
Analysis and simulation of frontal affinity chromatography of proteins
Upload
independentCategory
view
0download
0
Protein Expression and Purification 60 (2008) 157–164
Contents lists available at ScienceDirect
Protein Expression and Purification
journal homepage: www.elsevier .com/locate /yprep
The 22.5 kDa spectrin-binding domain of ankyrinR binds spectrin with highaffinity and changes the spectrin distribution in cells in vivo
Adam Kolondra, Michal Grzybek, Anna Chorzalska, Aleksander F. Sikorski *
Laboratory of Cytobiochemistry, Biotechnology Faculty, University of Wroclaw, ul Przybyszewskiego 63/77, 51148 Wroclaw, Poland
a r t i c l e i n f o a b s t r a c t
Article history:Received 7 January 2008and in revised form 27 March 2008Available online 14 April 2008
Keywords:AnkyrinR (Ankyrin-1)SpectrinSpectrin-binding domainAnkyrin–spectrin interactionExpression in E. coli
1046-5928/$ - see front matter � 2008 Elsevier Inc. Adoi:10.1016/j.pep.2008.04.002
* Corresponding author. Fax: +48 71 375 62 08.E-mail address: [email protected] (A.F. Sikor
1 Abbreviations used: SBD, spectrin-binding domain;nate; InsP3R, inositol 1,4,5-triphosphate receptor; L1CAmolecules; DD, death-domain.
It was previously shown that ankyrins play a crucial role in the membrane skeleton arrangement. Puri-fying ankyrinR obtained from erythrocytes is a time-consuming process. Therefore, cloned and bacteriallyexpressed ankyrinR–spectrin-binding domain (AnkSBD) is a demanded tool for studying spectrin–anky-rin interactions. In this communication, we report on the cloning and purification of AnkSBD and describethe results of binding experiments, in which we showed high-affinity interactions between the AnkSBDconstruct and isolated erythrocyte or non-erythroid spectrins. pEGFP–AnkSBD-transfected cells co-local-ised with non-erythroid spectrin in HeLa cells. The functional interactions of the AnkSBD construct in vivoand in vitro open many possibilities to study the structure and function of this domain, which has not yetbeen as extensively studied when compared to the aminoterminal domain of this protein.
� 2008 Elsevier Inc. All rights reserved.
Ankyrins are membrane proteins that are involved in the properassembly of the membrane skeleton [1–3]. The three main groupsof ankyrin isoforms, encoded by three known ankyrin genes(ank1—ankyrinR [4,5], ank2—ankyrinB [6] and ank3—ankyrinG[7]), and the alternative splicing of these genes’ primary transcriptsproduce the numerous isoforms found in human cells [8]. Ankyrinscan be divided into canonical, giant (with the insertion of up to2500 amino acid residues) and small variants (lacking one or moreof the main ankyrin domains) [2]. Erythrocyte ankyrin that belongsto the canonical group consist of three main domains: the N-termi-nal, central SBD (spectrin-binding domain)1 and C-terminal regula-tory domains.
The N-terminal membrane-binding domain consists of 24; 33amino acid residue motifs (ANK repeats) folded into four-indepen-dent subdomains of six repeats each [1,8–10]. The ANK repeatwhich is proposed to act as a generalised protein-binding motif[11] is similar to an earlier identified motif that was found inSwi6/Cdc10—a yeast cell cycle regulator protein—and in Notch sig-nalling protein from Drosophila melanogaster [12]. It was also foundin many other proteins, including the p105 precursor of the NFjBp50 subunit [13,14] and the NFjB inhibitor family members: a sub-unit IjBa [15], b subunit IjBb [16] and c subunit IjBc [17] or Bcl-3[18]. Another group of proteins that contain ANK repeats consists
ll rights reserved.
ski).RITC, rhodamine isothiocya-
Ms, L1 family of cell-adhesion
of 53BP2, the family of INK4 tumour suppressors, GABP-b tran-scription factor cell and the cycle inhibitors p16, p18 and p19 [19].
The main role of the 24ANK repeat motif in the ankyrin family isto anchor the spectrin–actin lattice to the cell membrane. It inter-acts with membrane integral proteins like a-Na+/K+-ATPase[20,21], H+/K+-ATPase [22], the L1 family of cell-adhesion mole-cules (L1CAMs) and CD44 [23–25], AE1-3 (a Cl�/HCO3-anion ex-changer) [26–28], the Na+ channel [29], clathrin heavy chain [30],Rh and RhAG [31], and the inositol 1,4,5-triphosphate receptor(InsP3R) [32]. Ankyrin was also shown to bind with high affinity di-rectly to CD45 in peripheral blood lymphocytes (PBLs) and Jurkat Tcells (KD = 4.3 ± 3.0 nM) [33]. The crystal structure of the 12ANK re-peats (D34) of the ankyrinR isoform showed that this fragment isfolded into antiparallel a-helices with a loop forming an L-shapedstructure [34].
The C-terminal regulatory domain, which seems to be the mostvariable ankyrin domain, is responsible for the regulation of thebinding activity of most ankyrin isoforms [35,36]. It allows partic-ular ankyrin isoforms to be distinguished [8].
In the ankyrinG isoform, SBD was localised within a sequenceencompassing residues 842–1420, homologous to rabbit ankyrinIII [22]. The ankyrinB isoform, first found in the brain [26], containsa 62 kDa SBD, and is known to associate with the bII-spectrin iso-form with high affinity (KD = 25 nM) [37]. The minimal SBD of ank-yrinB consists of a sequence spanning amino acid residues 966 and1125 [38]. In erythroid ankyrinR, SBD was previously shown to belinked directly to 24ANK repeat motif (residues 828–1386, 72 kDadomain), and was shown to bind erythrocyte spectrin with highaffinity (KD = 7 nM). In addition, a shorter ankyrinR fragment62 kDa (residues 862–1386) was found to bind spectrin with a

158 A. Kolondra et al. / Protein Expression and Purification 60 (2008) 157–164
KD 10 times higher, while a fragment encompassing residues 898–1386 with a significantly higher KD (200 nM) [39]. All these ankyrinSBD sequences include a ZU5 domain which plays an as yet un-known role function and was also found in the NFjB/Rel transcrip-tion factor family of proteins. Outside the spectrin-binding domain,most of the ankyrin isoforms contain the death-domain (DD),which was found in proapoptotic signalling proteins such as Fas.Proteins that contain both a ZU5 domain and DD domain have beenpostulated to be grouped as ZUD (ZU5 and death-domain contain-ing proteins), which play an inhibitory role in NFjB-dependenttranscription and sensitise the HEK 293 cell line to TNF andTRAIL-induced apoptosis [40]. On the other hand, Platt et al. [41]reported an erythroid ankyrin sequence including residues 1136–1192 with a highly positively charged motif that was also foundto bind spectrin on ligand blots.
The isolation of ankyrin is a time-consuming procedure and dueto the presence of many isoforms and high sensitivity to cellularproteases, it is difficult to obtain intact protein. We undertook aneffort to clone and express a functional spectrin-binding domainof erythrocyte ankyrin. The obtained product bears high sequencehomology to the spectrin-binding domains of all three main anky-rin isoforms therefore can be used to study the ankyrin–spectrininteractions in other cell models. The cloned product was also usedto follow the behaviour of ankyrin in HeLa cells, where AnkSBD–GFP was found to dramatically change the localisation of spectrinand to affect the shape of cells and their viability.
Materials and methods
All primers were synthesised by IBB Oligo Synthesis (Warsaw,Poland) (Table 1). The Taq Polymerase, One-Step RT-PCR andRNA Tissue kits were from Qiagen (Germany). The restriction en-zymes were from Promega (USA). The T4 DNA Ligase and Fugenetransfection reagent were from Roche (Switzerland). Talon� resinwas purchased from Clontech (USA), DNA molecular markers andloading dye were from Fermentas (Lithuania). The Plasmid Isola-tion Kit was from Eppendorf (Germany).
Plasmids and bacterial strains
The plasmid pEGFP-C1, the cloning host Escherichia coli XL1-Blueand the JM109 strain were from Stratagene (USA), and the plasmidpRSETC and E. coli BL21(DE3)pLysE strain were from Invitrogen(USA).
RNA preparation from reticulocyte-enriched fraction of human bloodreticulocyte cDNA synthesis and cloning of AnkSBD cDNA in pRSETC
RNA was isolated from the reticulocyte-enriched fraction,which was prepared according to Downey et al. [42] using RNA Tis-sue Kit (Qiagen), from freshly drawn human blood given by one ofthe authors. The erythrocyte lysis step was omitted from the prep-aration protocol. The cDNA, forward (FN) and reverse primer (RN)
Table 1The primers used in the experiment
FN (2337) 50-TACATTTCTGTCACCGACGTGC-30
RN (4218) 50-CTTCATCTCTGCCTGCTCTGTC-30
FP (2742) 50-CTGGTGAGCTTCATGGTTGACGC-30
RP (3113) 50-TCGTCCATCCCGTTGAGGATCTG-30
T7 50-GTAATACGACTCACTATAG-30
F2C909XhoI 50-CGACCTCGAGTGCATACAGGGTTTCTGGTGAGC-30
R2C1050HindIII 50-CGGATCAAGCTTCCTCTTCTTCTCTAGCTCCTCCAG-30
The position of the primers in the ankyrin mRNA coding sequence is in brackets, andthe restriction sites are underlined.
(see Table 1) were used to amplify the human ankyrinR cDNA frag-ment from the reticulocytic mRNA. In detail, 5 ll of RNA was mixedin a final volume of 25 ll of the PCR cocktail and was used underthe following conditions: 50 �C for 30 min, 95 �C for 15 min, 40 cy-cles of [94 �C for 30 s, 60 �C for 1 min and 72 �C for 2 min], with afinal extension for 10 min at 72 �C in a BIOMETRA thermocycler.The cDNA was diluted 1000 times and 1 ll was used for the PCRamplification of the AnkSBD fragment using a pair of primers:F2C909XhoI and R2C1050HindIII (Table 1) in a 50 ll volume ofthe PCR cocktail. The following conditions were used: 94 �C for5 min, 35 cycles of [94 �C for 30 s, 60 �C for 1 min and 72 �C for30 s], with a final extension as above. The PCR product and plasmidwere doubly digested with XhoI/HindIII restriction enzymes and li-gated at a 13:1 ratio (1 h at 20 �C in a 10 ll volume) (Fig. 1b).
Positive clone screening and expression of the pRSETC–AnkSBDconstruct in the E. coli BL21(DE3)pLysE strain
Transformed E. coli JM109 strains were screened via PCR using aconstruct-specific primer FP and commercial primer T7, in a vol-ume of 20 ll with bacterial stocks as a template, under the follow-ing conditions: 94 �C for 15 min, 35 cycles of [94 �C for 45 s, 54 �Cfor 1 min, and 72 �C for 1 min], with a final extension of 10 min.Plasmids from the positive clones were isolated and digested,and the identity of inserts was finally confirmed by sequencing. Po-sitive pRSETC–AnkSBD plasmid was transformed into the E. coliBL21(DE3)pLysE strain. Bacteria were plated into LB Agar mediumcontaining 50 lg/ml ampicillin and 34 lg/ml chloramphenicol.Expression was performed as follows: BL21(DE3)pLysE bacteriawere cultured in LB medium containing 50 lg/ml ampicillin and34 lg/ml chloramphenicol at 37 �C until OD600nm reached 0.6.The expression of AnkSBD protein was induced by adding IPTG toa final concentration of 0.75 mM. After 3 h at 37 �C, cells were har-vested by centrifugation and stored at �70 �C.
Purification of expressed proteins by affinity chromatography
Bacteria were thawed on ice and treated with the lysis buffer(20 mM Tris–HCl, 100 mM NaCl, 8 M urea, pH 8.0), pushed 10times through a 25 G needle and centrifuged (35,000g for 20 minat 4 �C). The supernatant was loaded onto a 2 ml bed volume Ta-lon� column which was pre-equilibrated with the lysis buffer.The resin was then washed with 15 ml of the washing buffer(20 mM Tris–HCl, 100 mM NaCl, 8 M urea, 1 mM imidazole, pH8.0) until A280nm was lower than 0.05, followed by washing with10 ml of the washing buffer without urea. Elution was performedwith: 20 mM Tris–HCl, 100 mM NaCl, 50 mM imidazole, pH 8.0(elution buffer), containing 8 M urea.
Cloning of an AnkSBD fragment into the pEGFP-C1 plasmid and HeLacells transfection
An insert clone was cut out from a positive pRSETC–AnkSBDwith XhoI/HindIII restriction enzymes and cloned into a pEG-FP-C1 transfection vector coding for Green Fluorescent Proteinas a tag protein (Fig. 1b). Positive clones were screened as de-scribed above by using FP/RP primer combination. pEGFP–AnkSBD plasmid was purified using Plasmid Mini Kit (Eppen-dorf) and used for transfection. HeLa cells were grown directlyon cover slips for 8 h in 6-well, 35 mm plates in MEMa mediumwith 10% FCS and antibiotics (100 lg/ml streptomycin and100 lg/ml penicillin), then transferred to MEMa medium (with-out FCS and antibiotics) for 3 h before transfection. Cells weretransfected with a Fugene transfection reagent (Roche):DNA(pEGFP–AnkSBD) 3:1 mixture, and after 3 h of incubation, 10%FCS and antibiotics were added. At specified time points, cells
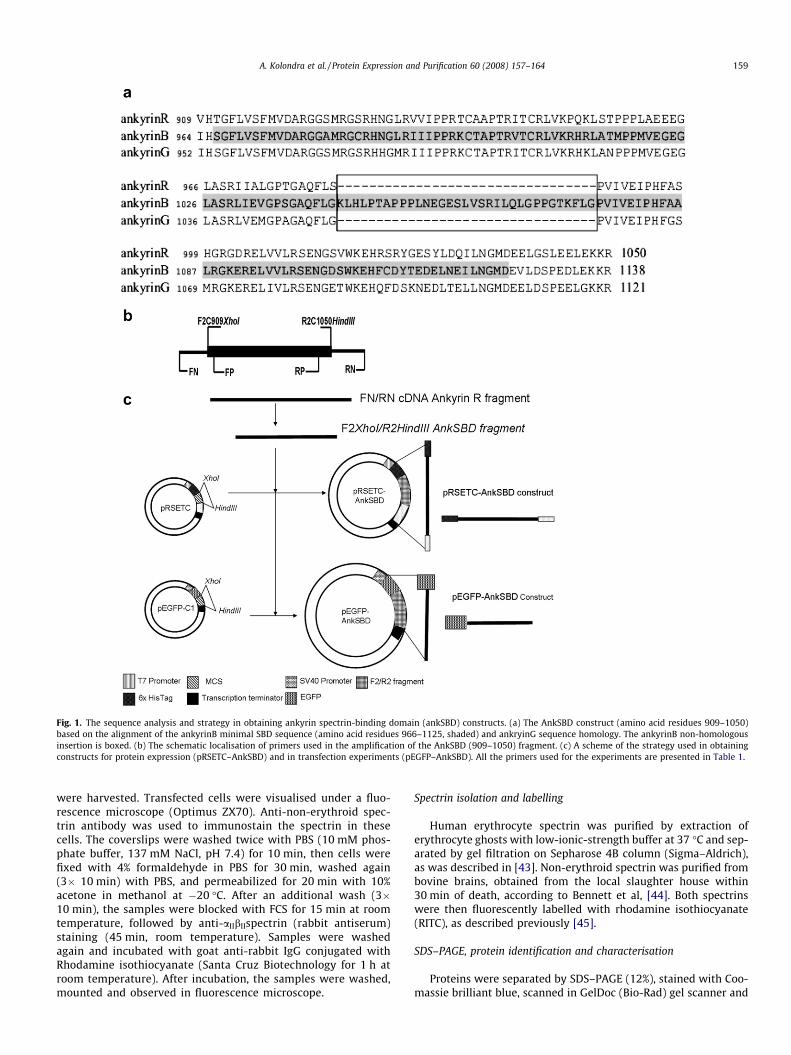
Fig. 1. The sequence analysis and strategy in obtaining ankyrin spectrin-binding domain (ankSBD) constructs. (a) The AnkSBD construct (amino acid residues 909–1050)based on the alignment of the ankyrinB minimal SBD sequence (amino acid residues 966–1125, shaded) and ankryinG sequence homology. The ankyrinB non-homologousinsertion is boxed. (b) The schematic localisation of primers used in the amplification of the AnkSBD (909–1050) fragment. (c) A scheme of the strategy used in obtainingconstructs for protein expression (pRSETC–AnkSBD) and in transfection experiments (pEGFP–AnkSBD). All the primers used for the experiments are presented in Table 1.
A. Kolondra et al. / Protein Expression and Purification 60 (2008) 157–164 159
were harvested. Transfected cells were visualised under a fluo-rescence microscope (Optimus ZX70). Anti-non-erythroid spec-trin antibody was used to immunostain the spectrin in thesecells. The coverslips were washed twice with PBS (10 mM phos-phate buffer, 137 mM NaCl, pH 7.4) for 10 min, then cells werefixed with 4% formaldehyde in PBS for 30 min, washed again(3� 10 min) with PBS, and permeabilized for 20 min with 10%acetone in methanol at �20 �C. After an additional wash (3�10 min), the samples were blocked with FCS for 15 min at roomtemperature, followed by anti-aIIbIIspectrin (rabbit antiserum)staining (45 min, room temperature). Samples were washedagain and incubated with goat anti-rabbit IgG conjugated withRhodamine isothiocyanate (Santa Cruz Biotechnology for 1 h atroom temperature). After incubation, the samples were washed,mounted and observed in fluorescence microscope.
Spectrin isolation and labelling
Human erythrocyte spectrin was purified by extraction oferythrocyte ghosts with low-ionic-strength buffer at 37 �C and sep-arated by gel filtration on Sepharose 4B column (Sigma–Aldrich),as was described in [43]. Non-erythroid spectrin was purified frombovine brains, obtained from the local slaughter house within30 min of death, according to Bennett et al, [44]. Both spectrinswere then fluorescently labelled with rhodamine isothiocyanate(RITC), as described previously [45].
SDS–PAGE, protein identification and characterisation
Proteins were separated by SDS–PAGE (12%), stained with Coo-massie brilliant blue, scanned in GelDoc (Bio-Rad) gel scanner and
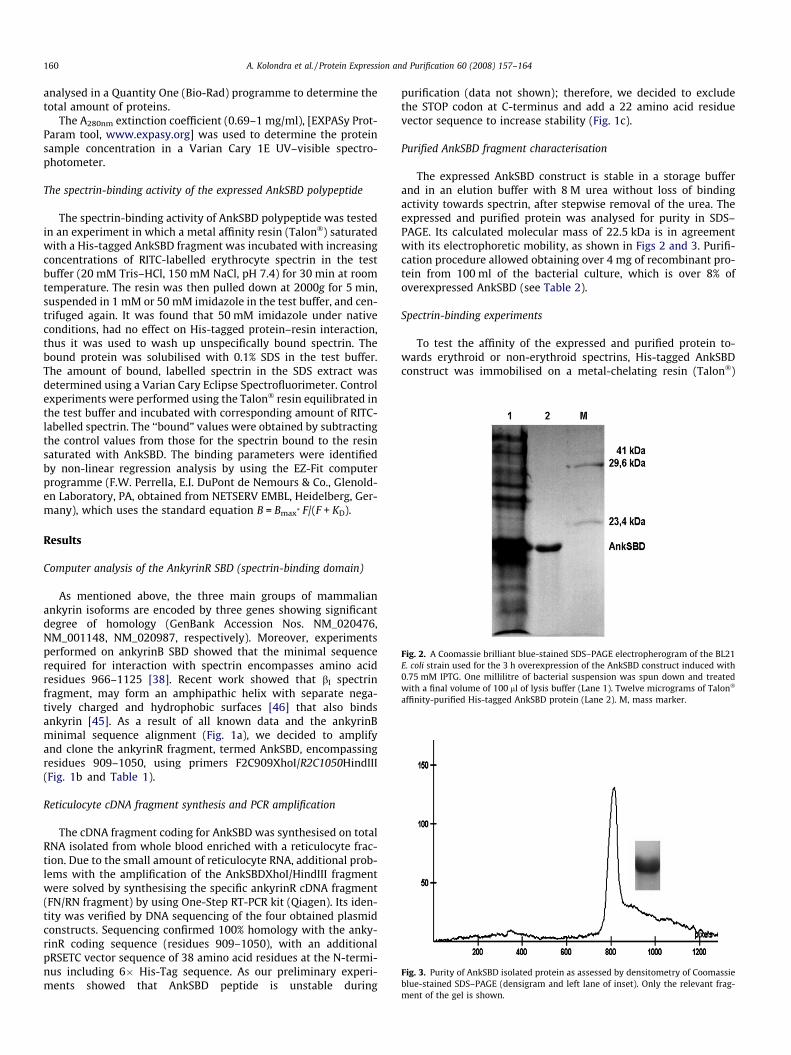
Fig. 2. A Coomassie brilliant blue-stained SDS–PAGE electropherogram of the BL21E. coli strain used for the 3 h overexpression of the AnkSBD construct induced with0.75 mM IPTG. One millilitre of bacterial suspension was spun down and treatedwith a final volume of 100 ll of lysis buffer (Lane 1). Twelve micrograms of Talon�
affinity-purified His-tagged AnkSBD protein (Lane 2). M, mass marker.
Fig. 3. Purity of AnkSBD isolated protein as assessed by densitometry of Coomassieblue-stained SDS–PAGE (densigram and left lane of inset). Only the relevant frag-
160 A. Kolondra et al. / Protein Expression and Purification 60 (2008) 157–164
analysed in a Quantity One (Bio-Rad) programme to determine thetotal amount of proteins.
The A280nm extinction coefficient (0.69–1 mg/ml), [EXPASy Prot-Param tool, www.expasy.org] was used to determine the proteinsample concentration in a Varian Cary 1E UV–visible spectro-photometer.
The spectrin-binding activity of the expressed AnkSBD polypeptide
The spectrin-binding activity of AnkSBD polypeptide was testedin an experiment in which a metal affinity resin (Talon�) saturatedwith a His-tagged AnkSBD fragment was incubated with increasingconcentrations of RITC-labelled erythrocyte spectrin in the testbuffer (20 mM Tris–HCl, 150 mM NaCl, pH 7.4) for 30 min at roomtemperature. The resin was then pulled down at 2000g for 5 min,suspended in 1 mM or 50 mM imidazole in the test buffer, and cen-trifuged again. It was found that 50 mM imidazole under nativeconditions, had no effect on His-tagged protein–resin interaction,thus it was used to wash up unspecifically bound spectrin. Thebound protein was solubilised with 0.1% SDS in the test buffer.The amount of bound, labelled spectrin in the SDS extract wasdetermined using a Varian Cary Eclipse Spectrofluorimeter. Controlexperiments were performed using the Talon� resin equilibrated inthe test buffer and incubated with corresponding amount of RITC-labelled spectrin. The ‘‘bound” values were obtained by subtractingthe control values from those for the spectrin bound to the resinsaturated with AnkSBD. The binding parameters were identifiedby non-linear regression analysis by using the EZ-Fit computerprogramme (F.W. Perrella, E.I. DuPont de Nemours & Co., Glenold-en Laboratory, PA, obtained from NETSERV EMBL, Heidelberg, Ger-many), which uses the standard equation B = Bmax* F/(F + KD).
Results
Computer analysis of the AnkyrinR SBD (spectrin-binding domain)
As mentioned above, the three main groups of mammalianankyrin isoforms are encoded by three genes showing significantdegree of homology (GenBank Accession Nos. NM_020476,NM_001148, NM_020987, respectively). Moreover, experimentsperformed on ankyrinB SBD showed that the minimal sequencerequired for interaction with spectrin encompasses amino acidresidues 966–1125 [38]. Recent work showed that bI spectrinfragment, may form an amphipathic helix with separate nega-tively charged and hydrophobic surfaces [46] that also bindsankyrin [45]. As a result of all known data and the ankyrinBminimal sequence alignment (Fig. 1a), we decided to amplifyand clone the ankyrinR fragment, termed AnkSBD, encompassingresidues 909–1050, using primers F2C909XhoI/R2C1050HindIII(Fig. 1b and Table 1).
Reticulocyte cDNA fragment synthesis and PCR amplification
The cDNA fragment coding for AnkSBD was synthesised on totalRNA isolated from whole blood enriched with a reticulocyte frac-tion. Due to the small amount of reticulocyte RNA, additional prob-lems with the amplification of the AnkSBDXhoI/HindIII fragmentwere solved by synthesising the specific ankyrinR cDNA fragment(FN/RN fragment) by using One-Step RT-PCR kit (Qiagen). Its iden-tity was verified by DNA sequencing of the four obtained plasmidconstructs. Sequencing confirmed 100% homology with the anky-rinR coding sequence (residues 909–1050), with an additionalpRSETC vector sequence of 38 amino acid residues at the N-termi-nus including 6� His-Tag sequence. As our preliminary experi-ments showed that AnkSBD peptide is unstable during
purification (data not shown); therefore, we decided to excludethe STOP codon at C-terminus and add a 22 amino acid residuevector sequence to increase stability (Fig. 1c).
Purified AnkSBD fragment characterisation
The expressed AnkSBD construct is stable in a storage bufferand in an elution buffer with 8 M urea without loss of bindingactivity towards spectrin, after stepwise removal of the urea. Theexpressed and purified protein was analysed for purity in SDS–PAGE. Its calculated molecular mass of 22.5 kDa is in agreementwith its electrophoretic mobility, as shown in Figs 2 and 3. Purifi-cation procedure allowed obtaining over 4 mg of recombinant pro-tein from 100 ml of the bacterial culture, which is over 8% ofoverexpressed AnkSBD (see Table 2).
Spectrin-binding experiments
To test the affinity of the expressed and purified protein to-wards erythroid or non-erythroid spectrins, His-tagged AnkSBDconstruct was immobilised on a metal-chelating resin (Talon�)
ment of the gel is shown.
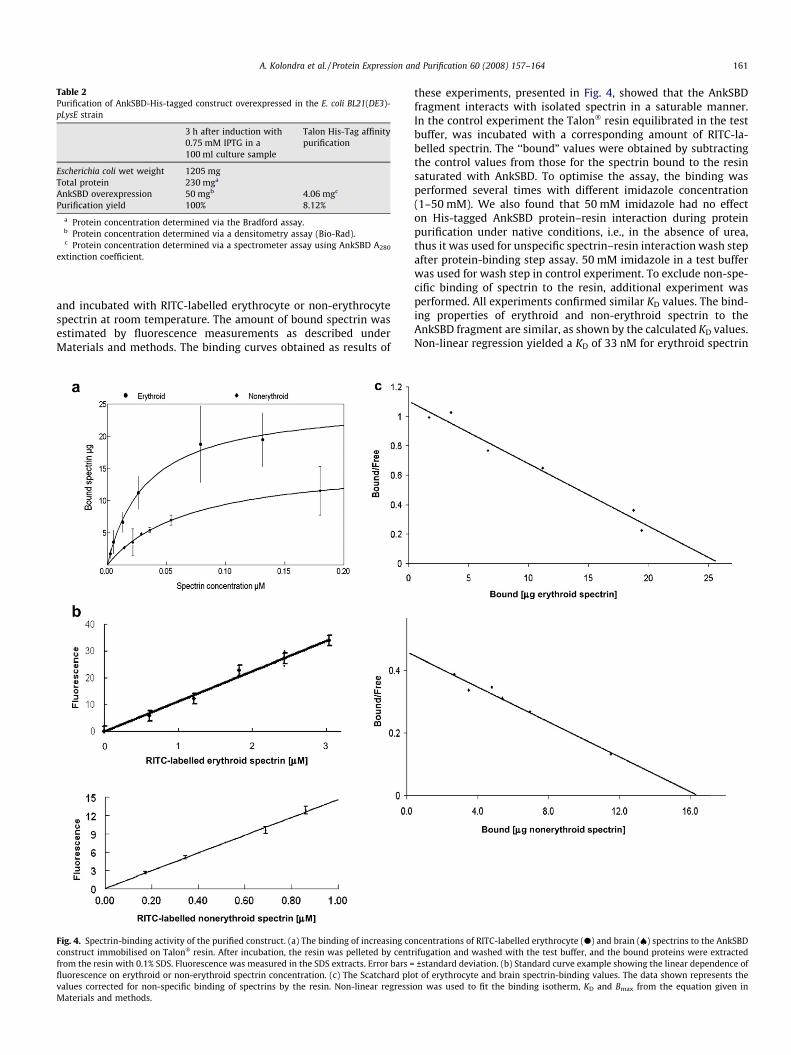
Table 2Purification of AnkSBD-His-tagged construct overexpressed in the E. coli BL21(DE3)-pLysE strain
3 h after induction with0.75 mM IPTG in a100 ml culture sample
Talon His-Tag affinitypurification
Escherichia coli wet weight 1205 mgTotal protein 230 mga
AnkSBD overexpression 50 mgb 4.06 mgc
Purification yield 100% 8.12%
a Protein concentration determined via the Bradford assay.b Protein concentration determined via a densitometry assay (Bio-Rad).c Protein concentration determined via a spectrometer assay using AnkSBD A280
extinction coefficient.
A. Kolondra et al. / Protein Expression and Purification 60 (2008) 157–164 161
and incubated with RITC-labelled erythrocyte or non-erythrocytespectrin at room temperature. The amount of bound spectrin wasestimated by fluorescence measurements as described underMaterials and methods. The binding curves obtained as results of
Fig. 4. Spectrin-binding activity of the purified construct. (a) The binding of increasing coconstruct immobilised on Talon� resin. After incubation, the resin was pelleted by centfrom the resin with 0.1% SDS. Fluorescence was measured in the SDS extracts. Error bars =fluorescence on erythroid or non-erythroid spectrin concentration. (c) The Scatchard plovalues corrected for non-specific binding of spectrins by the resin. Non-linear regressiMaterials and methods.
these experiments, presented in Fig. 4, showed that the AnkSBDfragment interacts with isolated spectrin in a saturable manner.In the control experiment the Talon� resin equilibrated in the testbuffer, was incubated with a corresponding amount of RITC-la-belled spectrin. The ‘‘bound” values were obtained by subtractingthe control values from those for the spectrin bound to the resinsaturated with AnkSBD. To optimise the assay, the binding wasperformed several times with different imidazole concentration(1–50 mM). We also found that 50 mM imidazole had no effecton His-tagged AnkSBD protein–resin interaction during proteinpurification under native conditions, i.e., in the absence of urea,thus it was used for unspecific spectrin–resin interaction wash stepafter protein-binding step assay. 50 mM imidazole in a test bufferwas used for wash step in control experiment. To exclude non-spe-cific binding of spectrin to the resin, additional experiment wasperformed. All experiments confirmed similar KD values. The bind-ing properties of erythroid and non-erythroid spectrin to theAnkSBD fragment are similar, as shown by the calculated KD values.Non-linear regression yielded a KD of 33 nM for erythroid spectrin
ncentrations of RITC-labelled erythrocyte (d) and brain (€) spectrins to the AnkSBDrifugation and washed with the test buffer, and the bound proteins were extracted
±standard deviation. (b) Standard curve example showing the linear dependence oft of erythrocyte and brain spectrin-binding values. The data shown represents the
on was used to fit the binding isotherm, KD and Bmax from the equation given in

Fig. 5. The inhibition of the binding of fluorescently labelled erythroid spectrin(0.04 lM) by increasing concentrations of unlabelled erythroid spectrin. The bind-ing conditions were as described in the legend to Fig. 4.
Fig. 6. HeLa cells transfected with GFP or GFP-AnkSBD. The control, GFP-transfec-ted cells (a) expressed the fluorescent protein continuously over the period of ob-servation (24 h). The protein was evenly distributed within the cell body (only asingle picture was chosen as an example of pEGFP-transfected cells, because theGFP-expressing cells looked very similar at different time points after transfection).In pEGFP–AnkSBD-transfected cells (b), the expressed fusion protein became agg-regated and the transfected cells changed their shapes. The cells rounded and de-tached from the cover slips; therefore, no pEGF–AnkSBD-transfected cells were seenafter 8 h of transfection. To compare the localisation of overexpressed protein withendogenous spectrin, the cells were immunostained with rabbit anti-spectrin ant-ibody followed by TRITC-labelled secondary antibody; (a) and (b) right panels.
162 A. Kolondra et al. / Protein Expression and Purification 60 (2008) 157–164
and 72 nM for non-erythroid spectrin (Fig 4a). The nanomolarrange of the equilibrium dissociation constant indicates thatAnkSBD-spectrin complexes of high affinity are formed. Bmax wasestimated at a 2:1 and a 4:1 ratio complex, respectively. Standardcurve showing correlation between fluorescence intensity and theamount of labelled spectrins (Fig. 4b) and Scatchard plots (correla-tion coefficients for the two plots: 0.97) (Fig. 4c) fitted well to thelinear characteristics indicating no cooperativity. The results of thebinding experiment in which unlabelled spectrin was used as acompetitor indicate strong inhibition, with a 10-fold excess ofunlabelled spectrin completely inhibiting the binding of labelledprotein to immobilised AnkSBD. This result indicates that thisbinding is not a result of labelling (Fig. 5).
HeLa cells transfection and fluorescent microscopy experiments
The transfection of HeLa cells with plasmid encoding EGFP-tagged AnkSBD indicated localisation of the expressed constructto the plasma membrane (Fig. 6). However, cells transfected withGFP–AnkSBD show low viability. The AnkSBD induces significantshape changes in the transfected cells compared to GFP onlyexpressing cells. Transfected cells became round and the expressedprotein was aggregated, while the control cells were flat and theEGFP protein was evenly distributed within the cells. Also, theexpression of EGFP alone is stable and easily seen even 24 h aftertransfection. In pEGFP–AnkSBD-transfected cells, the expressioncould be detected for 2–8 h after transfection. After 4 h of incuba-tion expressed protein was uniformly distributed near the cellmembrane although some small clusters were already seen inthe vicinity of the nuclei. The clustering increases over more then6 h of transfection when most of the AnkSBD–GFP forms immerseaggregates. Such aggregates were observed in almost all of thetransfected cells. After 12 h, no cells expressing EGFP–AnkSBDcould be detected. Immunostaining with anti-spectrin antibodyshowed that spectrin distribution dramatically changed comparedto that in EGFP only transfected cells (compare Fig. 6a and b) and tountransfected cells (not shown). Moreover, spectrin was to the rea-sonable extent redistributed to colocalise in the above-mentionedEGFP–AnkSBD aggregates. This may indicate that EGFP–AnkSBDprotein specifically binds endogenous spectrin and disrupts cyto-skeletal protein–protein interactions.
Discussion
Purification of ankyrin, whether from erythrocytes or other cellsand tissues is a time-consuming process, and the purified protein isusually difficult to store for an extended period of time withoutloss of activity as it is sensitive to cellular proteases contaminating
the preparation. It is also difficult to obtain a single ankyrin iso-form. The anion exchanger protein-binding domain of erythrocyteankyrin that was cloned and expressed facilitated important func-tional and structural studies carried out in Low’s laboratory [47].Our results show that it is possible to construct, clone and expressa functional spectrin-binding domain of ankyrinR.
Platt et al. [41] suggested that important for spectrin-bindingregion is located between amino acid residues 1136 and 1192. It

A. Kolondra et al. / Protein Expression and Purification 60 (2008) 157–164 163
should be noted however, that their bacterially expressed, GST-conjugated fragments showed much lower capacity to inhibit spec-trin-binding to the inside-out vesicles compared to the ‘‘native”62 kDa fragment. On the other hand, Davis and Bennett [39] sug-gested an importance of the entire region: residues 828–1386. Re-cent work from Bennett’s laboratory [38] on ankyrinB indicatedthat residues 966–1125 are responsible for this binding. These dataobtained by site-directed mutagenesis and two hybrid experi-ments were the starting point of our work which was focused onfragment between amino acid residues 909 and 1050 (see the se-quence comparison in Fig. 1). Moreover, expressed ankyrin frag-ment binds spectrin with similar affinity to the cloned andexpressed full-length SBD domain encompassing residues 798–1390 obtained recently in our laboratory (not shown).
Theoretical structure of AnkSBD construct was predicted withRobetta Full-chain Protein Structure Prediction Server (http://rob-etta.bakerlab.org). This putative structure contains ZU5 domainpresented in Fig. 7 shows an antiparallel b-strand structure whichmay play main role in ankyrin–spectrin interactions.
The applied strategy allowed us to purify milligram quantitiesof bacterially expressed protein which is stable at pH 5.5 and spe-cifically binds spectrins. Immobilised on Co2+ affinity resin AnkSBDbound purified red blood cell and brain spectrin with high affinity(KD in the nanomolar range) at an apparent 2:1 and 4:1 molar ratio,respectively. The increased peptide-to-spectrin molar ratio (thereis one ankyrin-binding site in the red blood cell spectrin dimerand two sites in one tetrameric non-erythroid spectrin molecule)may be due to the assay having been carried out on an immobilisedprotein, thus restricting its mobility and availability for the high-molecular weight ligand. Obtained KD values of AnkSBD–spectrincomplexes highly corresponded to values obtained previously forinteractions between spectrin and ankyrins with a value of �10–20 nM (reviewed in [8]). This binding was strongly inhibited byunlabelled spectrin; therefore, it was not the result of the labellingprocedure. Our experiments on binding the labelled ankyrin byimmobilised b-spectrin fragment corresponding to the ankyrin-binding domain gave the value close to 200 nM [45]. Also, the inhi-bition of spectrin–phospholipid monolayer binding experiments
Fig. 7. AnkSBD protein prediction structure performed with Robetta Full-chainProtein Structure Prediction Server (http://robetta.bakerlab.org).
carried out in a similar way to those with purified ankyrin andspectrin–lipid-binding site [45,46] suggest functional propertiesfor the pRSET vector expressed protein (not shown). As this domainis conservative among ankyrins, and the experiment indicated theability to bind brain spectrin, it was anticipated to also be func-tional in non-erythroid system in vivo. In the transfected HeLa cellsexpressing AnkSBD–GFP protein, large aggregates that form insidecells were observed. Immunostaining with anti aIIbII-spectrin anti-bodies revealed that AnkSBD–GFP colocalises with spectrin andmoreover, spectrin is clustered to these aggregates. It should alsobe noted that no such clusters were observed in GFP only express-ing cells that were used as a control. It was also surprising to notethat the number of AnkSBD–GFP expressing cells decreased dra-matically in time. This was probably caused by enhanced mortalityof these cells due to the presence of AnkSBD product. As AnkSBDproduct most probably acts as a competitor for endogenous anky-rin in spectrin-binding activity. We can only hypothesise thatbreaking of the ‘‘native” ankyrin–spectrin connections underliescell death. Whether this event concerns mechanical membraneproperties or apoptotic signal (ankyrin possesses the death-do-main) should be a subject of further studies. The fact is that after12 h of transfection no AnkSBD–GFP expressing cells were ob-served. Large, green fluorescent cell-size-like bodies devoid of nu-cleus, which we treat as cell debris and large nuclei with smallcytoplasmic space were occasionally seen which might suggestthat expression of AnkSBD fragment caused expelling nucleus fromthe cell.
In hereditary spherocytosis, the inappropriate binding of spec-trin and ankyrin is highly correlated [48,49]; therefore, under-standing the conditions and requirements for this binding isdemanded. Our construct binds both erythroid and non-erythroidspectrin with high affinity. The protein is easy to purify and handle,so the obtained construct allows functional studies on the spectrin-binding domain of this ankyrin isoform either in vitro or in vivo.This method allows expression of this fragment in large quantitieswhat may allow the structural studies. Together with the b-spec-trin ankyrin-binding domain constructs obtained in our laboratory[45], it is a promising tool to study spectrin–ankyrin interactions.In addition, this construct is an easy matrix for site-directed muta-genesis studies, which in the case of AnkSBD have yet to be carriedout.
Acknowledgments
This study was supported by Grant No. 2P04 021 27 from thePolish Ministry of Science and Higher Education and Grant No.2750/W/WBT/07 from the University of Wroclaw.
References
[1] P.J. Mohler, A.O. Gramolini, V. Bennett, Ankyrins, J. Cell Sci. 115 (2002) 1565–1566.
[2] V. Bennett, A.J. Baines, Spectrin and ankyrin-based pathways: metazoaninventions for integrating cells into tissues, Physiol. Rev. 81 (2001) 1353–1392.
[3] V. Nicolas, I. Mouro-Chanteloup, C. Lopez, P. Gane, A. Gimm, N. Mohandas, J.P.Cartron, C. Le Van Kim, Y. Colin, Functional interaction between Rh proteinsand the spectrin-based skeleton in erythroid and epithelial cells, Transfus. Clin.Biol. 13 (2006) 23–28.
[4] S. Lambert, H. Yu, J.T. Prchal, J. Lawler, P. Ruff, D. Speicher, M.C. Cheung, Y.W.Kan, J. Palek, cDNA sequence for human erythrocyte ankyrin, Proc. Natl. Acad.Sci. USA 87 (1990) 1730–1734.
[5] S.E. Lux, K.M. John, V. Bennett, Analysis of cDNA for human erythrocyte ankyrinindicates a repeated structure with homology to tissue-differentiation andcell-cycle control proteins, Nature 344 (1990) 36–42.
[6] E. Otto, M. Kunimoto, T. McLaughlin, V. Bennett, Isolation and characterizationof cDNAs encoding human brain ankyrins reveal a family of alternativelyspliced genes, J. Cell. Biol 114 (1991) 241–253.
[7] E. Kordeli, S. Lambert, V. Bennett, AnkyrinG. A new ankyrin gene with neural-specific isoforms localized at the axonal initial segment and node of Ranvier, J.Biol. Chem. 270 (1995) 2352–2359.

164 A. Kolondra et al. / Protein Expression and Purification 60 (2008) 157–164
[8] A. Hryniewicz-Jankowska, A. Czogalla, E. Bok, A.F. Sikorski, Ankyrins,multifunctional proteins involved in many cellular pathways, FoliaHistochem. Cytobiol. 40 (2002) 239–249.
[9] P. Michaely, V. Bennett, The membrane-binding domain of ankyrin containsfour independently folded subdomains, each comprised of six ankyrin repeats,J. Biol. Chem. 268 (1993) 22703–22709.
[10] L. Davis, S.E. Lux, V. Bennett, Mapping the ankyrin-binding site of the humanerythrocyte anion exchanger, J. Biol. Chem. 264 (1989) 9665–9672.
[11] P. Michaely, V. Bennett, The ANK repeat: a ubiquitous motif involved inmacromolecular recognition, Trends Cell Biol. 2 (1992) 127–129.
[12] L. Breeden, K. Nasmyth, Similarity between cell-cycle genes of budding yeastand fission yeast and the Notch gene of Drosophila, Nature 329 (1987)651–654.
[13] S. Ghosh, A. Gifford, L. Riviere, P. Tempst, G. Nolan, D. Baltimore, Cloning of thep50 DNA binding subunit of NF-kappa B: homology to rel and dorsal, Cell 62(1990) 1019–1029.
[14] M. Kieran, V. Blank, F. Logeat, J. Vandekerckhove, F. Lottspeich, O. Le Bail, M.Urban, P. Kourlisky, P. Baeuerle, A. Israel, The DNA binding subunit of NF-kappa B is identical to factor KBF1 and homologous to the rel oncogeneproduct, Cell 62 (1990) 1007–1018.
[15] S. Haskill, A. Beg, S.M. Tompkins, J. Morris, A. Yurochko, A. Sampson-Johannes,K. Mondal, P. Ralph, A. Baldin, Characterization of an immediate-early geneinduced in adherent monocytes that encodes I kappa B-like activity, Cell 65(1991) 1281–1289.
[16] N. Davis, S. Ghosh, D. Simmons, P. Tempst, H. Liou, D. Baltimore, H. Bose, Rel-associated pp40: an inhibitor of the rel family of transcription factors, Science253 (1991) 1268–1271.
[17] J. Inoue, L. Kerr, D. Rashid, N. Davis, H. Bose, I. Verma, Direct association ofpp40/I kappa B beta with rel/NF-kappa B transcription factors: role of ankyrinrepeats in the inhibition of DNA binding activity, Proc. Natl. Acad. Sci. USA 89(1992) 4333–4337.
[18] H. Ohno, G. Takimoto, T. McKeithan, The candidate proto-oncogene bcl-3 isrelated to genes implicated in cell lineage determination and cell cycle control,Cell 60 (1990) 991–997.
[19] L.K. Mosavi, T.J. Cammett, D.C. Desrosiers, Z.Y. Peng, The ankyrin repeat asmolecular architecture for protein recognition, Prot. Sci. 13 (2004) 1435–1448.
[20] J.Q. Davis, V. Bennett, The anion exchanger and Na + K(+)-ATPase interact withdistinct sites on ankyrin in in vitro assays, J. Biol. Chem. 265 (1990)17252–17256.
[21] S. Thevananther, A.H. Kolli, P. Devarajan, Identification of a novel ankyrinisoform (AnkG190) in kidney and lung that associates with the plasmamembrane and binds alpha-Na, K-ATPase, J. Biol. Chem. 273 (1998)23952–23958.
[22] F. Festy, J.C. Robert, R. Brausseur, A. Thomas, Interaction between the N-terminal domain of gastric H,K-ATPase and the spectrin binding domain ofankyrin III, J. Biol. Chem. 276 (2001) 7721–7726.
[23] J.Q. Davis, T. McLaughlin, V. Bennett, Ankyrin-binding proteins related tonervous system cell adhesion molecules: candidates to providetransmembrane and intercellular connections in adult brain, J. Cell. Biol 121(1993) 121–133.
[24] J.Q. Davis, V. Bennett, Ankyrin binding activity shared by the neurofascin/L1/NrCAM family of nervous system cell adhesion molecules, J. Biol. Chem. 269(1994) 27163–27166.
[25] P. Michaely, V. Bennett, The ANK repeats of erythrocyte ankyrin form twodistinct but cooperative binding sites for the erythrocyte anion exchanger, J.Biol. Chem. 270 (1995) 22050–22057.
[26] J.Q. Davis, V. Bennett, Brain ankyrin. Purification of a 72,000 Mr spectrin-binding domain, J. Biol. Chem. 259 (1983) 1874–1881.
[27] L.H. Davis, E. Otto, V. Bennett, Specific 33-residue repeat(s) of erythrocyteankyrin associate with the anion exchanger, J. Biol. Chem. 266 (1991) 11163–11169.
[28] C.W. Morgans, R.R. Kopito, Association of the brain anion exchanger, AE3, withthe repeat domain of ankyrin, J. Cell Sci. 105 (1993) 1137–1142.
[29] Y. Srinivasan, M. Lewallen, K. Angelides, Mapping the binding site on ankyrinfor the voltage-dependent sodium channel from brain, J. Biol. Chem. 267(1992) 7483–7489.
[30] P. Michaely, A. Kamal, R.G. Anderson, V. Bennett, A requirement for ankyrinbinding to clathrin during coated pit budding, J. Biol. Chem. 274 (1999) 35908–35913.
[31] V. Nicolas, C. Le Van Kim, P. Gane, C. Birkenmeier, J.P. Carton, Y. Colin, I. Mouro-Chanteloup, Rh-RhAG/ankyrin-R, a new interaction site between themembrane bilayer and the red cell skeleton, is impaired by Rh(null)-associated mutation, J. Biol. Chem. 278 (2003) 25526–25533.
[32] P.J. Mohler, J.Q. Davis, L.H. Davis, J.A. Hoffman, P. Michaely, V. Bennett, Inositol1,4,5-trisphosphate receptor localization and stability in neonatalcardiomyocytes requires interaction with ankyrin-B, J. Biol. Chem. 279(2004) 12980–12987.
[33] D. Pradhan, J.S. Morrow, The spectrin–ankyrin skeleton controls CD45 surfacedisplay and interleukin-2 production, Immunity 17 (2002) 303–315.
[34] P. Michaely, D.R. Tomchick, M. Machius, R.G.W. Anderson, Crystal structure ofa 12 ANK repeat stack from human ankyrinR, EMBO J. 21 (2002) 6387–6396.
[35] K.M. Abdi, P.J. Mohler, J.Q. Davis, V. Bennett, Isoform specificity of ankyrin-B: asite in the divergent C-terminal domain is required for intramolecularassociation, J. Biol. Chem. 281 (2006) 5741–5749.
[36] P.J. Mohler, A.O. Gramolini, V. Bennett, The ankyrin-B C-terminal domaindetermines activity of ankyrin-B/G chimeras in rescue of abnormal inositol1,4,5-trisphosphate and ryanodine receptor distribution in ankyrin-B (�/�)neonatal cardiomyocytes, J. Biol. Chem. 277 (2002) 10599–10607.
[37] J.Q. Davis, V. Bennett, Brain ankyrin. A membrane-associated protein withbinding sites for spectrin, tubulin, and the cytoplasmic domain of theerythrocyte anion channel, J. Biol. Chem. 259 (1984) 13550–13559.
[38] P.J. Mohler, W. Yoon, V. Bennett, Ankyrin-B targets beta2-spectrin to anintracellular compartment in neonatal cardiomyocytes, J. Biol. Chem. 279(2004) 40185–40193.
[39] L.H. Davis, V. Bennett, Mapping the binding sites of human erythrocyteankyrin for the anion exchanger and spectrin, J. Biol. Chem. 265 (1990) 10589–10596.
[40] J. Zhang, L.G. Xu, K.J. Han, H.B. Shu, Identification of a ZU5 and death domain-containing inhibitor of NF-jB, J. Biol. Chem. 279 (2004) 17819–17825.
[41] O.S. Platt, S.E. Lux, J.F. Falcone, A highly conserved region of human erythrocyteankyrin contains the capacity to bind spectrin, J. Biol. Chem. 268 (1993)24421–24426.
[42] K.M. Downey, J.J. Byrnes, B.S. Jurmark, A.G. So, Reticulocyte RNA-dependentRNA polymerase, Proc. Natl. Acad. Sci. USA 70 (1973) 3400–3404.
[43] K. Michalak, M. Bobrowska, A.F. Sikorski, Interaction of bovine erythrocytespectrin with aminophospholipid liposomes, Gen. Physiol. Biophys. 12 (1993)163–170.
[44] V. Bennett, A.J. Baines, J. Davis, Purification of brain analogs of red blood cellsmembrane skeletal proteins: ankyrin, protein 4.1 (synapsin), spectrin, andspectrin subunits, Methods Enzymol. 134 (1986) 55–68.
[45] A. Hryniewicz-Jankowska, E. Bok, P. Dubielecka, A. Chorzalska, W. Diakowski,A. Jezierski, M. Lisowski, A.F. Sikorski, Mapping of an ankyrin-sensitive,phosphatidylethanol-amine/phosphatidylcholine mono- and bilayer bindingsite in erythroid beta-spectrin, Biochem. J. 382 (2004) 677–685.
[46] A. Czogalla, A.R. Jaszewski, W. Diakowski, E. Bok, A. Jezierski, A.F. Sikorski,Structural insight into an ankyrin-sensitive lipid-binding site of erythroidbeta-spectrin, Mol. Membr. Biol. 24 (2007) 215–224.
[47] C.C. Wang, R. Moriyama, C.R. Lombardo, P.S. Low, Partial characterization ofthe cytoplasmic domain of human kidney band 3, J. Biol. Chem. 270 (1995)17892–17897.
[48] S.W. Eber, J.M. Gonzales, M.L. Lux, A.L. Scarpa, W.T. Tse, M. Dornwell, J.Herbers, W. Kugler, R. Ozcan, A. Pekrun, P.G. Gallagher, W. Schroter, B.G.Forget, S.E. Lux, Ankyrin-1 mutations are a major cause of dominant andrecessive hereditary spherocytosis, Nat. Genet. 13 (1996) 214–218.
[49] M.J. Broderick, S.J. Winder, Spectrin, alpha-actinin, and dystrophin, Adv. Prot.Chem. 70 (2005) 203–246.
Copyright © 2022 FDOKUMEN




















