
Peptidoglycan recognition proteins: pleiotropic sensors and effectors of antimicrobial defences
14
Peptidoglycan is an essential component of the cell wall of virtually all bacteria (BOX 1) and is not only a well- known target for antibiotics but is also a target for the host immune response through recognition by pattern recognition receptors (PRRs), including the peptido- glycan recognition proteins (PGRPs). The name PGRP was first introduced by Ashida’s group 1 . They purified a 19 kDa protein present in the haemolymph of a silk- worm (Bombyx mori) that bound peptidoglycan and activated the prophenoloxidase cascade, an antimicrobial defence mechanism in insects 1 . A PGRP orthologue was then cloned in the moth Trichoplusia ni 2 and in B. mori 3 . The cloning of insect PGRPs led to the dis- covery of mouse and human PGRP orthologues by Steiner’s group 2 , which showed that PGRPs are highly conserved from insects to mammals. A protein with the same sequence, named Tag7 (tumour-associated gene 7), was also cloned independently from a mouse adenocarcinoma 4 . The sequencing of the Drosophila melanogaster, Anopheles gambiae and human genomes led to the discovery of families of highly diversified PGRP homo- logues in both insects and mammals 5–7 . This was an intriguing finding because, unlike insect cells, mamma- lian cells do not possess the prophenoloxidase cascade. Subsequently, PGRPs have also been identified in mol- luscs, echinoderms and several groups of vertebrates 8–11 . However, plants and lower metazoa such as nematodes (Caenorhabditis elegans) do not have PGRPs. As the functions of PGRPs were being discovered, it became clear that all PGRPs function in the host response to infection. They accomplish this goal, how- ever, by different strategies. In insects, PGRPs are mainly recognition proteins that sense microorganisms and trigger defence pathways that generate antimicrobial effectors. By contrast, mammalian PGRPs function as both microbial recognition proteins and antimicrobial effectors. General features of PGRP genes and proteins Insect PGRP genes are classified into short (S) and long (L) transcripts and are often alternatively spliced. For example, D. melanogaster has 13 PGRP genes spliced into 19 proteins 5 (TABLE 1), and A. gambiae has 7 PGRP genes spliced into 9 proteins 7 . Mammals have a family of 4 PGRPs, which were initially named PGRP-S, PGRP-L, PGRP-Iα and PGRP-Iβ (for short, long and intermediate α and β transcripts, respectively), analogous to insect PGRPs 6 . Subsequently, these names were changed to peptidoglycan recognition protein 1, 2, 3 and 4 (PGLYRP-1, PGLYRP-2, PGLYRP-3 and PGLYRP-4), respectively (TABLE 1). PGRP genes usually form clusters, which indicates their origin by gene duplication. All PGRP proteins have at least one C-terminal PGRP domain (~165 amino-acid residues) that is homologous to the bacteriophage and bacterial type 2 amidases 2,4,6,8 . This homology indicates that PGRPs and prokaryotic type 2 amidases might have evolved from a *Institut de Biologie du Développement de Marseille- Luminy, UMR 6216 CNRS, Université de la Méditérannée Aix-Marseille II, Marseille, France. ‡ Indiana University School of Medicine, Northwest Campus, Gary, Indiana 46408, USA. Correspondence to J.R. and R.D. e-mails: Julien.Royet@ibdm. univ-mrs.fr; [email protected] doi:10.1038/nrmicro1620 Haemolymph Circulatory fluid found in insects, which bathes the organs directly. Prophenoloxidase cascade Serine-proteinase-based cascade leading to activation of a prophenoloxidase-activating proteinase, which converts inactive prophenoloxidase to active phenoloxidase and results in the generation of quinone and quinone intermediates. Polymerization of quinone intermediates can form melanin. Peptidoglycan recognition proteins: pleiotropic sensors and effectors of antimicrobial defences Julien Royet* and Roman Dziarski ‡ Abstract | Peptidoglycan recognition proteins (PGRPs) are innate immunity molecules that are present in most invertebrate and vertebrate animals. All PGRPs function in antimicrobial defence and are homologous to the prokaryotic peptidoglycan-lytic type 2 amidases. However, only some PGRPs have the catalytic activity that protects the host from excessive inflammation, and most PGRPs have diversified to carry out other host-defence functions. Insect and mammalian PGRPs defend host cells against infection through very different mechanisms. Insect PGRPs activate signal transduction pathways in host cells or trigger proteolytic cascades in the haemolymph, both of which generate antimicrobial effectors. By contrast, mammalian PGRPs are directly bactericidal. Here, we review these contrasting modes of action. REVIEWS 264 | APRIL 2007 | VOLUME 5 www.nature.com/reviews/micro © 2007 Nature Publishing Group
Transcript of Peptidoglycan recognition proteins: pleiotropic sensors and effectors of antimicrobial defences

Peptidoglycan is an essential component of the cell wall of virtually all bacteria (BOX 1) and is not only a well-known target for antibiotics but is also a target for the host immune response through recognition by pattern recognition receptors (PRRs), including the peptido-glycan recognition proteins (PGRPs). The name PGRP was first introduced by Ashida’s group1. They purified a 19 kDa protein present in the haemolymph of a silk-worm (Bombyx mori) that bound peptidoglycan and activated the prophenoloxidase cascade, an antimicrobial defence mechanism in insects1. A PGRP orthologue was then cloned in the moth Trichoplusia ni 2 and in B. mori3. The cloning of insect PGRPs led to the dis-covery of mouse and human PGRP orthologues by Steiner’s group2, which showed that PGRPs are highly conserved from insects to mammals. A protein with the same sequence, named Tag7 (tumour-associated gene 7), was also cloned independently from a mouse adenocarcinoma4.
The sequencing of the Drosophila melanogaster, Anopheles gambiae and human genomes led to the discovery of families of highly diversified PGRP homo-logues in both insects and mammals5–7. This was an intriguing finding because, unlike insect cells, mamma-lian cells do not possess the prophenoloxidase cascade. Subsequently, PGRPs have also been identified in mol-luscs, echinoderms and several groups of vertebrates8–11. However, plants and lower metazoa such as nematodes (Caenorhabditis elegans) do not have PGRPs.
As the functions of PGRPs were being discovered, it became clear that all PGRPs function in the host response to infection. They accomplish this goal, how-ever, by different strategies. In insects, PGRPs are mainly recognition proteins that sense microorganisms and trigger defence pathways that generate antimicrobial effectors. By contrast, mammalian PGRPs function as both microbial recognition proteins and antimicrobial effectors.
General features of PGRP genes and proteinsInsect PGRP genes are classified into short (S) and long (L) transcripts and are often alternatively spliced. For example, D. melanogaster has 13 PGRP genes spliced into 19 proteins5 (TABLE 1), and A. gambiae has 7 PGRP genes spliced into 9 proteins7. Mammals have a family of 4 PGRPs, which were initially named PGRP-S, PGRP-L, PGRP-Iα and PGRP-Iβ (for short, long and intermediate α and β transcripts, respectively), analogous to insect PGRPs6. Subsequently, these names were changed to peptidoglycan recognition protein 1, 2, 3 and 4 (PGLYRP-1, PGLYRP-2, PGLYRP-3 and PGLYRP-4), respectively (TABLE 1). PGRP genes usually form clusters, which indicates their origin by gene duplication.
All PGRP proteins have at least one C-terminal PGRP domain (~165 amino-acid residues) that is homologous to the bacteriophage and bacterial type 2 amidases2,4,6,8. This homology indicates that PGRPs and prokaryotic type 2 amidases might have evolved from a
*Institut de Biologie du Développement de Marseille-Luminy, UMR 6216 CNRS, Université de la Méditérannée Aix-Marseille II, Marseille, France. ‡Indiana University School of Medicine, Northwest Campus, Gary, Indiana 46408, USA. Correspondence to J.R. and R.D. e-mails: [email protected]; [email protected]:10.1038/nrmicro1620
Haemolymph Circulatory fluid found in insects, which bathes the organs directly.
Prophenoloxidase cascadeSerine-proteinase-based cascade leading to activation of a prophenoloxidase-activating proteinase, which converts inactive prophenoloxidase to active phenoloxidase and results in the generation of quinone and quinone intermediates. Polymerization of quinone intermediates can form melanin.
Peptidoglycan recognition proteins: pleiotropic sensors and effectors of antimicrobial defencesJulien Royet* and Roman Dziarski‡
Abstract | Peptidoglycan recognition proteins (PGRPs) are innate immunity molecules that are present in most invertebrate and vertebrate animals. All PGRPs function in antimicrobial defence and are homologous to the prokaryotic peptidoglycan-lytic type 2 amidases. However, only some PGRPs have the catalytic activity that protects the host from excessive inflammation, and most PGRPs have diversified to carry out other host-defence functions. Insect and mammalian PGRPs defend host cells against infection through very different mechanisms. Insect PGRPs activate signal transduction pathways in host cells or trigger proteolytic cascades in the haemolymph, both of which generate antimicrobial effectors. By contrast, mammalian PGRPs are directly bactericidal. Here, we review these contrasting modes of action.
R E V I E W S
264 | APRIL 2007 | VOLUME 5 www.nature.com/reviews/micro
© 2007 Nature Publishing Group

O
OH
OHO
NHO O
OH
ONH
CO
CH3
CO
CH3
HC(CH3)
MurNAcβ(1,4)GlcNAc
n
n
O
OH
OHO
n
NHO O
OH
ONH
CO
CH3
CO
CH3
HC(CH3)
CO
l-Ala
d-iGln
l-Lys
d-Ala
d-Ala
l-Lys
d-iGln
l-Ala
MurNAcβ(1,4)GlcNAc
n
(Gly)5
a DAP-type petidoglycan b Lys-type petidoglycan
CO
l-Ala
d-Glu
m-DAP
d-Ala
d-Ala
m-DAP
d-Glu
l-Ala
Type 2 amidaseAn enzyme that hydrolyses the lactyl-amide bond between the N-acetylmuramic acid and the first amino acid (L-alanine) of the stem peptide in bacterial peptidoglycan.
Teichoic acidA phosphate-rich, anionic polysaccharide that is attached to the peptidoglycan of Gram-positive bacteria. Most are polyglycerol phosphate or polyribitol phosphate and, in the case of lipoteichoic acids, have lipid modifications that allow association with the cytoplasmic membrane.
common primordial ancestor gene. Some PGRPs, most notably mammalian PGLYRP-3 and PGLYRP-4 and D. melanogaster PGRP-LF, have two PGRP domains, however, these domains are not identical. All animals have PGRP domains that are approximately 42% iden-tical and 55% similar. The short PGRPs (invertebrate PGRP-S and vertebrate PGLYRP-1) comprise ~200 amino-acid residues, with a signal peptide and a single PGRP domain. Most long or intermediate size PGRPs (invertebrate PGRP-L and vertebrate PGLYRP-2) are at least twice as large as this and have one C-terminal PGRP domain and an N-terminal sequence of vari-able length that is not conserved and is unique for a
given PGRP. These N-terminal sequences lack easily identifiable functional motifs and therefore might have unique — and so far unknown — functions5–8.
Almost all PGRPs have two closely spaced, conserved Cys residues in the centre of the PGRP domain that form a disulphide bond, which is required for PGRP struc-tural integrity and activity12–20. Most vertebrate PGLYRPs and some invertebrate PGRPs have two additional con-served Cys residues that form a second disulphide bond, and many mammalian PGLYRPs (PGLYRP-1 and the C-terminal PGRP domain of PGLYRP-3 and PGLYRP-4) have another conserved pair of Cys residues that form a third disulphide bond.
Box 1 | Peptidoglycan structure and function
Peptidoglycan is a polymer of β(1-4)-linked N-acetylglucosamine (GlcNAc) and N-acetylmuramic acid (MurNAc), with all lactyl groups of MurNAc substituted with stem peptides, typically containing four alternating l- and d-amino acids. The stem peptides from adjacent strands are often crosslinked, either directly or through short peptides. The structure of the polysaccharide backbone of peptidoglycan is conserved in all bacteria, with only minor variations, such as a different extent of O-acetylation, which provides resistance against lysozyme86, a peptidoglycan-lytic enzyme. By contrast, the composition of the peptide varies: Gram-negative bacteria and Gram-positive bacilli have meso-diaminopimelic acid (DAP) as the third amino acid (DAP-type peptidoglycan), whereas most other Gram-positive bacteria (including Gram-positive cocci) have l-lysine as the third amino acid (Lys-type peptidoglycan; see figure). The DAP-type peptides are usually directly crosslinked, and the Lys-type peptides are usually crosslinked through an interpeptide bridge that varies in length and amino-acid composition in different bacteria.
Peptidoglycan is highly abundant in Gram-positive bacteria, in which it accounts for approximately half of the mass of the cell wall. In Gram-negative bacteria, a relatively thin peptidoglycan layer surrounds the cytoplasmic membrane underneath the lipopolysaccharide (LPS)-containing outer membrane; the peptidoglycan forms one large macromolecule (sacculus) surrounding the entire cell.
The two main functions of peptidoglycan in bacteria are to maintain the shape of the cell and to counteract the high osmotic pressure of the bacterial protoplast — without peptidoglycan or when the integrity of peptidoglycan is compromised, the bacteria undergo immediate osmotic lysis. For this reason, peptidoglycan is a well-known target for almost all clinically useful antibiotics that inhibit bacterial cell-wall synthesis. Peptidoglycan also regulates the diffusion of molecules into cells, participates in the regulation of cell division and is an anchoring structure for cell-wall teichoic acid, proteins and polysaccharides, which are covalently linked to peptidoglycan. Bacteria also often release peptidoglycan fragments, which function as signals in cell–cell communication87.
The three-dimensional arrangement of peptidoglycan in the cell wall is controversial: one model proposes parallel arrangement of the glycan strands in relation to the cell surface88, whereas a second model proposes a perpendicular arrangement of the glycan strands relative to the bacterial surface with a honeycomb-like crosslinking through the peptides89. The second model, which postulates the presence of ~50 to ~500 Å pores in the cell wall, is supported by visualizing such pores by atomic force microscopy90 and by nuclear magnetic resonance analysis of synthetic peptidoglycan fragments91.
Peptidoglycan or its fragments are recognized by the host and induce many biological effects, including inflammation, accumulation of pus, arthritis, fever, sleepiness, decreased appetite, hypotension, leukocytosis, thrombocytopenia, toxicity to tracheal epithelial cells (caused by tracheal cytotoxin, a peptidoglycan fragment released by Bordetella pertussis) and enhanced immune responses (adjuvant effect)92,93. A combination of peptidoglycan and lipoteichoic acid induces shock and multiple organ failure94. Most of these effects are induced indirectly by stimulation of the production of various mediators, such as cytokines and chemokines. Peptidoglycan is also active in invertebrates, for example, in the squid Euprymna scolopes peptidoglycan fragments, such as tracheal cytotoxin, trigger development of the light-emitting organ that contains a symbiotic fluorescent bacterium, Vibrio fischeri87,95. The figure shows the structures of DAP-type (a) and Lys-type (b) peptidoglycan.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | APRIL 2007 | 265
© 2007 Nature Publishing Group

PlasmatocyteThe main cell type found in insect haemolymph.
Some insect PGRPs, for example, D. melanogaster PGRP-LC (FIG. 1), are transmembrane proteins whereas most other insect PGRPs either have a signal peptide and are secreted or do not have a signal peptide and are therefore either intracellular or are secreted by another mechanism. All mammalian PGLYRPs are secreted and form homodimers that, for PGLYRP-1, PGLYRP-3 and PGLYRP-4, are disulphide-linked21–23. Moreover, if PGLYRP-3 and PGLYRP-4 are expressed in the same cells, they form disulphide-linked heterodimers almost exclusively22. Insect PGRPs have not been shown to form disulphide-linked dimers, but ligand binding can induce dimerization or oligomerization24,25 (FIG. 1d,e).
Insect PGRPs — inducers of signalling cascadesThe insect immune system comprises three major defence mechanisms26. The first mechanism is the activation of the prophenoloxidase pathway. Phenoloxidase enzymatic activity is required to synthesize melanin, which is depos-ited at the injury site. This process is thought to release cytotoxic reactive oxygen species that affect invading microorganisms. The second mechanism is the cellular arm of the response, which disposes of microorganisms by phagocytosis27. This task is carried out by the major haemolymph cell type in insects, the plasmatocyte. The third defence mechanism is the humoral response28. Its effectors are small cationic antimicrobial peptides that
Table 1 | Expression sites and functions of Drosophila melanogaster and human PGRPs
PGRP Expression site Function
D. melanogaster
PGRP-LA-C* Haemocytes ND
PGRP-LA-D (a) Haemocytes ND
PGRP-LA-E Haemocytes ND
PGRP-LA-F (b) Haemocytes ND
PGRP-LB-A Fat body (induced), haemolymph Amidase16,50
PGRP-LB-B Fat body (induced), haemolymph Predicted amidase‡
PGRP-LB-C Fat body (induced), haemolymph Predicted amidase
PGRP-LC-A (x) Haemocytes, fat body Imd activation25,36–41, phagocytosis38
PGRP-LC-B (a) Haemocytes, fat body Imd activation25,36–38,40,41
PGRP-LC-C (y) Haemocytes, fat body Imd activation40
PGRP-LD-A Haemocytes ND
PGRP-LE Gut, haemocytes, haemolymph, trachea Imd and PPO activation42
PGRP-LF ND ND
PGRP-SA Fat body, haemolymph, epidermis Toll activation20, carboxypeptidase13,
phagocytosis49
PGRP-SB1 Fat body (induced), haemolymph Amidase, bactericidal53||
PGRP-SB2 ND Predicted amidase
PGRP-SC1a§ Gut Amidase52, Toll activation49, phagocytosis49
PGRP-SC1b§ Gut Amidase52
PGRP-SC2 Gut, fat body (induced) Predicted amidase
PGRP-SD Fat body (induced), haemolymph Toll activation35
Human
PGLYRP-1 (PGRP-S) PMN granules Bactericidal22,62
PGLYRP-2 (PGRP-L) Liver (secreted into blood), induced in
epithelial cells (skin, oral, intestinal)
Amidase19,21,23
PGLYRP-3 (PGRP-Iα) Skin, sweat glands, sebaceous glands, mouth,
intestinal tract, eyes
Bactericidal22,62
PGLYRP-4 (PGRP-Iβ) Skin, sweat glands, sebaceous glands, salivary
glands, mouth, intestinal tract, eyes
Bactericidal22,62
*Current nomenclature of D. melanogaster PGRP-LA, -LB and -LC isoforms (-A, -B) is indicated. Previous names are also included, indicated by lower case letters in parentheses. For PGRP-LD, isoforms -A, -B and -C have the same amino-acid sequence, and only isoform A is shown. ‡Amidase activities were predicted based on the presence of all four Zn2+-binding amino acids and other amino acids required for the amidase activity as described19,51,52. §D. melanogaster PGRP-SC1a and -SC1b are coded by two adjacent genes translated into proteins with identical amino-acid sequences. ||Narrow spectrum bactericidal activity owing to its amidase activity. ND, not determined; PMN, polymorphonuclear leukocytes; PPO, prophenoloxidase; PRGP or PGLYRP, peptidoglycan recognition proteins.
R E V I E W S
266 | APRIL 2007 | VOLUME 5 www.nature.com/reviews/micro
© 2007 Nature Publishing Group

d PGRP-LCx–PGRP-LCa–TCT e PGRP-LCx–PGN
b PGLYRP-3 c PGLYRP-3–MPP
90°
N terminus of PGRP domain
PGRP-LCx
IMD pathway signalling
a PGRP-LE–TCT
Polymeric PGN
Sugar chainPeptide stem
Cell membraneCytoplasmicdomain
α2
Figure 1 | PGRP structure. The figure shows PGRPs complexed with peptidoglycan fragments and the ligand-induced conformational changes, dimerization and oligomerization. A ribbon diagram of the Drosophila melanogaster PGRP-LE carboxy terminus complexed with a monomeric peptidoglycan fragment, tracheal cytotoxin (TCT; GlcNAc-1,6-anhydro-MurNAc-l-Ala-d-isoGlu-DAP-d-Ala) (a). The molecular surface of the carboxy-terminal PGRP domain of human PGLYRP-3 in a free form (b) and complexed with N-acetylmuramic acid (MurNAc)-pentapeptide (MPP) (c), which induces a conformational change in the PGRP domain (green) that locks the ligand in the peptidoglycan-binding groove. TCT induces dimerization between the ectodomains of D. melanogaster PGRP-LCx (green, space filling) and PGRP-LCa (blue ribbon) by binding to the peptidoglycan-binding groove of PGRP-LCx and the second α-helix (α2) of PGRP-LCa, viewed perpendicular (top) and parallel (bottom) to the surface of the cell membrane (d). Binding of polymeric peptidoglycan (PGN) to D. melanogaster PGRP-LCx on the cell surface induces PGRP-LCx oligomerization, viewed perpendicular (top) and parallel (bottom) to the surface of the cell membrane. PGRP-LC dimerization or oligomerization induces activation of the IMD (for immune deficiency) pathway (e). PGRP or PGLYRP, peptidoglycan recognition protein. Parts a and e reproduced with permission from REF. 79 © American Society for Biochemistry and Molecular Biology. Parts b and c reproduced with permission from REF. 17 © American Association for the Advancement of Science. Part d reproduced with permission from REF. 80 © Cold Spring Harbor Laboratory Press.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | APRIL 2007 | 267
© 2007 Nature Publishing Group

TollSpaetzle
Pro-spaetzle
Spe
DIF
DIFCactus
Kinasecomplex
Cactus
SphinxSpheroideSpirit
PGRP-SA
PGRP-SDGNBPI
GrassPersephone
PAMP?
Lys-typepeptidoglycan
Fungalcell
Antimicrobial peptides
Drosomycin
DIFProteasome
P
dMyD88
TubePelle
are produced by the fat body (the insect equivalent of the vertebrate liver), and by haemolymph and epithelial cells. The peptides are then released into the circulation, where they destroy invading microorganisms by disrupting their cell membrane29.
Antimicrobial peptides can be divided into three groups, according to their main microbial targets: antifungal peptides, which include drosomycin and metchnikowin; peptides that are active against Gram-negative bacteria, such as diptericin, attacins, cecropins and drosocin; and peptides that are active against Gram-positive bacteria, such as defensin. Interestingly, it seems that the type of antimicrobial peptide produced by D. melanogaster can be adapted, to some degree, to the invading microorganism. For example, fungi and Gram-positive bacteria are better elicitors of Drosomycin gene expression than Gram-negative bacteria30. By con-trast, infection by Gram-negative bacteria preferentially induces the expression of the Diptericin, Attacin, Cecropin and Drosocin genes.
Activation of any of these three immune mechanisms requires that the invading microorganisms are identi-fied and PGRPs have a role in microbial recognition upstream of each of these facets of the insect immune response.
PRRs upstream of immune signalling pathways. In D. melanogaster, the expression of the genes encoding most of the proteins involved in immunity is under the control of two distinct signalling pathways, both of which activate members of the nuclear factor κB (NF-κB) family of transcription factors31 (BOX 2). The Toll pathway (FIG. 2) responds to Gram-positive bacteria and fungal pathogens, whereas the IMD (for immune deficiency) pathway (FIG. 3) preferentially recognizes Gram-negative bacteria. Both of these pathways have similarities with the signalling cas-cades involved in mammalian innate immunity: the Toll pathway resembles the Toll-like receptor/interleukin 1 receptor (TLR/IL-1R) pathway, whereas the IMD path-way is more similar to the tumour necrosis factor (TNF) receptor cascade32. However, unlike their mammalian counterparts, the Toll and IMD pathways in insects rely on PGRPs to detect bacterial and fungal pathogens.
Box 2 | Drosophila melanogaster immune signalling pathways
The D. melanogaster Toll receptor is activated on binding its mature Spaetzle ligand (FIG. 2). Spaetzle maturation requires an upstream serine protease cascade that culminates in the activation of the Spaetzle processing enzyme (Spe). Some proteases are specifically required to transduce the signal from Gram-positive bacteria (Grass) or fungi (Persephone). Others such as Spirit, Sphinx and Spheroide, integrate signals from both types of microorganism. After binding to Spaetzle, Toll dimerizes and interacts with the D. melanogaster homologue dMyD88 through its TIR domain. dMyD88 recruits Tube, which in turn activates Pelle through their death domains. This leads to the activation of an as-yet-unidentified kinase complex that phosphorylates the IκB inhibitor Cactus, which becomes degraded, releasing the Rel transcription factor DIF.
The D. melanogaster IMD (for immune deficiency) pathway is initiated by dimerization of the PGRP-LC transmembrane receptor after peptidoglycan recognition (FIG. 3). Intracellular signalling requires the adaptor protein IMD, dFADD and the caspase Dredd. Dredd has a dual function in the IMD pathway. It is required for the activation of the IKK signalosome complex, but it is also directly involved in the cleavage of the nuclear factor κB (NF-κB) family member Relish. Signalling also requires the MAPKKK Tak1 and its partner TAB2, which are needed for activation of both the NF-κB and the JNK MAPK branches. Activated TAK1 is thought to act on the IKK signalosome to promote Relish phosophorylation, an essential step in Dredd-mediated cleavage of Relish. Once cleaved, the N-terminal region of Relish translocates into the nucleus to stimulate anti-bacterial gene expression. The JNK pathway regulates the expression of immune-induced genes that are involved in apoptosis, wound healing and the stress response.
Figure 2 | Activation of the Drosophila melanogaster Toll pathway. The Toll receptor is activated after binding its mature ligand Spaetzle. Spaetzle is cleaved by proteolytic cascades that are preferentially triggered by Gram-positive (the Grass protease) or by fungal (the Persephone protease) infection. Upstream of the Grass protease, Lys-type peptidoglycan is recognized either by the PGRP-SA–GNBP1 complex or by PGRP-SD. Intracellular signal transduction requires three death-domain-containing adaptor molecules. Formation of this protein complex leads to the activation of an as-yet-unidentified kinase complex that phosphorylates the IκB inhibitor Cactus, which is then degraded, releasing the Rel transcription factor DIF for nuclear uptake. The transcription of antimicrobial peptides, such as drosomycin, that are active against Gram-positive bacteria and fungi is induced, along with hundreds of additional genes. See BOX 2 for more details.
R E V I E W S
268 | APRIL 2007 | VOLUME 5 www.nature.com/reviews/micro
© 2007 Nature Publishing Group

PGRP-LC
IKK signalosome
DAP-type peptidoglycan
JNK
TAK1TAB2
IMDFADD
Dredd
Apoptotic genes
Antimicrobial peptides
Diptericin
P
P
P
Relish
Monomericpeptidoglycan
Polymeric peptidoglycanInactivepeptidoglycan
PGRP-LE
PGRP-LCa PGRP-LCx
PGRP-LCy
PGRP-LB PGRP-SCI/2
a
b
PGRP-LCx
The first in vivo functional analysis of a D. mela-nogaster PRR was reported by Michel and colleagues20. They isolated a genetically null mutant in the PGRP-SA gene, which was impaired in its ability to activate the Toll pathway and was highly susceptible to infection by Gram-positive bacteria. This mutation dissected the Toll pathway into a fungal and a bacterial branch, as the mutant responded normally to fungal challenge. Induction of the IMD-pathway-dependent antimicro-bial peptides was also normal in the PGRP-SA mutant, demonstrating that PGRP-SA is specifically required for the detection of Gram-positive bacteria and that this protein functions upstream of the Toll pathway (FIG. 2). Subsequent binding studies and structure resolution at 2.2 and 1.56 Å have shown that PGRP-SA is a bona fide PRR, which binds specifically to peptidoglycan13,15.
Detection of Gram-positive bacteria upstream of the Toll pathway involves other molecules in addition to PGRP-SA (FIG. 2). A mutation in a circulating protein named Gram-negative binding protein 1 (GNBP1) gave rise to an unexpected PGRP-SA-like phenotype. GNBP1, which shares sequence homology with gluca-nases, is probably an enzyme that degrades Gram-posi-tive peptidoglycan and generates free reducing ends of N-acetylmuramic acid (MurNAc), which are then recognized by PGRP-SA33,34. It is also clear that some Gram-positive species activate the Toll pathway in a PGRP-SA-independent, GNBP1-independent manner, indicating that alternative receptors exist. Consistent with this notion, PGRP-SD cooperates with PGRP-SA and GNBP1 to allow the detection of some Gram-positive bacterial species, such as Staphylococcus aureus35.
Activation of the IMD pathway also results in the gen-eration of antimicrobial peptides. The strong similarities between the mammalian TNF and D. melanogaster IMD pathways suggested that a TNF-receptor-like molecule could be the upstream component of this arm of the D. melanogaster immune response to Gram-negative bacteria. However, three independent studies demon-strated that a member of the PGRP family, PGRP-LC, is the main transmembrane receptor that activates the IMD pathway36–38 (FIGS 1,3). D. melanogaster mutants that carry a mutation in the PGRP-LC gene are unable to transcribe IMD-pathway-dependent genes after micro-bial challenge and are highly susceptible to infection with Gram-negative bacteria.
The activation of PGRP-SA by Gram-positive bac-teria, and PGRP-LC by Gram-negative bacteria, raised the question: what is the actual discriminating factor in peptidoglycan recognition by these distinct proteins? The presence of a lysine residue in the stem peptides of peptidoglycan (Lys-type peptidoglycan) in Gram-posi-tive bacteria (BOX 1) favours interaction with PGRP-SA, and therefore Toll activation, whereas the replacement of lysine by meso-diaminopimelic acid (DAP) in Gram-negative bacterial peptidoglycan (DAP-type peptidog-lycan) leads to an interaction with PGRP-LC and the subsequent activation of the IMD pathway39. Therefore, the ability of D. melanogaster to discriminate between Gram-positive and Gram-negative bacteria does not rely on the detection of lipopolysaccharide (LPS; a unique
Figure 3 | Activation of the Drosophila melanogaster IMD pathway by bacteria. a | DAP-type peptidoglycan is recognized by PGRP-LC, which contains a transmembrane domain and activates the IMD (for immune deficiency) pathway. IMD regulates at least three downstream pathways. The first involves activation of TAK1, which then stimulates IKK-dependent cleavage and activation of Relish. The second is the FADD–Dredd pathway that also activates Relish. Both pathways induce the transcription of the genes encoding several antimicrobial peptides, including diptericin (which is active against Gram-negative bacteria), and many other genes. The third event is the activation of the JNK pathway through TAK1, which regulates early response genes, wound healing and apoptosis. b | The D. melanogaster immune system can discriminate between various polymeric and monomeric peptidoglycan fragments. Polymeric peptidoglycan is recognized by a heterodimeric complex of PGRP-LCa and PGRP-LCx. Monomeric peptidoglycan fragments (such as the tracheal cytotoxin, TCT) bind to PGRP-LCx, inducing a conformation change that allows the formation of PGRP-LCx–PGRP-LCa heterodimers. TCT is also detected intracellularly by PGRP-LE. An extracellular form of PGRP-LE containing only the PGRP domain is thought to participate in peptidoglycan recognition in conjunction with PGRP-LC. The ligand(s) and function(s) of PGRP-LCy are unknown. Removal of the stem peptides from peptidoglycan by the amidases PGRP-SC1/2 and PGRP-LB abolishes the activity of the peptidoglycan fragments. DAP, meso-diaminopimelic acid; PGRP, peptidoglycan recognition protein.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | APRIL 2007 | 269
© 2007 Nature Publishing Group

Type IV secretion systemA syringe-like proteinaceous machinery that can transport bacterial protein or DNA effector molecules directly into a eukaryotic cell.
component of the outer membrane that is present only in Gram-negative bacteria), but rather on the recognition of specific forms of peptidoglycan39,40. Because DAP-type peptidoglycan is also found in Gram-positive rod-shaped bacteria, PGRP-LC can also recognize these bacteria in addition to all Gram-negative bacteria.
Two different elicitors have been shown to activate the IMD pathway in vivo. In adult D. melanogaster, strong antimicrobial peptide production is triggered by the injection of either a polymeric DAP-type peptidogly-can or a monomeric DAP-type peptidoglycan fragment, such as tracheal cytotoxin (TCT), which is secreted by Bordetella pertussis and has the following composition: GlcNAc-1,6-anhydro-MurNAc-l-Ala-γ-d-Glu-meso-DAP-d-Ala40,41. Interestingly, these two muropeptides are recognized in vivo by two different alternatively
spliced forms of PGRP-LC5, PGRP-LCx and PGRP-LCa (FIGS 1,3b; discussed later). A function and ligand have not yet been identified for the third PGRP-LC isoform, PGRP-LCy.
Gram-negative bacteria and DAP-type peptidoglycan are also sensed in vivo by the non-membrane-bound PGRP-LE receptor, which seems to have two distinct functions42,43. Similar to the role of the Nod1 and Nod2 mammalian proteins (BOX 3), full-length PGRP-LE is an intracellular receptor for monomeric peptidoglycan that is independent of the transmembrane PGRP-LC44 (FIG. 3b). It is not known how peptidoglycan fragments such as TCT gain access to intracellular PGRP-LE. Participation of the bacterial type IV secretion system, which has been shown to deliver peptidoglycan fragments from Helicobacter pylori into mammalian cells45, can be proposed. In addition, an
Box 3 | Peptidoglycan recognition in mammals
Peptidoglycan is an excellent target for recognition by the eukaryotic innate immune system, because it is an essential and unique component of the cell walls of virtually all bacteria and is not present in eukaryotic cells. Higher eukaryotes, including mammals, have several peptidoglycan recognition molecules, which are cell surface or intracellular pattern recognition receptors (PRRs) and soluble recognition and effector molecules (FIG. 6).
Extracellular recognition of peptidoglycanExtracellular recognition of peptidoglycan is mediated by membrane-bound CD14 (REFS 96,97) and Toll-like receptor 2 (TLR2)98–100. CD14 and TLR2 are cell-surface PRRs mostly present on monocytes, macrophages and dendritic cells (FIG. 6). Their activation results in the secretion of pro-inflammatory cytokines (such as tumour necrosis factor, interleukin (IL)-1 and IL6) and chemokines (such as IL8). This inflammatory response helps to limit infection before the adaptive immune response develops. Activation of TLRs on antigen-presenting cells (dendritic cells, macrophages and B cells) is also required for the initiation of adaptive immune responses, by inducing maturation of antigen-presenting cells, secretion of cytokines and expression of co-stimulatory molecules that are required for antigen presentation and activation of helper T cells. CD14 and TLR2 recognize multiple microbial components. High molecular weight polymeric peptidoglycan is recognized by CD14 and TLR2 (REFS 96,101,102). The recognition of peptidoglycan by TLR2 has been questioned103, but a more recent re-evaluation confirmed that peptidoglycan is a TLR2 activator104. In intact bacteria, it is likely that peptidoglycan acts as a scaffold for other TLR2 activators, such as lipopeptides105.
Intracellular recognition of peptidoglycanIntracellular recognition of peptidoglycan is mediated by two members of the Nod-like receptor (NLR) family, Nod1 and Nod2 (FIG. 6). Nod1 senses Gram-negative bacteria and some Gram-positive rods by recognizing peptidoglycan fragments containing meso-diaminopimelic acid (DAP). l-Ala-d-Glu-DAP is the minimum peptidoglycan fragment recognized by Nod1 (REFS 106,107). The minimum peptidoglycan fragment recognized by Nod2 is muramyl dipeptide (N-acetylmuramyl-l-alanyl-d-isoglutamine; MDP)106,108,109. Because MDP is present in all peptidoglycans, Nod2 can sense peptidoglycan from all bacteria. It is still unclear whether Nods directly bind peptidoglycan or whether they interact only with peptidoglycan fragments or polymeric peptidoglycan. Many bacteria secrete soluble peptidoglycan fragments that could be Nod activators87, and Nods can detect intracellular bacteria that escape into the cytoplasm, such as Salmonella, Shigella and Listeria species. It is also still unclear how extracellular bacteria and extracellular peptidoglycan fragments reach cytoplasmic Nods, although some bacteria, such as Helicobacter pylori, can deliver peptidoglycan into the host cell cytoplasm using a secretion system45.
Nod1 is ubiquitously expressed in many tissues and cell types, whereas Nod2 is highly expressed in monocytes, macrophages, dendritic cells and, to a lesser extent, in epithelial cells110. Nod1 and Nod2 mediate activation of nuclear factor-κB through an association with a serine-threonine kinase, RICK110. This activation is TLR-independent and is therefore an alternative pro-inflammatory pathway, and these two activation pathways are often synergistic. The susceptibility to some pro-inflammatory diseases that are probably triggered by bacteria, most notably Crohn’s disease and Blau syndrome, is associated with mutations in the Nod2 gene111–113.
Soluble peptidoglycan recognition moleculesSoluble peptidoglycan recognition molecules (FIG. 6) include: peptidoglycan recognition proteins (PGRPs; amidase and bactericidal PGRPs), which are the subject of this Review; soluble CD14, which is present in serum and enhances cell activation by peptidoglycan114; C-type lectins, which are specific for the glycan region of peptidoglycan, including mannose-binding lectin, which activates complement and modulates responses to peptidoglycan115,116, and mouse RegIIIγ and human HIP/PAP, which are secreted from Paneth cells in the intestine and are bactericidal117; and lysozyme, which is a muramidase that is present in phagocytic granules, serum and body secretions70 that hydrolyses the glycosidic bond between N-acetylmuramic acid (MurNAc) and N-acetylglucosamine (GlcNAc) in peptidoglycan. O-acetylation makes peptidoglycan resistant to lysozyme86. Lysozyme also has an antibacterial effect that is independent of its peptidoglycan-hydrolytic activity118.
R E V I E W S
270 | APRIL 2007 | VOLUME 5 www.nature.com/reviews/micro
© 2007 Nature Publishing Group

PGRP-LB
PGRP-SB1
PGLYRP-2
PGRP-SC1
(–GlcNAc–MurNAc–)n
(–GlcNAc–MurNAc–)n
L-Ala
D-Ala
D-Ala
D-Glu
m-DAP
m-DAP
D-Glu
L-Ala
PGRP-SA
DrosophilaN-acetylmuramoyl-L-Ala amidases
DrosophilaCarboxypeptidase
MammalianN-acetylmuramoyl-L-Ala amidase
OpsoninA substance that binds to microorganisms and other cells and enhances their phagocytosis.
RNA interference(RNAi). The use of double-stranded RNAs with sequences that precisely match a given gene, to ‘knock-down’ the expression of that gene by directing RNA-degrading enzymes to destroy the encoded mRNA transcript.
extracellular version of PGRP-LE containing only the PGRP domain seems to facilitate peptidoglycan recog-nition by PGRP-LC, although the molecular mechanism involved in this is unknown44.
Insect PGRPs and the prophenoloxidase pathway. In addition to the antimicrobial peptides that are gener-ated through the activation of the Toll and IMD path-ways, PGRPs from various insects, such as B. mori1, the mealworm Tenebrio molitor46, the beetle Holotrichia diomphalia47 and D. melanogaster43, trigger the proph-enoloxidase cascade. Overexpression of the PGRP-LE receptor in D. melanogaster is sufficient to induce the prophenoloxidase cascade, whereas PGRP-LE loss-of-function mutants are unable to activate this melanization cascade after Escherichia coli infection42,43. In vitro recon-stitution experiments have shown that T. molitor PGRP functions as a common recognition molecule upstream of the Lys-type and DAP-type peptidoglycan-depend-ent melanization cascades46. It is still unclear, however, how the recognition of microbial motifs by these PGRPs translates into activation of this protease cascade.
Insect PGRPs and phagocytosis? In addition to the above-mentioned cell-activating functions, PGRP-LC and PGRP-SC1 might also be involved in phagocytosis. Downregulation of PGRP-LC levels in D. melanogaster S2 blood-like cells diminishes uptake of E. coli but not S. aureus, although the extent of this inhibition is much less than that observed with the well-characterized phago-cytic receptor Eater38,48. By contrast, in D. melanogaster mutants with reduced PGRP-SC1 levels, phagocytosis of S. aureus is impaired49. However, it remains to be elucidated how peptidoglycan, which in Gram-negative bacteria is buried beneath the LPS-containing outer membrane, can be recognized by the transmembrane receptor PGRP-LC to allow phagocytosis to take place. It is also unclear how the secreted protein PGRP-SC1 functions in phagocytosis. One possibility could be that PGRP-SC1 functions as an opsonin that binds bacteria, and then enables or enhances recognition of this com-plex by a transmembrane phagocytic receptor. Further work is needed to test these hypotheses and to define the mechanisms through which insect PGRPs influence phagocytosis.
PGRPs with peptidoglycan-hydrolytic activityThe only conserved common function of insect and mammalian PGRPs is the N-acetylmuramoyl-l-alanine amidase enzymatic activity, which hydrolyses the amide bond between MurNAc and l-alanine in peptidoglycan and removes the stem peptides from the glycan chain (FIG. 4). D. melanogaster PGRP-SC1, PGRP-SB1 and PGRP-LB, and mammalian (human, mouse and swine) PGLYRP-2 have proven amidase activity9,19,50–53. Many other PGRPs are predicted to be amidases based on conserved amino acids in the amidase active site.
Although the role of PGRPs with PRR function has been well documented in D. melanogaster, the role of the hydrolytic PGRPs has been elusive. Steiner’s group has shown that in vitro hydrolysis of peptidoglycan by
PGRP-SC1 produces muropeptides that have no immu-nostimulatory properties52. They suggested that amidase PGRPs could function as scavengers in vivo, leading to the termination of the immune response. Recent in vivo studies support this hypothesis. D. melanogaster in which the soluble catalytic PGRP-SC1 or PGRP-LB have been downregulated by RNA interference present with selective over-activation of the IMD pathway after bacterial challenge50,54. More interestingly, systemic tolerance to ingested Gram-negative bacteria or to pep-tidoglycan was observed in wild-type D. melanogaster but not in D. melanogaster depleted of PGRP-SC1/2 or PGRP-LB. This indicates that these proteins, which are mainly expressed in the gut, maintain the homeo-static balance of the immune response to commensal and pathogenic bacteria. Therefore, in addition to their function as PRRs, some PGRPs can control the level of activation of the IMD signalling pathway by decreasing the immunostimulatory potential of pepti-doglycan (FIG. 5). In the absence of this control, infection can lead to developmental defects and larval death by over-activation of the immune pathway or induction of apoptosis54. In this respect, PGRPs with amidase activity act as ‘detoxifying’ enzymes for bacterial peptidoglycan in D. melanogaster. Similar functions have been attrib-uted to enzymes that reduce the immunostimulatory potential of LPS in vertebrates55.
One D. melanogaster amidase, PGRP-SB1, is also directly bactericidal, but only against one bacterial spe-cies (Bacillus megaterium) and not against other, even closely related, bacterial species53. This bactericidal activity is probably the result of hydrolysis of peptidog-lycan in the cell wall, and the selective sensitivity of this one bacterium is probably related to the accessibility of
Figure 4 | The enzymatic activity of PGRPs. The bonds cleaved in bacterial peptidoglycan by Drosophila melanogaster and mammalian PGRPs with N-acetylmuramoyl-l-alanine amidase or carboxypeptidase activities are shown. PGRP, peptidoglycan recognition protein.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | APRIL 2007 | 271
© 2007 Nature Publishing Group

Fat body cells(express Diptericin)
PGRP-LCPGRP-LC
PGRP-LB PGRP-SC1/2
Active peptidoglycan
Inactive peptidoglycan
Apoptotic cell death
Non-immune tissues
b
a
Bacteria
Gut epithelium
Chitin
Epidermis
Polymorphonuclear leukocyte (PMN). White blood cell with multilobed nuclei and cytoplasmic granules, which is involved in inflammatory responses.
M cell(Microfold cell). Cell type located in the Peyer’s patches of the small intestine. Involved in antigen transport and interaction with bacterial pathogens.
Peyer’s patches Specialized lymphoid follicles localized in the submucosa of the small intestine. They contain B-cell follicles and interfollicular T-cell areas, with an outer epithelial layer containing M cells.
its peptidoglycan to the enzyme. This contrasts with the bactericidal activity of mammalian PGRPs (described below), which have broad-spectrum activity against most Gram-positive and Gram-negative bacteria that does not rely on amidase activity.
One PGRP that is not an amidase, D. melanogaster PGRP-SA, has an l,d-carboxypeptidase activity with specificity for the bond between m-DAP and d-alanine of the stem peptide present in peptidoglycan of all Gram-negative bacteria and Gram-positive rods13 (FIG. 4). The biological significance of this carboxypeptidase activity is unclear.
When mammalian PGLYRP-2 was first cloned, it was thought to be a transmembrane cell-activat-ing receptor6. However, PGLYRP-2 turned out to be constitutively expressed in the liver and secreted into the bloodstream19,21,51. PGLYRP-2 is identical with the earlier identified but, at the time, not cloned serum N-acetylmuramoyl-l-alanine amidase21,23. PGLYRP-2 expression can be induced by bacteria and cytokines in epithelial cells, such as skin keratinocytes56 or intestinal
follicle-associated epithelial cells57. The constitutive expression of PGLYRP-2 in the liver and induced expres-sion in epithelial cells is regulated by different transcrip-tion factors, the binding sequences for which are located in different regions of the pglyrp2 promoter58. The func-tion of mammalian PGLYRP-2 is likely to be similar to the function of insect amidases, that is, to eliminate the pro-inflammatory peptidoglycan and, therefore, prevent over-activation of the immune system and excessive inflammation.
All amidase-active PGRPs (in invertebrates and vertebrates) have a conserved Zn2+-binding site in the peptidoglycan-binding groove, which is also present in bacteriophage type 2 amidases and comprises two His residues, one Tyr residue and one Cys residue (Cys168 in D. melanogaster PGRP-SC1 and Cys530 in human PGLYRP-2). In non-amidase PGRPs, this Cys residue is substituted with Ser19,51,52. Therefore, the amidase activ-ity of PGRPs can be predicted based on the presence of this Cys residue, other Zn2+-binding amino acids and four other required amino acids. In the carboxypepti-dase-active PGRPs however, these Zn2+-binding amino acids are not conserved13. The substrate requirements for amidase activity correspond to the binding capacity of PGRPs for peptidoglycan fragments: the minimum pep-tidoglycan fragment hydrolysed by human PGLYRP-2 is the MurNAc-tripeptide19, similar to the minimal PGRP-binding fragment17,59,60.
Mammalian PGRPs — bactericidal proteinsHumans, mice, rats and probably other mammals have four PGLYRPs. PGLYRP-2, as described above, is an amidase but what are the functions of the remaining mammalian PGLYRPs?
PGLYRP-1 is expressed primarily in polymorpho-nuclear leukocyte (PMN) granules and is directly bac-tericidal for both Gram-positive and Gram-negative bacteria10,22,61,62. PGLYRP-1 knockout mice are more susceptible to infections with some Gram-positive bacte-ria, and their PMNs are defective in killing and digesting Gram-positive bacteria63. Therefore, PGLYRP-1 is part of the antibacterial arsenal of PMNs (FIG. 6). PGLYRP-1 is also expressed in M cells57, which are present in intestinal Peyer’s patches and which transport bacteria and antigens to the intestinal submucosal antigen-presenting cells and lymphocytes. The role of PGLYRP-1 in M cells is unknown, but it might protect M cells and the intestinal submucosa from infections.
PGLYRP-3 and PGLYRP-4 are selectively expressed in tissues that come into contact with the environ-ment22,64, such as skin, eyes, oral cavity and intesti-nal tract. Whereas both PGLYRP-3 and PGLYRP-4 are expressed in sebaceous and sweat glands, only PGLYRP-4 is expressed in the salivary glands. Bacteria and bacterial products increase the expression of PGLYRP-3 and PGLYRP-4 in keratinocytes56,58 and oral epithelial cells65.
Human PGLYRP-1, PGLYRP-3, PGLYRP-4 and PGLYRP-3:4 are directly bactericidal for many patho-genic and non-pathogenic Gram-positive and Gram-negative bacteria22,62 (FIG. 6). They are also active in vivo
Figure 5 | Downregulation of the immune response by amidase-active PGRPs in Drosophila melanogaster. D. melanogaster larvae and adults feed on decaying fruits contaminated with bacteria. Actively dividing bacteria present in the gut release peptidoglycan. In wild-type flies (a), the PGRP-SC1/2 and PGRP-LB amidases, which are mainly produced in the gut, hydrolyze this peptidoglycan into fragments that are unable to activate the IMD (for immune deficiency) pathway. This restricts the activation of the immune response by peptidoglycan released from commensal and ingested bacteria. In the absence of such control (b), intact peptidoglycan accumulates in the gut and crosses the gut epithelium by an unknown mechanism. As D. melanogaster has an open circulatory system, peptidoglycan can bind and stimulate the membrane-bound receptor PGRP-LC present on the surface of fat body cells and in non-immune tissue (such as imaginal discs). This is detrimental to the host, as it induces apoptotic cell death, probably through activation of the JNK pathway, which branches downstream of the IMD cascade (FIG. 3a). PGRP-SC1/2 and PGRP-LB are also present in the haemolymph, where they participate in downregulation of the immune response by degrading peptidoglycan (FIG. 3b).
R E V I E W S
272 | APRIL 2007 | VOLUME 5 www.nature.com/reviews/micro
© 2007 Nature Publishing Group

C-type lectinPGLYRP-3:4
PGLYRP-3PGLYRP-4
PGLYRP-2
Complementactivation
Serum MBL
Opsonization
Lysozyme
Amidaseactivity
Bacterial killing
Nod1
TLR2CD14
Synergisticactivation
Production of inflammatory mediators
PGLYRP-1
Nod2
α/β defensin PLA2
Synergism
on mucous membranes in an intranasal S. aureus lung infection in mice22. Each PGLYRP has a different range of bactericidal activity; for example, Gram-nega-tive bacteria are killed most efficiently by PGLYRP-3 and PGLYRP-1, and least efficiently by PGLYRP-4 (REFS 22,62). PGLYRPs are bactericidal (kill 99% of bac-teria) at 0.1-1 µM, and therefore, on a per mol basis, are more active than most antibacterial peptides. PGLYRPs are less bactericidal or only bacteriostatic against normal flora Gram-positive bacteria, which might prevent over-growth of normal flora bacteria on the skin and mucous membranes without causing their complete elimination. The bactericidal activity is probably a general character-istic of these PGLYRPs in all mammals, because their sequences are highly conserved, because they show simi-lar patterns of expression and because bovine PGLYRP-1 is also bactericidal for both Gram-positive and Gram-negative bacteria10,61. Bovine PGLYRP-1, however, at higher concentrations, also has some activity against fungi10.
Bactericidal activity of all human PGLYRPs for both Gram-positive and Gram-negative bacteria requires physiological concentrations of Zn2+ (~2–5 µM), similar to the Zn2+ concentrations in sweat (~17 µM), saliva (~2 µM) and plasma (~15 µM)62. Zn2+ is an essential component of many proteins, and host immu-nity requires Zn2+ — Zn2+-deficient individuals have increased susceptibility to infections and Zn2+ supple-mentation restores or enhances resistance to infections and innate and acquired immune functions66–69.
Body surfaces, as well as phagocytic cells, con-tain numerous antimicrobial peptides70–72. Human PGLYRPs synergistically kill both Gram-positive and Gram-negative bacteria together with several of these antimicrobial peptides, such as phospholipase A2 (PLA2) and α- and β-defensins, when both compounds are present at sub-bactericidal concentrations62. This synergism is likely to greatly enhance antibacterial defences in vivo, because both PGLYRPs and antimi-crobial peptides are found in the same locations in the body, such as PMN granules, the skin, eyes, oral cavity and intestinal tract8,10,22,61,63,70–76. PGLYRPs also func-tion synergistically with bacteriolytic enzymes such as lysozyme and lysostaphin22,62,77.
Mammalian PGLYRP-1, PGLYRP-3 and PGLYRP-4 therefore constitute a new class of bactericidal protein that have a different structure, mechanism of action and expression from the currently known mammalian antimicrobial peptides. PGLYRPs are much larger than vertebrate antimicrobial peptides: they are disulphide-linked glycosylated 44–115 kDa dimers22, in contrast to vertebrate antimicrobial peptides, which are typically 3–15 kDa. PGLYRPs kill bacteria probably by interact-ing with their cell wall, whereas antibacterial peptides kill bacteria at high concentrations by inserting into their cytoplasmic membranes and causing loss of membrane permeability71,72. PGLYRPs require Zn2+ and N-glycosylation for their bactericidal activity whereas antimicrobial peptides usually do not, and PGLYRPs are insensitive to salt, whereas antimicrobial peptides are often salt-sensitive. Moreover, the expression patterns of mammalian PGLYRPs and antimicrobial peptides are different, and some cells that produce large amounts of these peptides, for example, intestinal Paneth cells (which produce defensins, PLA2 and lysozyme), do not express PGLYRPs22,71,72.
Peptidoglycan recognition by PGRPsPGRPs function as peptidoglycan scavengers, direct bac-tericidal molecules and receptors that link peptidoglycan recognition to the activation of proteolytic cascades or other intracellular signalling pathways. To carry out these functions, PGRPs have evolved several structurally distinct sites: a highly conserved peptidoglycan-binding site and other more variable sites for interacting with other PGRP molecules and with signalling or effector molecules.
The crystal structures of several free and ligated D. melanogaster and human PGRPs revealed a peptidog-lycan-binding groove on the face of the PGRP domain, the walls of which are formed by two α-helices and the floor by a β-sheet platform12–18,78–80 (FIG. 1). The minimum
Figure 6 | The many functions of mammalian peptidoglycan recognition molecules. The secreted peptidoglycan recognition molecules include: PGLYRP-3, PGLYRP-4 and PGLYRP-3:4 homodimers and heterodimers, which directly kill bacteria and act synergistically with membrane-damaging defensins and phospholipase A2 (PLA2); the C-type lectins (which are also directly bactericidal); the PGLYRP-2 amidase, which hydrolyses peptidoglycan in the positions shown; and mannose-binding lectin (MBL), which activates complement and induces opsonization and inflammation. Extracellular polymeric peptidoglycan is recognized by cell-surface receptors TLR2/6 and CD14, which induce secretion of pro-inflammatory mediators and initiate an adaptive immune response. Cytoplasmic receptors Nod1 and Nod2 recognize intracellular peptidoglycan fragments and activate cells synergistically with TLR2. Nod, nucleotide-binding oligomerization domain (containing protein); PGLYRP, peptidoglycan recognition protein; TLR, Toll-like receptor.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | APRIL 2007 | 273
© 2007 Nature Publishing Group
Dm PGRP-LEDm PGRP-LCxHs PGLYRP-1Hs PGLYRP-2Hs PGLYRP-3Hs PGLYRP-3Hs PGLYRP-4Hs PGLYRP-4
21237167
41957
21489
246
273432129
480118
275150307
DAPDAPDAPDAPDAP/LysLys?/Lys?/Lys
SEKRAINVRLIRDMQCFHIESRGWNDIAYNFLVGCDGNIYEGRGWKTVGAHTLG-YNRISLGI
CESREVCSARVNVVQSFHMDSWGWDHIGYNFLVGGDGRVYEGRGWDYVGAHTKG-YNRGSIGI
CNTPASCQQQARNVQHYHMKTLGWCDVGYNFLIGEDGLVYEGRGWNFTGAHSGHLWNPMSIGI
CNISDECRLLVRDIQSFYIDRLKSCDIGYNFLVGQDGAIYEGVGWNVQGSSTPG-YDDIALGI
CTDFTRCAANMRSMQRYHQDTQGWGDIGYSFVVGSDGYVYEGRGWHWVGAHTLG-HNSRGFGV
CQQQSVCSQMLRGLQSHSVYTIGWCDVAYNFLVGDDGRVYEGVGWNIQGLHTQG-YNNISLGI
CTVSTDCQTVVRNIQSFHMDTRNFCDIGYHFLVGQDGGVYEGVGWHIQGSHTYG-FNDIALGI
CHDQTVCSQRLRELQAHHVHNNSGCDVAYNFLVGDDGRVYEGVGWNIQGVHTQG-YNNISLGF
Specificity
peptidoglycan fragment that binds to PGRPs is a muramyl-tripeptide12–16,18,78–80, whereas a muramyl-dipeptide or a peptide without MurNAc does not bind to PGRPs17,59,60. A muramyl-tripeptide fills approximately one-third of the binding cleft and makes extensive contacts with the 16 residues that line the cleft18. The interface is predomi-nantly hydrophilic but solvent is excluded. Most of the interactions occur with the peptide moiety, but MurNAc is also required for binding. The remaining space in the peptidoglycan-binding groove can be occupied by a longer peptide and by GlcNAc, which in peptidoglycan is bound to MurNAc. Indeed, muramyl-pentapeptide binds with higher affinity than muramyl-tripeptide and induces a conformational change in the PGRP molecule that locks the ligand in the binding cleft, as shown for the C-terminal fragment of PGLYRP-3 (REF. 17) (FIG. 1c).
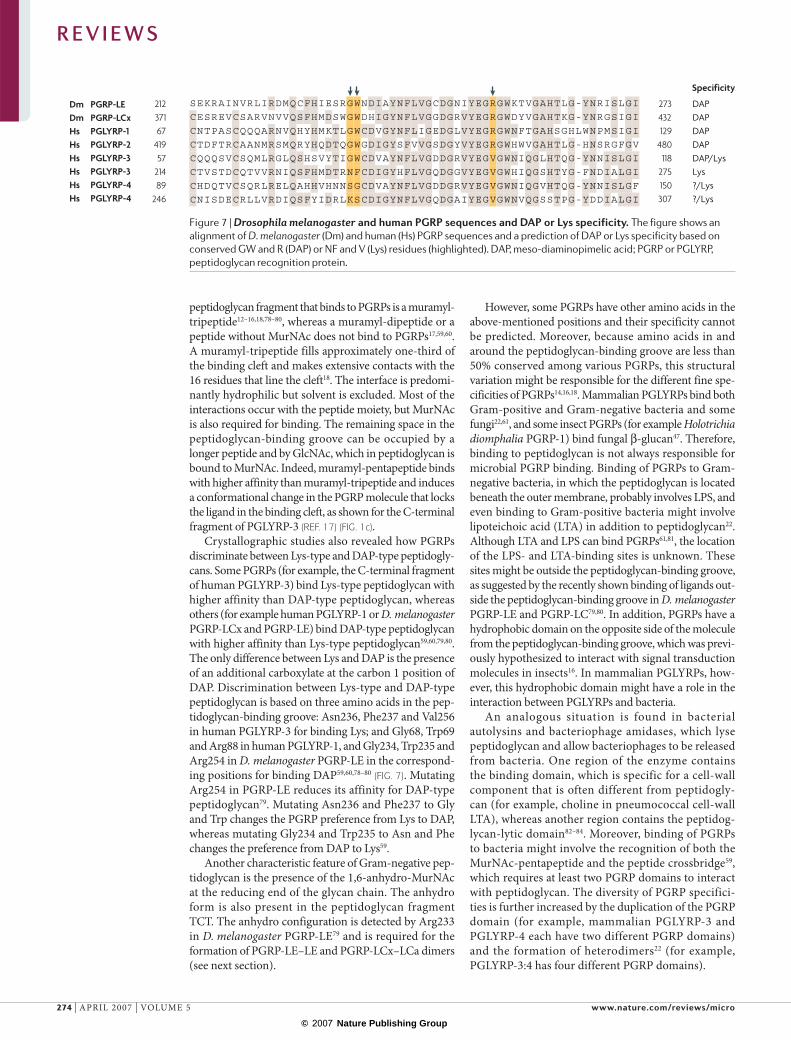
Crystallographic studies also revealed how PGRPs discriminate between Lys-type and DAP-type peptidogly-cans. Some PGRPs (for example, the C-terminal fragment of human PGLYRP-3) bind Lys-type peptidoglycan with higher affinity than DAP-type peptidoglycan, whereas others (for example human PGLYRP-1 or D. melanogaster PGRP-LCx and PGRP-LE) bind DAP-type peptidoglycan with higher affinity than Lys-type peptidoglycan59,60,79,80. The only difference between Lys and DAP is the presence of an additional carboxylate at the carbon 1 position of DAP. Discrimination between Lys-type and DAP-type peptidoglycan is based on three amino acids in the pep-tidoglycan-binding groove: Asn236, Phe237 and Val256 in human PGLYRP-3 for binding Lys; and Gly68, Trp69 and Arg88 in human PGLYRP-1, and Gly234, Trp235 and Arg254 in D. melanogaster PGRP-LE in the correspond-ing positions for binding DAP59,60,78–80 (FIG. 7). Mutating Arg254 in PGRP-LE reduces its affinity for DAP-type peptidoglycan79. Mutating Asn236 and Phe237 to Gly and Trp changes the PGRP preference from Lys to DAP, whereas mutating Gly234 and Trp235 to Asn and Phe changes the preference from DAP to Lys59.
Another characteristic feature of Gram-negative pep-tidoglycan is the presence of the 1,6-anhydro-MurNAc at the reducing end of the glycan chain. The anhydro form is also present in the peptidoglycan fragment TCT. The anhydro configuration is detected by Arg233 in D. melanogaster PGRP-LE79 and is required for the formation of PGRP-LE–LE and PGRP-LCx–LCa dimers (see next section).
However, some PGRPs have other amino acids in the above-mentioned positions and their specificity cannot be predicted. Moreover, because amino acids in and around the peptidoglycan-binding groove are less than 50% conserved among various PGRPs, this structural variation might be responsible for the different fine spe-cificities of PGRPs14,16,18. Mammalian PGLYRPs bind both Gram-positive and Gram-negative bacteria and some fungi22,61, and some insect PGRPs (for example Holotrichia diomphalia PGRP-1) bind fungal β-glucan47. Therefore, binding to peptidoglycan is not always responsible for microbial PGRP binding. Binding of PGRPs to Gram-negative bacteria, in which the peptidoglycan is located beneath the outer membrane, probably involves LPS, and even binding to Gram-positive bacteria might involve lipoteichoic acid (LTA) in addition to peptidoglycan22. Although LTA and LPS can bind PGRPs61,81, the location of the LPS- and LTA-binding sites is unknown. These sites might be outside the peptidoglycan-binding groove, as suggested by the recently shown binding of ligands out-side the peptidoglycan-binding groove in D. melanogaster PGRP-LE and PGRP-LC79,80. In addition, PGRPs have a hydrophobic domain on the opposite side of the molecule from the peptidoglycan-binding groove, which was previ-ously hypothesized to interact with signal transduction molecules in insects16. In mammalian PGLYRPs, how-ever, this hydrophobic domain might have a role in the interaction between PGLYRPs and bacteria.
An analogous situation is found in bacterial autolysins and bacteriophage amidases, which lyse peptidoglycan and allow bacteriophages to be released from bacteria. One region of the enzyme contains the binding domain, which is specific for a cell-wall component that is often different from peptidogly-can (for example, choline in pneumococcal cell-wall LTA), whereas another region contains the peptidog-lycan-lytic domain82–84. Moreover, binding of PGRPs to bacteria might involve the recognition of both the MurNAc-pentapeptide and the peptide crossbridge59, which requires at least two PGRP domains to interact with peptidoglycan. The diversity of PGRP specifici-ties is further increased by the duplication of the PGRP domain (for example, mammalian PGLYRP-3 and PGLYRP-4 each have two different PGRP domains) and the formation of heterodimers22 (for example, PGLYRP-3:4 has four different PGRP domains).
Figure 7 | Drosophila melanogaster and human PGRP sequences and DAP or Lys specificity. The figure shows an alignment of D. melanogaster (Dm) and human (Hs) PGRP sequences and a prediction of DAP or Lys specificity based on conserved GW and R (DAP) or NF and V (Lys) residues (highlighted). DAP, meso-diaminopimelic acid; PGRP or PGLYRP, peptidoglycan recognition protein.
R E V I E W S
274 | APRIL 2007 | VOLUME 5 www.nature.com/reviews/micro
© 2007 Nature Publishing Group

Mechanism of receptor activation and signallingHow is the recognition of ligands by PGRPs trans-lated into a cell-activating signal? The answer to this question came from combining the results of functional and crystallographic studies. First, it was demonstrated that signalling involves the forma-tion of ligand-induced dimers by D. melanogaster PGRP-LC24,25. Subsequent crystallization studies of PGRP-LE in the presence of TCT demonstrated that PGRP-LE–TCT forms oligomers in which TCT is bound in the peptidoglycan-binding cleft (FIG. 1a) of the first PLRP-LE molecule, and through its GlcNAc-1-6-anhydro-MurNAc to the α1-helix of the second PGRP-LE molecule outside the pepti-doglycan-binding cleft79. The peptidoglycan-binding cleft of the second PGRP-LE molecule binds another TCT molecule, which then binds the third PGRP-LE molecule and so on, thereby forming an infinite oli-gomer of PGRP-LE–TCT complexes. The interaction of the α1-helix of the second PGRP-LE molecule with the PGRP-LE–TCT locks the TCT ligand in the pep-tidoglycan-binding cleft and increases the affinity of TCT binding by approximately 30-fold79. Although binding of the second PGRP-LE molecule to the PGRP-LE complex requires that its α1-helix interacts with TCT, it also involves interactions of four residues in the α1-helix and three residues in other parts of the second PGRP-LE molecule with the first PGRP-LE molecule79. However, the functional significance of PGRP-LE oligomerization is still unclear.
Similar interactions of two splice variants of D. melanogaster PGRP-LC (LCx and LCa) with TCT were soon discovered. PGRP-LCx binds TCT in its peptidoglycan-binding cleft, but its GlcNAc-1-6-anhy-dro-MurNAc forms dimers with PGRP-LCa through a similar interaction with the PGRP-LCa α-helix80 (α2 in FIG. 1d). Binding of TCT to PGRP-LCx also induces a conformational change in the PGRP-LCx molecule that allows it to bind several residues of PGRP-LCa. Because both PGRP-LCx and PGRP-L are transmem-brane molecules, the formation of PGRP-LCx–TCT–PGRP-LCa dimers brings their cytoplasmic signalling domains together (FIG. 1d), which is presumed to generate the IMD pathway-activating signal79,80. TCT induces the formation of PGRP-LCx–TCT–PGRP-LCa dimers, but not oligomers, because PGRP-LCa is a splice form with a non-functional (obstructed) peptidoglycan-binding groove that is unable to bind TCT. These results are consistent with earlier func-tional studies showing a requirement for the entire TCT molecule (including its GlcNAc-1-6-anhydro-MurNAc and DAP) for the efficient activation of the IMD pathway 40,41. This dimer formation also increases the affinity of PGRP-LC for TCT, thereby allowing a more sensitive detection of small amounts of this peptidoglycan fragment. PGRP-LC is also activated by polymeric DAP-type peptidoglycan. The current model (FIG. 1e) proposes that polymeric peptidoglycan causes a clustering of PGRP-LCx molecules on the cell surface, which brings the cytoplasmic domains together79.
Little information is yet available on how ligand rec-ognition and/or clustering of PGRPs triggers activation of downstream signalling, for example, how PGRP-LC recruits the death-domain-containing IMD protein. However, all PGRPs contain an N-terminal segment of at least 22–23 residues (often longer) that is characterized by high sequence variability. In PGRP-LC, this segment forms the cytoplasmic signalling domain. In soluble PGRPs, this segment could work together with the α2 helix that is located next to the peptidoglycan-binding site, which is absent from bacteriophage amidases. These highly variable domains could function as binding sites for host effectors or signalling proteins such as GNBP1.
Perspectives and future directionsBoth insect and mammalian PGRPs function in anti-microbial defences and have similar peptidoglycan-recognizing PGRP domains, but they accomplish their functions through different mechanisms. Insect PGRPs activate signal transduction pathways in cells or proteo-lytic cascades in the haemolymph that generate antimi-crobial effectors. By contrast, mammalian PGRPs are directly bactericidal. Only PGRPs with amidase activity have similar anti-inflammatory scavenger functions in both insects and mammals.
Many questions, however, still need to be answered. How do insect PGRPs transduce the Toll-, IMD- and prophenoloxidase-activating signals? What is the in vivo significance of discrimination between monomeric and polymeric peptidoglycans? Do PGRPs have any other functions? The functions of many insect PGRPs are still unknown, and almost nothing is known about the functions of PGRPs in other invertebrates and non-mammalian vertebrates. Moreover, PGRPs might also have multiple functions. For example, D. melanogaster PGRP-SA triggers the pathway that activates Toll, but it also has carboxypeptidase activity. Similarly, the pig PGLYRP-2 is an amidase, but might also function in regulating β-defensin expression9. Moreover, the in vivo role of mammalian and other vertebrate PGRPs and their overall contribution to antimicrobial host defences must be established, taking into account the recently discov-ered synergistic activities of PGRPs and antimicrobial peptides. Possible associations of PGRPs with diseases also need to be explored. For example, the human pglyrp3 and pglyrp4 genes are located in the epidermal differen-tiation gene cluster in the psoriasis sensitivity (psors4) locus. Mutations in pglyrp3 and pglyrp4 genes might therefore contribute to the pathogenesis of psoriasis85 or other skin diseases.
PGRPs probably evolved from primordial amidases, but it is not known when these amidases acquired broad bactericidal activity and which amidases in inverte-brates or lower vertebrates are also bactericidal. The exact mechanism of bactericidal activity of PGRPs is also not known. Moreover, human PGRPs were shown to prevent lung infection in mice22, but can they be applied for prevention or treatment of infections in humans? Future research into these, and many other questions, should keep peptidoglycan researchers busy for years to come.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | APRIL 2007 | 275
© 2007 Nature Publishing Group

1. Yoshida, H., Kinoshita, K. & Ashida, M. Purification of a peptidoglycan recognition protein from hemolymph of the silkworm, Bombyx mori. J. Biol. Chem. 271, 13854–13860 (1996).The first characterization of a PGRP protein.
2. Kang, D., Liu, G., Lundstrom, A., Gelius, E. & Steiner, H. A peptidoglycan recognition protein in innate immunity conserved from insects to humans. Proc. Natl Acad. Sci. USA 95, 10078–10082 (1998).The first cloning of insect and mammalian PGRPs, showing that PGRPs are conserved throughout evolution.
3. Ochiai, M. & Ashida, M. A pattern recognition protein for peptidoglycan. Cloning the cDNA and the gene of the silkworm, Bombyx mori. J. Biol. Chem. 274, 11854–11858 (1999).
4. Kiselev, S. L. et al. Molecular cloning and characterization of the mouse tag7 gene encoding a novel cytokine. J. Biol. Chem. 273, 18633–18639 (1998).
5. Werner, T. et al. A family of peptidoglycan recognition proteins in the fruit fly Drosophila melanogaster. Proc. Natl Acad. Sci. USA 97, 13772–13777 (2000).
6. Liu, C., Xu, Z., Gupta, D. & Dziarski, R. Peptidoglycan recognition proteins: a novel family of four human innate immunity pattern recognition molecules. J. Biol. Chem. 276, 34686–34694 (2001).Describes the discovery of a family of PGRPs in mammals.
7. Christophides, G. K. et al. Immunity-related genes and gene families in Anopheles gambiae. Science 298, 159–165 (2002).
8. Dziarski, R. & Gupta, D. The peptidoglycan recognition proteins (PGRPs). Genome Biol. 7, 232 (2006).
9. Sang, Y., Ramanathan, B., Ross, C. R. & Blecha, F. Gene silencing and overexpression of porcine peptidoglycan recognition protein long isoforms: involvement in β-defensin-1 expression. Infect. Immun. 73, 7133–7141 (2005).
10. Tydell, C. C., Yount, N., Tran, D., Yuan, J. & Selsted, M. E. Isolation, characterization, and antimicrobial properties of bovine oligosaccharide-binding protein. A microbicidal granule protein of eosinophils and neutrophils. J. Biol. Chem. 277, 19658–19664 (2002).
11. Goodson, M. S. et al. Identifying components of the NF-kappaB pathway in the beneficial Euprymna scolopes–Vibrio fischeri light organ symbiosis. Appl. Environ. Microbiol. 71, 6934–6946 (2005).
12. Guan, R., Wang, Q., Sundberg, E. J. & Mariuzza, R. A. Crystal structure of human peptidoglycan recognition protein S (PGRP-S) at 1.70 Å resolution. J. Mol. Biol. 347, 683–691 (2005).
13. Chang, C. I. et al. A Drosophila pattern recognition receptor contains a peptidoglycan docking groove and unusual L,D-carboxypeptidase activity. PLoS Biol. 2, e277 (2004).
14. Guan, R., Malchiodi, E. L., Wang, Q., Schuck, P. & Mariuzza, R. A. Crystal structure of the C-terminal peptidoglycan-binding domain of human peptidoglycan recognition protein Iα. J. Biol. Chem. 279, 31873–31882 (2004).
15. Reiser, J. B., Teyton, L. & Wilson, I. A. Crystal structure of the Drosophila peptidoglycan recognition protein (PGRP)-SA at 1.56 Å resolution. J. Mol. Biol. 340, 909–917 (2004).
16. Kim, M. S., Byun, M. & Oh, B. H. Crystal structure of peptidoglycan recognition protein LB from Drosophila melanogaster. Nature Immunol. 4, 787–793 (2003).The first crystal structure of a PGRP molecule.
17. Guan, R. et al. Crystal structure of human peptidoglycan recognition protein Iα bound to a muramyl pentapeptide from Gram-positive bacteria. Protein Sci. 15, 1199–1206 (2006).
18. Guan, R. et al. Structural basis for peptidoglycan binding by peptidoglycan recognition proteins. Proc. Natl Acad. Sci. USA 101, 17168–17173 (2004).
19. Wang, Z. M. et al. Human peptidoglycan recognition protein-L is an N-acetylmuramoyl-L-alanine amidase. J. Biol. Chem. 278, 49044–49052 (2003).
20. Michel, T., Reichhart, J. M., Hoffmann, J. A. & Royet, J. Drosophila Toll is activated by Gram-positive bacteria through a circulating peptidoglycan recognition protein. Nature 414, 756–759 (2001).The first in vivo demonstration of the role of a PRR in the Drosophila immune response. Details the discovery of a PGRP that triggers the activation of a signal transduction pathway and the discovery of the role of PGRPs in resistance to infections.
21. Zhang, Y. et al. Identification of serum N-acetylmuramoyl-L-alanine amidase as liver peptidoglycan recognition protein 2. Biochim. Biophys. Acta 1752, 34–46 (2005).
22. Lu, X. et al. Peptidoglycan recognition proteins are a new class of human bactericidal proteins. J. Biol. Chem. 281, 5895–5907 (2006).Describes the discovery of the function of mammalian PGLYRP-3 and PGLYRP-4 as a new class of bactericidal proteins.
23. De Pauw, P., Neyt, C., Vanderwinkel, E., Wattiez, R. & Falmagne, P. Characterization of human serum N-acetylmuramyl-L-alanine amidase purified by affinity chromatography. Protein Expr. Purif. 6, 371–378 (1995).
24. Mellroth, P. et al. Ligand-induced dimerization of Drosophila peptidoglycan recognition proteins in vitro. Proc. Natl Acad. Sci. USA 102, 6455–6460 (2005).Explains how different PGRP-LC isoforms detect monomeric versus polymeric peptidoglycan. Demonstrates that PGRP-LCa is unable to bind to polymeric peptidoglycan.
25. Choe, K. M., Lee, H. & Anderson, K. V. Drosophila peptidoglycan recognition protein LC (PGRP-LC) acts as a signal-transducing innate immune receptor. Proc. Natl Acad. Sci. USA 102, 1122–1126 (2005).
26. Hoffmann, J. A. The immune response of Drosophila. Nature 426, 33–38 (2003).
27. Meister, M. Blood cells of Drosophila: cell lineages and role in host defence. Curr. Opin. Immunol. 16, 10–15 (2004).
28. Royet, J., Reichhart, J. M. & Hoffmann, J. A. Sensing and signaling during infection in Drosophila. Curr. Opin. Immunol. 17, 11–17 (2005).
29. Bulet, P. & Stocklin, R. Insect antimicrobial peptides: structures, properties and gene regulation. Protein Pept. Lett. 12, 3–11 (2005).
30. Lemaitre, B., Reichhart, J. M. & Hoffmann, J. A. Drosophila host defense: differential induction of antimicrobial peptide genes after infection by various classes of microorganisms. Proc. Natl Acad. Sci. USA 94, 14614–14619 (1997).
31. Kurata, S., Ariki, S. & Kawabata, S. Recognition of pathogens and activation of immune responses in Drosophila and horseshoe crab innate immunity. Immunobiol. 211, 237–249 (2006).
32. Royet, J. Infectious non-self recognition in invertebrates: lessons from Drosophila and other insect models. Mol. Immunol. 41, 1063–1075 (2004).
33. Filipe, S. R., Tomasz, A. & Ligoxygakis, P. Requirements of peptidoglycan structure that allow detection by the Drosophila Toll pathway. EMBO Rep. 6, 327–333 (2005).
34. Gobert, V. et al. Dual activation of the Drosophila Toll pathway by two pattern recognition receptors. Science 302, 2126–2130 (2003).
35. Bischoff, V. et al. Function of the Drosophila pattern-recognition receptor PGRP-SD in the detection of Gram-positive bacteria. Nature Immunol. 5, 1175–1180 (2004).
36. Choe, K. M., Werner, T., Stoven, S., Hultmark, D. & Anderson, K. V. Requirement for a peptidoglycan recognition protein (PGRP) in Relish activation and antibacterial immune responses in Drosophila. Science 296, 359–362 (2002).
37. Gottar, M. et al. The Drosophila immune response against Gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature 416, 640–644 (2002).Shows that a PGRP family member functions as a transmembrane receptor upstream of the IMD pathway (also see Refs 36 and 38).
38. Ramet, M., Manfruelli, P., Pearson, A., Mathey-Prevot, B. & Ezekowitz, R. A. Functional genomic analysis of phagocytosis and identification of a Drosophila receptor for E. coli. Nature 416, 644–648 (2002).
39. Leulier, F. et al. The Drosophila immune system detects bacteria through specific peptidoglycan recognition. Nature Immunol. 4, 478–484 (2003).Explains why Lys-type peptidoglycan activates the Toll pathway, whereas DAP-type peptidoglycan mainly induces the IMD pathway.
40. Kaneko, T. et al. Monomeric and polymeric Gram-negative peptidoglycan but not purified LPS stimulate the Drosophila IMD pathway. Immunity 20, 637–649 (2004).Demonstrates that different PGRP-LC receptor combinations are required to detect monomeric versus polymeric peptidoglycan.
41. Stenbak, C. R. et al. Peptidoglycan molecular requirements allowing detection by the Drosophila immune deficiency pathway. J. Immunol. 173, 7339–7348 (2004).
42. Takehana, A. et al. Peptidoglycan recognition protein (PGRP)-LE and PGRP-LC act synergistically in Drosophila immunity. EMBO J. 23, 4690–4700 (2004).
43. Takehana, A. et al. Overexpression of a pattern-recognition receptor, peptidoglycan-recognition protein-LE, activates Imd/Relish-mediated antibacterial defense and the prophenoloxidase cascade in Drosophila larvae. Proc. Natl Acad. Sci. USA 99, 13705–13710 (2002).
44. Kaneko, T. et al. PGRP-LC and PGRP-LE have essential yet distinct functions in the Drosophila immune response to monomeric DAP-type peptidoglycan. Nature Immunol. 7, 715–723 (2006).Demonstrates that PGRP-LE functions as an intracellular receptor for bacteria.
45. Viala, J. et al. Nod1 responds to peptidoglycan delivered by the Helicobacter pylori cag pathogenicity island. Nature Immunol. 5, 1166–1174 (2004).
46. Park, J. W. et al. A synthetic peptidoglycan fragment as a competitive inhibitor of the melanization cascade. J. Biol. Chem. 281, 7747–7755 (2006).
47. Lee, M. H. et al. Peptidoglycan recognition proteins involved in 1,3-β-D-glucan-dependent prophenoloxidase activation system of insect. J. Biol. Chem. 279, 3218–3227 (2004).
48. Kocks, C. et al. Eater, a transmembrane protein mediating phagocytosis of bacterial pathogens in Drosophila. Cell 123, 335–346 (2005).
49. Garver, L. S., Wu, J. & Wu, L. P. The peptidoglycan recognition protein PGRP-SC1a is essential for Toll signaling and phagocytosis of Staphylococcus aureus in Drosophila. Proc. Natl Acad. Sci. USA 103, 660–665 (2006).
50. Zaidman-Remy, A. et al. The Drosophila amidase PGRP-LB modulates the immune response to bacterial infection. Immunity 24, 463–473 (2006).Demonstrates that a PGRP with catalytic activity is required to control the level of activation of the IMD pathway after infection.
51. Gelius, E., Persson, C., Karlsson, J. & Steiner, H. A mammalian peptidoglycan recognition protein with N-acetylmuramoyl-L-alanine amidase activity. Biochem. Biophys. Res. Commun. 306, 988–994 (2003).
52. Mellroth, P., Karlsson, J. & Steiner, H. A scavenger function for a Drosophila peptidoglycan recognition protein. J. Biol. Chem. 278, 7059–7064 (2003).First demonstration that a Drosophila PGRP molecule can cleave peptidoglycan and decrease its immunostimulatory activity.
53. Mellroth, P. & Steiner, H. PGRP-SB1: an N-acetylmuramoyl-L-alanine amidase with antibacterial activity. Biochem. Biophys. Res. Commun. 350, 994–999 (2006).
54. Bischoff, V. et al. Downregulation of the Drosophila immune response by peptidoglycan-recognition proteins SC1 and SC2. PLoS Pathog. 2, e14 (2006).Shows that PGRPs with amidase activity expressed in the gut function as peptidoglycan detoxifying enzymes in vivo.
55. Munford, R. S. Detoxifying endotoxin: time, place and person. J. Endotoxin Res. 11, 69–84 (2005).
56. Wang, H., Gupta, D., Li, X. & Dziarski, R. Peptidoglycan recognition protein 2 (N-acetylmuramoyl-L-Ala amidase) is induced in keratinocytes by bacteria through the p38 kinase pathway. Infect. Immun. 73, 7216–7225 (2005).
57. Lo, D. et al. Peptidoglycan recognition protein expression in mouse Peyer’s patch follicle associated epithelium suggests functional specialization. Cell. Immunol. 224, 8–16 (2003).
58. Li, X., Wang, S., Wang, H. & Gupta, D. Differential expression of peptidoglycan recognition protein 2 in the skin and liver requires different transcription factors. J. Biol. Chem. 281, 20738–20748 (2006).
59. Swaminathan, C. P. et al. Dual strategies for peptidoglycan discrimination by peptidoglycan recognition proteins (PGRPs). Proc. Natl Acad. Sci. USA 103, 684–689 (2006).
60. Kumar, S. et al. Selective recognition of synthetic lysine and meso-diaminopimelic acid-type peptidoglycan fragments by human peptidoglycan recognition proteins Iα and S. J. Biol. Chem. 280, 37005–37012 (2005).
61. Tydell, C. C., Yuan, J., Tran, P. & Selsted, M. E. Bovine peptidoglycan recognition protein-S: antimicrobial activity, localization, secretion, and binding properties. J. Immunol. 176, 1154–1162 (2006).
R E V I E W S
276 | APRIL 2007 | VOLUME 5 www.nature.com/reviews/micro
© 2007 Nature Publishing Group

62. Wang, M. et al. Human peptidoglycan recogntion proteins require zinc to kill both Gram-positive and Gram-negative bacteria and are synergistic with antibacterial peptides. J. Immunol. 178, 3116–3125 (2007).
63. Dziarski, R., Platt, K. A., Gelius, E., Steiner, H. & Gupta, D. Defect in neutrophil killing and increased susceptibility to infection with nonpathogenic Gram-positive bacteria in peptidoglycan recognition protein-S (PGRP-S)-deficient mice. Blood 102, 689–697 (2003).
64. Mathur, P. et al. Murine peptidoglycan recognition proteins PglyrpIα and PglyrpIβ are encoded in the epidermal differentiation complex and are expressed in epidermal and hematopoietic tissues. Genomics 83, 1151–1163 (2004).
65. Uehara, A. et al. Chemically synthesized pathogen-associated molecular patterns increase the expression of peptidoglycan recognition proteins via Toll-like receptors, NOD1 and NOD2 in human oral epithelial cells. Cell. Microbiol. 7, 675–686 (2005).
66. Fraker, P. J. & King, L. E. Reprogramming the immune system during zinc deficiency. Annu. Rev. Nutr. 24, 277–298 (2004).
67. Walker, C. F. & Black, E. E. Zinc and the risk of infectious disease. Am. J. Clin. Nutr. 68, 447S–463S (2004).
68. Ibs, K. H. & Rink, L. Zinc-altered immune function. J. Nutr. 133, 1452S–1456S (2003).
69. Shankar, A. H. & Prasad, A. S. Zinc and immune function: the biological basis of altered resistance to infection. Am. J. Clin. Nutr. 68, 447S–463S (1998).
70. Dziarski, R. in Shaechter’s Mechanisms of Microbial Disease 4th Edn (eds R. Engelberg, and V. DiRita) 66–89 (Lippincott, Williams & Wilkins 2006).
71. Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 415, 389–395 (2002).
72. Ganz, T. Defensins: antimicrobial peptides of innate immunity. Nature Rev. Immunol. 3, 710–720 (2003).
73. Qu, X. D. & Lehrer, R. I. Secretory phospholipase A2 is the principal bactericide for staphylococci and other Gram-positive bacteria in human tears. Infect. Immun. 66, 2791–2797 (1998).
74. Koduri, R. S. et al. Bactericidal properties of human and murine groups I, II, V, X, and XII secreted phospholipases A2. J. Biol. Chem. 277, 5849–5857 (2002).
75. Weinrauch, Y., Elsbach, P., Madsen, L. M., Foreman, A. & Weiss, J. The potent anti-Staphylococcus aureus activity of a sterile rabbit inflammatory fluid is due to a 14-kD phospholipase A2. J. Clin. Invest. 97, 250–257 (1996).
76. Harwig, S. S. et al. Bactericidal properties of murine intestinal phospholipase A2. J. Clin. Invest. 95, 603–610 (1995).
77. Cho, J. H. et al. Human peptidoglycan recognition protein S is an effector of neutrophil-mediated innate immunity. Blood 106, 2551–2558 (2005).
78. Chang, C. I. et al. Structure of the ectodomain of Drosophila peptidoglycan-recognition protein LCa suggests a molecular mechanism for pattern recognition. Proc. Natl Acad. Sci. USA 102, 10279–10284 (2005).
79. Lim, J. H. et al. Structural basis for preferential recognition of diaminopimelic acid-type peptidoglycan by a subset of peptidoglycan recognition proteins. J. Biol. Chem. 281, 8286–8295 (2006).
80. Chang, C. I., Chelliah, Y., Borek, D., Mengin-Lecreulx, D. & Deisenhofer, J. Structure of tracheal cytotoxin in complex with a heterodimeric pattern-recognition receptor. Science 311, 1761–1764 (2006).
81. Liu, C., Gelius, E., Liu, G., Steiner, H. & Dziarski, R. Mammalian peptidoglycan recognition protein binds peptidoglycan with high affinity, is expressed in neutrophils, and inhibits bacterial growth. J. Biol. Chem. 275, 24490–24499 (2000).
82. Garcia, P., Gonzalez, M. P., Garcia, E., Lopez, R. & Garcia, J. L. LytB, a novel pneumococcal murein hydrolase essential for cell separation. Mol. Microbiol. 31, 1275–1277 (1999).
83. Loeffler, J. M., Nelson, D. & Fischetti, V. A. Rapid killing of Streptococcus pneumoniae with a bacteriophage cell wall hydrolase. Science 294, 2170–2172 (2001).
84. Garcia, P., Martin, A. C. & Lopez, R. Bacteriophages of Streptococcus pneumoniae: a molecular approach. Microb. Drug Resist. 3, 165–176 (1997).
85. Sun, C. et al. Peptidoglycan recognition proteins Pglyrp3 and Pglyrp4 are encoded from the epidermal differentiation complex and are candidate genes for the Psors4 locus on chromosome 1q21. Hum. Genet. 119, 113–125 (2006).
86. Bera, A., Herbert, S., Jakob, A., Vollmer, W. & Gotz, F. Why are pathogenic staphylococci so lysozyme resistant? The peptidoglycan O-acetyltransferase OatA is the major determinant for lysozyme resistance of Staphylococcus aureus. Mol. Microbiol. 55, 778–787 (2005).
87. Cloud-Hansen, K. A. et al. Breaching the great wall: peptidoglycan and microbial interactions. Nature Rev. Microbiol. 4, 710–716 (2006).
88. Vollmer, W. & Holtje, J. V. The architecture of the murein (peptidoglycan) in gram-negative bacteria: vertical scaffold or horizontal layer(s)? J. Bacteriol. 186, 5978–5987 (2004).
89. Dmitriev, B. A., Toukach, F. V., Holst, O., Rietschel, E. T. & Ehlers, S. Tertiary structure of Staphylococcus aureus cell wall murein. J. Bacteriol. 186, 7141–7148 (2004).
90. Touhami, A., Jericho, M. H. & Beveridge, T. J. Atomic force microscopy of cell growth and division in Staphylococcus aureus. J. Bacteriol. 186, 3286–3295 (2004).
91. Meroueh, S. O. et al. Three-dimensional structure of the bacterial cell wall peptidoglycan. Proc. Natl Acad. Sci. USA 103, 4404–4409 (2006).
92. Dziarski, R., Ulmer, A. J. & Gupta, D. in Glycomicrobiology (ed. R. J. Doyle) 145–186 (Kluwer Academic/Plenum Publishers, 2000).
93. Doyle, R. J. & Dziarski, R. in Molecular Medical Microbiology (ed. M. Sussman) 137–154 (Academic Press, 2001).
94. De Kimpe, S. J., Kengatharan, M., Thiemermann, C. & Vane, J. R. The cell wall components peptidoglycan and lipoteichoic acid from Staphylococcus aureus act in synergy to cause shock and multiple organ failure. Proc. Natl Acad. Sci. USA 92, 10359–10363 (1995).
95. Koropatnick, T. A. et al. Microbial factor-mediated development in a host-bacterial mutualism. Science 306, 1186–1188 (2004).
96. Dziarski, R., Tapping, R. I. & Tobias, P. S. Binding of bacterial peptidoglycan to CD14. J. Biol. Chem. 273, 8680–8690 (1998).
97. Gupta, D., Kirkland, T. N., Viriyakosol, S. & Dziarski, R. CD14 is a cell-activating receptor for bacterial peptidoglycan. J. Biol. Chem. 271, 23310–23316 (1996).
98. Yoshimura, A. et al. Cutting edge: recognition of Gram-positive bacterial cell wall components by the innate immune system occurs via Toll-like receptor 2. J. Immunol. 163, 1–5 (1999).
99. Takeuchi, O. et al. Differential roles of TLR2 and TLR4 in recognition of Gram-negative and Gram-positive bacterial cell wall components. Immunity 11, 443–451 (1999).
100. Schwandner, R., Dziarski, R., Wesche, H., Rothe, M. & Kirschning, C. J. Peptidoglycan- and lipoteichoic acid-induced cell activation is mediated by Toll-like receptor 2. J. Biol. Chem. 274, 17406–17409 (1999).
101. Mitsuzawa, H. et al. Extracellular Toll-like receptor 2 region containing Ser40-Ile64 but not Cys30–Ser39 is critical for the recognition of Staphylococcus aureus peptidoglycan. J. Biol. Chem. 276, 41350–41356 (2001).
102. Iwaki, D. et al. The extracellular Toll-like receptor 2 domain directly binds peptidoglycan derived from Staphylococcus aureus. J. Biol. Chem. 277, 24315–24320 (2002).
103. Travassos, L. H. et al. Toll-like receptor 2-dependent bacterial sensing does not occur via peptidoglycan recognition. EMBO Rep. 5, 1000–1006 (2004).
104. Dziarski, R. & Gupta, D. Staphylococcus aureus peptidoglycan is a Toll-like receptor 2 activator: a reevaluation. Infect. Immun. 73, 5212–5216 (2005).
105. Bubeck Wardenburg, J., Williams, W. A. & Missiakas, D. Host defenses against Staphylococcus aureus infection require recognition of bacterial lipoproteins. Proc. Natl Acad. Sci. USA 103, 13831–13836 (2006).
106. Girardin, S. E. et al. Peptidoglycan molecular requirements allowing detection by Nod1 and Nod2. J. Biol. Chem. 278, 41702–41708 (2003).
107. Chamaillard, M. et al. An essential role for NOD1 in host recognition of bacterial peptidoglycan containing diaminopimelic acid. Nature Immunol. 4, 702–707 (2003).
108. Inohara, N. et al. Host recognition of bacterial muramyl dipeptide mediated through NOD2. Implications for Crohn’s disease. J. Biol. Chem. 278, 5509–5512 (2003).
109. Girardin, S. E. et al. Nod2 is a general sensor of peptidoglycan through muramyl dipeptide (MDP) detection. J. Biol. Chem. 278, 8869–8872 (2003).
110. Strober, W., Murray, P. J., Kitani, A. & Watanabe, T. Signalling pathways and molecular interactions of NOD1 and NOD2. Nature Rev. Immunol. 6, 9–20 (2006).
111. Miceli-Richard, C. et al. CARD15 mutations in Blau syndrome. Nature Genet. 29, 19–20 (2001).
112. Ogura, Y. et al. A frameshift mutation in NOD2 associated with susceptibility to Crohn’s disease. Nature 411, 603–606 (2001).
113. Hugot, J. P. et al. Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn’s disease. Nature 411, 599–603 (2001).
114. Dziarski, R., Viriyakosol, S., Kirkland, T. N. & Gupta, D. Soluble CD14 enhances membrane CD14-mediated responses to peptidoglycan: structural requirements differ from those for responses to lipopolysaccharide. Infect. Immun. 68, 5254–5260 (2000).
115. Nadesalingam, J., Dodds, A. W., Reid, K. B. & Palaniyar, N. Mannose-binding lectin recognizes peptidoglycan via the N-acetyl glucosamine moiety, and inhibits ligand-induced proinflammatory effect and promotes chemokine production by macrophages. J. Immunol. 175, 1785–1794 (2005).
116. Ma, Y. G. et al. Human mannose-binding lectin and L-ficolin function as specific pattern recognition proteins in the lectin activation pathway of complement. J. Biol. Chem. 279, 25307–25312 (2004).
117. Cash, H. L., Whitham, C. V., Behrendt, C. L. & Hooper, L. V. Symbiotic bacteria direct expression of an intestinal bactericidal lectin. Science 313, 1126–1130 (2006).
118. Ibrahim, H. R., Matsuzaki, T. & Aoki, T. Genetic evidence that antibacterial activity of lysozyme is independent of its catalytic function. FEBS Lett. 506, 27–32 (2001).
AcknowledgementsWe thank members of the Royet laboratory for comments on the manuscript. Research in the authors’ laboratories was supported by the National Institutes of Health (R.D.) and the Ministère de l’Education Nationale de la Recherche et de la Technologie, the Agence National pour la Recherche and the Fondation pour la Recherche Médicale (J.R.).
Competing interests statementThe authors declare no competing financial interests.
DATABASESThe following terms in this article are linked online to:Entrez Genome Project: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=genomeprjAnopheles gambiae | Caenorhabditis elegans | Drosophila melanogaster | Escherichia coli | Helicobacter pylori | Staphylococcus aureusUniProtKB: http://ca.expasy.org/sprotGNBP1 | Nod1 | Nod2 | PGLYRP-1 | PGLYRP-2 | PGLYRP-3 | PGLYRP-4 | PGRP-LB | PGRP-LC | PGRP-LE | PGRP-LF | PGRP-SA | PGRP-SB1 | PGRP-SC1 | PGRP-SD
FURTHER INFORMATIONJulien Royet’s homepage: http://www.ibdml.univ-mrs.fr/equipesRoman Dziarski’s homepage: http://iusm-nw.medicine.iu.edu/smpledsk.htmlAccess to this links box is available online.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | APRIL 2007 | 277
© 2007 Nature Publishing Group




















