
Bacterial peptidoglycan binds to tubulin
10
Bacterial peptidoglycan binds to tubulin Roman Dziarski a ; *, Mark M. Rasenick b , Dipika Gupta a a Northwest Center for Medical Education, Indiana University School of Medicine, 3400 Broadway, Gary, IN 46408, USA b Department of Physiology and Biophysics, University of Illinois, College of Medicine, Chicago, IL 60612, USA Received 21 June 2000; received in revised form 29 August 2000; accepted 31 August 2000 Abstract A search for cellular binding proteins for peptidoglycan (PGN), a CD14- and TLR2-dependent macrophage activator from Gram- positive bacteria, using PGN-affinity chromatography and N-terminal micro-sequencing, revealed that tubulin was a major PGN-binding protein in mouse macrophages. Tubulin also co-eluted with PGN from anti-PGN vancomycin affinity column and bound to PGN coupled to agarose. Tubulin-PGN binding was preferential under the conditions that promote tubulin polymerization, required macromolecular PGN, was competitively inhibited by soluble PGN and tubulin, did not require microtubule-associated proteins, and had an affinity of 100^ 150 nM. By contrast, binding of tubulin to lipopolysaccharide (LPS) had 2^3 times lower affinity, faster kinetics of binding, and showed positive cooperativity. PGN enhanced tubulin polymerization in the presence of 4 M glycerol, but in the absence of glycerol, both PGN and LPS decreased microtubule polymerization. These results indicate that tubulin is a major PGN-binding protein and that PGN modulates tubulin polymerization. ß 2000 Elsevier Science B.V. All rights reserved. Keywords : Lipopolysaccharide ; Microtubule ; Peptidoglycan ; Tubulin 1. Introduction Peptidoglycan (PGN) is present in the cell walls of all bacteria and is the major constituent of the cell walls of Gram-positive bacteria. PGN can reproduce most of the major clinical manifestations of bacterial infections, in- cluding fever, in£ammation, hypotension, leukocytosis, sleepiness, decreased appetite, malaise, and arthritis [1^ 3]. These clinical manifestations are induced by PGN in- directly, through the activation of macrophages and stim- ulation of secretion of mediators of in£ammation, primar- ily cytokines and chemokines [1^3]. The ¢rst step in macrophage activation by PGN is the binding of PGN to its speci¢c receptor, membrane CD14 [2^5]. CD14 also serves as the receptor for other bacterial macrophage activators, including lipopolysaccharide (LPS or endotoxin) from Gram-negative bacteria, lipoteichoic acid from Gram-positive bacteria, lipoproteins from spi- rochetes, lipoarabinomannan from mycobacteria, and others [2,3]. The second step in macrophage activation is transmission of the activating signal from the receptor into the cell. However, CD14 is a glycosylphosphatidylinositol- linked rather than a transmembrane molecule and by itself CD14 cannot transmit an activating signal into the cell through the membrane. Therefore, Toll-like receptors, TLR2 and TLR4, serve as the cell-activating co-receptors for Gram-positive bacteria and PGN [6,7], and Gram-neg- ative bacteria and LPS [8^11], respectively. TLRs, how- ever, do not seem to bind these bacterial products with su/cient a/nity that could account for cell activation ([6,12]; R. Dziarski, C.J. Kirschning, unpublished). There- fore, TLRs appear to function as signal-transducing rather than bacterial cell wall-binding molecules [12]. While looking for other molecules in mammalian cells that bind bacterial PGN, we discovered that PGN binds to tubulin, which we report in this paper. Tubulin is a heter- odimer consisting of K and L chains, which forms micro- tubules and is present in all eucaryotic cells [13]. Polymer- ization of tubulin into microtubules requires GTP, and in the cell, most microtubules undergo continuous rapid as- sembly and disassembly. Microtubules participate in mor- phogenesis, cell division, and transport of vesicles and or- 0304-4165 / 00 / $ ^ see front matter ß 2000 Elsevier Science B.V. All rights reserved. PII:S0304-4165(00)00137-9 Abbreviations : PGN, peptidoglycan ; HBSS, Hank’s balanced salt so- lution ; LPS, lipopolysaccharide ; ReLPS, LPS from Salmonella minnesota Re mutant 595; MDP, muramyl dipeptide (MurNAc-L-Ala-D-isoglut- amine); MAP, microtubule-associated proteins ; sPGN, soluble PGN; TLR, Toll-like receptor * Corresponding author. Fax: +1-219-980-6566; E-mail : [email protected] Biochimica et Biophysica Acta 1524 (2000) 17^26 www.elsevier.com/locate/bba
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Bacterial peptidoglycan binds to tubulin

Bacterial peptidoglycan binds to tubulin
Roman Dziarski a;*, Mark M. Rasenick b, Dipika Gupta a
a Northwest Center for Medical Education, Indiana University School of Medicine, 3400 Broadway, Gary, IN 46408, USAb Department of Physiology and Biophysics, University of Illinois, College of Medicine, Chicago, IL 60612, USA
Received 21 June 2000; received in revised form 29 August 2000; accepted 31 August 2000
Abstract
A search for cellular binding proteins for peptidoglycan (PGN), a CD14- and TLR2-dependent macrophage activator from Gram-positive bacteria, using PGN-affinity chromatography and N-terminal micro-sequencing, revealed that tubulin was a major PGN-bindingprotein in mouse macrophages. Tubulin also co-eluted with PGN from anti-PGN vancomycin affinity column and bound to PGN coupledto agarose. Tubulin-PGN binding was preferential under the conditions that promote tubulin polymerization, required macromolecularPGN, was competitively inhibited by soluble PGN and tubulin, did not require microtubule-associated proteins, and had an affinity of 100^150 nM. By contrast, binding of tubulin to lipopolysaccharide (LPS) had 2^3 times lower affinity, faster kinetics of binding, and showedpositive cooperativity. PGN enhanced tubulin polymerization in the presence of 4 M glycerol, but in the absence of glycerol, both PGN andLPS decreased microtubule polymerization. These results indicate that tubulin is a major PGN-binding protein and that PGN modulatestubulin polymerization. ß 2000 Elsevier Science B.V. All rights reserved.
Keywords: Lipopolysaccharide; Microtubule; Peptidoglycan; Tubulin
1. Introduction
Peptidoglycan (PGN) is present in the cell walls of allbacteria and is the major constituent of the cell walls ofGram-positive bacteria. PGN can reproduce most of themajor clinical manifestations of bacterial infections, in-cluding fever, in£ammation, hypotension, leukocytosis,sleepiness, decreased appetite, malaise, and arthritis [1^3]. These clinical manifestations are induced by PGN in-directly, through the activation of macrophages and stim-ulation of secretion of mediators of in£ammation, primar-ily cytokines and chemokines [1^3].
The ¢rst step in macrophage activation by PGN is thebinding of PGN to its speci¢c receptor, membrane CD14[2^5]. CD14 also serves as the receptor for other bacterialmacrophage activators, including lipopolysaccharide (LPSor endotoxin) from Gram-negative bacteria, lipoteichoic
acid from Gram-positive bacteria, lipoproteins from spi-rochetes, lipoarabinomannan from mycobacteria, andothers [2,3]. The second step in macrophage activation istransmission of the activating signal from the receptor intothe cell. However, CD14 is a glycosylphosphatidylinositol-linked rather than a transmembrane molecule and by itselfCD14 cannot transmit an activating signal into the cellthrough the membrane. Therefore, Toll-like receptors,TLR2 and TLR4, serve as the cell-activating co-receptorsfor Gram-positive bacteria and PGN [6,7], and Gram-neg-ative bacteria and LPS [8^11], respectively. TLRs, how-ever, do not seem to bind these bacterial products withsu¤cient a¤nity that could account for cell activation([6,12]; R. Dziarski, C.J. Kirschning, unpublished). There-fore, TLRs appear to function as signal-transducing ratherthan bacterial cell wall-binding molecules [12].
While looking for other molecules in mammalian cellsthat bind bacterial PGN, we discovered that PGN binds totubulin, which we report in this paper. Tubulin is a heter-odimer consisting of K and L chains, which forms micro-tubules and is present in all eucaryotic cells [13]. Polymer-ization of tubulin into microtubules requires GTP, and inthe cell, most microtubules undergo continuous rapid as-sembly and disassembly. Microtubules participate in mor-phogenesis, cell division, and transport of vesicles and or-
0304-4165 / 00 / $ ^ see front matter ß 2000 Elsevier Science B.V. All rights reserved.PII: S 0 3 0 4 - 4 1 6 5 ( 0 0 ) 0 0 1 3 7 - 9
Abbreviations: PGN, peptidoglycan; HBSS, Hank's balanced salt so-lution; LPS, lipopolysaccharide ; ReLPS, LPS from Salmonella minnesotaRe mutant 595; MDP, muramyl dipeptide (MurNAc-L-Ala-D-isoglut-amine); MAP, microtubule-associated proteins; sPGN, soluble PGN;TLR, Toll-like receptor
* Corresponding author. Fax: +1-219-980-6566;E-mail : [email protected]
BBAGEN 25093 1-11-00
Biochimica et Biophysica Acta 1524 (2000) 17^26
www.elsevier.com/locate/bba

ganelles [13]. Numerous signal transduction molecules arealso associated with microtubules, including mitogen-acti-vated protein kinases [14,15] and G proteins [16,17]. Tu-bulin, therefore, may also participate in signal transduc-tion events during cell activation.
Interest in the interaction of bacterial LPS with tubulinstarted when microtubule-stabilizing agent, taxol, wasfound to mimic LPS-induced activation of macrophages,and when this e¡ect was not observed in LPS-hyporespon-sive C3H/HeJ mice [18^20]. Moreover, the genes control-ling the responsiveness to taxol and LPS are closely linked[18], and LPS antagonists block the responsiveness to tax-ol [21,22]. LPS also binds to tubulin [23,24] and micro-tubules [24], and all LPS-stimulated mitogen-activatedprotein kinases are associated with microtubules [15]. Fur-thermore, during cell activation, LPS is rapidly internal-ized and transported to the perinuclear Golgi apparatus[25,26]. On route to the Golgi complex or after reaching it,LPS could either intercalate into vesicle membranes orleave the vesicles for the cytoplasm [26], which wouldgive it an opportunity to interact with microtubules. Sinceboth PGN and LPS activate cells through the same CD14and TLR receptors, we hypothesized that both PGN andLPS may interect with tubulin during cell activation andintracellular transport.
Therefore, the objectives of this study were to: (i) de-termine if PGN binds to tubulin, (ii) characterize PGNbinding to tubulin in detail, (iii) compare PGN and LPSbinding to tubulin, including determining the a¤nity ofbinding, and the requirement for tubulin polymerizationand for microtubule-associated proteins, and (iv) deter-mine the e¡ects of PGN and LPS on tubulin polymeriza-tion in vitro.
2. Materials and methods
2.1. Materials
Tubulin containing microtubule-associated proteins(MAP) was puri¢ed from sheep brains by two cycles ofassembly and disassembly [17], and tubulin free of MAPwas puri¢ed from tubulin by phosphocellulose chromatog-raphy [17]. For some experiments (co-elution from vanco-mycin a¤nity columns, see below), tubulin was puri¢edfrom bovine brain by temperature-dependent assembly-disassembly cycles [27] and contained 15% MAP (obtainedfrom Sigma, St. Louis, MO).
Soluble PGN (sPGN) from Staphylococcus aureus, apolymeric non-cross-linked PGN of approximate averageMr = 125 000, was puri¢ed by vancomycin a¤nity chroma-tography [28]. For some experiments, sPGN was biosyn-thetically labeled with [14C]alanine [28]. Insoluble PGNwas isolated from the cell walls of S. aureus 845 and so-nicated [28]. By quantitative chemical analysis, both sPGN
and insoluble PGN had amino acid and amino sugar com-position characteristic of S. aureus PGN [28], and con-tained 6 24 pg endotoxin mg31 determined by the Limu-lus lysate assay [28,29]. For some experiments, sPGN wasdigested with lysostaphin [5].
Synthetic analogs of PGN fragments, PGN pentapep-tide (L-Ala-D-isoglutaminyl-L-Lys-D-Ala-D-Ala) and mur-amyl dipeptide (MurNAc-L-Ala-D-isoglutamine, MDP)were from Sigma, and a disaccharide-dipeptide (GlcNAc-L1-4-MDP) was from Calbiochem (La Jolla, CA). No sig-ni¢cant endotoxin contamination of these preparationswas detected (91 ng endotoxin mg31) [5].
LPS from Salmonella minnesota Re 595 (ReLPS,Mr = 2000^3000), a mutant devoid of O-polysaccharideand most of the core-polysaccharide, is a minimal natu-rally occurring endotoxic structure of LPS. It was ob-tained by phenol^chloroform^petroleum ether extractionby Sigma, and its purity was analyzed as before [5]. Puri-¢ed smooth (complete) LPS, obtained from Escherichiacoli O113 by phenol-water extraction (re¢ned endotoxinstandard, approximate average Mr = 15 000) was obtainedfrom Ribi Immunochem Research (Hamilton, MT). Taxol(paclitaxel from Taxus brevifolia) and all other chemicalswere obtained from Sigma, unless otherwise indicated.
2.2. Cells, cell lysates, and cell membranes
For the cell activation assays, mouse macrophageRAW264.7 cells were cultured in DMEM medium with10% fetal calf serum [4]. For obtaining cell lysates andcell membranes for the binding assays, RAW264.7 cellsadapted to growth in serum-free HL-1 medium [30] wereused, to avoid contamination with potential PGN-bindingproteins from serum. To obtain cell lysates, 2U109 cellswere lysed at 20³C in 40 ml of Hanks Balanced Salt So-lution (HBSS) with 1.2% octylglucoside, 5% glycerol,1 mM Hepes, 230 Wg ml31 PMSF, and 25 Wg ml31 apro-tinin, and undissolved cell fragments were removed by 30min centrifugation at 3000Ug, 20³C. To obtain cell mem-branes, 3.5U109 cells were suspended at 4³C in 60 ml of25 mM Tris^HCl bu¡er (pH 7.4) with 5 mM MgCl2, 1 mMEGTA, 230 Wg ml31 PMSF, and 25 Wg ml31 aprotinin,and homogenized for 20 s in a Polytrone (Brinkman In-struments, Westburg, NY) homogenizer on ice, until 75^85% of cells were broken with no visible damage to nuclei[31]. Unbroken cells, nuclei, and debris were removed by10 min centrifugation at 500Ug at 4³C, and the mem-branes were sedimented by 60 min centrifugation at240 000Ug at 4³C [31]. Membranes were suspended bysonication on ice in 0.2 M phosphate bu¡er (pH 7.4)with 5% glycerol, 425 Wg ml31 PMSF, and 45 Wg ml31
aprotinin, solubilized in 1% octylglucoside, and dilutedto 80 ml with HBSS with 0.1% octylglucoside and 1%glycerol. Undissolved membrane aggregates were removedby centrifugation at 25 000Ug at 4³C.
BBAGEN 25093 1-11-00
R. Dziarski et al. / Biochimica et Biophysica Acta 1524 (2000) 17^2618

2.3. PGN-a¤nity chromatography and N-terminalsequencing
In the ¢rst approach, cell lysate (40 ml) was diluted with200 ml of HBSS with 0.2% octylglucoside and 1% glycerol,mixed with 4 ml of sPGN coupled to agarose [5] for 18 hat 4³C, and poured into a column. The sPGN^agarose waswashed with 200 ml of HBSS with 0.2% octylglucosideand 1% glycerol, and agarose-bound proteins were elutedwith 9 M urea with 2% NP40 and 5 mg ml31 DTT at70³C, precipitated with 90% ethanol at 320³C for 18 h,sedimented by centrifugation at 3000Ug, and dissolved inpolyacrylamide gel electrophoresis (PAGE) sample bu¡erwith 1 N NaCl, 2% sodium dodecyl sulfate (SDS), and 1%2-mercaptoethanol. In the second approach, cell lysatesdiluted as above were centrifuged at 4000Ug for 30 min,the supernatants were mixed with 4 mg insoluble PGN for24 h at 4³C, the PGN was sedimented by centrifugation at3000Ug for 20 min at 4³C, the sediment was washed withHBSS with 0.2% octylglucoside and 1% glycerol, andPGN-bound proteins were eluted with 9 M urea with2% NP40 and DTT, precipitated with ethanol, and dis-solved in PAGE bu¡er as described above. In the thirdapproach, dissolved membranes were mixed with 1.2 mginsoluble PGN for 6 h at 4³C, PGN was sedimented at4000Ug, washed with HBSS with 0.2% octylglucoside and1% glycerol, and PGN-bound proteins were eluted anddissolved as in the second approach.
Eluted PGN-binding proteins were subjected to SDS^PAGE on 11% gels, blotted onto a PVDF Immobilon P(Millipore, Bedford, MA), and stained with Coomassieblue. The bands were cut out and subjected to the auto-matic N-terminal micro-sequencing at the BiochemistryBiotechnology Facility at Indiana University School ofMedicine, Indianapolis, IN. In some experiments, theeluted PGN-binding proteins were separated by two-di-mensional electrophoresis (isoelectric focusing and SDS^PAGE) [29], blotted onto Immobilon P, and sequenced asabove.
2.4. Vancomycin-a¤nity chromatography and Westernblots
Vancomycin coupled to agarose was used for sPGNa¤nity chromatography [28]. Vancomycin speci¢callybinds to the D-Ala-D-Ala part of PGN, the structureuniquely found in non-cross-linked PGN and biosyntheticPGN precursors [28]. Tubulin (20 Wg/100 Wl, from Sigma)was incubated either in the polymerizing bu¡er (0.05 Mphosphate or 0.1 M PIPES (pH 6.7) with 1 mM EGTA,5 mM MgCl2, and 1 mM GTP) with 20 WM taxol for 20min at 37³C, or in the de-polymerizing bu¡er (0.05 Mphosphate (pH 7.2) with 5 mM CaCl2) for 1 h at 4³C,followed by 1 h incubation with 20 Wg sPGN (labeledwith [14C]alanine) at 30³C for polymerizing conditions or at20³C for de-polymerizing conditions. Tubulin polymeriza-
tion is favored at 37³C in the presence of GTP and Mg2�
at pH6 7 with no Ca2�, and tubulin de-polymerizationoccurs at 4³C in the presence of Ca2� at pHs 7. Thecontrols included sPGN alone or tubulin alone. Vancomy-cin^agarose columns (1 ml) were washed with either thepolymerizing or de-polymerizing bu¡er with 0.1% Tween-20 at 20³C, the mixture of tubulin with sPGN in eitherpolymerizing or de-polymerizing bu¡er was loaded on thecolumn, the columns were washed with 12 ml of either thepolymerizing or de-polymerizing bu¡er with 0.1% Tween-20, and the sPGN was eluted from the columns withNH4OH in H2O (pH 10) with 0.1% Tween-20, and col-lected into 0.6-ml fractions. The concentration of sPGN inthe fractions was monitored by scintillation counting of14C and by absorbance at 218 nm. To detect the presenceof tubulin, the fractions were concentrated by evaporationat 52³C, and subjected to SDS^PAGE on 11% gels, trans-ferred to Immobilon P, and probed with a mixture ofmouse anti-tubulin-K (clone B-5-1-2) and anti-tubulin-L(clone JDR.3B8) monoclonal antibodies (mAbs) (both re-acting with tubulins from a variety of species, includingmouse and bovine, obtained from Sigma), followed byincubation with peroxidase-labeled anti-mouse IgG, anddetection by the enhanced chemiluminescence (ECL fromAmersham, Chicago Heights, IL). The speci¢cities andselectivities of the mAbs were con¢rmed using Westernblots with puri¢ed tubulin and crude cell lysates (seeFig. 2B below).
In some experiments, cell lysates were used instead oftubulin. 15U106 RAW264.7 cells, grown in serum-freeHL-1 medium, were washed twice with HBSS withoutCa2� and Mg2�, and lysed by sonication in 200 Wl ofpolymerizing bu¡er with 1.2% NP-40, 30 Wg ml31 aproti-nin and 250 Wg ml31 PMSF on ice. Insoluble material wasremoved by centrifugation at 12 000Ug at 4³C, and thesupernatant was incubated with 1 mM GTP and 20 WMtaxol at 37³C for 1.5 h, followed by incubation with sPGNand vancomycin a¤nity chromatography under polymer-izing conditions, and by detection of sPGN and tubulin inthe fractions, as described above. The controls includedsPGN alone and cell lysate alone.
2.5. Binding of 125I-labeled tubulin to sPGN^agarose andReLPS^agarose
Tubulin (100 Wg) and MAP-free tubulin (50 Wg) werelabeled with 2 mCi 125I using pre-coated iodogen tubes(from Pierce, Rockford, IL) and checked for functionalintegrity [16]. Unbound 125I was removed by dialysis at4³C against 100 mM PIPES bu¡er (pH 6.9) with 1 mMMgCl2, 1 mM EDTA, and 1 mM GTP. The aliquots ofthe iodinated preparations were stored at 380³C andfreshly thawed out for each experiment. The speci¢c activ-ities were 220^240 Ci mmol31 for tubulin, and 525^545 Cimmol31 for MAP-free tubulin. Both 125I-labeled tubulinsyielded one 50 kDa band detected by autoradiography on
BBAGEN 25093 1-11-00
R. Dziarski et al. / Biochimica et Biophysica Acta 1524 (2000) 17^26 19

SDS^PAGE gels, that co-migrated with a Coomassie blue-stained band of unlabeled tubulin, con¢rming iodinationof tubulin and not any potential contaminants, and min-imal iodination of MAPs (see Fig. 3B below).
sPGN, ReLPS, smooth LPS, GlcNAc-MDP, MDP, orPGN-pentapeptide were coupled to 40^165 Wm agarosebeads [5], and the binding of iodinated tubulin to theseagarose-immobilized preparations was performed as de-scribed before for the binding of 32P-sCD14 [5]. For thestandard binding assay under polymerizing conditions,[125I]tubulin was pre-incubated in 10 Wl of the polymeriz-ing bu¡er with 1 mM GTP at 37³C for 20 min, and thendiluted to 60 Wl to yield ¢nal concentration of 1 Wg ml31
([125I]tubulin) or 0.5 Wg ml31 ([125I]MAP-free tubulin), andincubated with 1.5 Wl of sPGN^agarose (20 min with[125I]tubulin, or 40 min with [125I]MAP-free tubulin), orReLPS^agarose (20 min for both [125I]tubulins) at 37³C.Then PBS with 0.5 M NaCl was added, the agarose wascentrifuged through 0.8 M sucrose, and the amount of 125Ibound to agarose was measured [5]. For the binding underde-polymerizing conditions, the assay was performed asabove but in the de-polymerizing bu¡er at 4³C, withpre-incubation of [125I]tubulin for 1 h, followed by incu-bation with sPGN^agarose or ReLPS^agarose for 3 h. Insome experiments, the amount of agarose, incubationtime, or tubulin concentrations were varied, as indicatedin Section 3. For inhibition of binding, unlabeled tubulinswere mixed with the iodinated tubulins and pre-incubatedas described above, before addition of sPGN^agarose orReLPS^agarose. In other competitive inhibition experi-ments, the competitors were ¢rst mixed with sPGN^aga-rose or ReLPS^agarose, followed by addition of iodinatedtubulin. The apparent dissociation constant (Kd) and themaximal binding at saturation (Bmax) were calculated asbefore [5].
2.6. Tubulin polymerization and electron microscopy
Tubulin (100 Wl, 2 mg ml31) was incubated in the poly-merizing bu¡er with 1 mM GTP, with or without 4 Mglycerol, with or without 25 Wg ml31 of sPGN or ReLPS,and with or without 0.8 mM colchicine, at 37³C in 96-wellmicrotiter plates, and the optical density (OD) at 405 nmwas measured using a BioRad ELISA plate reader. Similarincubation in de-polymerizing bu¡er at 4³C did not causean increase in OD (the OD values under de-polymerizingconditions were subtracted from the OD values underpolymerizing conditions to obtain net OD). 25 Wg ml31
of sPGN and ReLPS was selected because in preliminaryexperiments it had the most consistent e¡ect on microtu-bule polymerization (out of 2.5, 25, and 250 Wg ml31). Forelectron microscopy, after 80 min of incubation in thepolymerizing bu¡er with GTP, with or without glycerol,and with or without 25 Wg ml31 sPGN or ReLPS, sampleswere diluted 4 times, applied to carbon-coated nickel
grids, stained with 2% uranyl acetate [17], and viewed ina JOEL-100CXII electron microscope.
3. Results
3.1. Tubulin is a major PGN-binding protein in cell lysatesand membrane fractions of RAW264.7 macrophages
The major PGN-binding protein, consistently present inlysates of RAW264.7 cells and solubilized membrane frac-tions, that bound both to agarose-coupled sPGN or in-soluble PGN, was L-tubulin (GenBank accession no.X04663) (Fig. 1). When similar samples were separatedby two-dimensional electrophoresis (isoelectric focusingand SDS^PAGE), in addition to L-tubulin (which wasthe major spot), K-tubulin was also detected (not shown).CD14, another PGN-binding protein [5], could be detectedby photoa¤nity cross-linking [5] or on Western blots ofPGN-binding proteins with anti-CD14 antibodies (notshown), but the amount of CD14 recovered from thecell lysates was not su¤cient to yield a band that couldbe stained with Coomassie blue and sequenced.
3.2. Polymerized tubulin binds to sPGN and co-elutes withsPGN from anti-PGN vancomycin a¤nity column
When tubulin was incubated with sPGN under polymer-izing conditions and then applied to a vancomycin^aga-rose column, both sPGN and tubulin were retained on thecolumn and were then co-eluted at high pH (Fig. 2A,C,fractions 23 and 24). When tubulin was applied to thecolumn by itself, it was not retained on the column. Sim-
Fig. 1. Tubulin is a major PGN-binding protein in cell lysates andmembrane fractions of RAW264.7 macrophages. Cell lysates (A,B) ormembrane fraction (C) were incubated with insoluble PGN (A,C) orsPGN^agarose (B). PGN-binding proteins were resolved by SDS^PAGE, blotted onto Immobilon, and stained with Coomassie blue. Themajor band was identi¢ed by N-terminal sequencing as L-tubulin. Threeother minor proteins, that bound to insoluble PGN, but not to sPGN,were GRP 78 (78 kDa glucose-regulated protein), actin, and glyceralde-hyde-3-phosphate dehydrogenase (G-3P-D). The amino acid sequencesare shown in the single-letter code, and the positions of molecular massstandards (kDa) are shown on the left.
BBAGEN 25093 1-11-00
R. Dziarski et al. / Biochimica et Biophysica Acta 1524 (2000) 17^2620
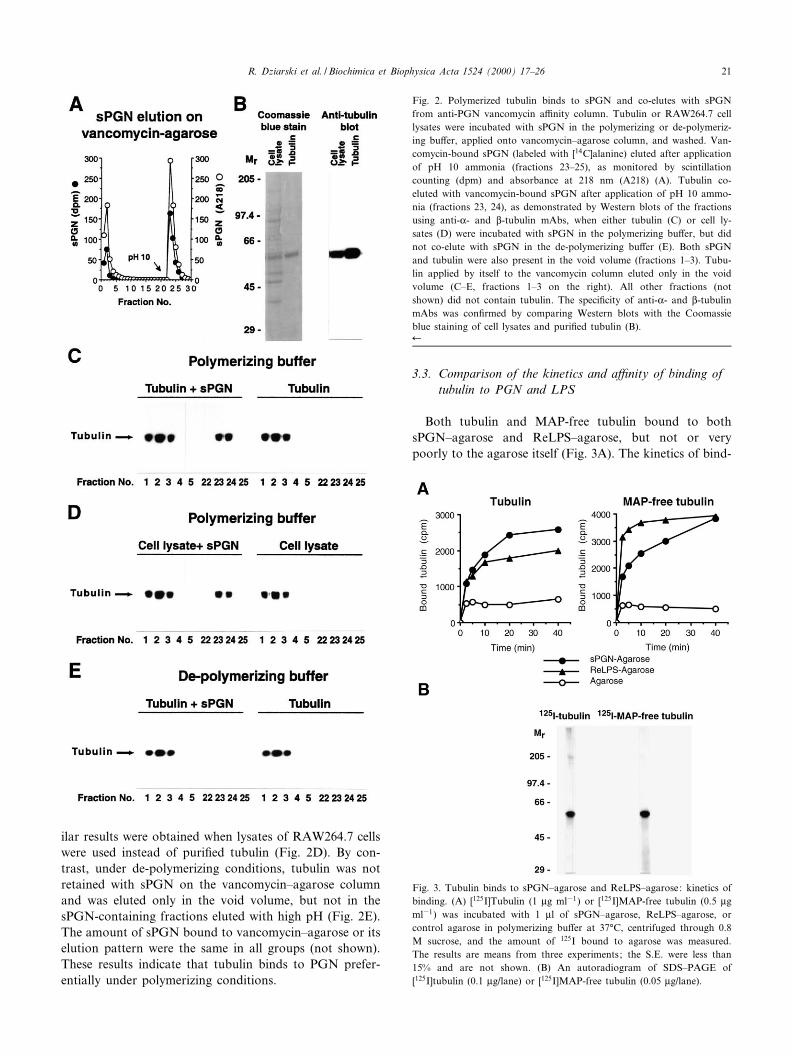
ilar results were obtained when lysates of RAW264.7 cellswere used instead of puri¢ed tubulin (Fig. 2D). By con-trast, under de-polymerizing conditions, tubulin was notretained with sPGN on the vancomycin^agarose columnand was eluted only in the void volume, but not in thesPGN-containing fractions eluted with high pH (Fig. 2E).The amount of sPGN bound to vancomycin^agarose or itselution pattern were the same in all groups (not shown).These results indicate that tubulin binds to PGN prefer-entially under polymerizing conditions.
3.3. Comparison of the kinetics and a¤nity of binding oftubulin to PGN and LPS
Both tubulin and MAP-free tubulin bound to bothsPGN^agarose and ReLPS^agarose, but not or verypoorly to the agarose itself (Fig. 3A). The kinetics of bind-
Fig. 2. Polymerized tubulin binds to sPGN and co-elutes with sPGNfrom anti-PGN vancomycin a¤nity column. Tubulin or RAW264.7 celllysates were incubated with sPGN in the polymerizing or de-polymeriz-ing bu¡er, applied onto vancomycin^agarose column, and washed. Van-comycin-bound sPGN (labeled with [14C]alanine) eluted after applicationof pH 10 ammonia (fractions 23^25), as monitored by scintillationcounting (dpm) and absorbance at 218 nm (A218) (A). Tubulin co-eluted with vancomycin-bound sPGN after application of pH 10 ammo-nia (fractions 23, 24), as demonstrated by Western blots of the fractionsusing anti-K- and L-tubulin mAbs, when either tubulin (C) or cell ly-sates (D) were incubated with sPGN in the polymerizing bu¡er, but didnot co-elute with sPGN in the de-polymerizing bu¡er (E). Both sPGNand tubulin were also present in the void volume (fractions 1^3). Tubu-lin applied by itself to the vancomycin column eluted only in the voidvolume (C^E, fractions 1^3 on the right). All other fractions (notshown) did not contain tubulin. The speci¢city of anti-K- and L-tubulinmAbs was con¢rmed by comparing Western blots with the Coomassieblue staining of cell lysates and puri¢ed tubulin (B).6
Fig. 3. Tubulin binds to sPGN^agarose and ReLPS^agarose: kinetics ofbinding. (A) [125I]Tubulin (1 Wg ml31) or [125I]MAP-free tubulin (0.5 Wgml31) was incubated with 1 Wl of sPGN^agarose, ReLPS^agarose, orcontrol agarose in polymerizing bu¡er at 37³C, centrifuged through 0.8M sucrose, and the amount of 125I bound to agarose was measured.The results are means from three experiments; the S.E. were less than15% and are not shown. (B) An autoradiogram of SDS^PAGE of[125I]tubulin (0.1 Wg/lane) or [125I]MAP-free tubulin (0.05 Wg/lane).
BBAGEN 25093 1-11-00
R. Dziarski et al. / Biochimica et Biophysica Acta 1524 (2000) 17^26 21

ing of tubulin to sPGN and ReLPS was similar, with mostof the binding completed in 20 min. MAP-free tubulinbound to ReLPS with a much faster kinetics than tosPGN: binding to ReLPS was completed in 10 min, butto sPGN it required 40 min (Fig. 3A).
Both tubulin and MAP-free tubulin bound to sPGN ina saturable manner with similar a¤nity. The binding couldbe ¢tted to a hyperbolic curve and into a single straightline in a Scatchard plot, yielding an apparent Kd = 147 þ 35nM for tubulin and Kd = 106 þ 16 nM for MAP-free tubu-lin (Fig. 4A). The Bmax, however, was twice as high fortubulin than for MAP-free tubulin (230 þ 32 vs 110 þ 9 ngml31).
In contrast to sPGN, binding of both tubulin andMAP-free tubulin to ReLPS showed cooperative binding(indicated by concave upwards Scatchard plots) (Fig. 4B).Also, the a¤nities of binding of tubulin and MAP-freetubulin to ReLPS (calculated from the descending slopesof the Scatchard plots) were lower than to sPGN. Thisdi¡erence in the a¤nity of binding to sPGN and ReLPSwas a function of the ligand (tubulin) and not of a speci¢cbatch of sPGN^agarose or ReLPS^agarose, because sim-ilar results were obtained with two independently preparedbatches of each agarose, and because the same agarosepreparations previously [5] gave the expected higher a¤n-
ity of binding of CD14 to ReLPS than to sPGN in thepresence of LPS-binding protein.
Under de-polymerizing conditions, the amount of tubu-lin or MAP-free tubulin speci¢cally bound to sPGN orReLPS (Bmax) decreased 10 or 5 times, respectively (datanot shown). De-polymerizing conditions reduced theamount of microtubules by over 95% (determined by elec-tron microscopy). These results explain why in Fig. 1 thebinding was detected in the solubilizing bu¡er (which doesnot promote tubulin polymerization): binding under de-polymerizing conditions is 10 times lower than under poly-merizing conditions, and in Fig. 1, cell lysate from 200times more cells than in Fig. 2D was used.
3.4. Di¡erential e¡ects of unlabeled tubulin, sPGN, andReLPS on the binding of tubulin to sPGN and ReLPS
Unlabeled tubulin or MAP-free tubulin competitivelyinhibited the binding of labeled [125I]tubulin or[125I]MAP-free tubulin to sPGN (Fig. 5). By contrast,the unlabeled tubulins not only did not inhibit the bindingto ReLPS, but the addition of unlabeled tubulins or to[125I]tubulins greatly enhanced the binding of labeled tu-bulins to ReLPS (Fig. 5). These results suggest thatReLPS induces or enhances tubulin polymerization (or
Fig. 4. Tubulin binds to sPGN^agarose with higher a¤nity than to ReLPS^agarose and binding of tubulin to ReLPS is cooperative. Increasing concen-trations of [125I]tubulin or [125I]MAP-free tubulin were incubated with 1.5 Wl of control agarose and sPGN^agarose (A) or ReLPS^agarose (B) in thepolymerizing bu¡er, and the amounts of [125I]tubulin bound to agarose and remaining unbound (free [125I]tubulin) were measured. Total binding wasthe amount of 125I associated with sPGN^agarose or ReLPS^agarose; non-speci¢c binding was the amount of 125I associated with control agarose; spe-ci¢c binding was total binding minus non-speci¢c binding. The results are means of 4^5 experiments. The Scatchard plots were ¢tted using the CricketGraph software (r2 = 0.82 þ 0.024, mean þ S.E.) ; mean apparent dissociation constants (Kd) and Bmax ( þ S.E.) are also shown.
BBAGEN 25093 1-11-00
R. Dziarski et al. / Biochimica et Biophysica Acta 1524 (2000) 17^2622

aggregation) that leads to continued increased binding ofaggregated tubulin to ReLPS (or to tubulin bound toReLPS). They also con¢rm the positive cooperative bind-ing of tubulin to ReLPS (presented in Fig. 4B). sPGN (insolution) inhibited the binding of [125I]tubulin to bothsPGN and ReLPS (Fig. 5). ReLPS (in solution) increasedthe binding of [125I]tubulin to ReLPS and showed onlyvery weak inhibition of tubulin binding to sPGN at high
concentrations (Fig. 5). These results further support thecooperative binding of tubulin to ReLPS.
3.5. Macromolecular sPGN is required for binding totubulin
Neither tubulin nor MAP-free tubulin bound to any ofthe low molecular mass synthetic PGN fragments (MDP,
Fig. 5. Di¡erential e¡ects of unlabeled tubulin, sPGN, and ReLPS on binding of tubulin to sPGN^agarose and ReLPS^agarose. Binding of [125I]tubulinor [125I]MAP-free tubulin to sPGN^agarose or ReLPS^agarose in the polymerizing bu¡er in the presence of indicated concentrations of unlabeled tubu-lin or MAP-free tubulin, sPGN, or ReLPS was measured. The results were calculated as the percent of binding without a competitor and are meansfrom three experiments; the S.E. were less than 18% and are not shown.
Fig. 6. Macromolecular sPGN is required for binding to tubulin. [125I]tubulin (A) or [125I]MAP-free tubulin (B) bound to sPGN^agarose, but not tolow Mr sPGN fragments (GlcNAc-MDP, MDP, and PGN pentapeptide) coupled to agarose. The ability of sPGN (at 50 Wg ml31) to inhibit the bindingof tubulin to sPGN^agarose was lost after digestion of sPGN with lysostaphin and dialysis (C). The extent of digestion of sPGN was con¢rmed by theloss of [14C]alanine from macromolecular biosynthetically labeled sPGN following lysostaphin digestion and dialysis (D). The results are from one oftwo similar experiments (A,B) or mean þ S.E. from four assays from two experiments (C,D).
BBAGEN 25093 1-11-00
R. Dziarski et al. / Biochimica et Biophysica Acta 1524 (2000) 17^26 23
GlcNAc-MDP, and PGN pentapeptide) coupled to aga-rose (Fig. 6A,B). These synthetic low Mr PGN fragments,in contrast to high Mr polymeric PGN (Fig. 5), also didnot inhibit tubulin binding to sPGN and ReLPS (data notshown). Morover, digestion of polymeric sPGN with aPGN-lytic enzyme, lysostaphin, abolished the ability ofsPGN to competitively inhibit the binding of both tubulinand MAP-free tubulin to sPGN^agarose (Fig. 6C,D).These results suggest that polymeric PGN is required forthe binding to tubulin.
3.6. Taxol diminishes binding of tubulin to sPGN andReLPS
Taxol (an agent that stabilizes microtubules and pro-motes microtubule assembly) did not further increase thebinding of tubulin and MAP-free tubulin to sPGN orReLPS, but it diminished the binding of tubulin to bothsPGN and ReLPS (Fig. 7).
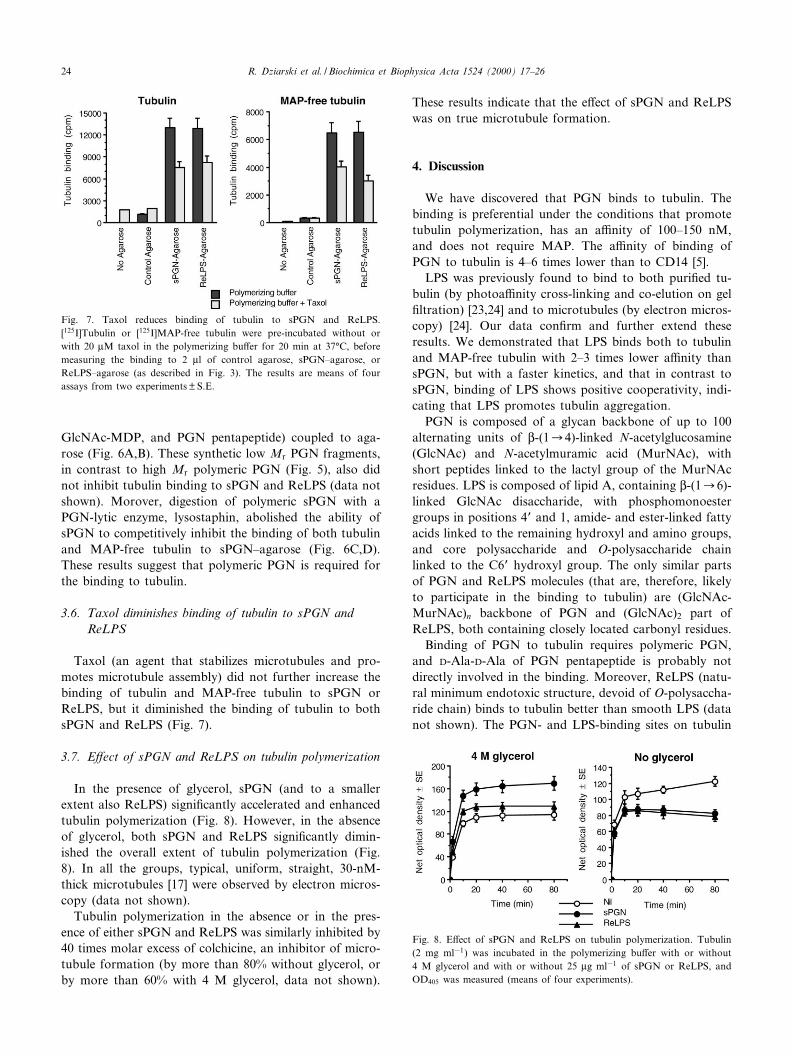
3.7. E¡ect of sPGN and ReLPS on tubulin polymerization
In the presence of glycerol, sPGN (and to a smallerextent also ReLPS) signi¢cantly accelerated and enhancedtubulin polymerization (Fig. 8). However, in the absenceof glycerol, both sPGN and ReLPS signi¢cantly dimin-ished the overall extent of tubulin polymerization (Fig.8). In all the groups, typical, uniform, straight, 30-nM-thick microtubules [17] were observed by electron micros-copy (data not shown).
Tubulin polymerization in the absence or in the pres-ence of either sPGN and ReLPS was similarly inhibited by40 times molar excess of colchicine, an inhibitor of micro-tubule formation (by more than 80% without glycerol, orby more than 60% with 4 M glycerol, data not shown).
These results indicate that the e¡ect of sPGN and ReLPSwas on true microtubule formation.
4. Discussion
We have discovered that PGN binds to tubulin. Thebinding is preferential under the conditions that promotetubulin polymerization, has an a¤nity of 100^150 nM,and does not require MAP. The a¤nity of binding ofPGN to tubulin is 4^6 times lower than to CD14 [5].
LPS was previously found to bind to both puri¢ed tu-bulin (by photoa¤nity cross-linking and co-elution on gel¢ltration) [23,24] and to microtubules (by electron micros-copy) [24]. Our data con¢rm and further extend theseresults. We demonstrated that LPS binds both to tubulinand MAP-free tubulin with 2^3 times lower a¤nity thansPGN, but with a faster kinetics, and that in contrast tosPGN, binding of LPS shows positive cooperativity, indi-cating that LPS promotes tubulin aggregation.
PGN is composed of a glycan backbone of up to 100alternating units of L-(1C4)-linked N-acetylglucosamine(GlcNAc) and N-acetylmuramic acid (MurNAc), withshort peptides linked to the lactyl group of the MurNAcresidues. LPS is composed of lipid A, containing L-(1C6)-linked GlcNAc disaccharide, with phosphomonoestergroups in positions 4P and 1, amide- and ester-linked fattyacids linked to the remaining hydroxyl and amino groups,and core polysaccharide and O-polysaccharide chainlinked to the C6P hydroxyl group. The only similar partsof PGN and ReLPS molecules (that are, therefore, likelyto participate in the binding to tubulin) are (GlcNAc-MurNAc)n backbone of PGN and (GlcNAc)2 part ofReLPS, both containing closely located carbonyl residues.
Binding of PGN to tubulin requires polymeric PGN,and D-Ala-D-Ala of PGN pentapeptide is probably notdirectly involved in the binding. Moreover, ReLPS (natu-ral minimum endotoxic structure, devoid of O-polysaccha-ride chain) binds to tubulin better than smooth LPS (datanot shown). The PGN- and LPS-binding sites on tubulin
Fig. 7. Taxol reduces binding of tubulin to sPGN and ReLPS.[125I]Tubulin or [125I]MAP-free tubulin were pre-incubated without orwith 20 WM taxol in the polymerizing bu¡er for 20 min at 37³C, beforemeasuring the binding to 2 Wl of control agarose, sPGN^agarose, orReLPS^agarose (as described in Fig. 3). The results are means of fourassays from two experiments þ S.E.
Fig. 8. E¡ect of sPGN and ReLPS on tubulin polymerization. Tubulin(2 mg ml31) was incubated in the polymerizing bu¡er with or without4 M glycerol and with or without 25 Wg ml31 of sPGN or ReLPS, andOD405 was measured (means of four experiments).
BBAGEN 25093 1-11-00
R. Dziarski et al. / Biochimica et Biophysica Acta 1524 (2000) 17^2624

are unknown, but they seem to be at least partially similar,because sPGN could competitively inhibit tubulin bindingto both sPGN and ReLPS. However, the binding sites forsPGN and LPS are not identical, because sPGN was abetter inhibitor for tubulin binding to sPGN than toReLPS, although the a¤nity of binding of tubulin tosPGN was higher than to ReLPS. Similar binding sitesfor sPGN and ReLPS are also suggested by diminishedbinding of tubulin to both sPGN and ReLPS in the pres-ence of taxol.
sPGN and ReLPS have a direct e¡ect on tubulin poly-merization in vitro. In the presence of glycerol, sPGNenhances tubulin polymerization signi¢cantly more thanLPS whereas, without glycerol, both sPGN and LPS in-hibit tubulin polymerization (Fig. 8). Our results withoutglycerol are, therefore, consistent with the previously ob-served fragmentation of microtubules induced by 10 timeshigher concentration of LPS in vitro [32]. Di¡erent e¡ectsin the medium with glycerol may be attributed to thestabilizing properties of glycerol for microtubules or itse¡ect on MAP binding.
Although the involvement of microtubules in cell acti-vation is not precisely known, microtubules serve as asca¡old for numerous signal transduction molecules thatare associated with tubulin. For example, most of theLPS-activable mitogen-activated protein kinases in macro-phages are associated with microtubules [14,15]. More-over, several other key signal transduction molecules,such as G proteins [16,17,33], ZAP-70 kinase [34], Vav[34], Syk [35], phosphoinositide 3-kinase [36,37], phospho-lipase CQ1 [36], c-Myc [38], and Rac1 [39] are associatedwith tubulin. Tubulin binds Src homology 2 domains(SH2) [36], and dynamin (one of MAPs) has binding sitesfor Src homology 3 domains (SH3) [40]. Both SH2 andSH3 are often present in signal transduction molecules.Furthermore, some cell surface receptors, e.g., glycine re-ceptor [41] or CD2 [42], are associated with tubulin, andupon cell activation tubulin is phosphorylated on a singletyrosyl residue by Syk [35]. Therefore, these results collec-tively suggest that tubulin and microtubules are importantparticipants in the cell activation processes. Because someof these proteins are involved in LPS- and PGN-inducedcell activation [2,3,15,30,31,43], it is possible that tubulinmay also play a role in LPS- and PGN-induced cell acti-vation. This possibility needs to be investigated.
Because both PGN and LPS bind to tubulin and have adirect e¡ect on tubulin polymerization, it is possible thatPGN and LPS could interact with tubulin in the cell andmodify microtubule function. Tubulin is located intracel-lularly mainly as a part of microtubules and also as a freeprotein pool. Therefore, extracellular ligands have to enterthe cell to interact with tubulin. LPS is rapidly (withinminutes) taken up by macrophages following interactionwith membrane CD14, although LPS uptake may not berequired for LPS-induced signal transduction [2,3]. How-ever, recent results indicate that endocytic transport of
LPS from the cell surface to the perinuclear Golgi appa-ratus is important for LPS-induced cell activation [25,26].The exact mechanism of this transport is still unknown,but one of the well-established functions of microtubules isunidirectional transport of vesicles along microtubuletracks [13]. sPGN is also e¤ciently taken up by monocytesand macrophages (A.J. Ulmer, R. Dziarski, unpublished).
It is still not known how PGN or LPS would gainaccess to microtubules in the cells. It has been suggestedthat LPS can intercalate into the cytoplasmic or vesiclemembranes [26] or that, similar to bacterial or plant tox-ins, LPS could cross the vesicle membranes into the cyto-plasm [26]. Both mechanisms would give LPS an oppor-tunity to interact with microtubules. It has been alsosuggested that tubulin may be exposed on the surface ofmonocytic cells and that LPS stimulation of these cellsresults in a marked reduction of the amount of cell sur-face-exposed tubulin [44]. This cell-surface tubulin wouldprovide an opportunity for LPS and PGN to interact withtubulin before their internalization. However, the presenceof cell-surface tubulin and its relevance to LPS- and PGN-induced cell activation need to be further con¢rmed. Nowthat the binding of PGN and LPS to tubulin and theire¡ects on microtubules have been demonstrated, themechanism of interaction of PGN and LPS with tubulinand microtubules in the cells needs to be investigated.
Acknowledgements
This work was supported by National Institute ofHealth Grants AI2879 (to R.D.) and MH39595 andAG15482 (to M.M.R.).
References
[1] B. Heymer, P.H. Seidl, K.H. Schleifer. Immunochemistry and biolog-ical activity of peptidoglycan, in: D.E.S. Stewart-Tull, M. Davis(Eds.), Immunology of the Bacterial Cell Envelope, Wiley, NewYork, 1985, pp. 11^46.
[2] R. Dziarski, A.J. Ulmer, D. Gupta, Interactions of bacterial lipopoly-saccharide and peptidoglycan with mammalian CD14, in: R.J. Doyle(Ed.), Glycomicrobiology, Kluwer Academic/Plenum, New York,2000, pp. 145^185.
[3] R. Dziarski, A.J. Ulmer, D. Gupta, Interactions of CD14 with com-ponents of Gram-positive bacteria, Chem. Immunol. 74 (2000) 83^107.
[4] D. Gupta, T.N. Kirkland, S. Viriyakosol, R. Dziarski, CD14 is a cell-activating receptor for bacterial peptidoglycan, J. Biol. Chem. 271(1996) 23310^23316.
[5] R. Dziarski, R.I. Tapping, P.S. Tobias, Binding of bacterial peptido-glycan to CD14, J. Biol. Chem. 273 (1998) 8680^8690.
[6] R. Schwandner, R. Dziarski, H. Wesche, M. Rothe, C.J. Kirschning,Peptidoglycan- and lipoteichoic acid-induced cell activation is medi-ated by Toll-like receptor 2, J. Biol. Chem. 274 (1999) 17406^17409.
[7] A. Yoshimura, E. Lien, R.R. Ingalls, E. Tuomanen, R. Dziarski, D.Golenbock, Recognition of Gram-positive bacterial cell wall compo-nents by the innate immune system occurs via Toll-like receptor 2,J. Immunol. 163 (1999) 1^5.
BBAGEN 25093 1-11-00
R. Dziarski et al. / Biochimica et Biophysica Acta 1524 (2000) 17^26 25

[8] R.-B. Yang, M.R. Mark, A. Gray, A. Huang, M.H. Xie, M. Zhang,A. Goddard, W.I. Wood, A.L. Gurney, P.J. Godowski, Toll-likereceptor-2 mediates lipopolysaccharide-induced cellular signaling,Nature 395 (1998) 284^288.
[9] C.J. Kirschning, H. Wesche, T.M. Ayres, M. Rothe, Human Toll-likereceptor 2 confers responsiveness to bacterial lipopolysaccharide,J. Exp. Med. 188 (1998) 2091^2097.
[10] A. Poltorak, X. He, I. Smirnova, M.-Y. Liu, C. Van Hu¡el, X. Du,D. Birdwell, E. Alejos, M. Silva, C. Galanos, M. Freudenberg, P.Riccardi-Castagnoli, B. Layton, B. Beutler, Defective LPS signalingin C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene, Sci-ence 282 (1998) 2085^2088.
[11] K. Hoshino, O. Takeuchi, T. Kawai, H. Sanjo, T. Ogawa, Y. Take-da, K. Takeda, S. Akira, Toll-like receptor 4 (TLR4)-de¢cient miceare hyporesponsive to lipopolysaccharide : evidence for TLR4 as theLps gene product, J. Immunol. 162 (1999) 3749^3752.
[12] S.D. Wright, Toll, a new piece in the puzzle of innate immunity,J. Exp. Med. 189 (1999) 605^609.
[13] L. Stryer, Biochemistry, 4th ed., W.H. Freeman, New York, 1995,pp. 405^411.
[14] A.A. Reszka, R. Seger, C.D. Diltz, E.G. Krebs, E.H. Fischer, Asso-ciation of mitogen-activated protein kinase with the microtubule cy-toskeleton, Proc. Natl. Acad. Sci. USA 92 (1995) 8881^8885.
[15] A. Ding, B. Chen, M. Fuortes, E. Bloom, Association of mitogen-activated protein kinases with microtubules in mouse macrophages,J. Exp. Med. 183 (1996) 1899^1904.
[16] N. Wang, K. Yan, M.M. Rasenick, Tubulin binds speci¢cally to thesignal-transducing proteins, GsK and GiK1, J. Biol. Chem. 265 (1990)1239^1242.
[17] S. Roychowdhury, M.M. Rasenick, G protein L1Q2 subunits promotemicrotubule assembly, J. Biol. Chem. 272 (1997) 31576^31581.
[18] A.H. Ding, F. Porteu, E. Sanchez, C.F. Nathan, Shared actions ofendotoxin and taxol on TNF receptors and TNF release, Science 248(1990) 370^372.
[19] A. Ding, E. Sanchez, C.F. Nathan, Taxol shares the ability of bac-terial lipopolysaccharide to induce tyrosine phosphorylation of mi-crotubule-associated protein kinase, J. Immunol. 151 (1993) 5596^5602.
[20] C.L. Manthey, M.E. Brandes, P.Y. Perera, S.N. Vogel, Taxol in-creases steady-state levels of lipopolysaccharide-inducible genes andprotein-tyrosine phosphorylation in murine macrophages, J. Immu-nol. 149 (1992) 2459^2465.
[21] C.L. Manthey, N. Qureshi, P.L. Stutz, S.N. Vogel, Lipopolysaccha-ride antagonists block taxol-induced signaling in murine macro-phages, J. Exp. Med. 178 (1993) 695^702.
[22] P.-Y. Perera, N. Qureshi, S.N. Vogel, Paclitaxel (taxol)-induced NF-UB translocation in murine macrophages, Infect. Immun. 64 (1996)878^884.
[23] A. Ding, E. Sanchez, M. Tancino, C. Nathan, Interactions of bacte-rial lipopolysaccharide with microtubule proteins, J. Immunol. 148(1992) 2853^2858.
[24] C. Risco, J.E. Dominguez, M.A. Bosch, J.L. Carrascosa, Biochemicaland electron microscopy analysis of the endotoxin binding to micro-tubules in vitro, Mol. Cell. Biochem. 121 (1993) 67^74.
[25] N. Thieblemont, R. Thieringer, S.D. Wright, Innate immune recog-nition of bacterial lipopolysaccharide: dependence on interactionswith membrane lipids and endocytic movement, Immunity 8 (1998)771^777.
[26] N. Thieblemont, S.D. Wright, Transport of bacterial lipopolysaccha-ride to the Golgi apparatus, J. Exp. Med. 190 (1999) 523^534.
[27] I. Ringel, S.B. Horwitz, E¡ect of alkaline pH on taxol-microtubuleinteractions, J. Pharmacol. Exp. Ther. 259 (1991) 855^860.
[28] R.S. Rosenthal, R. Dziarski, Isolation of peptidoglycan and solublepeptidoglycan fragments, Methods Enzymol. 235 (1994) 253^286.
[29] R. Dziarski, Demonstration of peptidoglycan-binding sites on lym-phocytes and macrophages by photoa¤nity cross-linking, J. Biol.Chem. 266 (1991) 4713^4718.
[30] D. Gupta, Y. Jin, R. Dziarski, Peptidoglycan induces transcriptionand secretion of TNF-K and activation of Lyn, extracellular signal-regulated kinase, and Rsk signal transduction proteins in mouse mac-rophages, J. Immunol. 155 (1995) 2620^2630.
[31] R. Dziarski, Correlation between ribosylation of pertussis toxin sub-strates and inhibition of peptidoglycan-, muramyl didpeptide- andlipopolysaccharide-induced mitogenic stimulation in B lymphocytes,Eur. J. Immunol. 19 (1989) 125^130.
[32] K.J. Bohm, W. Vater, S. Russwurm, K. Reinhart, E. Unger, Lipo-polysaccharide-caused fragmentation of individual microtubules invitro observed by video-enhanced di¡erential interference contrastmicroscopy, FEBS Lett. 425 (1998) 134^136.
[33] S. Roychowdhury, N. Wang, M.M. Rasenick, G protein binding andG protein activation by nucleotide transfer involve distinct domainson tubulin: regulation of signal transduction by cytoskeletal ele-ments, Biochemistry 32 (1993) 4955^4961.
[34] R.D.J. Huby, G.W. Carlile, S.C. Ley, Interactions between the pro-tein-tyrosine kinase ZAP-70, the proto-oncoprotein Vav, and tubulinin Jurkat T cells, J. Biol. Chem. 270 (1995) 30241.
[35] J.D. Peters, M.T. Furlong, D.J. Asai, M.L. Harrison, R.L. Geahlen,Syk, activated by cross-linking the B-cell antigen receptor, localizes tothe cytosol where it interacts with and phosphorylates K-tubulin ontyrosine, J. Biol. Chem. 271 (1996) 4755^4762.
[36] T. Itoh, K. Miura, H. Miki, T. Takenawa, L-tubulin binds Src ho-mology 2 domains through a region di¡erent from the tyrosine-phos-phorylated protein-recognizing site, J. Biol. Chem. 271 (1996) 27931^27935.
[37] R. Kapeller, A. Toker, L.C. Cantley, C.L. Carpenter, Phosphoinosi-tide 3-kinase binds constitutively to K/L-tubulin and binds to Q-tubu-lin in response to insulin, J. Biol. Chem. 270 (1995) 25985^25991.
[38] N. Alexandrova, J. Niklinski, V. Bliskovsky, G.A. Otterson, M.Blake, F.J. Kaye, M. Zajac-Kaye, The N-terminal domain of c-Myc associates with K-tubulin and microtubules in vivo and in vitro,Mol. Cell. Biol. 15 (1995) 5188^5195.
[39] A. Best, S. Ahmed, R. Kozma, L. Lim, The Ras-related GTPaseRac1 binds tubulin, J. Biol. Chem. 271 (1996) 3756^3762.
[40] J.S. Herskovits, H.S. Shpetner, C.C. Burgess, R.B. Vallee, Microtu-bules and Src homology 3 domains stimulate the dynamin GTPasevia its C-terminal domain, Proc. Natl. Acad. Sci. USA 90 (1993)11468^11472.
[41] J. Kirsch, D. Langosch, P. Prior, U.Z. Littauer, B. Schmitt, H. Betz,The 93-kDa glycine receptor-associated protein binds to tubulin,J. Biol. Chem. 266 (1991) 22242^22245.
[42] R. O¡ringa, B.E. Bierer, Association of CD2 with tubulin: Evidencefor a role of the cytoskeleton in T cell activation, J. Biol. Chem. 268(1993) 4979^4988.
[43] Y. Jin, D. Gupta, R. Dziarski, Endothelial and epithelial cells do notrespond to complexes of peptidoglycan with soluble CD14, but areactivated indirectly by peptidoglycan-induced tumor necrosis factor-Kand interleukin-1 from monocytes, J. Infect. Dis. 177 (1998) 1629^1638.
[44] S.B. Por, M.A. Cooley, S.N. Breit, R. Penny, P.W. French, Anti-bodies to tubulin and actin bind to the surface of a human monocyticcell line, U937, J. Histochem. Cytochem. 39 (1991) 981^985.
BBAGEN 25093 1-11-00
R. Dziarski et al. / Biochimica et Biophysica Acta 1524 (2000) 17^2626




















