
The neurofilament infrastructure of a developing presynaptic calyx
Upload
independentCategory
view
3download
0
TOXICOLOGY AND APPLIED PHARMACOLOGY 142, 95–105 (1997)ARTICLE NO. TO968028
CS2-Mediated Cross-linking of Erythrocyte Spectrin andNeurofilament Protein: Dose Response and Temporal
Relationship to the Formation of Axonal Swellings1
WILLIAM M. VALENTINE,*,2 VENKATARAMAN AMARNATH,* DOYLE G. GRAHAM,*DANIEL L. MORGAN,† AND ROBERT C. SILLS
†
*Department of Pathology, Vanderbilt University Medical Center, Nashville, Tennessee 37232; and †Environmental Toxicology Program,NIEHS, Research Triangle Park, North Carolina 27709
Received May 24, 1996; accepted September 9, 1996
Carbon disulfide is a clean air act chemical with emissionsCS2-Mediated Cross-linking of Erythrocyte Spectrin and Neuro- in the U.S. of 50–60 ktons/year. Human exposure may occur
filament Protein: Dose Response and Temporal Relationship to through the use of CS2 in the production of rayon and cello-the Formation of Axonal Swellings. VALENTINE, W. M., AMAR-
phane or through release of CS2 from dithiocarbamates usedNATH, V., GRAHAM, D. G., MORGAN, D. L., AND SILLS, R. C.
as pesticides or therapeutic agents (Johnson et al., 1996).(1997). Toxicol. Appl. Pharmacol. 142, 95–105.The toxicity of CS2 was first recognized in humans duringthe mid-nineteenth century in Europe where CS2 was usedUsing model proteins, a mechanism for CS2-mediated covalent
cross-linking of proteins has been demonstrated previously. The in the cold vulcanization of rubber (Davidson and Feinlab,biologic importance of CS2-promoted protein cross-linking is ap- 1972) and remains a contemporary issue (Chu et al., 1995;parent as a possible dosimeter of CS2 exposure and as a potential Ruijten et al., 1993). Animal studies have demonstrated thatmechanism to account for the identical neuropathies produced subchronic or longer exposures to CS2 result in the develop-by 2,5-hexanedione and CS2. The present investigation examines ment of a distal axonopathy that is identical to the neuropa-the utility of erythrocyte spectrin cross-linking as a biomarker
thy produced by 2,5-hexanedione, the neurotoxic metaboliteof effect for inhalation exposure to CS2 and examines the abilityof hexane3 (Graham et al., 1995). Both toxicants requireof CS2 to cross-link neurofilament proteins, a potential neurotoxicfrequent and prolonged exposures in the occupational settingtarget. Rats were exposed to CS2 via inhalation at control, 50-,and demonstrate a predilection for long large-diameter my-500-, and 800-ppm levels for 2, 4, 8, and 13 weeks and spectrinelinated axons in the central and peripheral nervous systemsdimer formation was quantified using denaturing gel electropho-
resis and densitometry. Neurofilament preparations were also (Szendzikowski et al., 1973; Gottfried et al., 1985; Cavanaghobtained from spinal cords and examined for cross-linking using and Bennetts, 1981; Cavanagh, 1982). A prominent and in-Western blotting methods. The results obtained for protein cross- cipient structural change produced by these compounds atlinking were compared to morphologic changes in the cervical subterminal locations of the axon is the development of pre-and lumbar spinal cord using light and electron microscopy. The nodal swellings containing increased numbers of malalignedspectrin dimer exhibited a cumulative dose response and was neurofilaments (Gottfried et al., 1985; Colombi et al., 1981;detectable at both the 50-ppm level employed that did not pro-
Spencer and Schaumburg, 1977). Axonal atrophy occursduce axonal swellings and prior to the development of axonalproximal to the swellings (Pyle et al., 1992; Pappolla et al.,swellings for the 500- and 800-ppm levels used. Neurofilament1987) and although it has been proposed that atrophy canprotein cross-linking involved all three subunits and the temporalbe produced exclusive of axonal swellings using subacuterelationship of cross-linking was consistent with a contributingip administration of 2,5-hexanedione (Lehning et al., 1995),role in the development of axonal swellings. These results estab-
lish the sensitivity of spectrin cross-linking for evaluating inhala- other investigators using similar exposure paradigms havetion exposures and extend the similarities observed for 2,5-hex- observed swellings in both the central (Pyle et al., 1992)anedione and CS2 in both clinical settings and in vitro models to and the peripheral (Anthony et al., 1983b) nervous systems.their effects exerted on neurofilaments in the axon. q 1997 Academic Greater exposure to either compound results in larger diame-Press ter swellings accompanied by Schwann cell proliferation,
myelin retraction, and axonal degeneration distal to theswellings (Schaumburg and Spencer, 1976; Jirmanova andLukas, 1984; Gottfried et al., 1985). At the ultrastructural1 This work was presented in part at the 35th Annual Meeting of the
Society of Toxicology in March 1996, Anaheim, CA.3 IUPAC nomenclature (common name, n-hexane).2 To whom correspondence should be addressed.
95 0041-008X/97 $25.00Copyright q 1997 by Academic Press
All rights of reproduction in any form reserved.
AID TOX 8028 / 6h13$$$101 12-17-96 18:35:32 toxa AP: Tox

96 VALENTINE ET AL.
level, changes common to both 2,5-hexanedione and CS2 analogs of 2,5-hexanedione, another model that includes pro-tein cross-linking subsequent to pyrrole formation has beeninclude decreased numbers of axonal microtubules, seques-advanced. Methyl-substituted analogs of 2,5-hexanedionetration of axoplasmic organelles, and expansion of Schwanncapable of forming pyrroles and cross-linking proteins morecell cytoplasm (Gottfried et al., 1985; Colombi et al., 1981;rapidly than 2,5-hexanedione have demonstrated enhancedSpencer and Schaumburg, 1977). In addition to structuralneurotoxicity relative to 2,5-hexanedione (Anthony et al.,alterations, similar effects regarding axonal transport (Mo-1983a,b,c; Monaco et al., 1990). Similarly, rats exposed tonaco et al., 1989; Pappolla et al., 1987; Watson et al., 1991)2,5-hexanedione and hyperbaric oxygen, conditions ex-and clinical presentation (Herskowitz et al., 1971; Davidsonpected to result in more rapid protein cross-linking, demon-and Feinlab, 1972) have been reported. When consideredstrate onset of neuropathy at lower cumulative exposurescollectively, these shared characteristics define a neuro-than animals maintained under normobaric oxygen (Rosen-filamentous axonopathy that can be distinguished fromberg et al., 1987). Further supportive evidence that pyrroleother toxicant-induced neuropathies involving abnormalitiesformation is not adequate for the neurotoxicity of g-dike-in neurofilament structure, e.g., 3,3*-iminodipropionitrile,tones was presented using 3-acetyl-2,5-hexanedione. Al-which requires only acute to subacute exposures and pro-though this derivative of 2,5-hexanedione can rapidly formduces proximal axonal swellings without distal degenerationpyrroles on proteins, it does not effect covalent cross-linking,(Clark et al., 1980).axonal swellings, neurofilament aggregations, or axonal de-The identical effects exerted by CS2 and 2,5-hexanedionegeneration (Genter St Clair et al., 1988; Pyle et al., 1992).on the structure and function of the axon suggest a common
A series of reactions responsible for CS2-mediated cova-molecular mechanism of toxicity for these two toxicants.lent cross-linking of proteins has been established previouslyThe ability of CS2 to react with amino groups of proteins tousing model proteins (Valentine et al., 1992, 1995). Thisgenerate dithiocarbamates has provided a basis for severalability of CS2 to covalently cross-link proteins presents apossible mechanisms of CS2 toxicity (Beauchamp et al.,novel mechanism having the potential to account for the1983). In one, chelation of metal cofactors by protein-associ-identical structural and functional changes incurred in theated dithiocarbamates was proposed, and another involvedaxon from CS2 and 2,5-hexanedione. Additionally, due tovitamin B6 deficiency resulting from derivatization of pyri-the stability of the thiourea structures produced by CS2,doxamine. Interestingly, both metal chelation and vitaminanalysis of cross-linking in a relatively easily obtained pro-deficiency have also been discussed in the context of g-tein appears promising as a mechanistically relevant dosime-diketone neurotoxicity (Spencer et al., 1980) but little sup-ter for exposures to CS2. In particular, previous investiga-port for these mechanisms regarding either CS2 or 2,5-hex-tions have suggested that erythrocyte spectrin may be ananedione has ensued. Due to the magnitude and early occur-appropriate protein for monitoring CS2-induced proteinrence of changes in neurofilament structure, neurofilamentcross-linking (Valentine et al., 1993). Those studies demon-proteins have drawn attention as potential neurotoxic targetsstrated distinct protein species in erythrocyte membranesof CS2 and 2,5-hexanedione. Transformation of neurofila-after CS2 exposure that were composed of covalently cross-ment triplet protein amino groups to 2,5-dimethylpyrroleslinked heterodimers of the a and b subunits of spectrin. Theor dithiocarbamates by either 2,5-hexanedione or CS2, re-investigations presented here evaluate the utility of erythro-spectively, has been presented as the event responsible forcyte spectrin cross-linking for detecting inhalation exposuresproducing axonal swellings (DeCaprio and O’Neill, 1985;to CS2 and examine the ability of CS2 to covalently cross-Sayre et al., 1985; Zhu et al., 1994). According to this model,link neurofilament proteins. Sodium dodecyl sulfate–poly-changes in solubility and/or net charge of neurofilament pro-acrylamide gel electrophoresis (SDS–PAGE)4 and densi-teins are thought to alter either the interactions among neuro-tometry were used to isolate and quantify spectrin dimerfilaments or the interaction of neurofilaments with other axo-formation in erythrocytes obtained from rats exposed to CS2plasmic components, leading to accelerated transport andby inhalation as a function of exposure level and duration.
accumulation of neurofilaments in the distal portion of ax-The quantity of spectrin cross-linking was compared to light-
ons. Contradictory to this hypothesis are the findings thatmicroscopic and ultrastructural changes in the spinal cord
accelerated transport continues even though exposure to 2,5- to evaluate the sensitivity of this biomarker for subneuro-hexanedione and neurofilament derivatization have ceased toxic, i.e., those that do not produce axonal swellings, expo-(Pyle et al., 1993) and that administration of 3-acetyl-2,5-hexanedione, which will adduct but not cross-link proteins,
4 Abbreviations used: SDS–PAGE, sodium dodecyl sulfate–polyacryl-is not accompanied by accelerated neurofilament transportamide gel electrophoresis; PVDF, polyvinylidene difluoride; NFL, low-(Pyle et al., 1992). These data appear more consistent withmolecular-weight neurofilament protein; NFM, medium-molecular-weightan interpretation that accelerated transport is a nonspecific neurofilament protein; NFH, high-molecular-weight neurofilament protein;
reparative or regenerative response to toxicant-induced in- TBST, 20 mM Tris, 0.5 M NaCl, pH 7.5, 0.05% Tween-20; TTCA, 2-thiothiazolidine-4-carboxylic acid.jury (McKerracher et al., 1990; Pyle et al., 1993). Utilizing
AID TOX 8028 / 6h13$$$101 12-17-96 18:35:32 toxa AP: Tox

97CS2 CROSS-LINKING OF SPECTRIN AND NEUROFILAMENTS
lowing which a complete necropsy was performed. Sections of cervical andsures of CS2 and neurotoxic exposures of CS2 prior to mor-lumbar spinal cord were postfixed in 2% osmium tetroxide, dehydrated inphological changes in the axon. Neurofilament preparationsgraded alcohols and propylene oxide, and embedded in epoxy resin. For
obtained from CS2-exposed rats were examined for cross- light microscopy, 1-mm-thick sections were stained with toluidine blue. Forlinking using Western blotting techniques, and the ability of electron microscopy, 60-nm ultra-thin sections were stained with lead citrate
and uranyl acetate and viewed on a Zeiss 10-A transmission electron micro-CS2 to covalently cross-link the individual triplet proteins ofscope. Four males and four females were examined at each exposure level,the neurofilament assessed. Additionally, to address whetherincluding controls, at the 2-, 4-, and 8-week time points and eight animalsneurofilament cross-linking is a contributing mechanism inof each sex were used for the 13-week exposure groups.
the formation of neurofilamentous axonal swellings or a pro-Isolation of erythrocyte spectrin. Spectrin was purified as described
cess that occurs secondary to axonal swelling, neurofilament previously (Genter St Clair et al., 1988) from erythrocytes of 5 randomlycross-linking was quantified as a function of exposure and selected male rats taken from each exposure group except for the 13-week
groups for which 10 male rats were used. Within 24 hr of the last exposurethe temporal relationship of neurofilament protein cross-link-rats were exsanguinated from the abdominal aorta under deep chloral hy-ing to the development of axonal swellings determined.drate anesthesia. Erythrocytes were washed four times with 310 mOsmTris–HCl, pH 7.6, and lysed by addition of 6 volumes of 20 mOsm Tris–
MATERIALS AND METHODS HCl, pH 7.6, to red cells at a hematocrit of 50. Ghosts were separated fromhemolysate by centrifugation (20,000g) at 47C for 40 min and washed three
Chemicals. Polyvinylidene difluoride membranes (PVDF) and chemi- times with 20 mOsm Tris–HCl, pH 7.6. Spectrin was eluted from red cellluminescence reagents were obtained from NEN (Boston, MA). Goat horse- ghosts in 0.1 mM EDTA, pH 8.0, at 377C and collected in the supernatantradish peroxidase-conjugated anti-rabbit and rabbit polyclonal anti-spectrin following centrifugation at 130,000g at 47C for 30 min.antibodies were obtained from Sigma Chemicals (St. Louis, MO). The Neurofilament isolation. Neurofilament proteins were isolated from ratmouse monoclonal antibodies anti-NFL and anti-NFH were obtained from spinal cord according to a published method (Liem and Hutchison, 1982).Sigma Chemicals (St. Louis, MO) and the mouse monoclonal anti-NFM was Three male rats were examined at each exposure level and duration. Imme-obtained from Boehringer Mannheim (Indianapolis, IN). Goat horseradish diately following exsanguination the spinal cords were blown from theperoxidase-conjugated anti-mouse IgG was obtained from Bio-Rad (Mel- spinal canal using cold saline. Following removal of the meninges the spinalville, NY). Rabbit polyclonal anti-ubiquitin was purchased from Dako Cor- cords were homogenized in a solution containing 10 mM phosphate buffer,poration (Carpinteria, CA). pH 6.8, 1 mM EDTA, 0.1 mM NaCl, 1 mM PMSF, 1 mg/ml leupeptin, 1%
Animals. This study was in accordance with the NIH Guide for Care Triton-X, and 0.85 M sucrose and stirred overnight at 47C in the homogeniz-and Use of Laboratory Animals and was approved by the animal care ing solution. A pellet containing neurofilament protein was obtained bycommittee. Male and female Fischer 344 rats, 6–7 weeks old (Charles centrifugation (90,000g for 30 min) which was washed with the same solu-River Breeding Laboratories, Raleigh, NC), were acclimated for 2 weeks tion minus sucrose and Triton-X. The neurofilament preparations were thenprior to the start of the CS2 exposures. During acclimation, rats were individ- solubilized for analysis by combining with an equal volume of sampleually identified by microchip implantation and randomized by weight to buffer containing 8 M urea, 1% SDS, 2% dithiothreitol, 2% glycerol, andtreatment groups. The exposure levels and durations examined were control, 0.5 M Tris–HCl, pH 6.8.50 ppm, 500 ppm, 800 ppm, and 2, 4, 8, and 13 weeks, respectively. SDS–PAGE. Covalent cross-linking of erythrocyte spectrin was evalu-Animals were selected randomly for analyses as follows: spectrin cross- ated by electrophoresis on SDS-containing 5% polyacrylamide gels andlinking, 5 males per exposure level for the 2-, 4-, and 8-week durations visualized with silver stain. Spectrin preparations (Ç10 mg total protein)and 10 males for the 13-week exposure groups; neurofilament cross-linking, were boiled for 5 min in sample buffer containing 1% SDS and 2% dithio-3 males were examined for each exposure level and period; morphologic threitol before electrophoresis. Monomer and dimer spectrin were quantifiedevaluations, 4 males and 4 females were examined at each exposure level by densitometry using an ISCO gel scanner, Model 1312, in combinationat the 2-, 4-, and 8-week time points and 8 animals of each sex examined with an ISCO UA-5 absorbance/fluorescence detector and analyzed usingfor the 13-week exposure groups. Animals were individually housed in HP Chem Station software. Data were collected as area under the curveHazleton 1000 inhalation exposure chambers for the duration of the study. for monomer and dimer spectrin. Statistical significance was determinedChlorinated, filtered tap water and NIH 07 diet were provided ad libitum; using analysis of variance for differences within exposure periods, p õhowever, food was removed during the 6-hr exposures and for 6 hr per day 0.01, and comparisons of individual treatment groups were performed usingon nonexposure days. Animals were observed twice a day for clinical signs Duncan’s multiple range test, p õ 0.01. Dimer as percentage of dimer plusof toxicity. monomer is presented as means { SE with n Å 5 (13-week samples,
Inhalation exposure. Carbon disulfide, purity ú99% (Aldrich Chemi- n Å 10).cal Co., Milwaukee, WI), was vaporized, mixed with conditioned air Immunoblotting of spectrin and neurofilament preparations. Erythro-(HEPA-filtered, charcoal-scrubbed, temperature and humidity controlled), cyte membrane proteins were separated using 5% homogeneous polyacryl-and delivered to the inhalation chambers at nominal concentrations of 0, amide gels containing SDS. Transfer to PVDF membranes was performed50, 500, or 800 ppm. Rats were exposed to either conditioned air (controls) using the wet transfer method at 60 V for 1 hr. Immunoblotting of mem-or CS2 for 6 hr per day, 5 days a week, for 2, 4, 8, or 13 weeks (weekdays branes was initiated by briefly washing the membrane in Tris-bufferedonly). Exposure chamber concentrations were continuously monitored by saline (20 mM Tris, 0.5 M NaCl, pH 7.5) with 0.05% Tween-20 (TBST).Fourier transform infrared spectrophotometers. The mean daily exposure The membranes were blocked with 3% bovine serum albumin or 3% nonfatconcentrations were within 3% of the target levels: 49 { 0.42, 512.6 { dry milk in TBST for 1 hr at room temperature. After incubating with1.14, and 814.0 { 1.73 ppm (means { SE). The mean daily temperature primary antibody (rabbit polyclonal anti-human erythrocyte spectrin at aand percentage relative humidity were not significantly different between dilution of 1:400) for 1 hr at room temperature the membranes were washedthe four chambers. The mean chamber temperature was 227C, and the mean three times for 5 min in TBST, incubated with goat anti-rabbit horseradishpercentage relative humidity was 49.1%. peroxidase-conjugated antibody (1:2000 dilution) in TBST at room temper-
ature, washed three times for 5 min in TBST, and visualized using chemilu-Morphology. Rats were whole-body perfused with phosphate-buffered4% paraformaldehyde followed by phosphate-buffered 4% glutaraldehyde, minescence.
Rat neurofilament preparations were separated using 7% homogeneouspH 7.4, at room temperature. Rats were refrigerated at 47C overnight, fol-
AID TOX 8028 / 6h13$$$102 12-17-96 18:35:32 toxa AP: Tox
98 VALENTINE ET AL.
jor bands occurring within the 200- to 250-kDa range wereobserved for all animals. In addition, several protein bandswere detected in the 410-kDa region of rats exposed to CS2
as reported previously for rats administered CS2 ip (Valen-tine et al., 1993).
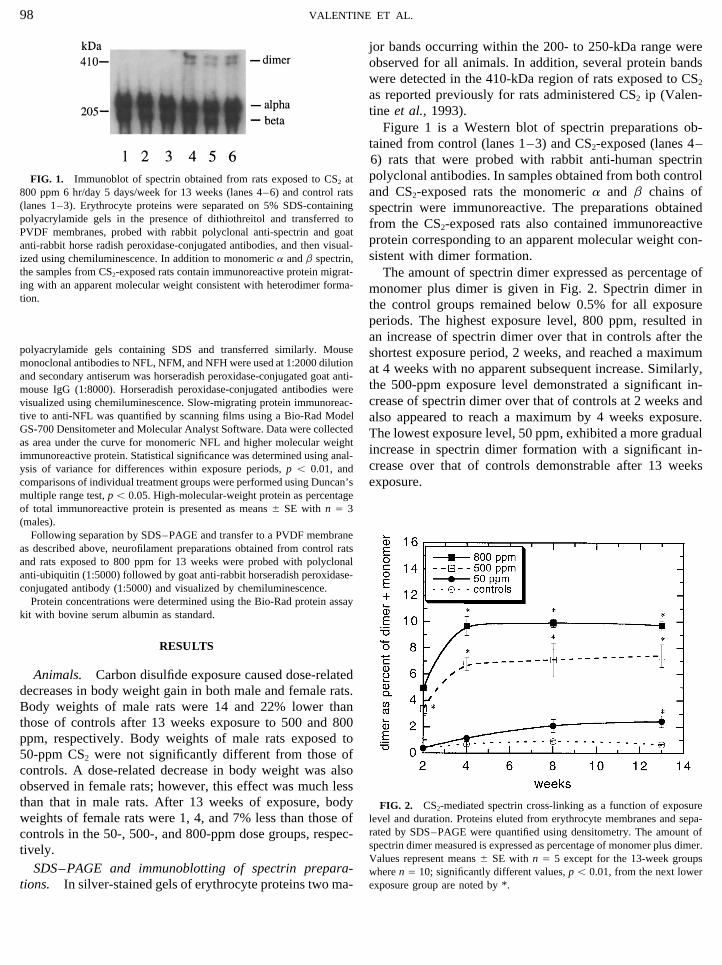
Figure 1 is a Western blot of spectrin preparations ob-tained from control (lanes 1–3) and CS2-exposed (lanes 4–6) rats that were probed with rabbit anti-human spectrinpolyclonal antibodies. In samples obtained from both controlFIG. 1. Immunoblot of spectrin obtained from rats exposed to CS2 at
800 ppm 6 hr/day 5 days/week for 13 weeks (lanes 4–6) and control rats and CS2-exposed rats the monomeric a and b chains of(lanes 1–3). Erythrocyte proteins were separated on 5% SDS-containing spectrin were immunoreactive. The preparations obtainedpolyacrylamide gels in the presence of dithiothreitol and transferred to from the CS2-exposed rats also contained immunoreactivePVDF membranes, probed with rabbit polyclonal anti-spectrin and goat
protein corresponding to an apparent molecular weight con-anti-rabbit horse radish peroxidase-conjugated antibodies, and then visual-sistent with dimer formation.ized using chemiluminescence. In addition to monomeric a and b spectrin,
the samples from CS2-exposed rats contain immunoreactive protein migrat- The amount of spectrin dimer expressed as percentage ofing with an apparent molecular weight consistent with heterodimer forma- monomer plus dimer is given in Fig. 2. Spectrin dimer intion. the control groups remained below 0.5% for all exposure
periods. The highest exposure level, 800 ppm, resulted inan increase of spectrin dimer over that in controls after the
polyacrylamide gels containing SDS and transferred similarly. Mouse shortest exposure period, 2 weeks, and reached a maximummonoclonal antibodies to NFL, NFM, and NFH were used at 1:2000 dilution at 4 weeks with no apparent subsequent increase. Similarly,and secondary antiserum was horseradish peroxidase-conjugated goat anti-
the 500-ppm exposure level demonstrated a significant in-mouse IgG (1:8000). Horseradish peroxidase-conjugated antibodies werecrease of spectrin dimer over that of controls at 2 weeks andvisualized using chemiluminescence. Slow-migrating protein immunoreac-
tive to anti-NFL was quantified by scanning films using a Bio-Rad Model also appeared to reach a maximum by 4 weeks exposure.GS-700 Densitometer and Molecular Analyst Software. Data were collected The lowest exposure level, 50 ppm, exhibited a more gradualas area under the curve for monomeric NFL and higher molecular weight
increase in spectrin dimer formation with a significant in-immunoreactive protein. Statistical significance was determined using anal-crease over that of controls demonstrable after 13 weeksysis of variance for differences within exposure periods, p õ 0.01, and
comparisons of individual treatment groups were performed using Duncan’s exposure.multiple range test, p õ 0.05. High-molecular-weight protein as percentageof total immunoreactive protein is presented as means { SE with n Å 3(males).
Following separation by SDS–PAGE and transfer to a PVDF membraneas described above, neurofilament preparations obtained from control ratsand rats exposed to 800 ppm for 13 weeks were probed with polyclonalanti-ubiquitin (1:5000) followed by goat anti-rabbit horseradish peroxidase-conjugated antibody (1:5000) and visualized by chemiluminescence.
Protein concentrations were determined using the Bio-Rad protein assaykit with bovine serum albumin as standard.
RESULTS
Animals. Carbon disulfide exposure caused dose-relateddecreases in body weight gain in both male and female rats.Body weights of male rats were 14 and 22% lower thanthose of controls after 13 weeks exposure to 500 and 800ppm, respectively. Body weights of male rats exposed to50-ppm CS2 were not significantly different from those ofcontrols. A dose-related decrease in body weight was alsoobserved in female rats; however, this effect was much lessthan that in male rats. After 13 weeks of exposure, body FIG. 2. CS2-mediated spectrin cross-linking as a function of exposure
level and duration. Proteins eluted from erythrocyte membranes and sepa-weights of female rats were 1, 4, and 7% less than those ofrated by SDS–PAGE were quantified using densitometry. The amount ofcontrols in the 50-, 500-, and 800-ppm dose groups, respec-spectrin dimer measured is expressed as percentage of monomer plus dimer.tively.Values represent means { SE with n Å 5 except for the 13-week groups
SDS–PAGE and immunoblotting of spectrin prepara- where n Å 10; significantly different values, p õ 0.01, from the next lowerexposure group are noted by *.tions. In silver-stained gels of erythrocyte proteins two ma-
AID TOX 8028 / 6h13$$$102 12-17-96 18:35:32 toxa AP: Tox

99CS2 CROSS-LINKING OF SPECTRIN AND NEUROFILAMENTS
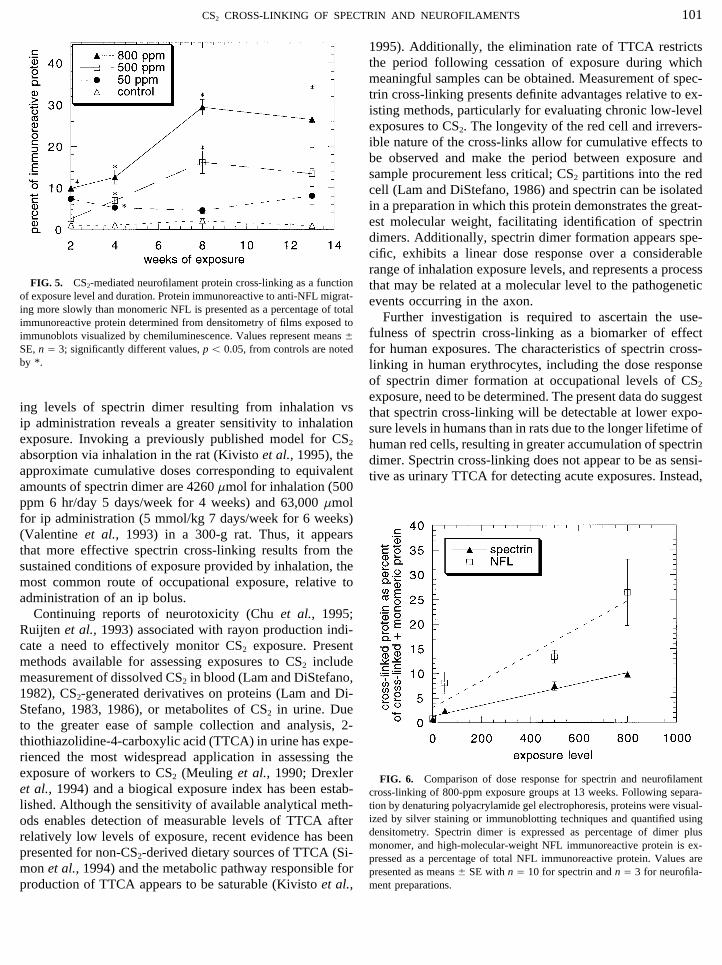
immunoreactive protein is presented in Fig. 5. The earliestsignificant increase over controls was observed at 2 weeksin the 800-ppm group. At 4 weeks significant increases overcontrols were present in all treatment groups. Backgroundlevels in controls remained below 2% for all exposureperiods.
Neither neurofilament preparations obtained from controlrats nor rats exposed to CS2 at 800 ppm for 13 weeks exhib-ited immunoreactivity when probed with polyclonal anti-ubiquitin using Western blot analysis.
Dose response of spectrin and NFL cross-linking. Theamount of spectrin dimer and the quantity of high-molecular-
FIG. 3. Immunoblots of neurofilament proteins obtained from spinal weight protein demonstrating NFL immunoreactivity for thecords of controls (lanes 1 and 2) and rats exposed to CS2 at 800 ppm 6 hr/ 13-week exposure groups were measured. In Fig. 6 spectrinday, 5 days/week for 8 weeks (lanes 3 and 4). Proteins were separated on dimer is expressed as percentage of dimer plus monomer7% homogeneous polyacrylamide gels in the presence of SDS, 8 M urea
and high-molecular-weight NFL immunoreactive protein isand dithiothreitol, transferred to PVDF, and probed with (A) mouse mono-expressed as percentage of total NFL immunoreactive pro-clonal anti-NFL primary antisera and horseradish peroxidase-conjugated
goat anti-mouse IgG secondary antisera, (B) mouse monoclonal anti-NFM tein. Values are presented as means { SE (n Å 10 for spec-primary antisera and horseradish peroxidase-conjugated goat anti-mouse trin and n Å 3 for neurofilament preparations). The slopesIgG secondary antisera, and (C) mouse monoclonal anti-NFH primary anti- for spectrin cross-linking and neurofilament cross-linkingsera and horseradish peroxidase-conjugated goat anti-mouse IgG secondary
were 0.011 { 0.001 (R2 Å 0.97) and 0.027 { 0.006 (R2 Åantisera. Proteins were visualized by chemiluminescence. The farthest mi-0.86), respectively.grating bands in A, B, and C (lanes 1 and 2) represent monomeric NFL,
NFM, and NFH, respectively. In addition to monomeric neurofilament sub- Morphology. The incidence and severity of spinal cordunits, the preparations from CS2-exposed rats also contained slower migrat- axonal swellings are presented in Table 1. No axonal swell-ing protein possessing immunoreactivity to all three neurofilament anti-
ings were observed in spinal cords of controls or rats exposedbodies.to 50-ppm CS2 at any time point examined. Similarly, swell-ings were not detected in sections obtained from animalsexposed to 500- or 800-ppm levels at 2 and 4 weeks expo-
Immunoblotting of neurofilament preparations. Figure sure. At the 500-ppm exposure level multifocal mild axonal3A is a Western blot of the neurofilament preparations ob- swellings were detected at 8 weeks exposure in the cervicaltained from control (lanes 1 and 2) and CS2-exposed rats and lumbar regions in 75 and 50% of the rats, respectively.(lanes 3 and 4) probed with anti-NFL. The lanes correspond- Thirteen weeks exposure at 500 ppm resulted in diffuse gianting to the proteins isolated from exposed rats contain immu- axonal swellings in both regions of the spinal cord in allnoreactive protein migrating more slowly than the NFL rats examined. Spinal cord sections obtained from all ratsmonomer. Higher apparent molecular weight protein in the exposed to 800-ppm CS2 demonstrated swellings in the cer-samples obtained from treated rats (lanes 3 and 4) was also vical and lumbar regions that increased in both distributionimmunoreactive for NFM and NFH antibodies as shown in and size from 8 to 13 weeks exposure.Figs. 3B and 3C, respectively. No bands in addition to the In the cervical spinal cord, axonal swelling was observedNFL, NFM, and NFH monomers were detected in controls primarily in the fasciculus gracilis nerve tracts and was char-(lanes 1 and 2). acterized by round, variable-size clear spaces surrounded by
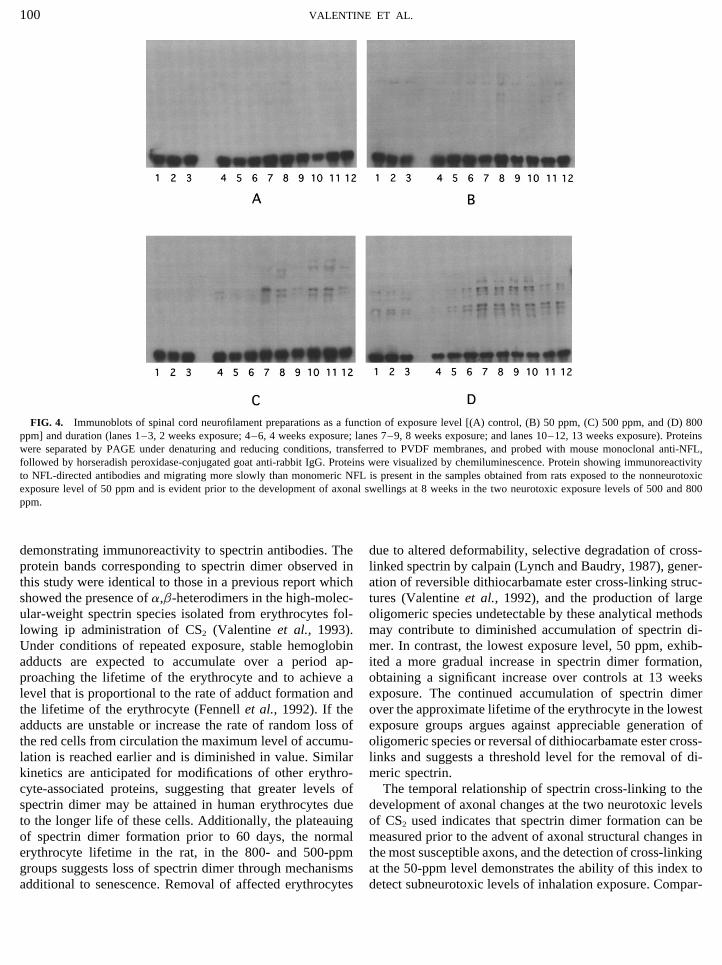
Figure 4 presents Western blots of neurofilament prepara- reduced myelin which were multifocal at 8 weeks and muchtions probed with anti-NFL according to level and duration more diffuse by 13 weeks (Fig. 7B). Axonal structuralof exposure to CS2. Figure 4A presents preparations obtained changes were more prominent in the lumbar spinal cord.from controls which do not display immunoreactive protein Focal axonal swelling was first detected in the ventral funicu-migrating slower than NFL. Figure 4B demonstrates the lus nerve tracts and progressed in severity and distributionpresence of immunoreactive protein in the high-molecular- with longer exposure (Fig. 7D). Electron-microscopic exam-weight region in animals exposed at the subneurotoxic, i.e., ination of swollen axons in the ventral funiculus nerve tracksno production of axonal swellings, level of 50 ppm. Both revealed fewer detectable microtubules per field and an in-Fig. 4C and Fig. 4D illustrate that immunoreactive high- crease in the density of neurofilaments with an accompa-molecular-weight protein can be detected prior to the devel- nying loss of their normal longitudinal orientation (Fig. 7F).opment of neurofilamentous swellings, which occurred at 8
DISCUSSIONweeks, for the two neurotoxic doses, 500 and 800 ppm. Theamount of immunoreactive protein migrating more slowly Exposure to CS2 by inhalation resulted in the formation of
high-molecular-weight erythrocyte membrane protein bandsthan monomeric NFL expressed as a percentage of total
AID TOX 8028 / 6h13$$$103 12-17-96 18:35:32 toxa AP: Tox
100 VALENTINE ET AL.
FIG. 4. Immunoblots of spinal cord neurofilament preparations as a function of exposure level [(A) control, (B) 50 ppm, (C) 500 ppm, and (D) 800ppm] and duration (lanes 1–3, 2 weeks exposure; 4–6, 4 weeks exposure; lanes 7–9, 8 weeks exposure; and lanes 10–12, 13 weeks exposure). Proteinswere separated by PAGE under denaturing and reducing conditions, transferred to PVDF membranes, and probed with mouse monoclonal anti-NFL,followed by horseradish peroxidase-conjugated goat anti-rabbit IgG. Proteins were visualized by chemiluminescence. Protein showing immunoreactivityto NFL-directed antibodies and migrating more slowly than monomeric NFL is present in the samples obtained from rats exposed to the nonneurotoxicexposure level of 50 ppm and is evident prior to the development of axonal swellings at 8 weeks in the two neurotoxic exposure levels of 500 and 800ppm.
demonstrating immunoreactivity to spectrin antibodies. The due to altered deformability, selective degradation of cross-linked spectrin by calpain (Lynch and Baudry, 1987), gener-protein bands corresponding to spectrin dimer observed in
this study were identical to those in a previous report which ation of reversible dithiocarbamate ester cross-linking struc-tures (Valentine et al., 1992), and the production of largeshowed the presence of a,b-heterodimers in the high-molec-
ular-weight spectrin species isolated from erythrocytes fol- oligomeric species undetectable by these analytical methodsmay contribute to diminished accumulation of spectrin di-lowing ip administration of CS2 (Valentine et al., 1993).
Under conditions of repeated exposure, stable hemoglobin mer. In contrast, the lowest exposure level, 50 ppm, exhib-ited a more gradual increase in spectrin dimer formation,adducts are expected to accumulate over a period ap-
proaching the lifetime of the erythrocyte and to achieve a obtaining a significant increase over controls at 13 weeksexposure. The continued accumulation of spectrin dimerlevel that is proportional to the rate of adduct formation and
the lifetime of the erythrocyte (Fennell et al., 1992). If the over the approximate lifetime of the erythrocyte in the lowestexposure groups argues against appreciable generation ofadducts are unstable or increase the rate of random loss of
the red cells from circulation the maximum level of accumu- oligomeric species or reversal of dithiocarbamate ester cross-links and suggests a threshold level for the removal of di-lation is reached earlier and is diminished in value. Similar
kinetics are anticipated for modifications of other erythro- meric spectrin.The temporal relationship of spectrin cross-linking to thecyte-associated proteins, suggesting that greater levels of
spectrin dimer may be attained in human erythrocytes due development of axonal changes at the two neurotoxic levelsof CS2 used indicates that spectrin dimer formation can beto the longer life of these cells. Additionally, the plateauing
of spectrin dimer formation prior to 60 days, the normal measured prior to the advent of axonal structural changes inthe most susceptible axons, and the detection of cross-linkingerythrocyte lifetime in the rat, in the 800- and 500-ppm
groups suggests loss of spectrin dimer through mechanisms at the 50-ppm level demonstrates the ability of this index todetect subneurotoxic levels of inhalation exposure. Compar-additional to senescence. Removal of affected erythrocytes
AID TOX 8028 / 6h13$$$103 12-17-96 18:35:32 toxa AP: Tox
101CS2 CROSS-LINKING OF SPECTRIN AND NEUROFILAMENTS
1995). Additionally, the elimination rate of TTCA restrictsthe period following cessation of exposure during whichmeaningful samples can be obtained. Measurement of spec-trin cross-linking presents definite advantages relative to ex-isting methods, particularly for evaluating chronic low-levelexposures to CS2. The longevity of the red cell and irrevers-ible nature of the cross-links allow for cumulative effects tobe observed and make the period between exposure andsample procurement less critical; CS2 partitions into the redcell (Lam and DiStefano, 1986) and spectrin can be isolatedin a preparation in which this protein demonstrates the great-est molecular weight, facilitating identification of spectrindimers. Additionally, spectrin dimer formation appears spe-cific, exhibits a linear dose response over a considerablerange of inhalation exposure levels, and represents a process
FIG. 5. CS2-mediated neurofilament protein cross-linking as a function that may be related at a molecular level to the pathogeneticof exposure level and duration. Protein immunoreactive to anti-NFL migrat- events occurring in the axon.ing more slowly than monomeric NFL is presented as a percentage of total
Further investigation is required to ascertain the use-immunoreactive protein determined from densitometry of films exposed tofulness of spectrin cross-linking as a biomarker of effectimmunoblots visualized by chemiluminescence. Values represent means {
SE, n Å 3; significantly different values, p õ 0.05, from controls are noted for human exposures. The characteristics of spectrin cross-by *. linking in human erythrocytes, including the dose response
of spectrin dimer formation at occupational levels of CS2
exposure, need to be determined. The present data do suggesting levels of spectrin dimer resulting from inhalation vs that spectrin cross-linking will be detectable at lower expo-ip administration reveals a greater sensitivity to inhalation sure levels in humans than in rats due to the longer lifetime ofexposure. Invoking a previously published model for CS2 human red cells, resulting in greater accumulation of spectrinabsorption via inhalation in the rat (Kivisto et al., 1995), the dimer. Spectrin cross-linking does not appear to be as sensi-approximate cumulative doses corresponding to equivalent tive as urinary TTCA for detecting acute exposures. Instead,amounts of spectrin dimer are 4260 mmol for inhalation (500ppm 6 hr/day 5 days/week for 4 weeks) and 63,000 mmolfor ip administration (5 mmol/kg 7 days/week for 6 weeks)(Valentine et al., 1993) in a 300-g rat. Thus, it appearsthat more effective spectrin cross-linking results from thesustained conditions of exposure provided by inhalation, themost common route of occupational exposure, relative toadministration of an ip bolus.
Continuing reports of neurotoxicity (Chu et al., 1995;Ruijten et al., 1993) associated with rayon production indi-cate a need to effectively monitor CS2 exposure. Presentmethods available for assessing exposures to CS2 includemeasurement of dissolved CS2 in blood (Lam and DiStefano,1982), CS2-generated derivatives on proteins (Lam and Di-Stefano, 1983, 1986), or metabolites of CS2 in urine. Dueto the greater ease of sample collection and analysis, 2-thiothiazolidine-4-carboxylic acid (TTCA) in urine has expe-rienced the most widespread application in assessing theexposure of workers to CS2 (Meuling et al., 1990; Drexler FIG. 6. Comparison of dose response for spectrin and neurofilamentet al., 1994) and a biogical exposure index has been estab- cross-linking of 800-ppm exposure groups at 13 weeks. Following separa-
tion by denaturing polyacrylamide gel electrophoresis, proteins were visual-lished. Although the sensitivity of available analytical meth-ized by silver staining or immunoblotting techniques and quantified usingods enables detection of measurable levels of TTCA afterdensitometry. Spectrin dimer is expressed as percentage of dimer plusrelatively low levels of exposure, recent evidence has beenmonomer, and high-molecular-weight NFL immunoreactive protein is ex-
presented for non-CS2-derived dietary sources of TTCA (Si- pressed as a percentage of total NFL immunoreactive protein. Values aremon et al., 1994) and the metabolic pathway responsible for presented as means { SE with n Å 10 for spectrin and n Å 3 for neurofila-
ment preparations.production of TTCA appears to be saturable (Kivisto et al.,
AID TOX 8028 / 6h13$$$103 12-17-96 18:35:32 toxa AP: Tox
102 VALENTINE ET AL.
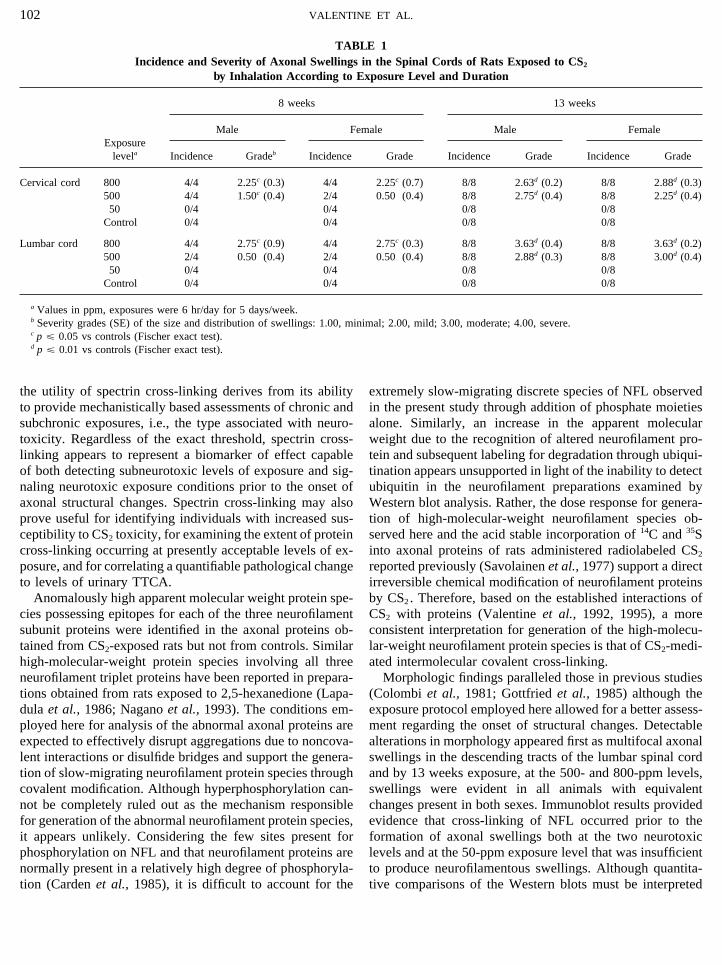
TABLE 1Incidence and Severity of Axonal Swellings in the Spinal Cords of Rats Exposed to CS2
by Inhalation According to Exposure Level and Duration
8 weeks 13 weeks
Male Female Male FemaleExposure
levela Incidence Gradeb Incidence Grade Incidence Grade Incidence Grade
Cervical cord 800 4/4 2.25c (0.3) 4/4 2.25c (0.7) 8/8 2.63d (0.2) 8/8 2.88d (0.3)500 4/4 1.50c (0.4) 2/4 0.50 (0.4) 8/8 2.75d (0.4) 8/8 2.25d (0.4)50 0/4 0/4 0/8 0/8
Control 0/4 0/4 0/8 0/8
Lumbar cord 800 4/4 2.75c (0.9) 4/4 2.75c (0.3) 8/8 3.63d (0.4) 8/8 3.63d (0.2)500 2/4 0.50 (0.4) 2/4 0.50 (0.4) 8/8 2.88d (0.3) 8/8 3.00d (0.4)50 0/4 0/4 0/8 0/8
Control 0/4 0/4 0/8 0/8
a Values in ppm, exposures were 6 hr/day for 5 days/week.b Severity grades (SE) of the size and distribution of swellings: 1.00, minimal; 2.00, mild; 3.00, moderate; 4.00, severe.c p £ 0.05 vs controls (Fischer exact test).d p £ 0.01 vs controls (Fischer exact test).
the utility of spectrin cross-linking derives from its ability extremely slow-migrating discrete species of NFL observedin the present study through addition of phosphate moietiesto provide mechanistically based assessments of chronic and
subchronic exposures, i.e., the type associated with neuro- alone. Similarly, an increase in the apparent molecularweight due to the recognition of altered neurofilament pro-toxicity. Regardless of the exact threshold, spectrin cross-
linking appears to represent a biomarker of effect capable tein and subsequent labeling for degradation through ubiqui-tination appears unsupported in light of the inability to detectof both detecting subneurotoxic levels of exposure and sig-
naling neurotoxic exposure conditions prior to the onset of ubiquitin in the neurofilament preparations examined byWestern blot analysis. Rather, the dose response for genera-axonal structural changes. Spectrin cross-linking may also
prove useful for identifying individuals with increased sus- tion of high-molecular-weight neurofilament species ob-served here and the acid stable incorporation of 14C and 35Sceptibility to CS2 toxicity, for examining the extent of protein
cross-linking occurring at presently acceptable levels of ex- into axonal proteins of rats administered radiolabeled CS2
reported previously (Savolainen et al., 1977) support a directposure, and for correlating a quantifiable pathological changeto levels of urinary TTCA. irreversible chemical modification of neurofilament proteins
by CS2. Therefore, based on the established interactions ofAnomalously high apparent molecular weight protein spe-cies possessing epitopes for each of the three neurofilament CS2 with proteins (Valentine et al., 1992, 1995), a more
consistent interpretation for generation of the high-molecu-subunit proteins were identified in the axonal proteins ob-tained from CS2-exposed rats but not from controls. Similar lar-weight neurofilament protein species is that of CS2-medi-
ated intermolecular covalent cross-linking.high-molecular-weight protein species involving all threeneurofilament triplet proteins have been reported in prepara- Morphologic findings paralleled those in previous studies
(Colombi et al., 1981; Gottfried et al., 1985) although thetions obtained from rats exposed to 2,5-hexanedione (Lapa-dula et al., 1986; Nagano et al., 1993). The conditions em- exposure protocol employed here allowed for a better assess-
ment regarding the onset of structural changes. Detectableployed here for analysis of the abnormal axonal proteins areexpected to effectively disrupt aggregations due to noncova- alterations in morphology appeared first as multifocal axonal
swellings in the descending tracts of the lumbar spinal cordlent interactions or disulfide bridges and support the genera-tion of slow-migrating neurofilament protein species through and by 13 weeks exposure, at the 500- and 800-ppm levels,
swellings were evident in all animals with equivalentcovalent modification. Although hyperphosphorylation can-not be completely ruled out as the mechanism responsible changes present in both sexes. Immunoblot results provided
evidence that cross-linking of NFL occurred prior to thefor generation of the abnormal neurofilament protein species,it appears unlikely. Considering the few sites present for formation of axonal swellings both at the two neurotoxic
levels and at the 50-ppm exposure level that was insufficientphosphorylation on NFL and that neurofilament proteins arenormally present in a relatively high degree of phosphoryla- to produce neurofilamentous swellings. Although quantita-
tive comparisons of the Western blots must be interpretedtion (Carden et al., 1985), it is difficult to account for the
AID TOX 8028 / 6h13$$$103 12-17-96 18:35:32 toxa AP: Tox
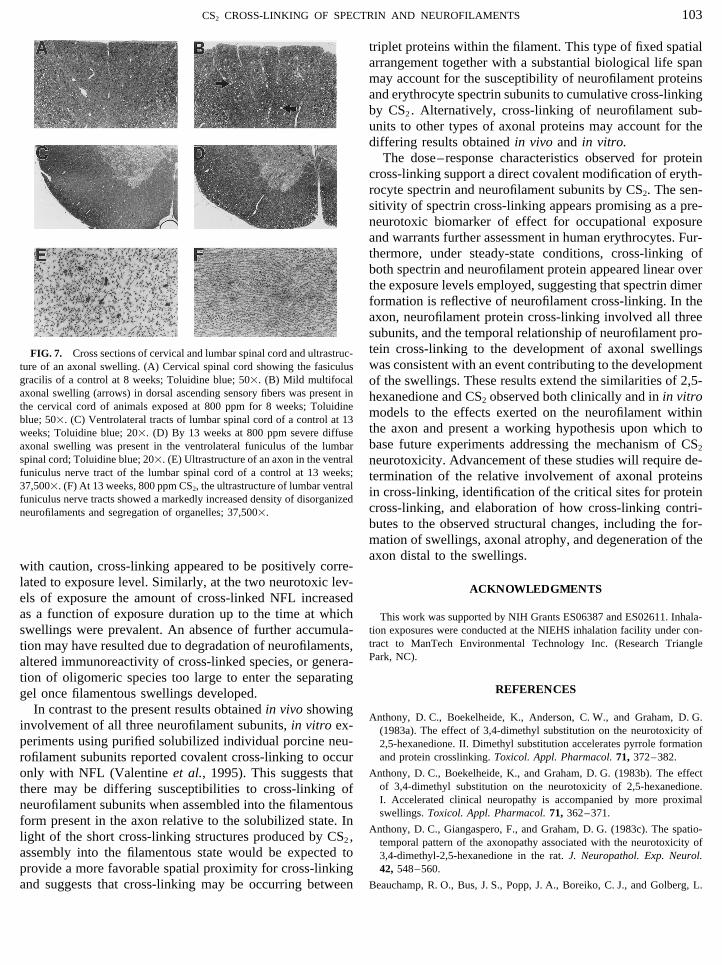
103CS2 CROSS-LINKING OF SPECTRIN AND NEUROFILAMENTS
triplet proteins within the filament. This type of fixed spatialarrangement together with a substantial biological life spanmay account for the susceptibility of neurofilament proteinsand erythrocyte spectrin subunits to cumulative cross-linkingby CS2. Alternatively, cross-linking of neurofilament sub-units to other types of axonal proteins may account for thediffering results obtained in vivo and in vitro.
The dose–response characteristics observed for proteincross-linking support a direct covalent modification of eryth-rocyte spectrin and neurofilament subunits by CS2. The sen-sitivity of spectrin cross-linking appears promising as a pre-neurotoxic biomarker of effect for occupational exposureand warrants further assessment in human erythrocytes. Fur-thermore, under steady-state conditions, cross-linking ofboth spectrin and neurofilament protein appeared linear overthe exposure levels employed, suggesting that spectrin dimerformation is reflective of neurofilament cross-linking. In theaxon, neurofilament protein cross-linking involved all threesubunits, and the temporal relationship of neurofilament pro-tein cross-linking to the development of axonal swellingsFIG. 7. Cross sections of cervical and lumbar spinal cord and ultrastruc-was consistent with an event contributing to the developmentture of an axonal swelling. (A) Cervical spinal cord showing the fasiculus
gracilis of a control at 8 weeks; Toluidine blue; 501. (B) Mild multifocal of the swellings. These results extend the similarities of 2,5-axonal swelling (arrows) in dorsal ascending sensory fibers was present in hexanedione and CS2 observed both clinically and in in vitrothe cervical cord of animals exposed at 800 ppm for 8 weeks; Toluidine models to the effects exerted on the neurofilament withinblue; 501. (C) Ventrolateral tracts of lumbar spinal cord of a control at 13
the axon and present a working hypothesis upon which toweeks; Toluidine blue; 201. (D) By 13 weeks at 800 ppm severe diffusebase future experiments addressing the mechanism of CS2axonal swelling was present in the ventrolateral funiculus of the lumbar
spinal cord; Toluidine blue; 201. (E) Ultrastructure of an axon in the ventral neurotoxicity. Advancement of these studies will require de-funiculus nerve tract of the lumbar spinal cord of a control at 13 weeks; termination of the relative involvement of axonal proteins37,5001. (F) At 13 weeks, 800 ppm CS2, the ultrastructure of lumbar ventral in cross-linking, identification of the critical sites for proteinfuniculus nerve tracts showed a markedly increased density of disorganized
cross-linking, and elaboration of how cross-linking contri-neurofilaments and segregation of organelles; 37,5001.butes to the observed structural changes, including the for-mation of swellings, axonal atrophy, and degeneration of theaxon distal to the swellings.
with caution, cross-linking appeared to be positively corre-lated to exposure level. Similarly, at the two neurotoxic lev- ACKNOWLEDGMENTSels of exposure the amount of cross-linked NFL increasedas a function of exposure duration up to the time at which This work was supported by NIH Grants ES06387 and ES02611. Inhala-
tion exposures were conducted at the NIEHS inhalation facility under con-swellings were prevalent. An absence of further accumula-tract to ManTech Environmental Technology Inc. (Research Triangletion may have resulted due to degradation of neurofilaments,Park, NC).altered immunoreactivity of cross-linked species, or genera-
tion of oligomeric species too large to enter the separatingREFERENCESgel once filamentous swellings developed.
In contrast to the present results obtained in vivo showingAnthony, D. C., Boekelheide, K., Anderson, C. W., and Graham, D. G.
involvement of all three neurofilament subunits, in vitro ex- (1983a). The effect of 3,4-dimethyl substitution on the neurotoxicity ofperiments using purified solubilized individual porcine neu- 2,5-hexanedione. II. Dimethyl substitution accelerates pyrrole formation
and protein crosslinking. Toxicol. Appl. Pharmacol. 71, 372–382.rofilament subunits reported covalent cross-linking to occurAnthony, D. C., Boekelheide, K., and Graham, D. G. (1983b). The effectonly with NFL (Valentine et al., 1995). This suggests that
of 3,4-dimethyl substitution on the neurotoxicity of 2,5-hexanedione.there may be differing susceptibilities to cross-linking ofI. Accelerated clinical neuropathy is accompanied by more proximalneurofilament subunits when assembled into the filamentousswellings. Toxicol. Appl. Pharmacol. 71, 362–371.
form present in the axon relative to the solubilized state. InAnthony, D. C., Giangaspero, F., and Graham, D. G. (1983c). The spatio-
light of the short cross-linking structures produced by CS2, temporal pattern of the axonopathy associated with the neurotoxicity ofassembly into the filamentous state would be expected to 3,4-dimethyl-2,5-hexanedione in the rat. J. Neuropathol. Exp. Neurol.
42, 548–560.provide a more favorable spatial proximity for cross-linkingand suggests that cross-linking may be occurring between Beauchamp, R. O., Bus, J. S., Popp, J. A., Boreiko, C. J., and Golberg, L.
AID TOX 8028 / 6h13$$$103 12-17-96 18:35:32 toxa AP: Tox

104 VALENTINE ET AL.
(1983). A critical review of the literature on carbon disulfide toxicity. indicator of carbon disulfide exposure, and its accumulation in repeatedlyexposed rats. Toxicol. Appl. Pharmacol. 70, 402–410.CRC Crit. Rev. Toxicol. 11, 169–278.
Lam, C. W., and DiStefano, V. (1986). Characterization of carbon disulfideCarden, M. J., Schlapfer, W. W., and Lee, V. M. Y. (1985). The structure,binding in blood and to other biological substances. Toxicol. Appl. Phar-biochemical properties and immunogenicity of neurofilament peripheralmacol. 86, 235–242.regions are determined by phosphorylation state. J. Biol. Chem. 260,
9805–9817. Lapadula, D. M., Irwin, R. D., Suwita, E., and Abou-Donia, M. B. (1986).Cross-linking of neurofilament proteins of rat spinal cord in vivo afterCavanagh, J. B. (1982). The pattern of recovery of axons in the nervousadministration of 2,5-hexanedione. J. Neurochem. 46, 1843–1850.system of rats following 2,5-hexanediol intoxication: A question of rheol-
ogy? Neuropathol. Appl. Neurobiol. 104, 297–318. Lehning, E. J., Dyer, K. S., Jortner, B. S., and LoPachin, R. M. (1995).Axonal atrophy is a specific component of 2,5-hexanedione peripheralCavanagh, J. B., and Bennetts, R. J. (1981). On the pattern of changes inneuropathy. Toxicol. Appl. Pharmacol. 135, 58–66.the rat nervous system produced by 2,5-hexanediol. Brain 104, 297–318.
Liem, R. K. H., and Hutchison, S. B. (1982). Purification of individual com-Chu, C. C., Huang, C. C., Chen, R. S., and Shih, T. S. (1995). Polyneuropa-ponents of the neurofilament triplet: Filament assembly from the 70 000-thy induced by carbon disulfide in viscose rayon workers. Occup. Environ.dalton unit. Biochemistry 21, 3221–3226.Med. 52, 404–407.
Lynch, G., and Baudry, M. (1987). Brain spectrin, calpain and long-termClark, A., Griffin, J. W., and Price, D. L. (1980). The axonal pathologychanges in synaptic efficacy. Brain Res. Bull. 18, 809–815.in chronic b,b*-iminodipropionitrile intoxication. J. Neuropathol. Exp.
Neurol. 39, 42–55. McKerracher, L., Vidal-Sanz, M., and Aguayo, A. J. (1990). Slow transportrates of cytoskeletal proteins change during regeneration of axotomizedColombi, A., Maroni, M., Picchi, E., Rota, E., Castano, P., and Foa, V.retinal neurons in adult rats. J. Neurosci. 10, 641–648.(1981). Carbon disulfide neuropathy in rats. A morphological and ultra-
structural study of degeneration and regeneration. Clin. Toxicol. 18, Meuling, W. J. A., Bragt, P. C., and Braun, C. L. J. (1990). Biological moni-toring of carbon disulfide. Am. J. Ind. Med. 17, 247–254.1463–1474.
Monaco, S., Jacob, J., Jenich, H., Patton, A., Autilo-Gambetti, L., andDavidson, M., and Feinlab, M. (1972). Carbon disulfide poisoning: A re-Gambetti, P. (1989). Axonal transport of neurofilament is accelerated inview. Am. Heart J. 83, 100–114.peripheral nerve during 2,5-hexanedione intoxication. Brain Res. 491,DeCaprio, A. P., and O’Neill, E. A. (1985). Alterations in rat cytoskeletal328–334.proteins induced by in vitro and in vivo 2,5-hexanedione exposure. Tox-
Monaco, S., Wongmongkolrit, T., Shearson, C. M., Patton, A., Schaetzle,icol. Appl. Pharmacol. 78, 235–247.B., Autilo-Gambetti, L., Gambetti, P., and Sayre, L. M. (1990). GiantDrexler, H., Goen, T., Angerer, J., Aboul-el-ela, S., and Lehnert, G. (1994).axonopathy characterized by intermediate location of axonal enlarge-Carbon disulphide. I. External and internal exposure to carbon disulphidements and acceleration of neurofilament transport. Brain Res. 519, 73–of workers in the viscose industry. Int. Arch. Occup. Environ. Health 65,81.359–365.
Nagano, M., Yamamoto, H., Harada, K., Miyamoto, E., and Futatsuka, M.Fennell, T. R., Sumner, S. C. J., and Walker, V. E. (1992). A model for(1993). Comparative study of modification and degradation of neurofil-the formation and removal of hemoglobin adducts. Cancer Epidemiol.ament proteins in rats subchronically treated with allyl chloride, acryl-Biomark. Prev. 1, 213–219.amide, or 2,5-hexanedione. Environ. Res. 63, 229–240.
Genter St Clair, M. B., Amarnath, V., Moody, M. A., Anthony, D. C., An-Pappolla, M., Penton, R., Weiss, H. S., Miller, C. H., Jr., Sahenk, Z., andderson, C. W., and Graham, D. G. (1988). Pyrrole oxidation and protein
Autilio-Gambetti, L., and Gambetti, P. (1987). Carbon disulfide axonopa-cross-linking as necessary steps in the development of g-diketone neurop-thy. Another experimental model characterized by acceleration of neuro-athy. Chem. Res. Toxicol. 1, 179–185.filament transport and distinct changes of axonal size. Brain Res. 424,
Gottfried, M. R., Graham, D. G., Morgan, M., Cases, H. W., and Bus, J. S. 272–280.(1985). The morphology of carbon disulfide neurotoxicity. Neurotoxicity
Pyle, S. J., Amarnath, V., Graham, D. G., and Anthony, D. C. (1992). The6, 89–96.role of pyrrole formation in the alteration of neurofilament transport
Graham, D. G., Amarnath, V., Valentine, W. M., Pyle, S. J., and Anthony, induced during exposure to 2,5-hexanedione. J. Neuropathol. Exp. Neu-D. C. (1995). Pathogenetic studies of hexane and carbon disulfide neuro- rol. 51, 451–458.toxicity. CRC Crit. Rev. Toxicol. 25, 91–112.
Pyle, S. J., Amarnath, V., Graham, D. G., and Anthony, D. C. (1993). De-Herskowitz, A., Ishii, N., and Schamburg, H. (1971). n-Hexane neuropathy: creased levels of the high molecular weight subunit of neurofilaments
A syndrome occurring as a result of industrial exposure. N. Engl. J. Med. and accelerated neurofilament transport during the recovery phase of 2,5-285, 82. hexanedione exposure. Cell Motil. Cytoskel. 26, 133–143.
Jirmanova, I., and Lukas, E. (1984). Ultrastructure of carbon disulfide neu- Rosenberg, C. K., Anthony, D. C., Szakal-Quin, G., Genter, M. B., andropathy. Acta Neuropathol. (Berl.) 63, 255–263. Graham, D. G. (1987). Hyperbaric oxygen accelerates the neurotoxicity
of 2,5-hexanedione. Toxicol. Appl. Pharmacol. 87, 374–379.Johnson, D. J., Graham, D. G., Amarnath, V., Amarnath, K., and Valentine,W. M. (1996). The measurement of 2-thiothiazolidine-4-carboxylic acid Ruijten, M. W., Salle, H. J., and Verberk, M. M. (1993). Verification ofas an index of the in vivo release of CS2 by dithiocarbamates. Chem. effects on the nervous system of low level occupational exposure to CS2.Res. Toxicol., 9, 910–916. Br. J. Ind. Med. 50, 301–307.
Kivisto, H., Elovaara, E., Riihimaki, V., and Aitio, A. (1995). Effect of Savolainen, H., Lehtonen, E., and Vaino, H. (1977). Carbon disulfide bind-cytochrome P450 isozyme induction and glutathione depletion on the ing to rat spinal neurofilaments. Acta Neuropathol. (Berl.) 37, 219–223.metabolism of CS2 to TTCA in rats. Arch. Toxicol. 69, 185–190. Sayre, L. M., Autilio-Gambetti, L., and Gambetti, P. (1985). Pathogenesis of
Lam, C. W., and DiStefano, V. (1982). Behavior and characterization of experimental giant neurofilamentous axonopathies: A unified hypothesisblood carbon disulfide in rats after inhalation. Toxicol. Appl. Pharmacol. based on chemical modification of neurofilaments. Brain Res. Rev. 18,64, 327–334. 69–75.
Schaumburg, H. H., and Spencer, P. S. (1976). Degeneration in central andLam, C. W., and DiStefano, V. (1983). Blood-bound carbon disulfide: An
AID TOX 8028 / 6h13$$$104 12-17-96 18:35:32 toxa AP: Tox

105CS2 CROSS-LINKING OF SPECTRIN AND NEUROFILAMENTS
peripheral nervous systems produced by pure n-hexane: An experimental D. G. (1995). Carbon disulfide mediated protein crosslinking by N, N-diethyldithiocarbamate. Chem. Res. Toxicol. 8, 96–102.study. Brain 99, 183–192.
Simon, P., Nicot, T., and Dieudonne, M. (1994). Dietary habits, a non- Valentine, W. M., Amarnath, V., Graham, D. G., and Anthony, D. C.negligible source of 2-thiothiazolidine-4-carboxylic acid and possible (1992). Covalent cross-linking of proteins by carbon disulfide. Chem.over estimation of carbon disulfide exposure. Int. Arch. Occup. Environ. Res. Toxicol. 5, 254–262.Health 66, 85–90. Valentine, W. M., Graham, D. G., and Anthony, D. C. (1993). Covalent
Spencer, P. S., and Schaumburg, H. H. (1977). Ultrastructural studies of cross-linking of erythrocyte spectrin by carbon disulfide in vivo. Toxicol.the dying back process III. The evolution of experimental giant axonal Appl. Pharmacol. 121, 71–77.degeneration. J. Neuropathol. Exp. Neurol. 36, 276–299. Watson, D. F., Fittro, K. P., Hoffman, P. N., and Griffin, J. W. (1991).
Spencer, P. S., Schaumburg, H. H., Sabri, M. I., and Veronesi, B. V. (1980). Phosphorylation-related immunoreactivity and the rate of transport ofThe enlarging view of hexacarbon neurotoxicity. CRC Crit. Rev. Toxicol. neurofilaments in chronic 2,5-hexanedione intoxication. Brain Res. 539,7, 279–356. 103–109.
Szendzikowski, S., Setkiewicz, J., Wronska-Nofer, T., and Zdrajkowska, I. Zhu, M., Spink, D. C., Yan, B., Bank, S., and DeCaprio, A. P. (1994).(1973). Structural aspects of experimental carbon disulfide intoxication. Formation and structure of cross-linking and monomeric pyrrole autoxi-Int. Arch. Arbeits Med. 31, 135–149. dation products in 2,5-hexanedione-treated amino acids, peptides, and
protein. Chem. Res. Toxicol. 7, 551–558.Valentine, W. M., Amarnath, V., Amarnath, K., Rimmele, F., and Graham,
AID TOX 8028 / 6h13$$$104 12-17-96 18:35:32 toxa AP: Tox
Copyright © 2022 FDOKUMEN




















