
GREEN'S FUNCTIONS OF FILAMENT SOURCES EMBEDDED IN STRATIFIED DIELECTRIC MEDIA

Journal of Neururhemistry Raven Press, New York 0 1986 International Society for Neurochemistry
Homology and Diversity Between Intermediate Filament Proteins of Neuronal and Nonneuronal Origin in Goldfish
Optic Nerve
Wolfgang Quitschke and Nisson Schechter
Department of Psychiatry and Behavioral Science, Health Sciences Center, State University of New York at Stony Brook, Stony Brook, New York U.S.A.
Abstract: The predominant intermediate filament pro- teins of the goldfish optic nerve have molecular weights of 58K. They can be separated into a series of four major isoelectric variants of neuronal (ON, and ON,) and non- neuronal (ON, and ON4) origin. The extent of homology between the goldfish 58K intermediate filament proteins themselves and to rat optic nerve vimentin and dial fi- brillary acidic protein (GFAP) was investigated. Unla- beled and [32P]orthophosphate-labeled proteins were sub- jected to partial hydrolysis by V8 protease, chymo- trypsin, and CNBr. The results show that the goldfish intermediate filament proteins share with vimentin and GFAP a 40K chymotrypsin-resistant core fragment. Phosphorylated moieties appear to be located outside the core region since they are preferentially cleaved off by chymotrypsin and not found associated with the 40K
core. In addition, the goldfish ON proteins contain the antigenic site within the core that is common to most intermediate filaments. V8 or CNBr digestion indicates that many fragments that are common to ON, and ON, are clearly distinct from fragments that are common to ON, and ON,. In addition, structural variability is ob- served between the goldfish intermediate filament pro- teins and vimentin and GFAF! The results are discussed in terms of intermediate filament structure and their pos- sible role in nerve growth. Key Words: Intermediate fil- aments-Optic nerve-Fragment patterns-Goldfish- Rat. Quitschke W. and Schechter N. Homology and di- versity between intermediate filament proteins of neu- ronal and nonneuronal origin in goldfish optic nerve. J . Neurochem. 46, 545-555 (1986).
Intermediate filaments are more diverse in pro- tein composition than the two other major cytoskel- eta1 structures, the microfilaments and the micro- tubules (Lazarides, 1982). According to their cel- lular localization and tissue specificity, at least five subclasses of proteins belonging to the intermediate filament complex (Franke et al., 1982; Lazarides 1982) have been delineated. Mammalian neurofila- ments are composed of three proteins having mo- lecular weights of 200,000 daltons (200K), 150K, and 70K (Hoffman and Lasek, 1975; Weber et al., 1983). In the CNS astrocytes contain glial fibrillary acidic protein (GFAP-51K) which is organized into the glial filaments. In addition, vimentin (58K), an intermediate filament protein which, in general, has been associated with mesenchymally derived cells,
can coexist with GFAP in astrocytes (Shaw et al., 1981; Dahl et al., 1982). It has also been observed that during embryogenesis vimentin is transiently expressed in neuronal precursor cells (Tapscott et al., 1981) and in developing central (Bignami et al., 1982) and peripheral (Cochard and Paulin, 1984) neurons.
The goldfish visual pathway can be viewed as being in a quasi-embryonic state. This pathway ex- hibits a capacity for continued addition of new neu- rons throughout its life (Meyer 1978; Easter and Stuermer, 1984) and a capacity for nerve regenera- tion after optic nerve crush (Sperry, 1%3). Molec- ular events associated with these phenomena can be studied with this system (Heacock and Agranoff, 1976; Schechter et al., 1979; Giulian et al., 1981;
Received April 5, 1985; accepted August 23, 1985. Address correspondence and reprint requests to Dr. N.
Schechter at Department of Psychiatry and Behavioral Science, Health Sciences Center, T-10, State University of New York, Stony Brook, NY 11794, U.S.A.
Abbreviations used: AIF-antibody, monoclonal antibody to in- termediate fdament proteins; BSA, bovine serum albumin; Ches, 2-(N-~yclohexylamino)ethanesulfonic acid; DIT, dithiothreitol; GFAP, dial fibrillary acidic protein; ON, goldfish optic nerve intermediate filament proteins (58K); SDS, sodium dodecyl sul- fate.
545

546 W . QUITSCHKE AND N . SCHECHTER
Benowitz and Lewis, 1983; Perry et al., 1985). in- cluding the synthesis of intermediate filament pro- teins (Quitschke and Schechter, 19836). The com- position of the intermediate filaments of the gold- fish optic nerve is different from the conventional protein composition of intermediate filaments found within the more stable mammalian nerves (Quitschke and Schechter, 1984). This observation can be extended further. In contrast to mammals, lower vertebrates in general express a diversity of intermediate filament proteins within homologous nerves of different species and within different nerves of the same species (Quitschke et al., 1985).
The predominant intermediate filament proteins of the goldfish optic nerve have molecular weights of 58K. They can be separated on two-dimensional gels into a series of isoelectric variants, previously designated as ON,, ON,, ON,, and ON, (Quitschke and Schechter, 1983~) . These proteins share with vben t in the same molecular weight; however, their isoelectric points are more basic. These proteins do not react with specific monoclonal antibodies to vi- mentin; however, they do react with general anti- bodies to intermediate filament proteins (Quitschke et al., 1985). Proteins ON, and ON, are neurofila- ment proteins, whereas ON, and ON, are nonneu- ronal intermediate filament proteins. The relative concentration of these proteins is much higher in the optic nerve than in other brain regions of the goldfish. Electrophoretic analysis shows that sub- components of ON, and ON, are specific to the goldfish retinotectal pathway. Furthermore, these proteins (ON,-ON,) are differentially phosphory- lated both in vivo and in vitro. On the basis of the above observations (Quitschke a n d Schechter, 1983a,b, 1984), the extent of homology between the goldfish 58K intermediate filament proteins them- selves and to vimentin and GFAP becomes an im- portant issue. Unlabeled and [32Plorthophosphate- labeled goldfish and rat optic nerve filament pro- teins were subjected to partial hydrolysis by V8 protease, chymotrypsin, and cyanogen bromide. The rat optic nerve was chosen because it contains substantial amounts of both GFAP and vimentin. The resulting fragment patterns from the studies presented here are compared and discussed in terms of intermediate filament structure and their possible role in nerve growth.
MATERIALS AND METHODS
Animals The optic nerves of goldfish (Carassius auratus) and
rat (Long-Evans, hooded) were used as sources for in- termediate filament proteins.
Phosphorylation Goldfish and rat optic nerves were incubated in vitro
for 1 h with 1 mCi of [32P]orthophosphate. The incubation medium was as described elsewhere (Quitschke and
Schechter, 19836), except that the concentration of K,HP, was reduced to 0.01 mM.
Tissue preparations A cytoskeletal preparation was obtained from optic
nerves by a modification of a method described by Chiu and Norton (1982). All operations were carried out at 4°C unless otherwise indicated. The tissues were first ho- mogenized (50 mg/ml) in 50 mM Tris-HC1 (pH 6.8),2 mM EDTA, 2 mM EGTA, 0.5% (voVvol) Triton X-100, and 30% (wtlvol) sucrose. The homogenate was centrifuged at 13,000 g for 15 min. The myelin layer and the super- natant were removed and the cytoskeletal pellet was ho- mogenized in 50 mM Tns-HCI (pH 6.8), 2 mM EDTA, 2 mM EGTA, and 0.5% (voUvol) Triton X-100 and recen- trifuged at 13,000 g for 15 min. The resulting pellet was rehomogenized in 0.4% sodium dodecyl sulfate (SDS) and centrifuged at 20,000 g for 10 min. The clear supernatant was removed and mixed with five volumes of acetone, cooled to -20°C. The mixture was stored for at least 3 h at - 20°C and the precipitated protein was pelleted by centrifugation at 10,000 g for 5 min. The pellet was air- dried and homogenized in 0.5 ml of water. Prior to ly- ophilization, aliquots were removed for protein determi- nation (Lowry et al., 1951) or liquid scintillation counting (Quitschke and Schechter, 19836). The lyophilized pro- tein was dissolved at a concentration of about 3.75 mg/ ml in 0.05 M 2-(N-cyclohexylamino)ethanesulfonic acid (Ches) (Sigma) buffer (pH 9.0) containing 2% SDS, 1% dithiothreitol (DTT), and 5% glycerol. This solution was stored at - 80°C until used for electrophoresis. Gel electrophoresis
’Itnro-dimensional gel electrophoresis was generally per- formed as previously described (O’Farrell, 1975; Quitschke and Schechter, 1984). Isoelectric focusing gels were loaded with 20 pl of solution containing 75 pg of cytoskeletal protein. After isoelectric focusing the pro- teins were subjected to either enzymatic digestion or chemical cleavage within the first dimension by a modi- fication of previously described methods (Cleveland et al., 1977; Lam and Kasper, 1980). In this study the acryl- amide concentration of the second dimension running gel was increased to 15% (acry1amide:bisacrylamide 30: 1).
After electrophoresis in the second-dimension, gels were stained with Coomassie Blue or processed for au- toradiography as described elsewhere (Quitschke and Schechter, 1984). Partial enzymatic digestion
In experiments where the proteins are fragmented under denaturing conditions (Figs. 3-6 below), the iso- electric focusing gels were preequilibrated in 5 ml of 0.1% SDS and 0.125 M Tris (pH 6.8) for 10 min with three changes of solution (Figs. 3-6). Thereafter, the gels were immersed in 2.5 ml of the same buffer containing 6 pg/ ml Staphylococcus aureus protease V8 or chymotrypsin at either low (20 pg/ml) or high (100 pg/ml) enzyme con- centration. The gels were exposed to the enzyme solution for 20 min at 25°C. The gels were then transferred to 5 ml of equilibration buffer containing 10% (wtlvol) glyc- erol, 5% (voUvol) p-mercaptoethanol, 2.3% (wtlvol) SDS, and 0.0625 M Tris-HC1 (PH 6.8) for 5 min and then loaded onto the second dimension.
Chymotrypsin digestion of intermediate filaments under nondenaturing conditions (see Fig. 2 below) was
J . Neurochem.. Vol. 46. No. 2. 1986

OPTIC NERVE fNTERMEDfATE FfLAMENT PROTEINS 547
performed essentially as described by Rueger e t al. (1981). The final cytoskeletal pellet of goldfish or rat optic nerve was homogenized in 1 ml of buffer containing 50 mM Tris (pH 7.8) at a concentration of 1 mg proteidml. Chymotrypsin was added to a final enzymelprotein ratio of 1:100 and incubated at 25°C. Aliquots of 0.4 ml were removed after 10 rnin and 30 min of incubation and then 0.1 ml of 1% SDS was added. The aliquots were precip- itated with acetone and lyophilized as described above. The lyophilized protein was dissolved in equilibration buffer at a concentration of 2 mg/ml. Ten microliters of the solution was loaded onto the lanes of 10% acrylamide gels as previously described (Quitschke and Schechter, 1984).
Partial chemical cleavage The isoelectric focusing gels were transferred to 5 ml
of 0.1 M HCI containing 10 mg/ml of CNBr. The gels were shaken at 25°C for 30 rnin with three changes of solution. Then the gels were incubated in 5 ml of equilibration buffer for 30 rnin with at least six changes of buffer prior to electrophoresis in the second dimension.
Electrophoretic transfer Protein transfer from acrylamide gels was modified
from the method described by Towbin et al. (1979). The chamber buffer contained 20% methanol (vol/vol), 2.5 mM Tris, and 0.01% SDS. The transfer was completed after 90 rnin at 200 mA constant current.
Antibody incubation After the transfer, the nitrocellulose sheets were
blocked for 1 h in 3% bovine serum albumin (BSA), 50 mM Tris, and 0.01% SDS. The transfer was completed clonal antibodies to all intermediate filament proteins (AIF-antibody) were obtained from the supernatant of a hybridoma culture (Pruss et al., 1981). The hybridoma supernatant was diluted at a ratio of I: 10 with 0.3% BSA, 0.05% NP-40, 50 mM Tns-HC1 (pH 7.4), and 200 mM NaCI. After the blocking, each nitrocellulose sheet (ap- proximate dimensions, 3.5 x 5 cm) was incubated for at least 2 h in 4 ml of the antibody solution. The sheets were then washed three times for 5 min in buffer without the antibody. The second antibody stock solution was diluted at a ratio of 1500. The second antibody consisted of a horseradish peroxidase-conjugated anti-mouse IgG. The sheets were incubated for at least 2 h and then washed three times for 5 min with 20 mM Tris-HCI (pH 7). The nitrocellulose sheets were developed as follows: 5 ml of chloronaphthol solution (3 mg/ml in methanol) was added to 25 ml of 20 mM Tris-HC1 (pH 7). Four to six sheets were placed in this solution and I5 pI of 30% H,O, were added. The color reaction was stopped after 5-20 rnin by rinsing in water and the sheets were immediately photo- graphed. All operations were carried out at room tem- perature.
RESULTS
The intermediate filament proteins of goldfish and rat optic nerve
Major goldfish cytoskeletal proteins are found at molecular weights of 58K and 48K (Fig. 1A). In addition, a very basic 48K protein with several iso- electric variants is consistently observed in goldfish
FIG. 1. Cytoskeletal proteins of goldfish and rat optic nerve. A: Goldfish, Coornassie Blue. B: Goldfish, autoradiograrn (32P-labeled). C: Rat. Coornassie Blue. D: Rat, autoradiograrn (32P-labeled).
cytoskeletal preparations (arrowhead). The group of proteins at 58K can be separated into at least four isoelectric variants designated as ON,, ON,, ON,, and ON,. To the acidic side of ON,, additional components of decreasing concentrations can be re- solved. The arrow between ON, and ON, indicates a slowly transported protein (Quitschke and Schechter, 1984).
After incubating intact optic nerves in vitro in the presence of [32P]orthophosphate, ON, and ON, are phosphorylated, whereas no label is detected in ON, (Fig. 1B). Some label can be observed in the vicinity of the lower-molecular-weight end of ON,. Phosphorylated components are also observed to the acidic side of ON, and between ON, and ON, (arrow).
The group of proteins at 48K consists of several isoelectric variants that are differentially phosphor- ylated. The components within the 48K, bracket are predominant when stained with Coomassie Blue (Fig. 1A). No phosphate label is detected in these components. However, the minor components at 48K (Fig. 1A) are the most phosphorylated (48K,, Fig. 1B). Two phosphorylated spots are observed to the basic side of 48K, (arrowheads). These spots are not detectable by Coornassie Blue staining (Fig. lA), and it is possible that they represent phos- phorylated isoelectric variants of the very basic protein indicated by the arrowhead in Fig. IA.
J . Neworhem.. Vol. 46. N o . 2, 1986

548 W . QUITSCHKE AND N . SCHECHTER
The two major nonneuronal intermediate filament proteins in rat optic nerve are GFAP at 51K and vimentin at 58K (Fig. 1C). Each protein can be sep- arated into three major isoelectric variants desig- nated within brackets (Fig. 1C). Of these three var- iants, only components 2 and 3 are phosphorylated both in vimentin and GFAP (Fig. 1D). Three phos- phorylated proteins are present at approximately 26K which are not detectable by Coomassie Blue staining (Fig. lD, arrowheads). A differentially phosphorylated group at 64K (arrow) has been im- plicated as a neurofilament-associated protein (Pruss et al., 1981; Dahl et al., 1982).
Partial chymotrypsin hydrolysis- undenatured conditions
Cytoskeletal preparations from goldfish and rat optic nerves were subjected to proteolytic digest with chymotrypsin. Since the cytoskeletal prepa- ration represents a mixture of intermediate fila- ments with different protein composition, it is not possible to distinguish easily fragments from indi- vidual proteins after digestion. Therefore, the frag- ment pattern is considered the sum from the diges- tion of all the intermediate filament proteins in the tissue.
Analysis of the normal cytoskeleton of the gold- fish optic nerve reveals two predominant bands at 58K and 48K. Lower amounts of the higher-molec- ular-weight neurofilament proteins are observed (McQuarrie and Lasek, 1981; Quitschke and Schechter, 1985) at 80K and 145K (arrows; Fig. 2A, lane 1). Exposure of the cytoskeletal preparation to chymotrypsin generates a doublet of about 40K and 42K and a lower-molecular-weight fragment at about 24K (Fig, 2A, lanes 2 and 3; arrows). As a
flG. 2. Chymotrypsin digest of un- denatured goldfish and rat cyto- skeletal proteins. A Lane 1, goldfish, Coomassie Blue (control); lane 2,lO- min incubation; lane 3, 30-min in- cubation. B: Lane 1, goldfish, auto- radiogram (32P-labeled. control); lane 2. l0-rnin incubation; lane3,3O- min incubation. C: Lane 1. rat, Coo- massie Blue (control); lane 2,lO-min incubation; lane 3, 30-min incuba- tion. D: Lane 1, rat; autoradiogram PP-labeled, control); lane 2. 10-min incubation; lane 3, 30-min incuba- tion.
consequence of the exposure to chymotrypsin, phosphorylated fragments are no longer observed (Fig. 2B).
The rat cytoskeletal preparation contains GFAP, vimentin, and the neurofilament triplet at 70K, 150K and 200K (Fig. 2C, lane 1; arrows). Exposure to chymotrypsin results in the collapse of the major intermediate filament proteins into three major bands at 37-40K. In addition, bands are also ob- served at about 24K and lSOK molecular weight (Fig. 2C, lanes 2 and 3; arrows). Phosphate labeling is primarily observed in the band at 150K (Fig. 2C, lane 3; arrow). The fragments at 37-40K most likely represent a chymotrypsin-resistant structural core. This core consists of two a-helical domains that are linked approximately in the center by a short nonhelical domain (Geisler e t al., 1982u, 1983). Cleavage through this nonhelical region would result in fragments of 20-25K. Such frag- ments previously observed (Rueger et al., 1981; Geisler et al., 1982~) are observed here for both rat and goldfish. Phosphorylated moieties are rapidly removed from both goldfish and rat intermediate filament proteins in agreement with the concept that these groups are located in the more labile periph- eral domains adjacent to the structural core (Stei- nert et al., 1982). One exception is a band at 150K in rat cytoskeletal preparations. A stable fragment at 160K has been described, that is derived from the 200K neurofilament protein (Julien and Mu- shynski, 1983). That phosphorylated fragment was relatively resistant to further degradation by chy- motrypsin. It is possible that the 150K fragment ob- served here, although reminiscent of the 150K neu- rofilament protein, may be derived from the 200K protein.
1. Neurochem., Vol. 46. No. 2, 1986

OPTIC NERVE INTERMEDIATE FILAMENT PROTEINS 549
Partial chymotrypsin hydrolysis- denaturing conditions
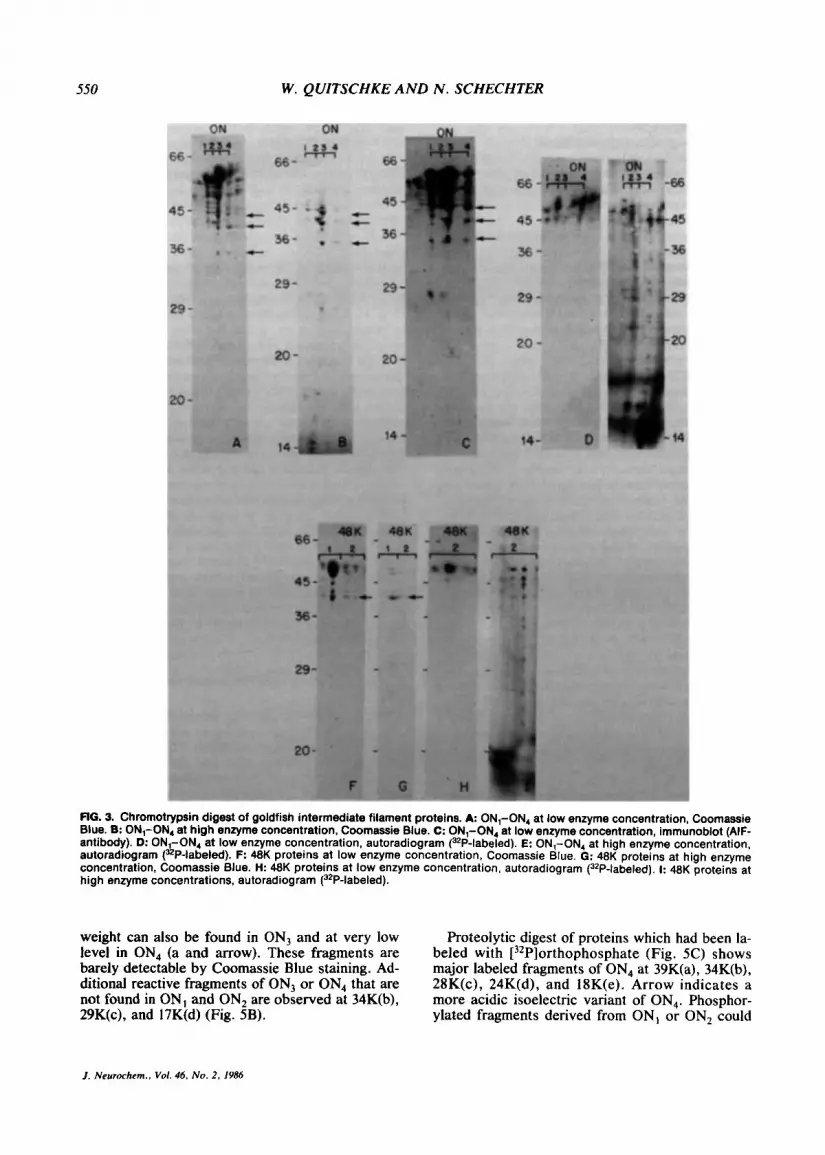
Digests within gels may be considered to be under denaturing conditions. It is likely that under such conditions cleavage sites are exposed which are not exposed under nondenaturing conditions. Digest patterns were compared at high and low chy- motrypsin concentrations for goldfish (Fig. 3) and rat (Fig. 4) optic nerve. The digest pattern of the goldfish proteins (ON,-ON,) is very similar at the lower chymotrypsin concentration (Fig. 3A). Nu- merous fragments are generated between 57K and 40K. When these fragments are transferred to ni- trocellulose paper and reacted with a monoclonal antibody which recognizes an epitope common to perhaps all intermediate filaments (AIF antibody, Pruss et al., 1981), they retain their reactivity (Fig. 3C). At the higher chymotrypsin concentration, the higher-molecular-weight fragments are rapidly con- verted to lower-molecular-weight fragments (Fig. 3B). Three major fragments of ON, and ON, are indicated by arrows in Fig. 3, A-C. The concen- tration of these fragments is relatively stable as the enzyme concentration is increased. It is therefore likely that the highest-molecular-weight fragments are converted to these three fragments before being digested into lower-molecular-weight fragments. The digest pattern of the phosphorylated proteins indicates that most of the phosphate label is re- tained in fragments which migrate close to the un- digested protein at low chymotrypsin concentra- tions (Fig. 3D). At high chymotrypsin concentra- tion, low levels of phosphate-labeled fragments are found over the entire molecular weight range, al- though most of the phosphate label is found at the bottom of the gel (Fig. 3E). This suggests that, al- though there is a wider range of cleavage sites avail- able to chymotrypsin digest in the denatured state, the phosphate-carrying portions of the molecule are still more likely to be cleaved off, as was observed in the undenatured state (Fig. 2A and B).
The goldfish 48K proteins are initially converted to fragments between 47K and 40K (Fig. 3F). A fragment at 40K is indicated (Fig. 3G) that appears more stable as the proteins are cleaved into smaller units at high enzyme concentrations. Digestion of the phosphorylated proteins follows a pattern sim- ilar to the 58K proteins. At low enzyme concentra- tion, the phosphorylated fragments are close to the undigested 48K proteins (Fig. 3H). At high enzyme concentration, numerous fragments are generated at low specific activity (Fig. 31).
Rat GFAP is relatively resistant to digestion at low chymotrypsin concentration (Fig. 4A). In ad- dition to the undigested protein, fragments are ob- served at 50-40K(a) and 32K(b). At higher enzyme concentrations, a similar pattern persists but the un- digested protein decreases in concentration and low-molecular-weight fragments appear at 28K(c)
and 18K(d) (Fig. 4B). Immunoreactive fragments are found at 50-40K(a), 32K(b), and 28K(c) (Fig. 4C). Phosphorylated fragments (Fig. 4D and E) are primarily observed at 50-40K(a) and in the smaller fragments at 22K(b) and 18K(c).
Digestion of rat vimentin at low chymotrypsin concentration yields fragments at 57-40K(a) and 33K(b) (Fig. 4A). At high enzyme concentration, a fragment at 52K(a) particularly stable as lower-mo- lecular-weight fragments appear at 15K(c) and 13K(d) (Fig. 4B). The reactivity of the AIF antibody is retained in the 57-40K(a) and 33K(b) fragments (Fig. 4C). Incorporated phosphate is also retained in these fragments, as in the lower-molecular- weight fragments at 18K(c) and 13K(d) (Fig. 4D and E). The fragment pattern of GFAP and vimentin is qualitatively similar to fragment patterns that have been obtained in other studies (Rueger et al., 1981; Evans and Fink, 1982). However, the relative con- centration of the chymotryptic fragments in this study is considerably lower. It is likely that there are factors that influence the digestion within gels that do not influence the digestion of proteins in solution. Digestion within gels requires the enzyme to diffuse into the gels for subsequent hydrolysis. Consequently, there is, at least initially, a concen- tration gradient between the peripheral and the cen- tral portions of the gel. Peripherally located protein will be more completely digested than centrally lo- cated protein, particularly if the generated frag- ments are susceptible to further hydrolysis. Inter- mediate-size fragments are rapidly converted to low-molecular-weight fragments as the enzyme dif- fuses into the gel.
Partial hydrolysis with Staphylococcus aureus protease V8
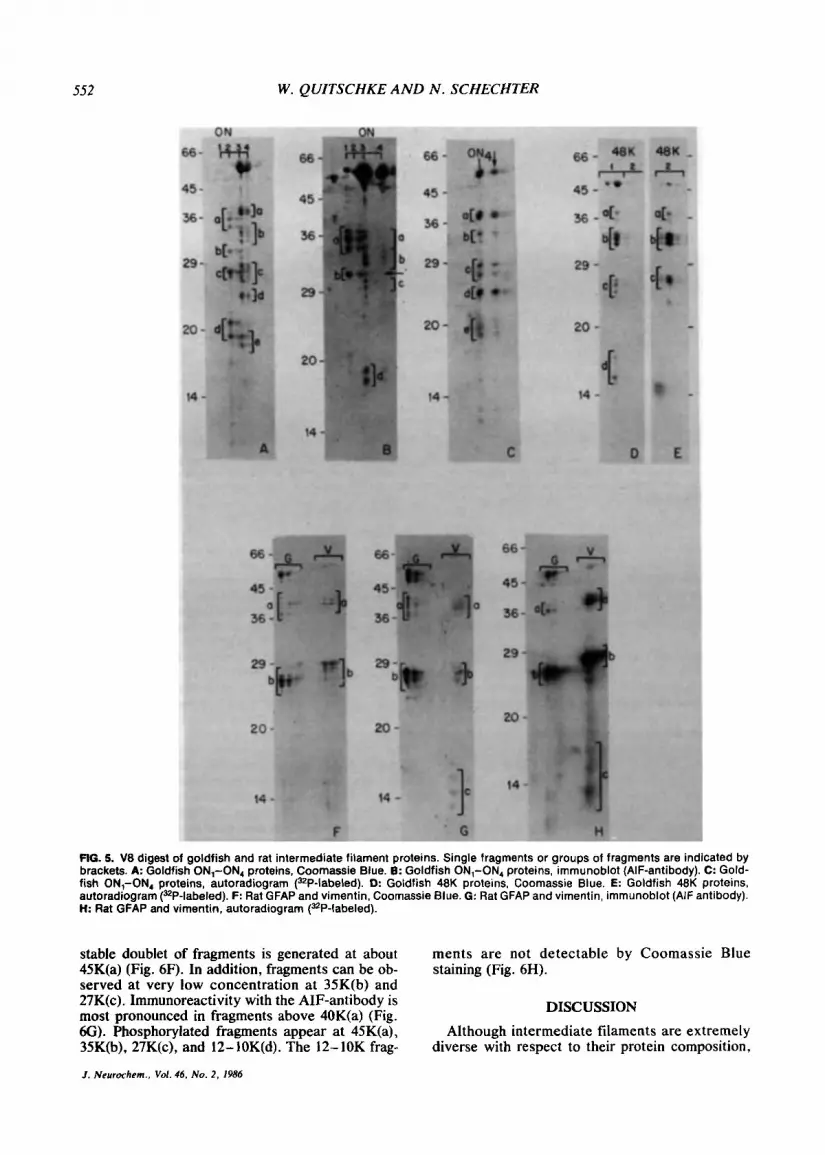
Partial proteolytic cleavage of goldfish ON, and ON, proteins (Fig. 5A) produced major groups of fragments at approximately 36K(a), 32K(b), 28K(c), and 20K(d). In contrast, ON, and ON, were more resistant to cleavage by V8 and residual undigested protein remained even at higher enzyme concentra- tions and longer digestion times (not shown). The major fragments derived from ON, and ON, were found at approximately 39K(a), 34K(b), 28K(c), 24K(d), and 18K(e). There is extensive overlap be- tween the fragment patterns of components ON, and ON,. It is also apparent that the fragment pat- terns of the neuronal ON, and ON, proteins are different from the fragment patterns of the nonneu- ronal ON, and ON, proteins, although several frag- ments appear to be common to all ON proteins.
When the digested fragments are transferred to nitrocellulose paper (Fig. 5B) and reacted with the AIF antibody, most of the reactivity of ON, and ON, is retained in the fragments at 36K(a) and 32K(b). Reactive fragments of identical molecular
J . Neurorhem., Vol. 46, No. 2. 1986
550 W . QUITSCHKE AND N . SCHECHTER
RG. 3. Chromotrypsin digest of goldfish intermediate filament proteins. A: ONl-ON4 at low enzyme concentration, Coomassie Blue. 0: ONl-ON, at high enzyme concentration, Coomassie Blue. C: ON,-ON4 at low enzyme concentration, immunoblot (AIF- antibody). D: ON -ON4 at low enzyme concentration, autoradiogram 132P-labeled). E: ON,-ON, at high enzyme concentration, autoradiogram (&P-labeled). F: 48K proteins at low enzyme concentration, Coomassie Blue. G: 48K proteins at high enzyme concentration, Coomassie Blue. H: 48K proteins at low enzyme concentration, autoradiogram (32P-labeled). I: 48K proteins at high enzyme concentrations, autoradiogram (3*P-labeled).
weight can also be found in ON, and at very low level in ON, (a and arrow). These fragments are barely detectable by Coomassie Blue staining. Ad- ditional reactive fragments of ON, or ON, that are not found in ON, and ON, are observed at 34K(b), 29K(c), and 17K(d) (Fig. 5B).
Proteolytic digest of proteins which had been la- beled with [32Plorthophosphate (Fig. 5C) shows major labeled fragments of ON, at 39K(a), 34K(b), 28K(c), 24K(d), and 18K(e). Arrow indicates a more acidic isoelectric variant of ON,. Phosphor- ylated fragments derived from ON, or ON, could
J. Neurorhem.. Vol. 46, N o . 2, 1986

OPTIC NERVE INTERMEDIATE FILAMENT PROTEINS 551
FIG. 4. Chymotrypsin digest of rat vimentin and GFAP. Single fragments or groups of fragments are indicated by brackets. A: Low enzyme concentration, Coomassie Blue. 8: High enzyme concentration, Coomassie Blue. C: Low enzyme concentration, immunoblot (AIF-antibody). D: Low enzyme concentration. autoradiogram (3zP-labeled). E: High enzyme concentration, auto- radiogram (32P-labeled).
not be conclusively demonstrated presumably be- cause of the low specific activity.
Partial digest with V8 of the goldfish 48K protein produces major fragments at 38K(a), 34K(b), 25K(c), and 16K(d) (Fig. 5D). Major phosphory- lated fragments are observed at 38K(a), 34K(b), and 25K(c).
Major fragments generated by the V8 digest of rat GFAP (Fig. 5F) are observed at 39K(a) and 27K(b). The AIF-antibody reactivity is strong with fragments close to the molecular weight range of the undigested protein (51 -48K), and somewhat lower with the fragments in the 39K(a) range (Fig. 5G). Strong reactivity is again observed with the fragments at 27K(b). The major phosphorylated fragments are also found at 27K(b) (Fig. 5H).
Vimentin in rat optic nerve shows major frag- ments at 41K(a) and 29K(b). In contrast to GFAP, all vimentin fragments have lost a significant part of their reactivity with the AIF antibody (Fig. 5G). Weak reactivity can be discerned at 41K(a), 29K(b), and also at 14- 18K(c). The major phosphorylated fragments (Fig. 2H) are found at 41K(a) and 29K(b).
Partial cleavage with cyanogen bromide Partial cleavage of the goldfish ON proteins
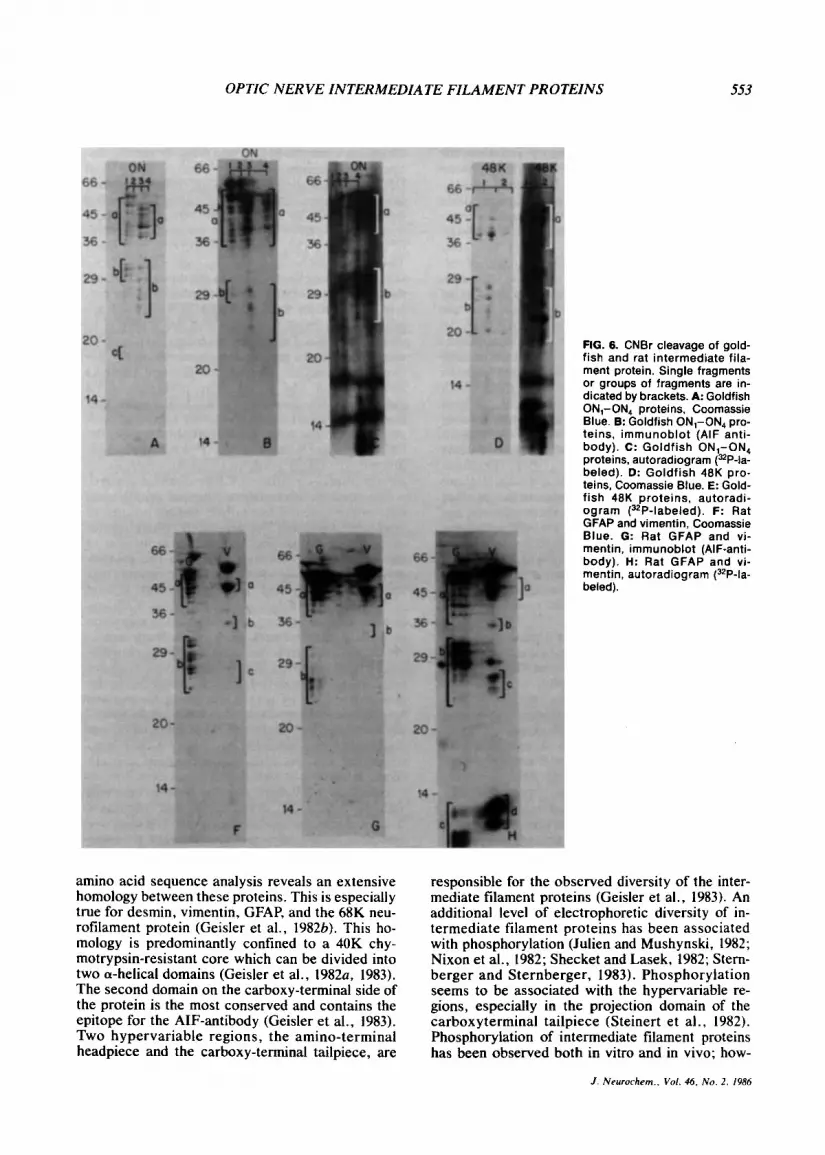
yields numerous fragments distributed over a wide molecular weight range (Fig. 6A). Proteins ON, and ON, display identical fragment patterns to each other as do proteins ON, and ON,. Although the electrophoretic mobility of fragments from ON, and
ON, are in many cases very similar to those of ON, and ON,, the two groups are clearly distinct. Pro- teins ON, and ON, display numerous fragments at 58-36K(a) and at 32-29K(b). In addition, one frag- ment is observed at 18K(c). Most fragments of ON, and ON, are concentrated in the region of 48- 37K(a) and 32-23K(b). [32P]Orthophosphate label is largely retained in the Coomassie Blue-stained fragments generated by partial CNBr cleavage of ON, and ON, (Fig. 6C). The reactivity of the AIF antibody is retained in all higher-molecular-weight fragments of ON,-ON, but diminishes in the lower- molecular-weight fragments (Fig. 6B).
The goldfish 48K protein generates several frag- ments with molecular weights of 47-38K(a) and 28- 20K(b). The phosphorylation pattern coincides with the pattern of the Coomassie Blue-stained frag- ments (Fig. 6E). In addition, phosphate-labeled fragments are observed below 10K. These frag- ments are barely detectable by Coomassie Blue staining.
Cleavage of rat GFAP results in fragments with decreasing concentration at 50-40K(a) and at 32- 23K(b) (Fig. 6F). The immunoreactivity of the frag- ments at 50-40K decreases with decreasing molec- ular weight (Fig. 6G). Immunoreactivity is also ob- served in the lower-molecular-weight fragments at 32-23K(b). Phosphorylated fragments are promi- nent at 50-40K(a) and at 37-23K(b) and <10K (c) (Fig. 6H).
CNBr cleaves rat vimentin so that a relatively
1. Neurochern.. Vol. 46, N o . 2. 1986
552 W . QUITSCHKE AND N . SCHECHTER
FIG. 5. V8 digest of goldfish and rat intermediate filament proteins. Single fragments or groups of fragments are indicated by brackets. A Goldfish ON,-ON, proteins, Coornassie Blue. 6: Goldfish ON,-ON, proteins, immunoblot (AIF-antibody). C: Gold- fish ON,-ON, proteins, autoradiogram (32P-labeled). D: Goldfish 48K proteins, Coomassie Blue. €: Goldfish 48K proteins, autoradiogram (=P-labeled). F: Rat GFAP and vimentin. Coomassie Blue. G: Rat GFAP and vimentin, immunoblot (AIF antibody). H: Rat GFAP and vimentin, autoradiogram (32P-labeled).
stable doublet of fragments is generated at about 45K(a) (Fig. 6F). In addition, fragments can be ob- served at very low concentration at 35K(b) and 27K(c). Immunoreactivity with the AIF-antibody is most pronounced in fragments above 40K(a) (Fig. 6G). Phosphorylated fragments appear at 45K(a), 35K(b), 27K(c), and 12-10K(d). The 12-10K frag-
ments are not detectable by Coomassie Blue staining (Fig. 6H).
DISCUSSION
Although intermediate filaments are extremely diverse with respect to their protein composition,
1. Neurochem., Vol. 46. No. 2 , 1986
OPTIC NERVE INTERMEDIATE FILAMENT PROTEINS 553
amino acid sequence analysis reveals an extensive homology between these proteins. This is especially true for desmin, vimentin, GFAP, and the 68K neu- rofilament protein (Geisler et al., 19826). This ho- mology is predominantly confined to a 40K chy- motrypsin-resistant core which can be divided into two a-helical domains (Geisler et al., 1982~. 1983). The second domain on the carboxy-terminal side of the protein is the most conserved and contains the epitope for the AIF-antibody (Geisler et al., 1983). Two hypervariable regions, the amino-terminal headpiece and the carboxy-terminal tailpiece, are
FIG. 6. CNBr cleavage of gold- fish and rat intermediate fila- ment protein. Single fragments or groups of fragments are in- dicated by brackets. A: Goldfish ON,-ON, proteins, Coomassie Blue. 8: Goldfish ON,-ON4 pro- teins, immunoblot (AIF anti- body). c: Goldfish ON,-ON4 proteins. autoradiogram (32P-la- beled). D: Goldfish 48K pro- teins, Coomassie Blue. E: Gold- fish 48K proteins, autoradi- ogram (32P-labeled). F: Rat GFAP and vimentin, Coomassie Blue. G: Rat GFAP and vi- mentin, immunoblot (AIF-anti- body). H: Rat GFAP and vi- mentin, autoradiogram (3ZP-la- beled).
responsible for the observed diversity of the inter- mediate filament proteins (Geisler et al., 1983). An additional level of electrophoretic diversity of in- termediate filament proteins has been associated with phosphorylation (Julien and Mushynski, 1982; Nixon et al., 1982; Shecket and Lasek, 1982; Stern- berger and Sternberger, 1983). Phosphorylation seems to be associated with the hypervariable re- gions, especially in the projection domain of the carboxyterminal tailpiece (Steinert et al., 1982). Phosphorylation of intermediate filament proteins has been observed both in vitro and in vivo; how-
J . Neurorhern.. Vol. 46, No. 2. 1986

554 W . QUITSCHKE AND N . SCHECHTER
ever, the amount of phosphate incorporated into the individual neurofilament proteins varies between the two procedures (Julien and Mushynski, 1981).
Although the goldfish optic nerve intermediate filament proteins (ONl-ON4) are unique interme- diate filament proteins (Quitschke and Schechter, 1984), the data presented here indicate that they share similar structural features with the higher ver- tebrate intermediate filament proteins. Chymo- trypsin digestion under nondenaturing conditions results in stable fragments at about 40K that are also observed for rat vimentin and GFAP (Fig. 2). Phosphorylated moieties appear to be located out- side the core region since they are preferentially cleaved off by chymotrypsin and not found associ- ated with the 40K core. This is true for the goldfish as well as the rat intermediate filament proteins (Fig. 2). In addition, the goldfish ON proteins con- tain the antigenic site within the core (Fig. 3) which is common to most intermediate filament proteins (Pruss et al., 1981; Geisler et al., 1983). Therefore, the molecular organization of the goldfish interme- diate filaments (ON,-ON,) is similar to that of other intermediate filaments. The same conclusion ap- plies to the goldfish proteins in the 48K molecular weight range except that these proteins do not dis- play the same degree of reactivity with the AIF an- tibody. The antigenic determinant within the core of this class of proteins may be significantly dif- ferent without altering the general molecular orga- nization of the filament.
Chymotrypsin digestion under denaturing condi- tions reveals a fragment pattern that is similar be- tween the goldfish ON proteins, the 48K proteins, rat GFAP, and vimentin. Although the molecular weight of the individual fragments vary, all the pro- teins studied here are cleaved into fragments having molecular weights between their respective undi- gested protein and 40K (Fig. 3). These fragments retain their reactivity with the AIF antibody. This suggests that under the denaturing conditions here, cleavage with chymotrypsin still preferentially takes place in the peripheral regions, leaving the core intact. Under harsher digestion conditions, fragments of lower molecular weights are gener- ated. With the exception of a few fragments be- tween 28K and 36K, these fragments do not react with this antibody, indicating the destruction of the epitope for the AIF antibody. A similar conclusion can be drawn from the fragment patterns of [32Plorthophosphate-labeled proteins. Although many phosphorylated fragments are detected over a wide molecular weight range, the specific activity of these fragments is comparatively low, suggesting that the phosphate-containing fragments have been largely cleaved off.
The cleavage points for CNBr and V8 may be considered more random along the protein than those of chyrnotrypsin. In these experiments the fragment patterns resulting from the hydrolysis of the goldfish ON proteins show that many fragments
which are common to ON, and ON, are clearly dis- tinct from fragments which are common to ON, and ON,, thus indicating structural differences between the two groups. The fragment patterns resulting from either CNBr or V8 hydrolysis clearly show that proteins ON, and ON, are structurally similar to each other. This is also true for ON, and ON,. The differences between the two groups (ON,, ON,/ ON3,0N4) most likely occur in the domains outside the central core. Despite the differences in the pat- tern of the major fragments, the AIF antibody ap- pears to recognize fragments that are common to all ON proteins. The fragments from CNBr and V8 cleavage reveal a greater structural variability be- tween the goldfish intermediate filament proteins and vimentin and GFAP. The preferential V8 cleavage points of vimentin and GFAP suggest that these two proteins are more closely related to each other than to the goldfish proteins. In contrast, the CNBr fragment pattern of GFAP is remotely remi- niscent to that of goldfish proteins whereas vi- mentin displays a different CNBr cleavage pattern.
The goldfish intermediate filament proteins, as well as rat vimentin and GFAP, display a range of isoelectric variants. The presented results cannot rule out differential phosphorylation as the source for some of the isoelectric variation. The goldfish 48K proteins consist of two isoelectric variants that are not phosphorylated. This suggests that, at least in this case, alternative posttranslational modifica- tion may be operating. Attempts to remove the phosphate enzymatically have so far been unsuc- cessful. In contrast to the 200K and 150K neurofil- ament proteins, the goldfish intermediate filament proteins, GFAP and vimentin, are resistant to de- phosphorylation by alkaline or acid phosphatase (unpublished observation). Also, the unphosphor- ylated isoelectric variants of these proteins do not display different solubility properties as has been suggested for the l5OK neurofilament protein (Wong et al., 1984).
Intermediate filament proteins have in common with the highly conserved actins and tubulins a structural core that is also conserved. The diversity associated with intermediate filament proteins stems from the variable regions adjacent to the core. It is interesting to speculate that the con- served core region meets structural requirements common to many cell types. The variable regions meet additional functions such as, for example, dif- ferentiation, development, and plasticity, which can be associated with specific cell types.
Acknowledgment: This research was supported by a grant from the National Institutes of Health (EY 05212) to Nisson Schechter.
REFERENCES Benowitz L. I. and Lewis E. R. (1983) Increased transport of
44,OOO- to 49,000-dalton acidic proteins during regeneration

OPTIC NERVE INTERMEDIATE FILAMENT PROTEINS 555
of the goldfish optic nerve: a two-dimensional gel analysis. J. Neurosci. 3, 2153-2163.
Bignami A, , Raju T., and Dahl D. (1982) Localization in vi- mentin, the nonspecific intermediate filament protein in em- bryonal glia and in early differentiating neurons. Dev. Biol.
Chiu F.-C. and Norton W. T. (1982) Bulk preparation of CNS cytoskeleton and the separation of individual neurofilament proteins by gel filtration: dye-binding characteristics and amino acid compositions. J . Neurochem. 39, 1252- 1260.
Cleveland D. W., Fischer S. G., Kuschner M. W., and Laemmli U. K. (1977) Peptide mapping by limited proteolysis in so- dium dodecyl sulfate and analysis by gel electrophoresis. J . Biol. Chem. 252, 1102-1 106.
Cochard P. and Paulin D. (1984) Initial expression of neurofila- ments and vimentin in the central and peripheral nervous system of the mouse embryo in vitro. J . Neurosci. 4, 2080- 2094.
Dahl D., Strocchi P., and Bignami A. (1982) Vimentin in the central nervous system. Differentiation 22, 189- 190.
Easter S. S. and Stuermer C. A. 0. (1984) An evaluation of the hypothesis of shifting terminals in goldfish optic tectum. J . Neurosci. 4, 1052-1063.
Evans R. M. and Fink L. M. (1982) An alteration in the phos- phorylation of vimentin-type intermediate filaments is as- sociated with mitosis in cultured mammalian cells. Cell 29, 43-52.
Franke W. W., Schmidt E., Schiller D. L., Winter S., Jarasch E. D., Moll R., Denk H., Jackson B. W., and Illmensee K. (1982) Differentiation-related patterns of expression of pro- teins of intermediate-size filaments in tissues and cultured cells. Cold Spring Harbor Symp. Quant. Biol. 46, 431-454
Geisler N., Kaufmann E., and Weber K. (1982a) Proteinchemical characterization of three structurally distinct domains along the protofilament unit of desmin 10 nm filaments. Cell 30, 277- 286.
Geisler N., Plessmann U., and Weber K. (19826) Related amino acid sequences in neurofilaments and non-neuronal inter- mediate filaments. Nature 2%, 448-450.
Geisler N., Kaufmann E., Fischer S., Plessman U. , and Weber K. (1983) Neurofilament architecture combines structural principles of intermediate filaments with carboxy-terminal extensions increasing in size between triplet proteins. EMBO J . 2, 1295-1302.
Giulian D., Ruisseaux H. D., and Cowburn D. (1981) Biosyn- thesis and intraaxonal transport of proteins during neuronal regeneration. J. B i d . Chem. 255, 6494-6501.
Heacock A. M. and Agranoff B. W. (1976) Enhanced labeling of retinal protein during regeneration of the optic nerve in gold- fish. Proc. Natl. Acad. Sci. USA 73, 878-882.
Hoffman P. N. and Lasek R. J. (1975) The slow component of axonal transport: identification of major structural polypep- tides of the axon and their generality among mammalian neurons. J . Cell Biol. 66, 351-366.
Julien J.-P. and Mushynski W. E. (1981) Comparison of in vitro and in vivo-phosphorylated neurofilament polypeptides. J . Neurochem. 37, 1579- 1585.
Julien J.-P. and Mushynski W. E. (1982) Multiple phosphoryla- tion sites in mammalian neurofilament polypeptides. J . Biol. Chem. 257, 10467-10470.
Julien J.-P. and Mushynski W. E. (1983) The distribution of phos- phorylation sites among identified proteolytic fragments of mammalian neurofilaments. J . B i d . Chem. 258, 4009-4025.
Lam K. S. and Kasper C. B. (1980) Sequence homology analysis of a heterogeneous protein population by chemical and en- zymatic digestion using a two-dimensional sodium dodecyl sulfate-polyacrylamide gel system. Anal. Biochem. 108, 220-226.
Lazarides E. (1982) Intermediate filaments: a chemically heter- ogeneous developmentally regulated class of proteins. Ann. Rev. Biochem. 51, 219-250.
Lowry 0. H., Rosebrough N. J., Farr A. L., and Randall R. J. (1951) Protein measurement with the Folin phenol reagent. J . Biol. Chem. 193, 265-275.
91, 286-295.
McQuame I. G. and Lasek R. J. (1981) Axonal transport of la- beled neurofilament proteins in goldfish optic axons. J. Cell Biol. 91, 234a.
Meyer R. L. (1978) Evidence from thymidine labeling for con- tinuing growth of retina and tectum in juvenile goldfish. Exp. Neurol. 59, 99- 11 1.
Nixon R. A., Brown B. A., and Marotta C. A. (1982) Post-trans- lational modification of a neurofilament protein during axo- plasmic transport: implications for regional specialization of CNS axons. J . Cell B i d . 94, 150-158.
O’Farrell P. H. (1975) High resolution two-dimensional electro- phoresis of proteins. J . B i d . Chem. 250, 4007-4021.
Perry G. W., Burmeister D. W., and Grafstein B. (1985) Changes in protein content of goldfish optic nerve during degenera- tion and regeneration following nerve crush. J . Neurochem. 44, 1142-1151.
Pruss R. M., Mirsky R., Raff M. C., Thorpe R., Dowding A. S., and Anderton B. H. (1981) All classes of intermediate fila- ments share a common antigenic determinant defined by a monoclonal antibody Cell 27, 419-428.
Quitschke W. and Schechter N. (1983b) In vitro protein synthesis teins during regeneration of the goldfish retinotectal pathway. Brain Res. 258, 69-78.
Quitschke W. and Schechter N. (19830) In virro protein synthesis in the goldfish retinotectal pathway during regeneration: Ev- idence for specific axonal proteins of retinal origin in the optic nerve. J . Neurochem. 41, 1137-1 142.
Quitschke W. and Schechter N. (1984) 58,000 Dalton interme- diate filament proteins of neuronal and nonneuronal origin in the goldfish visual pathway. J . Neurochem. 42, 569-576.
Quitschke W., Jones P. S., and Schechter N. (1985) Survey of intermediate filament proteins in optic nerve and spinal cord: evidence for differential expression. J . Neurochem. 44, 165-1476.
Rueger D. C., Gardner E. E., Simonian H. D., Dahl D., and Bignami A. (1981) Purified glial fibrillary acidic protein and desmin are distinct intermediate filament proteins exhibiting similar properties. J . Biol. Chem. 256, 10606-10612.
Schechter N., Francis A., Deutsch D. G., and Gazzaniga M. S. (1979) Recovery of tectal nicotinic-cholinergic receptor sites during optic nerve regeneration in goldfish. Brain Res. 166, 57-64.
Shaw G., Osborn M., and Weber K. (1981) An immunofluores- cence microscopical study of the triplet proteins, vimentin and glial fibrillary acidic protein within the adult rat brain. Eur. J . Cell Biol. 26, 68-82.
Shecket G. and Lasek R. S. (1982) Neurofilament protein phos- phorylation: species generality and reaction characteristics. J. Biol. Chem. 257, 4788-4795.
S p e w R. W. (1963) Chemoaffinity in the orderly growth of nerve fiber patterns and connections. Proc. Natl. Acad. Sci. USA 50, 703-710.
Steinert P. M., Wantz M. L., and Idler W. W. (1982) 0-Phos- phoserine content of intermediate filament subunits. Bio- chemistry 21, 177-183.
Sternberger L. A. and Sternberger N. H. (1983) Monoclonal an- tibodies distinguish phosphorylated and nonphosphorylated forms of neurofilaments in siru. Proc. Natl. Acad. Sci. USA
Tapscott S. S.. Bennett G. S., Toyama Y., Kleinbart F.. and Holtzer H. (1981) Intermediate filament proteins in the de- veloping chick spinal cord. Dev. B i d . 86, 40-54.
Towbin H., Staehelin T., and Gordon J. (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellu- lose sheets: procedure and some application. Proc. Natl. Acad. Sci USA 76, 4350-4354.
Weber K., Shaw G., Osborn M., Debus E., and Geisler N. (1983) Neurofilaments, a subclass of intermediate filaments: struc- ture and expression. Cold Spring Harbor Symp. Quant. Biol. 48, 717-719.
Wong J., Hutchinson S. B., and Liem R. K. H. (1984) An iso- electric variant of the 150,000-dalton neurofilament poly- peptide. J . B i d . Chem. 259, 10867- 10874.
80, 6126-6130.
Copyright © 2022 FDOKUMEN




















