 au 71 5 Ile 263 His 4 His 38 6 Glfl 15 Cln 58 7 Leu 27 Leu 52 8 Leu 34 Leu 82 9 His 9 His 9
10 Asp 7 Asp 25
KEY: HPLC = high-pressure liquid chromatography; PiWrP = parathyroid hormone-related protein.
TABLE II. Molecular weight of HPLC separated PTHrP (l-34) fragments after hydrolysis by PSA as determined by mass spectrometry
Sequence 1 Sequence 2 Sequence 3 Sequence 4
aa 24-34 aa l-22 aa l-23 aa l-34 M (talc) = 1254.44 M (talc) = 2633.97 M (talc) = 278 1 .15 M (talc) = 4017.59 M (det) = 1252.4 M (det) = 2635.9 M (det) = 2784.1 M (det) = 403 1.7
KEY: aa = number and sequence of amino acids; M (talc) = theoretical molecular weight; M (det) = aperimentally determined molecular weight.
acid sequence of peptide 4 may represent intact PTHrP (l-34), but both peptides 2 and 3 also show a sequence corresponding to the amino ter- minus of PTHrP ( l-34). However, because the two peptides eluted at different retention times they are not identical with peptide 4, which eluted at a position identical with intact PTHrP (l-34). This indicates that the different elution pattern shown by fragments 2 and 3 is probably the result of differences in their carboxy-terminal regions,
For further characterization, the different pep- tide fragments were subjected to Matrix Assisted Laser Desorption Mass Spectrometry. The results from this analysis are summarized in Table II. It was possible to match the calculated (theoretical) molecular weights of suggested PTHrP peptide fragments almost exactly with the experimentally determined molecular weights of the different pep- tides. These results are consistent with the fact that the difference between peptide 2 and 3 is related to an additional carboxy-terminal phenylalanine (corresponding to residue 23 > in peptide 3. Taken together, our data show that PSA proteolytically cleaves PTHrP (l-34) after either residue 22 or 23, In both cases, the cleavage occurs carboxy termi- nally of a phenylalanine residue.
COMMENT
In the present study, we have demonstrated that a peptide corresponding to residues l-34 in PTHrP
322
is a substrate for enzymatically active PSA. The proteolytic cleavage occurs carboxy terminally of either phenylalanine 22 or phenylalanine 23, which is consistent with earlier studies that have established PSA as a protease with chymotrypsin- like activity. i4-18 Previously reported data on pro- cessing by catalytically active PSA have demon- strated it to occur in a carboxy terminal of a leucine residue or a tyrosine residue, and cleavage at a site preceded by two phenylalanine residues has not been regarded as preferred. However, PSA has been shown to cleave the RSA-lysozyme car- boxy terminal of phenylalanine residues at a rapid hydrolysis rate.14
As a result of the processing of PTHrP ( l-34) by PSA, the “PTH-like” induction of CAMP produc- tion in UMR-106 cells by exogenously added PTHrP (l-34) was abolished, which is consistent with studies showing that when PTHrP is reduced to 24 or fewer residues, the latter are devoid of bioactivity. 26 It is clear that the effect is entirely due to the enzymatic activity of PSA because heat- inactivated, denatured PSA showed no effect on PTHrP (l-34) activity. The bioassay was per- formed with PSA and PTHrP ( l-34) at concentra- tions normally found in seminal plasma, that is, 0.5 to 2 mg/mLig and 10 ng/mL,7 respectively. Although the cleavage of PTHrP by PSA was in- vestigated mainly in an in vitro system using a syn- thetic peptide corresponding to residues l-34, it is
UROLOGY 48 (21, 1996

possible that the results presented represent a physiologic process similar to that which may oc- cur in the prostate itself. This view is further sup- ported by the finding that the production of CAMP by UMR-106 cells, induced by a culture-condi- tioned medium of BEN cells, was decreased by prior incubation with enzymatically active PSA.
The source used to isolate PSA was seminal plasma collected after ejaculation. This material does not represent the fluid present in the prostatic ducts and acini. Instead, it is a mix of secretions originating from the different accessory sex organs and has probably been subjected to several phys- iologic changes. Thus, the form of PSA isolated from ejaculated seminal plasma may be very dif- ferent from the PSA originally present in the pros- tatic fluid. PSA is most likely secreted in the form of an inactive proenzyme, zymogen, which is ac- tivated through a specific proteolytic cleavage of the amino terminus2’ When and where this crit- ical activation takes place has yet to be elucidated. Not all PSA found in seminal plasma is free. Some PSA is found in a complex at a 1:l molar ratio with protein C inhibitor (PCI) .28 Seminal PC1 is believed to be produced mainly by the seminal ves- icles. In the blood, a major fraction of PSA circu- lates as a complex, either with alphai-antichymo- trypsin (ACT > or alpha2-macroglobulin.16 In analogy, it is plausible that a complex incorporat- ing ACT and PSA may be formed in the prostate ducts/seminal fluid, for it has been shown that ACT is expressed by cells of the prostate secretory epithelium. 29 It is imp0 r ta nt to determine the ex- tent to which ACT is involved in the regulation of the proteolytic activity of PSA through complex formation at the local prostatic level.
Our data suggest that PTHrP is a physiologic substrate for PSA, and we believe that once PSA has been activated and translated into a functional protease, it may be involved in the regulation of PTHrP activity, specifically in the prostate and in seminal plasma. If the activation of PSA is an early postsecretional event, resulting in the presence of an active protease in the prostatic ducts and acini, it may have important consequences for the par- acrine/autocrine activities of PTHrP.
The regulation of the bioactivity of PTHrP is very complex and takes place at several levels. Ste- roid hormones and polypeptide growth factors as well as other factors have been shown to be in- volved at the transcriptional leve1.30-32 These fac- tors may also affect translation and/or secretion of the different isoforms of PTHrP. Moreover, several studies have conceptualized PTHrP as a prohor- mone or precursor that contains several peptide regions, each of which has a distinct bioactivity and each of which is generated post-translationally through intracellular proteolytic processing.4V5
This processing of the PTHrP prohormone is prob- ably highly regulated in a manner specific to cell and/or tissue. Several fragments representing dif- ferent portions of PTHrP have been detected in blood samples from patients suffering from HHM, including one consisting of the 36 amino acid res- idues of the amino terminus.3-5
Further support for the prohormone concept can be found in functional studies. Although many studies have focused attention on the amino-ter- minal domain of PTHrP that results from its struc- tural and functional similarity to PTH, some unique “PTH-dissimilar” activities have been as- cribed to other domains. For instance, PTHrP (residues 67-86 > appears to be involved in the ac- tive transport of calcium in the placental’ and in- duces the transcription of mRNA for epidermal growth factor receptor,33 suggesting a role in cal- cium regulation as well as in cell growth and dif- ferentiation. In addition, a carboxy-terminal frag- ment consisting of residues 107-111 has been shown to be a potential inhibitor of osteoclasts and probably suppresses osteoclastic bone resorp- tion.6,34 This is contrary to the PTH-like activity of the amino terminus. In the present study, our data suggest an additional level of regulation of the bio- activity of PTHrP involving a postsecretional pro- cessing of PTHrP peptide fragments. This form of regulation may be critical either to elicit or to sup- press certain bioactivities of PTHrP and may be mediated locally in a manner specific to tissue or site.
In an earlier work, we have immunohistochem- ically characterized the widespread expression of PTHrP in prostatic adenocarcinomas using a monoclonal antibody raised against the carboxy- terminal fragment 109-141.35 In addition, human prostate cancer cell lines secrete immunodetecta- ble levels of PTHrP in culture-conditioned me- dium.36 It is known that hypercalcemia associated with prostate cancer occurs in less than 2% of pa- tients, as compared with its common occurrence in other PTHrP-producing tumors.37 This discrep- ancy does not appear to reflect a quantitative dif- ference in PTHrP levels, because the incidence of hypercalcemia seems low even in Stage D2 patients with multiple metastatic lesions. Instead, the PTH- like activity of PTHrP may be neutralized by fac- tors concomitantly produced by prostate cancer cells. One such factor is calcitonin. Calcitonin has been shown to be secreted by prostate NE cells and may functionally counteract the PTH-like activi- ties of PTHrP.38 An alternative explanation relates to the activity of PSA. The potential humoral hy- percalcemic action of PTHrP, secreted by prostate cancer cells, may be abated by the local enzymatic activity of PSA because they are commonly co-se- creted by most adenocarcinoma cells in the pros-
UROLOGY 48 (21, 1996 323

tate.35’39 This mechanism may perhaps also be valid for metastatic lesions.
In this study, the cleavage of PTHrP by PSA was investigated using the amino-terminal fragment of PTHrP. PTHrP contains several putative sites for cleavage by PSA located in the middle and car- boxy1 part. The previously mentioned PTHrP frag- ment consisting of amino acid residues 107-111, which has been shown to be a potential inhibitor of osteoclastic bone resorption, does not have any obvious putative cleavage site for PSA. Therefore, it is likely that the activity of PTHrP (107-111) is not affected by PSA. Instead, its activity may ac- tually be enhanced by a presence of proteolytic PSA activity inasmuch as this PTHrP fragment has the opposite function of the amino terminus. This may be particularly important in the establishment and formation of bone metastases by prostate can- cer cells. In contrast to the behavior of other ma- lignant tumors, most metastatic bone lesions of prostate cancer are accompanied by “osteoblastic” changes. This phenomenon may be explained, at least in part, by the effect of PSA on PTHrP se- creted by prostate cancer cells in bone.
In conclusion, our study demonstrates that PTHrP is a physiologic substrate for PSA. PSA may play an important role in the post-translational regulation of prostatic PTHrP activities, which in- clude regulation of prostate growth and differen- tiation as well as regulation of the calcium concentration in semen. We are currently inves- tigating the interaction between PSA and PTHrP in semen and its effect on the PTH-like activity of PTHrP. Further studies will be needed to elucidate the effects of PSA on other region-specific activi- ties of PTHrP.
ACKNOWLEDGMENT. To Susan Schoen for her expert tech-
nical assistance.
REFERENCES
1. Martin TJ: Properties of parathyroid hormone-related protein and its role in malignant hypercalcemia. Q J Med 76: 771-786, 1990.
2. Martin TJ, Moseley JM, and Gillespie MT: Parathyroid hormone-related protein: biochemistry and molecular biol- ogy. Crit Rev Biochem Mol Biol 26: 377-395, 1991.
3. Burtis WJ: Parathyroid hormone-related protein: struc- ture, function, and measurment. Clin Chem 38: 2171-2183,
1992. 4. Orloff JJ, Reddy D, dePapp AE, Yang KH, Soifer NE,
and Stewart AF: Parathyroid hormone-related protein as a prohormone: posttranslational processing and receptor inter- actions. Endocr Rev 15: 40-60, 1994.
5. Yang KH, dePapp AE, Soifer NE, Dreyer BE, Wu TL, Porter SE, Bellantoni M, Burtis WJ, Insogna KL, Broadus AE,
et al: Parathyroid hormone-related protein: evidence for iso- form- and tissue-specific posttranslational processing. Bio- chemistry 33: 7460-7469,1994.
6. Evely RS, Bonomo A, Schneider H-G, Moseley JM, Gal- lagher J, and Martin TJ: Structural requirements for the action
of parathyroid hormone-related protein ( PTHrP) on bone re- sorption by isolated osteoclasts. J Bone Miner Res 6: 85-93, 1991.
7. Iwamura M, Abrahamsson P-A, Schoen SR, Cockett ATK, and Deftos LJ: Immunoreactive parathyroid hormone- related protein is present in human seminal plasma and is of
prostate origin. J Androl 15: 410-414, 1994. 8. Iwamura M, Wu G, Abrahamsson P-A, di Sant’Agnese
PA, Cockett ATK, and Deftos LJ: Parathyroid hormone-related protein is expressed by prostatic neuroendocrine cells. Urol-
ogy 43: 667-674, 1994. 9. Abrahamsson P-A, and di Sant’Agnese PA: Neuroen-
docrine cells in the human prostate gland. J Androl 5: 307-
309, 1993. 10. Bonkhoff H, Wernert N, Dhom G, and Remberger K:
Relation of endocrine-paracrine cells to cell proliferation in normal, hyperplastic, and neoplastic human prostate. Prostate 19: 91-98, 1991.
11. Barlet JP, Champredon C, Coxam V, Davicco MJ, and Tress01 JC: Parathyroid hormone-related peptide might stim-
ulate calcium secretion into the milk of goats. J Endocrinol 132: 353-359,1992.
12. Care AD, Caple IW, and Martin TJ: Evidence for a novel parathyroid hormone-related protein in fetal lamb par-
athyroid glands and sheep placenta: comparisons with a sim- ilar protein implicated in humoral hypercalcemia of malig- nancy. J Endocrinol 117: 261-271, 1988.
13. Zhou LX, Nemere I, and Norman AW: A parathyroid hormone-related peptide induces transcaltachia (the rapid, hormonal stimulation of intestinal Ca’+ transport), Biochem
Biophys Res Commun 186: 69-73, 1992. 14. Akiyama K, Nakamura T, Iwanaga S, and Hara M: The
chymotrypsin-like activity of human prostate-specific antigen, y-seminoprotein. FEBS Lett 225: 168-172, 1987.
15. Watt KWK, Lee P-J, M’Timkulu TM, Chan W-P, and Loor R: Human prostate-specific antigen: structural and func- tional similarity with serine proteases. Proc Nat1 Acad Sci
U S A 83: 3166-3170, 1986. 16. Christensson A, Laurel1 C-B, and Lilja H: Enzymatic
activity of prostate-specific antigen and its reactions with ex-
tracellular serine proteinase inhibitors. Eur J Biochem 194: 755-763,199O.
17. Lilja H: A kallikrein-like serine protease in prostatic fluid cleaves the predominant seminal vesicle protein. J Clin Invest 76: 1899-1903, 1985.
18. Ban Y, Wang MC, Watt KW, Loor L, and Chu TM: The proteolytic activity of human prostate-specific antigen. Biochem Biophys Res Commun 123: 482-488, 1984.
19. Bilhartz DL, Tindall DJ, and Oesterling JE: Prostate- specific antigen and prostatic acid phosphatase: biomolecular and physiological characteristics. Urology 38: 95-102, 1991.
20. Oesterling JE: Prostate-specific antigen: a critical as- sessment of the most useful tumor marker for adenocarci- noma of the prostate. J Urol 145: 907-923, 1991.
21. Cohen P, Graves HCB, Peehl DM, Kamarei M, Giudice LC, and Rosenfeld RG: Prostate-specific antigen (PSA) is an insulin-like growth factor binding protein-3 protease found
in seminal plasma. J Clin Endocrinol Metab 75: 1046-1053, 1992.
22. Cohen P, Peehl DM, Graves HC, and Rosenfeld RG:
Biological effects of prostate specific antigen as an insulin-like growth factor binding protein-3 protease. J Endocrinol 142: 407-415, 1994.
23. Ellison M, Woodhouse D, Hillyard CJ, Dowsett M, Coombes RC, Gilby ED, Greenberg PB, and Neville AM: Im- munoreactive calcitonin production by human lung carci- noma cells in culture. Br J Cancer 32: 373-379, 1975.
24. Moseley JM, Kubota M, Diefenbach-Jagger H, Wetten- hall REH, Kemp BE, Suva LJ, Rodda CP, Ebeling PR, Hudson
324 UROLOGY 48 (2), 1996

PJ, Zajac JD, et al: Parathyroid hormone-related protein pu- rified from a human lung cancer cell line. Proc Nat1 Acad Sci U S A 84: 5048-5052, 1987.
25. Schermer DT, Bradley MS, Bambino TH, Nissenson RA, and Strewler GJ: Functional properties of a synthetic chicken parathyroid hormone-related protein l-36 fragment. J Bone Miner Res 9: 1041- 1046,1994.
26. Kemp BE, Moseley JM, Rodda CP, Ebeling PR, Wetten- hall REH, Stapleton D, Diefenbach-Jagger H, Ure F, Miche- langeli VP, Simmons HA, et al: Parathyroid hormone-related protein of malignancy: active synthetic fragments. Science 238:1568-1570,1987.
27. Lundwall A, and Lilja H: Molecular cloning of human prostate specific antigen cDNA. FEBS Lett 214: 317-322, 1987.
28. Laurel1 M, Christensson A, Abrahamsson P-A, Stenflo J, and Lilja H: Protein C inhibitor in human body fluids. Sem- inal plasma is rich in inhibitor antigen deriving from cells throughout the male reproductive system. J Clin lnvest 89: 1094-1101,1992.
29. Bjartell A, Bjork T, Matikainen M-T, Abrahamsson P-A, di Sant’Agnese A, and Lilja H: Production of alfa-l-an- tichymotrypsin by PSA-containing cells of human prostate ep- ithelium. Urology 42: 502-510, 1993.
30. Casey ML, Mibe M, and MacDonald PC: Regulation of parathyroid hormone-related protein gene expression in hu- man endometrial stromal cells in culture. J Clin Endocrinol Metab 77: 188-194, 1993.
31. Merryman JI, DeWille JW, Werkmeister JR, Capen CC, and Rosol TJ: Effects of transforming growth factor-p on par- athyroid hormone-related protein production and ribonucleic acid expression by a squamous carcinoma cell line in vitro. Endocrinology 134: 2424-2430, 1994.
32. Li X, and Drucker DJ: Parathyroid hormone-related peptide is a downstream target for ras and SYC activation, J Biol Chem 269: 6263-6266, 1994.
33. Alsat E, Haziza J, Scippo M-L, Frankenne F, and Evain- Brion D: Increase in epidermal growth factor receptor and its mRNA levels by parathyroid hormone (l-34) and parathyroid hormone-related protein ( l-34) during differentiation of human trophoblast cells in culture. J Cell Biochem 53: 32-42, 1993.
34. Fenton AJ, Martin TJ, and Nicholson GC: Carboxy- terminal parathyroid hormone-related protein inhibits bone resorption by isolated chicken osteoclasts. J Bone Miner Res 9: 515-519,1994.
35. lwamura M, di Sant’Agnese PA, Wu G, Benning CM, Cockett ATK, Deftos LJ, and Abrahamsson P-A: Immuno- histochemical localization of parathyroid hormone-related protein in human prostate cancer. Cancer Res 53: 1724- 1726, 1993.
36. Iwamura M, Abrahamsson P-A, Foss KA, Wu G, Cock- ett ATK, and Deftos LJ: Parathyroid hormone-related protein: a potential autocrine growth regulator in human prostate can- cer cell lines. Urology 43: 675-679, 1994.
37. Smith DC, Tucker JA, and Trump DL: Hypercalcemia and neuroendocrine carcinoma of the prostate: a report of three cases and a review of the literature. J Clin Oncol 10: 499-505,1992.
38. di Sant’Agnese PA, de Mesy Jensen KL, and Ackroyd RK: Calcitonin, katacalcin, and calcitonin gene-related pep- tide in the human prostate. Arch Path01 Lab Med 113: 790- 796, 1989.
39. Bjork T, Bjartell A, Abrahamsson P-A, Hulkko S, di Sa- nt’Agnese A, and Lilja H: Alfar-antichymotrypsin production in PSA-producing cells is common in prostate cancer but rare in benign prostatic hyperplasia. Urology 43: 427-434, 1994.
UROLOGY 48 (2)) 1996 325




















