
Studies on the Terminal Nerve and Its Central Connections in Goldfish
12
Studies on the Terminal Nerve and Its Central Connections in Goldfish” HARRY E. SLOAN AND LEO S. DEMSKI University of Kentucky School of Biological Sciences Lexington, Kentucky 40506 INTRODUCTION In goldfish, pheromones play an important role in sexual behavior and it has been proposed that the terminal nerve (TN) detects these stimuli and triggers sexual behavior including sperm release (SR).’ TN ganglion cells lie in a medioventral area of the olfactory bulb and send processes rostrally toward the olfactory epithelium and caudally in the medial olfactory tract (MOT) to the ventral telencephalon, ventral nucleus of area ventralis telencephali (Vv), supracommissural nucleus of area ventralis telencephali (Vs) and preoptic area (POA). Some cells also project to the retina.’-’ Several functional studies suggest TN involvement in sexual responses. Electrical stimulation of the medial bundle of MOT, the area containing the TN, elicits repro- ductive behavior in cod4 and sperm release in goldfi~h.~ Moreover, bilateral transection of MOT6 and lesions of the caudal Vv-rostra1 Vs complex and the POA produce deficits in sexual behavior’.’ whereas stimulation of the POA evokes sex behavior and gamete Both forebrain targets contain sex-steroid concentrating neurons and fibers with luteinizing-hormone releasing hormone (LHRH), a peptide that fa- cilitates reproductive behavior.” At least some of these fibers probably originate from the LHRH-containing ganglion cells of the TN. It has been suggested that in goldfish TN fibers release LHRH in the POA and ventral telencephalon in response to sex pheromones, thereby facilitating sex responses controlled by the nuclei. The POA has direct connections to spinal areas thought to mediate SR and perhaps other sexual responses.’* Thus, the preopticospinal system may be the final mediator of TN-induced responses. Connections between the ventral telencephalon and the POA have been suggested in landlocked red ~almon,’”’~ but have not been adequately demonstrated in goldfish. To further examine the possible role of the T N in sexual behavior, we have carried out brain lesion, stimulation and neuroanatomical studies in goldfish. The results of three experiments are generally consistent with the hypothesis that the T N influences sexual activity, i.e., (1) TN cells can be labeled with HRP placed on the cut optic nerve in preparations with only mMOT intact; (2) intracranial LHRH appears to ‘This work was supported in part by Grant NS19431 from the National Institutes of Health and by Biomedical Research Support Funds. 42 1
Transcript of Studies on the Terminal Nerve and Its Central Connections in Goldfish

Studies on the Terminal Nerve and Its Central Connections in Goldfish”
HARRY E. SLOAN AND LEO S. DEMSKI
University of Kentucky School of Biological Sciences Lexington, Kentucky 40506
INTRODUCTION
In goldfish, pheromones play an important role in sexual behavior and it has been proposed that the terminal nerve ( T N ) detects these stimuli and triggers sexual behavior including sperm release (SR).’ T N ganglion cells lie in a medioventral area of the olfactory bulb and send processes rostrally toward the olfactory epithelium and caudally in the medial olfactory tract (MOT) to the ventral telencephalon, ventral nucleus of area ventralis telencephali (Vv), supracommissural nucleus of area ventralis telencephali (Vs) and preoptic area (POA). Some cells also project to the retina.’-’ Several functional studies suggest TN involvement in sexual responses. Electrical stimulation of the medial bundle of MOT, the area containing the TN, elicits repro- ductive behavior in cod4 and sperm release in goldf i~h.~ Moreover, bilateral transection of MOT6 and lesions of the caudal Vv-rostra1 Vs complex and the POA produce deficits in sexual behavior’.’ whereas stimulation of the POA evokes sex behavior and gamete Both forebrain targets contain sex-steroid concentrating neurons and fibers with luteinizing-hormone releasing hormone (LHRH), a peptide that fa- cilitates reproductive behavior.” At least some of these fibers probably originate from the LHRH-containing ganglion cells of the TN. It has been suggested that in goldfish TN fibers release LHRH in the POA and ventral telencephalon in response to sex pheromones, thereby facilitating sex responses controlled by the nuclei. The POA has direct connections to spinal areas thought to mediate SR and perhaps other sexual responses.’* Thus, the preopticospinal system may be the final mediator of TN-induced responses. Connections between the ventral telencephalon and the POA have been suggested in landlocked red ~a lmon, ’”’~ but have not been adequately demonstrated in goldfish.
To further examine the possible role of the T N in sexual behavior, we have carried out brain lesion, stimulation and neuroanatomical studies in goldfish. The results of three experiments are generally consistent with the hypothesis that the T N influences sexual activity, i.e., (1) T N cells can be labeled with H R P placed on the cut optic nerve in preparations with only mMOT intact; (2) intracranial LHRH appears to
‘This work was supported in part by Grant NS19431 from the National Institutes of Health and by Biomedical Research Support Funds.
42 1

422 ANNALS NEW YORK ACADEMY OF SCIENCES
modulate SR; and (3) connections between steroid-concentrating ventral telencephalic nuclei and the POA can be demonstrated using H R P tracing.
METHODS AND MATERIALS
Male goldfish, Curussius uumtus, were maintained at 20°C on a 15-hr light cycle and given intraperitoneal injections of 100-300 IU chorionic gonadotropin (Sigma) to produce spermiation. For brain stimulation (Experiments 1 and 2), fish were anesthetized in 2% urethane and curarized with an injection of 1.36 ng/g body weight before being secured in a modified stereotaxic holder. Silver hook electrodes were used to stimulate the olfactory tracts using 10-300 p A at 50 Hz with 1-msec duration stimuli. In Experiment 1 prelesion threshold values for evoking SR were determined. The lateral olfactory tract (LOT) and the lateral portion of the medial olfactory tract (IMOT) were then sectioned, leaving the medial portion of MOT (mMOT) intact, and postlesion threshold values were obtained for comparison. SR was also evoked from the exposed but unsectioned contralateral OT. After testing, H R P type VI (Sigma) crystals were applied to both sectioned optic nerves. The dorsal cranium and optic capsules were sealed with plastic and cyanoacrylate glue and covered by dental cement. Animals were allowed to survive 4-6 days and their brains processed for HRP histochemistry.” The olfactory bulbs, epithelium, tracts and brain were hori- zontally sectioned as a unit a t 40 pm. The number of HRP-filled T N cells was determined. Only cells with a nucleus in the section were counted.
In Experiment 2, threshold values for SR from stimulation of the intact mMOT were determined and 100-pg LHRH/ml (Sigma) was injected into the third ventricle. Postinjection thresholds were then determined. In several cases, SR was evoked by a POA stimulation protocol9 and milt discharges were measured through a hollow tube placed under the urogenital opening. The tube contained a light source and a pho- tosensitive cadmium-sulfide resistor as part of a bridge circuit in which blockage of light results in a higher voltage output to an analog-to-digital converter on a Nicolet 1 170 signal averager.
Behavioral experiments (Experiment 1) tested the effect of olfactory lesions on male courtship and then determined the number of T N cells that filled when HRP was applied to the optic nerve. Courtship behavior was triggered using a stimulus female prepared by injection (females) with 200-ng prostaglandin F2 a / g body weight (Upjohn).’ Males that spawned within 15 minutes after the introduction of the stimulus female were selected for the experiment. Treatment of males involved transection of an OT and section of the opposite LOT, thus leaving one MOT intact. Posttreatment tests for components of spawning behavior (i.e., following, quivering, rising and spawning) were carried out either 2 or 5 days later. H R P was then applied to the optic nerves, the animals allowed to survive for 5 days, and tissue processed as described earlier.
In Experiment 3, ventral telencephalic projections to the POA were examined. Preliminary findings were obtained from two animals. Four percent H R P (type VI Sigma) was deposited in the POA by either pressure injection of 4 -80 nl in phosphate buffer (pH 7.4) using a hydraulic system” or iontophoresis in 0.15 M Tris buffer (Sigma) for 30 minutes at 600 nA + current.I6 H R P was placed into the third ventricle as a control against spurious uptake. Survival times and tissue processing were as described above. Cell diameter was determined by averaging the long axis

SLOAN Br DEMSKI: TERMINAL NERVE IN GOLDFISH 423
and perpendicular short axis of the soma. Standardized nomenclatures were adapted for telencephalic ” and diencephalic structures.
RESULTS
Experiment I : HRP Tracing of Functional OT Pathways
The relationship between the ability to evoke SR from OT stimulation and the integrity of the TN projections to the retinae was examined. SR was elicited by
STIMULATION
Optk‘ N.
FIGURE 1. Dorsal view of goldfish brain showing the terminal nerve (TN) pathway via the medial olfactory tract (MOT) to the retina and the supracommissural nucleus of the ventral telencephalon (Vs). Sperm release (SR) was evoked by simulation of the olfactory tract before and after transection (arrowhead) of the ipsilateral lateral olfactory tract (LOT) and in some cases the lateral part of the MOT. TN cells were retrogradely labeled by application of HRP to the optic nerve.
stimulation of the intact olfactory tracts and, following surgery, the medial portion of the left MOT (mMOT in FIGURE 1). Prelesion thresholds of the left OT were 10 pA (fish 3), 41 pA (fish 4), 65 pA (fish 6), 150 pA (fish 9) and 20 pA (fish 13). Following section of the left LOT and the lateral portion of MOT (IMOT), i.e., only mMOT remaining, thresholds were 50 pA (fish 3), 62 pA (fish 4), 120 pA (fish 6), 160 pA (fish 9), and 45 pA (fish 13). The increases could be due to damaging some axons controlling SR and/or, perhaps, differences in the effectiveness of the original and final electrode placement. Retrograde transport of HRP from the optic nerves to

424 ANNALS NEW YORK ACADEMY OF SCIENCES
TN cells of the experimental and ipsilateral sides was successfully demonstrated in two animals. Ratios of ipsilaterally /contralaterally filled cells were 18/41 in animal 4 and 11/57 in fish 13. The decrease in ipsilaterally labeled cells could be a result of accidentally damaging axons of TN within the lateral portion of mMOT or loss of fibers in LOT and IMOT. Thus, in preparations in which mMOT sustained SR, some of the TN cells that project to the retina via mMOT are intact. Rostrally directed processes of labeled T N cells were not found after application of H R P to sectioned optic nerves.
The relationship of OT damage to T N fillings and chemically mediated sexual behavior was also studied. Two males that spawned with a stimulus female within 15 minutes later received OT sections resulting in a single intact mMOT, i.e., one complete OT transection and the other LOT and IMOT cut. The fish were tested at either 2 or 5 days posttreatment and failed to spawn. They did, however, show “following” and “quivering, ” two major components of sexual behavior, thus indicating an interest in the stimulus female. Application of HRP to cut optic nerves revealed labeled cells ipsilateral to the intact mMOT (28 and 38). As expected, labeled cells were not present on the side of the transected OT.
Experiment 2: LHRH Effects on SR
As an extension of earlier preliminary findings: we studied the effects of intracranial LHRH on evoked SR. Initially, SR was consistently evoked by stimulation of the intact OT (threshold of 250 pA) in an anesthetized and immobilized fish (TABLE 1). Triggering levels dropped to 20 p A within 16 minutes following injection of 0.5 pI of LHRH ( 1 mg/ml) into the third ventricle. Fifteen minutes later (31 minutes postapplication) threshold levels increased to 100 PA. A second injection of LHRH again lowered the threshold to 20 p A within 15 minutes.
TABLE 1. Effects of LHRH upon Thresholds for Evoking Sperm Release by Olfactory Tract Stirnulation in Fish 13
Time (min)
Responses to Stimulation of Left Olfactory Tract (thresholds in pA)
0 No response at 100 Weak response at 175-200 Moderate response at 250
4 Moderate response at 250 9 5 p1 of LHRH (1 mg/ml) in teleost Ringer’s into the 3rd ventricle
10- 12 Strong response by 10 sec at 250 Good response by 10 sec at 200
19 Some milt at 90 20 Some milt at 90 21 24 Weak response at 30 25 26 No response at 10
Weak response at 50 (repeated three times)
Response at end of test at 20

SLOAN 61 DEMSKI: TERMINAL NERVE IN GOLDFISH 425
l K 1 IK I
A
FIGURE 2. Averaged sperm release (SR) re- sponses before (A) and after (B) intracranial application of LHRH (100 pg/ml). Eight preinjection tests and eight postinjection tests were run. Note the decreased latency of SR onset and increased milt discharge in (B) com- pared to (A).
I 0 20 40
Seconds
In several other animals, the effects of LHRH upon the latency of SR and the relative amount of milt released were measured using a photodetector input into a signal averager. A typical example is illustrated in FIGURE 2 and described below. The average of eight stimulations of the preoptic area before and after the application of LHRH to the intracranial cavity (as above) were compared. The LHRH treatment resulted in an average 6.2-sec reduction in latency and an increase in the amount of milt released. The latter was estimated by the relative blockage of light to photosensitive resistor and subsequent change in voltage to the signal averager. The increase was obvious under visual inspections as well as in the semiquantitative recording. Analog voltages were digitized and converted to a number directly proportional to the thickness of the SR stream. The counts were integrated over the period of SR and averaged for the eight tests. The total count for the untreated controls (71,736) was substantially less than for the LHRH-treated fish (105,818). Similar decreases in latency of response and increases in the amount of milt were not observed after the intraventricular application of teleost ringers, the vehicle used to deliver the LHRH.
Experiment 3: Anatomic Studies of Basal Forebrain Systems
Cells in the telencephalon and diencephalon were labeled following H R P injections into the POA. Iontophoretic delivery resulted in discrete applications of H R P with minimal diffusion beyond the designated border of the nucleus. Larger amounts of HRP were delivered by pressure injection with greater diffusion into adjacent nuclei. The observations presented are primarily from the iontophoretic material, with con- firmation by results using injection.
HRP was deposited into the nucleus preopticus parvocellularis posterioris (PPp) of the POA. The periventricular POA has been found to concentrate sex steroids and to be an area from which SR and sexual behavior can readily be evoked.'.'' All labeled cells within the telencephalon and diencephalon were ipsilateral to the injection site.

426 ANNALS NEW YORK ACADEMY OF SCIENCES
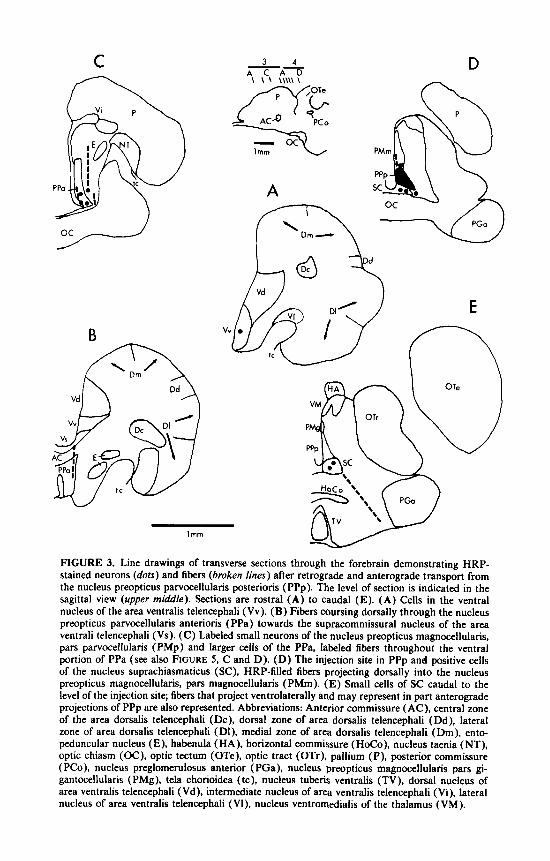
The most rostral telencephalic group labeled was the ventral nucleus of the area ventralis telencephali (Vv; FIGS. 3A and 5A). In transverse sections filled cells were oval with diameters of 7 p m (n = 2 cells, 6-8 p m ) whereas while in horizontally sectioned material, the cells measured 9 pm in diameter (n = 7 cells, range 6-10 p m ) and appeared bipolar with processes extending rostrally and caudally. At the level of the anterior commissure, labeled fibers of unknown origin were found lateral to the nucleus preopticus parvocellularis anterioris (PPa) and were directed dorsally toward the supracommissural nucleus of the area ventralis telencephali (Vs) (FIG. 3B). No other telencephalic nuclei were labeled.
Diencephalic nuclei containing filled cells were rostral, at the level of, and caudal to the injection site. The former were located in nucleus preopticus magnocellularis pars parvocellularis (PMp) and the adjacent nucleus preopticus parvocellularis an- terioris (PPa) (FIG. 3C). Labeled cells of PMp and PPa were in the ventral portions of their respective regions. In transverse sections PMp cells were round to oval (diameter of 8pm; n = 41 cells; range, 5-1 1 p m ) while labeled PPa cells were round to spindle shaped (8 pm in diameter; n = 18 cells, range, 6-12 pm). A labeled PPa neuron with both dorsally and ventrally oriented processes is shown in FIGURE 5C,D.
At the level of the injection site, a few cells of nucleus preopticus magnocellularis pars magnocellularis (PMm) and nucleus suprachiasmaticus (SC) were labeled. HRP- containing neurons and fibers of unknown origin were located in the ventral half of the PMm and cells were 11 p m in diameter (n = 6 cells; range, 8-14 pm). Most labeled SC neurons were ventral to the injection site (FIG. 3D). Other SC cells were caudal to the injection site, ranged in size from 6-13 p m with an average of 8 p m in diameter (n = 41 cells), and were otherwise similar to those in the PMp. Labeled fibers extend ventrolaterally from the SC (FIG. 3E). They were predominantly ipsi- lateral. The fibers traced a pathway similar to the preoptico-pituitary projection (per- sonal observations and those of others”).
Caudal to the injection site, labeled cells were also present in the dorsal hypotha- lamic zone (Hd), ventral hypothalamic zone (Hv) and the central posterior thalamic nucleus (CP). Filled neurons of Hd were located immediately lateral to the ependyma and dorsal to the anterior tuberal nucleus (TA). At this level, labeled fibers were also dorsolateral to Hd, while other HRP-containing axons were ventrolateral to TA and dorsal to the ventromedial nucleus of the thalamus (VM; FIG. 4A and B). Hd cells have processes extending dorsally and ventrally and measure 8 p m in diameter (n = 4, range, 8-9 p m ) (FIG. 5F). Filled neurons were also found lateral to the ependyma in Hv in the zone ventral to TA, and in the rostral and caudal portions of CP (FIG. 4C and D). HV cells were oval (10 p m in diameter; n = 2; range, 8- 12 p m ) with dorsally and ventrally directed processes (FIGS. 4A and 5B). Rostra1 C P neurons were subependymal while more caudal cells in the nucleus were more lateral. Labeled C P neurons were 6 p m in diameter (n = 3, range, 5-9 p m ) (FIG. 5E). HRP-filled cells were not found caudal to CP.
DISCUSSION
Our results generally support a T N modulation of sexual behavior’ including SR.5 We have demonstrated that: ( 1 ) the TN-retinal projection can be labeled following either elicited courtship behavior or evoked SR with the mMOT as the only connection between olfactory epithelium and the brain (Experiment 1 ); (2) intracranial LHRH
C 3 4 -- A C A D \ \ \ \\\\\ \
A
1 mm
D
E
FIGURE 3. Line drawings of transverse sections through the forebrain demonstrating HRP- stained neurons (dots) and fibers (broken lines) after retrograde and anterograde transport from the nucleus preopticus parvocellularis posterioris (PPp). The level of section is indicated in the sagittal view (upper middle). Sections are rostra1 (A) to caudal (E). (A) Cells in the ventral nucleus of the area ventralis telencephali (Vv). (B) Fibers coursing dorsally through the nucleus preopticus parvocellularis anterioris (PPa) towards the supracommissural nucleus of the area ventrali telencephali (Vs). (C) Labeled small neurons of the nucleus preopticus magnocellularis, pars parvocellularis (PMp) and larger cells of the PPa, labeled fibers throughout the ventral portion of PPa (see also FIGURE 5 , C and D). (D) The injection site in PPp and positive cells of the nucleus suprachiasmaticus (SC), HRP-filled fibers projecting dorsally into the nucleus preopticus magnocellularis, pars magnocellularis (PMm). (E) Small cells of SC caudal to the level of the injection site; fibers that project ventrolaterally and may represent in part anterograde projections of PPp are also represented. Abbreviations: Anterior commissure ( AC), central zone of the area dorsalis telencephali (Dc), dorsal zone of area dorsalis telencephali (Dd), lateral zone of area dorsalis telencephali (DI), medial zone of area dorsalis telencephali (Dm), ento- peduncular nucleus (E), habenula (HA), horizontal commissure (HoCo), nucleus taenia (NT), optic chiasm (OC), optic tectum (OTe), optic tract (OTr), pallium (P), posterior commissure ( PCo), nucleus preglomerulosus anterior (PGa), nucleus preopticus magnocellularis pars gi- gantocellularis (PMg), tela chorioidea (tc), nucleus tuberis ventralis (TV), dorsal nucleus of area ventralis telencephali (Vd), intermediate nucleus of area ventralis telencephali (Vi), lateral nucleus of area ventralis telencephali (Vl), nucleus ventromedialis of the thalamus (VM).
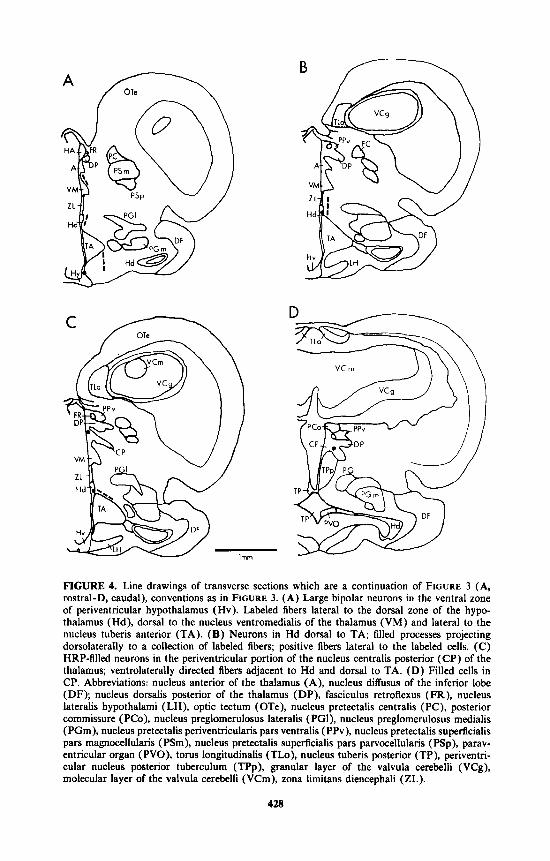
FIGURE 4. Line drawings of transverse sections which are a continuation of FIGURE 3 (A, rostral-D, caudal), conventions as in FIGURE 3. (A) Large bipolar neurons in the ventral zone of periventricular hypothalamus (Hv). Labeled fibers lateral to the dorsal zone of the hypo- thalamus (Hd), dorsal to the nucleus ventromedialis of the thalamus (VM) and lateral to the nucleus tuberis anterior (TA). (B) Neurons in Hd dorsal to TA; filled processes projecting dorsolaterally to a collection of labeled fibers; positive fibers lateral to the labeled cells. (C) HRP-filled neurons in the penventricular portion of the nucleus centralis posterior (CP) of the thalamus; ventrolaterally directed fibers adjacent to Hd and dorsal to TA. (D) Filled cells in CP. Abbreviations: nucleus anterior of the thalamus (A), nucleus diffusus of the inferior lobe (DF); nucleus dorsalis posterior of the thalamus (DP), fasciculus retroflexus (FR), nucleus lateralis hypothalami (LH), optic tectum ( OTe), nucleus pretectalis centralis (PC), posterior commissure (PCo), nucleus preglomerulosus lateralis (PGI), nucleus preglomerulosus medialis (PGm), nucleus pretectalis periventricularis pars ventralis (PPv), nucleus pretectalis superficialis pars magnocellularis (PSm), nucleus pretectalis superficialis pars parvocellularis (PSp), parav- entricular organ (PVO), torus longitudinalis (TLo), nucleus tuberis posterior (TP), periventri- cular nucleus posterior tuberculum (TPp), granular layer of the valvula cerebelli (VCg), molecular layer of the valvula cerebelli (VCm), zona limitans diencephali (ZL).
428
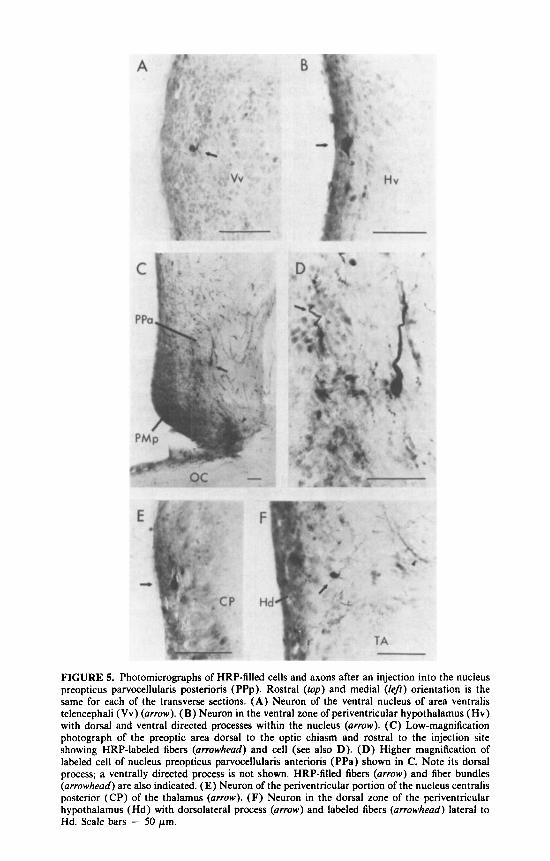
FIGURE 5. Photomicrographs of HRP-filled cells and axons after an injection into the nucleus preopticus parvocellularis posterioris (PPp). Rostra1 (top) and medial (left) orientation is the same for each of the transverse sections. ( A ) Neuron of the ventral nucleus of area ventralis telencephali (Vv) (arrow). (B) Neuron in the ventral zone of periventricular hypothalamus (Hv) with dorsal and ventral directed processes within the nucleus (arrow). (C) Low-magnification photograph of the preoptic area dorsal to the optic chiasm and rostra1 to the injection site showing HRP-labeled fibers (arrowhead) and cell (see also D). (D) Higher magnification of labeled cell of nucleus preopticus parvocellularis anterioris (PPa) shown in C. Note its dorsal process; a ventrally directed process is not shown. HRP-filled fibers (arrow) and fiber bundles (arrowhead) are also indicated. (E) Neuron of the periventricular portion of the nucleus centralis posterior (CP) of the thalamus (arrow). (F) Neuron in the dorsal zone of the periventricular hypothalamus (Hd) with dorsolateral process (arrow) and labeled fibers (arrowhead) lateral to Hd. Scale bars = 50 pm.

430 ANNALS NEW YORK ACADEMY OF SCIENCES
has a stimulatory effect upon SR (Experiment 2); and (3) the ventral telencephalic nuclei which receive TN inputs project to POA nuclei from which SR and sexual behavior can be evoked in goldfish (Experiment 3).
The labeling of the TN-retinal projection paired with evoked SR and courtship behavior is consistent with a TN relay of pheromonal information via fibers in the mMOT. A single intact mMOT, is sufficient for evoked SR and elicited courtship behavior. This confirms earlier studies indicating that a single intact mMOT (1 ) can mediate pheromone-induced courtship behavior similar to behavior exhibited by fish with one OT intact6 and (2) is the “necessary” component for SR evoked by OT stimulation.’ The HRP-labeled, TN-retinal projection verifies that a reasonable number of TN fibers are intact in preparations sufficient to sustain sexual behavior and SR.
The stimulatory effects of LHRH upon SR are supportive of TN mediation of reproductive behavior and are consistent with LHRH facilitation of reproductive behavior in tetrapods and other teleosts.“ The TN is an LHRH-containing system in many vertebrates.” In goldfish and other teleosts, LHRH fibers have been found in Vv and Vs of the ventral telencephalon and the PPa of the POA?’ areas that receive TN projections and areas known to concentrate sex-steroids and control repro- ductive behavior and SR? Presumably at least some of the peptide-containing axons in this area are of TN origin. The stimulatory effects of LHRH application to the cranial cavity and third ventricle may thus mimic a normal TN action upon these nuclei. Alternatively, olfactory input in the SR “area” could be mediated by LHRH released from ventral telencephalic, preoptic or other cells known to contain the peptide.20 Additional experiments are needed to further test these hypotheses.
The application of HRP to PPp of the POA demonstrates projections from cells of the ventral telencephalic zone that receive TN afferents (Vv-Vs complex) and contain steroid-concentrating neurons.” Labeling of Vv cells of the ventral telence- phalon is consistent with an earlier study in landlocked red salmon’‘ which reports Vv projections to PPa and the rostral nucleus preopticus of the POA. The VV-POA connection demonstrates a pathway that interconnects areas known to control sexual responses with afferents from the olfactory epithelium (TN and olfactory). This connection is undoubtedly only one of several in the “sex-control area” that most likely mediate sensorimotor integrations during courtship and spawning.
Similarities in the goldfish VV-POA connections to limbic systems in mammals controlling reproduction are striking.Z2 It has been proposed that Vv and Vs are homologous to the mammalian medial septum and the basal amygdala, respe~tively.”.~’ Another proposed homology suggests that the caudal Vv-rostra1 Vs complex is ho- mologous to the mammalian medial amygdala.‘ Both mammalian areas, septa1 and amygdala, have projections to the hypothalamus, portions of which are presumed to be homologous to the goldfish preoptic area.”
Some of the other areas (SC, nucleus suprachiasmaticus; Hv, ventral zone of the periventricular hypothalamus; CP, nucleus centralis posterior of the thalamus) that have inputs into the POA may also be part of systems involved in reproductive physiology and behavior. These preliminary results are discussed below with the intention of providing additional insight into the integrative capacity of the basal forebrain complex through which the TN may exert its control of sexual activity.
It is known that changes in day-length trigger gonadal growth and spawning in goldfish.24 The SC is involved in this type of seasonal breeding in other vertebrates2’ and thus may also serve a similar function in goldfish. Consistent with this idea are the observations that the SC in goldfish receives a retinal projection as it does in mammals.” A similar projection from the nucleus anterioris periventricularis (NAPv), which is, in part, equivalent to SC,” to the PPa and the rostral nucleus preopticus has been reported in the landlocked red salmon.’‘ The SC-POA connection may be the critical link between light-detection systems and pituitary gonadotropin secretion.

SLOAN & DEMSKI: TERMINAL NERVE IN GOLDFISH 431
The periventricular portion of the ventral hypothalamus (Hv) may be equivalent to part of the nucleus lateralis tuberis (NLT).l8 Lesions of NLT by monosodium L- glutamate produce short-term increased levels of gonadotropin,2" and NLT is believed to be involved in gonadal recrudescence in goldfish?' The Hv-to-PPp projection may thus influence reproductive development and behavior.
Thalamic cells (CP) were found to project to PPp. Recent studies in hermaphroditic sea bass have implicated the dorsal thalamus in the control of three distinct sex-color patterns and gamete r e l e a ~ e . ~ ~ . * ~ The thalamus is also known to concentrate sex-steroids and to contain LHRH cells and fibers in some fish and other vertebrates." Whereas color banding is not applicable to the goldfish, the thalamus (CP) may influence the preopticospinal pathway for SR. Studies on the role of the thalamus in goldfish sexual behavior are needed to substantiate the idea.
Overall, our results are consistent with the hypothesis that the T N influences sexual behavior and SR. However, overlapping of projections of the TN and the olfactory system still remains a problem in determination of the relative roles of the two pathways in chemosensory control of sexual behavior. The effect of LHRH upon SR strongly argues for the involvement of the TN, an LHRH-containing system. Specific forebrain nuclei which receive both T N and olfactory inputs are involved in reproductive behavior, as seen in and stimulation ~tudies.~'*~" Our current and previous anatomical work," along with other studies on these n ~ c l e i , ' ~ . ' ~ demonstrate anatomical pathways likely to mediate chemosensory effects on sexual behavior. Additional func- tional studies are needed to detail the mechanisms involved.
REFERENCES
1.
2.
3.
4.
5 .
6.
7.
8.
9.
10.
11.
12.
13.
DEMSKI, L. S. & R. G. NORTHCUTT. 1983. The terminal nerve: A new chemosensory system in vertebrates? Science 220 435-437.
STELL, W. K. & S. E. WALKER. 1987. Functional-anatomical studies on the terminal nerve projection to the retina of bony fishes. Ann. N.Y. Acad. Sci. This volume.
VON BARTHELD, C. S. 1987. Central connections of the terminal nerve in ray-finned fishes. Ann. N.Y. Acad. Sci. This volume.
DBVING, K. B. & R. SELSET. 1980. Behavior patterns in cod released by electrical stim- ulation of olfactory tract bundlets. Science 207: 559-560.
DEMSKI. L. S. & J. G. DULKA. 1984. Functional-anatomical studies on sperm release evoked by electrical stimulation of the olfactory tract in goldfish. Brain Res. 291: 241 -247.
STACEY, N. E. & A. L. KYLE. 1983. Effects of olfactory tract lesions on sexual and feeding behavior in the goldfish. Physiol. Behav. 3 0 621 -628.
KYLE, A. L. & R. E. PETER. 1982. Effects of forebrain lesions on spawning behavior in the male goldfish. Physiol. Behav. 28: 1103- 1109.
KOYAMA, Y., M. SATOU, Y. OKA & K. UEDA. 1984. Involvement of the telencephalic hemispheres and the preoptic area in sexual behavior of the male goldfish, Curussius oururus: A brain-lesion study. Behav. Neural Biol. 40: 70-86.
DEMSKI, L. S. & P. J. HORNBY. 1982. Hormonal control of fish reproductive behavior: Brain-gonadal steroid interactions. Can. J. Fish. Aquat. Sci. 39 36-47.
DEMSKI, L. S. 1983. Behavioral effects of electrical stimulation of the brain. In Fish Neurobiology. R. E. Davis & R. G. Northcutt. Eds. Vol 2: 317-359. The University of Michigan Press. Ann Arbor, MI.
DEMSKI, L. S. 1984. The evolution of neuroanatomical substrates of reproductive behavior: Sex steroid and LHRH-specific pathways including the terminal nerve. Amer. Zool. 24:
DEMSKI, L. S. & H. E. SLOAN. 1985. A direct magnocellular-preopticospinal pathway in
SHIGA, T., Y. OKA, M. OKUMOTO & K. UEDA. 1985. Efferents from the supracommissural
809 - 830.
goldfish: implications for control of sex behavior. Neurosci. Letts. 5 5 283-288.

432 ANNALS NEW YORK ACADEMY OF SCIENCES
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
ventral telencephalon in the hime salmon (landlocked red salmon, Oncorhynchus nerku): an anterograde degeneration study. Brain Res. Bull. 14 55-61.
SHIGA, T., Y. OKA, M. SATOU, N. OKUMOTO & K. UEDA. 1985. An HRP study of afferent connections of the supracommissural ventral telencephalon and the medial preoptic area in hime salmon (landlocked red salmon, Oncorhynchus nerku). Brain Res. 361: 162- 177.
SHIPLEY, M. T. 1982. A simple, low cost hydraulic pressure device for making microin- jections in the brain. Brain Res. Bull. 8 237-239.
COOK, J. E. & E. C. C. COOK. 1984. Use of a lectin-peroxidase conjugate (WGA-HRP) to assess the retinotopic precision of goldfish optic terminals. Neurosci. Lett. 48: 61 -66.
NORTHCUTT, R. G. & R. E. DAVIS. 1983. Telencephalic organization in ray-finned fishes. In Fish Neurobiology. R. E. Davis & R. G. Northcutt, Eds. Vol. 2 203-236. The University of Michigan Press. Ann Arbor, MI.
BRAFORD, M. R., JR., & R. G. NORTHCUTT. 1983. Organization of the diencephalon and pretectum of the ray-finned fishes. In Fish Neurobiology. R. E. Davis & R. G. Northcutt, Eds. Vol. 2: 117-163. The University of Michigan Press. Ann Arbor, MI.
FRYER, J. N. & L. MALER. 1981. Hypophysiotropic neurons in the goldfish hypothalamus demonstrated by retrograde transport of horseradish peroxidase. Cell Tiss. Res. 218:
KAH, O., B. BRETON, J. G. DULKA, J. NUNEZ-RODRIGUEZ, R. E. PETER, A. CORRICAN, J. E. RIVIER & W. W. VALE. 1986. A reinvestigation of the Gn-RH (gonadotrophin- releasing hormone) systems in the goldfish brain using antibodies to salmon Gn-RH. Cell Tiss. Res. 244: 327-337.
VON BARTHELD, C. S. & D. L. MEYER. 1986. Tracing of single fibers of the nervus terminalis in the goldfish brain. Cell Tiss. Res. 245: 143-158.
KOSTARCZYK, E. W. 1986. The amygdala and male reproductive functions: I. anatomical and endocrine bases. Neurosci. Biobehav. Rev. 1 0 67-77.
NORTHCUTT, R. G. & M. R. BRAFORD, JR. 1980. New observations on the organization and evolution of the telencephalon of actinopterygian fishes. In Comparative Neurology of the Telencephalon. S. 0. E. Ebbesson, Ed.: 41-98. Plenum. New York, NY.
FENWICK, J. C. 1970. The pineal organ: photoperiod and reproductive cycles in the goldfish, Curussius uurutus L. J. Endocrinol. 46: 101-111.
LEHMAN, M. N., E. L. BITTMAN & S. W. NEWMAN. 1984. Role of the hypothalamic paraventricular nucleus in neuroendocrine responses to daylength in the golden hamster. Brain Res. 308 25-32.
PETER, R. E., 0. KAH, C. R. PAULENCU, H. COOK & A. L. KYLE. 1980. Brain lesions and short-term endocrine effects of monosodium L-glutamate in goldfish, Curassius uu- rutus. Cell Tiss. Res. 212 429-442.
PETER, R. E. & J . N. FRYER. 1983. Endocrine functions of the hypothalamus of the actinopterygians. In Fish Neurobiology. R. E. Davis & R. G. Northcutt, Eds. Vol. 2: 165-201. The University of Michigan Press. Ann Arbor, MI.
DEMSKI, L. S. & J. G. DULKA. 1986. Thalamic stimulation evokes color change and gamete release in a vertebrate hermaphrodite. Experientia. 42: 1285- 1287.
DEMSKI, L. S. 1987. Diversity in reproductive patterns and behavior in teleost fishes. In Psychobiology of Reproductive Behavior. D. Crews, Ed.: 1-27. Prentice-Hall. Englewood Cliffs, NJ.
SATOU, M., Y. OKA, M. KUSUNOKI, T. MATSUSHIMA, M. KATO, I. FUJITA & K. UEDA. 1984. Telencephalic and preoptic areas integrate sexual behavior in hime salmon (land- locked red salmon, Oncorhynchus nerku): Results of electrical brain stimulation exper- iments. Physiol. Behav. 33: 441-447.
93- 102.




















