
RISK-SENSITIVE ANTIPREDATOR BEHAVIOR IN THE TRINIDADIAN GUPPY, POECILIA RETICULATA

Available online at www.sciencedirect.com
ANIMAL BEHAVIOUR, 2008, 75, 113e121doi:10.1016/j.anbehav.2007.04.019
Pleiotropic antipredator strategies, fleeing and feigning death,
correlated with dopamine levels in Tribolium castaneum
TAKAHISA MIYATAKE* , KEN TABUCHI†, KEN SASAKI‡, KENSUKE OKADA* ,
KOHJI KATAYAMA* & SEIICHI MORIYA†
*Graduate School of Environmental Science, Okayama University
yNational Agricultural Research Center, Tsukuba
zDepartment of Brain and Bioinformation Science, Kanazawa Institute of Technology
(Received 20 October 2006; initial acceptance 20 November 2006;
final acceptance 16 April 2007; published online 27 September 2007; MS. number: 9154)
A prey animal has the alternative of fleeing or feigning death to survive when it encounters predators. Wefound that fleeing by an artificial threat, locomotion and feigning death are pleiotropically correlated witha genetic factor related to a biogenic amine in the red flour beetle, Tribolium castaneum. Walking distance ofadults was significantly lower in strains artificially selected for longer (L strains) than shorter duration(S strains) of death-feigning. Crosses showed that S-strain adults were dominant in the frequency and du-ration of death-feigning and locomotor activity compared to those of L strains, suggesting that death-feigning and activity have the same genetic basis. S-strain adults fled, but L-strain adults feigned death,when they encounter artificial threat. Not only adults that were directly selected for the duration ofdeath-feigning, but also the larvae of L strains frequently showed tonic immobility, when they were drop-ped onto the ground: the larvae of S strains showed this behaviour less often. This suggests that chemicalmodulators of behaviour present in these insects before and after metamorphosis control both general lo-comotor activity and death-feigning. Brain levels of the candidate neuromodulator dopamine were, in fact,found to be significantly higher in S strains compared to L strains in the two selection replications. Thus,we suggest that two alternative behaviours related to antipredator strategies, fleeing or feigning death, areassociated with the pleiotropic effects of a neuroactive substance in T. castaneum.
� 2007 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
Keywords: animal hypnosis; artificial selection; biogenic amine; death-feigning; defence; genetic correlation; locomotion;thanatosis; tonic immobility; Tribolium castaneum
Predation is a key selection pressure in shaping preybehaviours (Lima & Dill 1990; Sih 1992; Lima 1998). Be-cause natural selection favours individuals that success-fully avoid predators, selection has favoured antipredatorbehaviour. When a prey perceives danger, it often usestwo alternative tactics to survive, immobilizing orrunning; it cannot adopt both simultaneously. Death-
Correspondence: T. Miyatake, Laboratory of Evolutionary Ecology,Graduate School of Environmental Science, Okayama University,Okayama 800-7530, Japan (email: [email protected]). K. Tabuchi is now at the Faculty of Forestry and EnvironmentalManagement, University of New Brunswick, Fredericton, New Bruns-wick E3B 6C2, Canada. K. Sasaki is at the Department of Brain andBioinformation Science, Kanazawa Institute of Technology, Hakusan,Ishikawa, 924-0838, Japan. S. Moriya is at National Agricultural Re-search Center, Tsukuba, Ibaraki 305-8666, Japan.
110003e3472/08/$30.00/0 � 2007 The Association for the S
feigning (sometimes called animal hypnosis, playingdead, or thanatosis), a kind of tonic immobility assumedby many animals in response to external stimuli (Ruxtonet al. 2004, Ruxton 2006), has been considered a defencemechanism against predators that is shared by mammals(Franq 1969), birds (Sargeant & Eberhardt 1975), amphib-ians (Gargaglioni et al. 2001), fishes (Howe 1991), reptiles(McCallum 1999), crustaceans (Fedotov et al. 2006), mites(Ebermann 1991) and insects (Fabre 1900; Edmunds 1974,Miyatake et al. 2004; Ruxton et al. 2004; Hozumi &Miyatake 2005; Honma et al. 2006). On the other hand,a prey can survive by fleeing from predators. Escaping byrunning from predators is a common tactic for many ani-mals (Edmunds 1974) and may be related to the level oflocomotor activity of the insect.
At a physiological level, negative phenotypic relation-ships between death-feigning and activity have been
3tudy of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.

ANIMAL BEHAVIOUR, 75, 1114
observed in some insects. In the parasitoid species Nasoniavitripennis, the females that were least active had the great-est tendency to show thanatosis (King & Leaich 2006). Inthe sweetpotato weevil Cylas formicarius, the frequencyand duration of death-feigning are influenced by the behav-iour before the weevil was startled: almost all resting indi-viduals feigned death, whereas the active ones seldom did(Miyatake 2001a). Adults of Cylas formicarius, a nocturnalspecies, seldom showed death-feigning behaviour duringthe night, when they are active for reproduction, whereasthey showed a higher frequency and longer duration ofdeath-feigning in the daytime (Miyatake 2001a). Thesestudies suggest the existence of two behavioural modes,active and resting, in which less and more death-feigning,respectively, occurs. If death-feigning and activity are ge-netically linked, then both traits may evolve together asa trade-off strategy. That is, individuals that are active atthe time of threat often adopt a fleeing strategy, while inac-tive individuals frequently adopt a death-feigning strategy.
Many insect behaviours are controlled by neuroactivesubstances or biogenic amines, including octopamine anddopamine, that are derived from the amino acid tyrosine(Evans 1980; Bicker & Menzel 1989; Stevenson et al. 2000;Libersat & Pfluger 2004). In crickets, dopamine and octop-amine are required for a successful aggressive encounter(Stevenson et al. 2000), octopamine and dopamine playroles in conveying positive and negative reinforcing sig-nals in olfactory learning (Unoki et al. 2005, 2006), and fly-ing and fighting abilities are also regulated by biogenicamines (Hofmann & Stevenson 2000; Stevenson et al.2005). In the cockroach, dopamine and octopamineenhances the escape response (Goldstein & Camhi 1991;Casagrand & Ritzmann 1992). In honeybees, dopamineenhances the activity level of the motor response (Bozic& Woodring 1998; Menzel et al. 1999), sucrose responsive-ness is increased by injection of octopamine, but decreasedby dopamine (Scheiner et al. 2002), and octopamine alsomodulate other behaviours, such as learning, memory for-mation and division of labour (Scheiner et al. 2006). InDrosophila melanogaster, dopamine provides a role in themodulation of arousal (Kume et al. 2005) and in stress re-sponse (Chentsova et al. 2002). However, in spite ofmuch neuro-physiological research, no study has exam-ined the genetic relationships between insect behaviourand neuroactive substances from the view point of antipre-dation strategies.
We here show a negative genetic relationship betweenlocomotor activity or fleeing and death-feigning, anda possible chemical that controls fleeing or death-feigningbehaviour, using strains selected for shorter (S strains) orlonger (L strains) duration of death-feigning in Triboliumcastaneum, a model insect for the adaptive significanceof death-feigning behaviour (Prohammer & Wade 1981;Miyatake et al. 2004). The death-feigning behaviour ofthis beetle increases its fitness for survival against a preda-tory spider (Miyatake et al. 2004). Heritable variation ofdeath-feigning intensity has also been reported in thisbeetle (Prohammer & Wade 1981; Miyatake et al. 2004).
In the present study, first we measured adult locomotoractivities, fleeing or feigning death behaviours of adultsand larval tonic immobility from the S and L strains, and
examined crosses between the two strains to identify thegenetic base for the locomotor activity and larval tonicimmobility. We used the term ‘tonic immobility’, notdeath-feigning for larval thanatosis, because the adaptivesignificance for the larval immobility was not known inT. castaneum while for the adults the significance hasbeen identified as avoiding attack by a model predator,the jumping spider (Miyatake et al. 2004). Second, wecompared the levels of brain dopamine expression of adultbeetles in selected strains, because dopamine is a biogenicamine associated with increased locomotor activity, es-cape behaviour and arousal in insects (Goldstein & Camhi1991; Casagrand & Ritzmann 1992; Stevenson et al. 2000;Libersat & Pfluger 2004; Kume et al. 2005).
METHODS
Insects and Culture
The Tribolium castaneum beetle culture used in thisstudy has been maintained in laboratories for more than25 years. The beetles were fed wholemeal (YoshikuraShokai, Tokyo, Japan) enriched with brewer’s yeast (AsahiBeer, Toyko, Japan) as the rearing medium and kept ina chamber (Sanyo, Tokyo, Japan) maintained at 25�Cand 60% RH under a photoperiod of 16:8 h light:darkcycle (lights on at 0700, light off at 2300).
Observation of Death-feigning
One day before observation, each beetle was placed ina well of a 48-well tissue culture plate (Falcon, BectonDickinson and Company, Lincoln Park, NJ, U.S.A.) toprevent disturbance by other beetles, which usuallyreduces the duration of death-feigning (Miyatake 2001a).The next day, each beetle was gently placed on its backin a white china saucer (140 mm diameter, 15 mmdeep). Death-feigning behaviour was induced by touchingthe abdomen of the beetle with a wooden stick. A trialconsisted of provoking the death-feigning behaviour andrecording its duration with a stopwatch. The behaviourduration was defined as the length of time between thestick touching the beetle and detection of its first visiblemovement. If the beetle did not respond, the touch wasrepeated. When the beetle feigned death, the durationwas recorded. All the trials were conducted between1300 and 1700 hours in the chamber described above.
Artificial Selection
One hundred males and 100 females (15 days old) wererandomly selected from the stock culture and their death-feigning behaviour was observed (F0 generation). Themales and females (10 each) with the shortest durationof death-feigning were selected to propagate short-dura-tion strains (S strains); similarly, 10 males and 10 femaleswith the longest duration were selected to propagate long-duration strains (L strains). The males and females of eachstrain were placed in a plastic cup (6.5 cm diameter,2.5 cm height) with 20 g of medium and allowed to

MIYATAKE ET AL.: PLEIOTROPIC ANTIPREDATOR STRATEGIES 115
copulate and lay eggs for 1 week. The pupae from the eggswere stored in separate-sex groups in other plastic cupsand allowed to emerge. When the adults reached 10e15days of age, 100 males and 100 females were randomlyselected from each strain and their death-feigning wasobserved again (F1 generation). The same procedure wascarried out in each generation. Two selection replicatesfor the short and long strains (S-1 and L-1; and S-2 andL-2) initiated at the same time were tested and maintainedin the chamber. The selection regimes were continued formore than 20 generations for all strains. During the 11thand 13th generations for S-1 and L-1 strains, and the 11thand 12th generations for S-2 and L-2 strains, the traitcould not be selected and thus the selection was relaxed.
Adult Locomotor Activity
At generation 12 of both S and L strains, locomotoractivity was measured for male adults. The method to trackadult locomotion was followed by Tabuchi et al. (2007) withsome modifications. A T. castaneum adult was gently placedon a piece of paper and covered by a transparent square plas-tic container (36 � 36 mm and 3 mm) to allow the insect towalk freely. Although the adults can walk on the sides andtop of the plastic container, vertical movement could be ig-nored because the height of the container was low enoughto allow the adult to walk in only a horizontal direction. Six-teen containers each containing one adult were arrayed ina 4 � 4 matrix at intervals of less than 7 mm. The imagewas recorded for 10 min at one frame/s by a CCD video cam-era (VCC-250A, CIS Corporation, Toyko, Japan) under desklight conditions. The images obtained were analysed by im-age analysing software (Image Hyper II, Digimo Corpora-tion, Osaka, Japan) to calculate the distance each beetletravelled. Each adult was distinguished by setting a searcharea for a respective adult with the coordinate system ofthe software. For each search area, the linear distance walkedin 1 s was calculated from the coordinates at a given timeand those recorded 1 s before. Adults were tracked underLD 16:8 h photoperiod and at 25 � 2�C. Adults were indi-vidually set into each container 5 min before the start oftracking. Food and water were not supplied after they wereplaced in the apparatus.
Crosses
At generation 11, reciprocal crosses were made betweenthe S-1 and L-1 strains. Ten virgin males and 10 virginfemales randomly chosen from each of the S-1 and L-1strains were allowed to copulate and lay eggs in 10 daysfor four typed crosses (S-1 females � S-1 males, S-1females � L-1 males, L-1 females � S-1 males and L-1females � L-1 males). The pupae from the eggs were storedin separate-sex groups in other plastic cups and allowed toemerge. When the adults reached 10e15 days of age,66e70 males were prepared for each cross and their dura-tions of death-feigning were observed. At generation 14for the S strains and 13 for the L strains, reciprocal crosseswere again made between the S-1 and the L-1 strains bythe same method described above. Walking distances of
the 58e64 male offspring from each cross were measuredby the method described above.
Response of Adults to Artificial Threat
At generations more than 20 of both S and L strains, anartificial threat was presented to some males of the beetles.Males were placed in the centre of a filter paper (a radius of55 mm; Toyo Roshi Kaisha Ltd, Tokyo), and we picked atthe abdominal part of the beetles using forceps. Theresponses of adults, running away from the centre of thepaper, i.e. fleeing, or death-feigning at the centre of the pa-per, during 1 min were recorded. All trials were conductedin the chamber described above.
Tonic Immobility of Larvae
At generation 16 of both S and L strains, tonicimmobility of larvae was observed. Larval immobilitywas induced by picking up a larva with tweezers andthen dropping it onto the small white china saucerdescribed above from a height of 15 cm. A trial consistedof provoking the behaviour and timing the duration ofimmobility, i.e. the length of time between the droppingof the larva and its first visible movement, with a stop-watch. If the larva failed to respond to the stimulus, thesame stimulus (picking up and dropping) was providedonce more or even a third time. If the larva showed tonicimmobility, the duration was recorded. All trials wereconducted in the chamber described above.
Expression of Brain Dopamine
Bodies of adult T. castaneum males (at generation 20)were frozen whole with liquid nitrogen, lyophilized over-night, and stored with silica gel at �80�C. The contents ofeach freeze-dried head including brain, suboesophagealganglion, frontal ganglion and mandibular muscle wereremoved out by peeling the head cuticle under a dissectingmicroscope, homogenized with a micro-glass homoge-nizer in 50 ml of ice-cold 0.1 M perchloric acid containing12.5 ng/ml 3,4-dihydroxybenzylamine (DHBA) as theinternal standard, and centrifuged in an Eppendorf tube.To specifically extract catecholamines including dopa-mine, the samples were subjected to alumina adsorptionprocedures (Anton & Sayre 1962; Refshauge et al. 1974;Sasaki & Nagao 2001). Each sample was transferred toa tube containing 20 mg alumina as an adsorbent and250 ml of 1.5 M Trisebuffer (pH 8.8) containing 10 mM2NaeEDTA, and stirred by a micro-stirring bar for 15 minat room temperature. After centrifugation at 3 000 rpm for2 min, the alumina adsorbent containing the catechol-amines was washed twice with 1 ml of 5 mM Trisebuffer(pH 8.8) and stirred with 150 ml of 0.1 M perchloric acidfor 20 min. The samples were centrifuged at 15 000 rpmfor 30 min at 0�C. The supernatant was transferred to amicro-vial for subsequent HPLC-ECD analysis.
An improved version (Harano et al. 2005) of the methodoriginally described by Sasaki & Nagao (2001, 2002) was

ANIMAL BEHAVIOUR, 75, 1116
used for the determination of dopamine. The HPLC-ECDsystem consisted of a solvent delivery pump (EP-300, EicomCorporation, Kyoto, Japan), refrigerated automatic injector(231XL-402, Gilson, Middleton, WI, U.S.A.), and a C18reversed-phase column (250 � 4.6 nm i.d., 5-mm averageparticle size, UG 120, Shiseido,Tokyo, Japan) maintainedat 35�C in a column oven. An electrochemical detector(ECD-300, Eicom Corporation, Kyoto, Japan) with a glassycarbon electrode (WE-GC, Eicom Corporation, Kyoto,Japan) was used. The detector potential was usually set at0.8 V referenced to an Ag/AgCl electrode. The detector cellwas held at a constant temperature of 35�C by placing it inthe column oven. Signals from the electrochemical detectorwere recorded and integrated by using data analysis software(PowerChrom, AD Instruments, Nagoya, Japan). Themobilephase contained 0.18 M monochloroacetic acid and 40 mMof 2NaeEDTA adjusted to pH 3.6 with NaOH. Both2.77 mM sodium-1-octanesulfonate as the ion-pair reagent,and 5% CH3CN as the organic modifier were added to thissolution. The mobile phase buffer was filtered througha 0.22-mm filter (Millipore, Billerica, U.S.A.) and degassed.The flow rate was kept constant at 0.7 ml/min. Externalstandards were run before and after sample runs to identifyand quantify dopamine. A peak of dopamine was identifiedby comparing both the retention time and hydrodynamicvoltamograms with the standards. Measurements were ob-tained by comparing peak heights of sample and standardchromatograms.
Statistical Analysis
Data for walking distances, the duration of tonicimmobility in larvae, and the expression of brain
dopamine were analysed using the ManneWhitney Utests to compare replicates within selection regimes atfirst, and if they were significantly different, then analysedS-1 versus L-1 and S-2 versus L-2, separately, with anotherManneWhitney U test. To compare the duration of death-feigning and the travel distances of adults among crossedpopulations, KruskaleWallis tests and corrected the signif-icance level (a ¼ 0.01) by the Holm’s sequential Bonfer-roni method (Rice 1989) after ManneWhitney U testsbetween combinations were used. To compare the per-centages of adult’s responses to artificial threat and oflarvae expressing tonic immobility between strains, logis-tic regression models were produced with two categoricalexplanatory variables: strain (S or L), replicates (1 or 2),and response (run or feign death for adults, and expressingor nonexpressing tonic immobility of larvae). In all thetreatments, observed frequencies were adjusted by adding0.5 to create an empirical logit (Haldane 1955, Gart et al.1985, Agresti 1990). All statistical analyses were conductedusing JMP version 6.03 (SAS Institute, 2005).
RESULTS
Responses to Selection
The duration of death-feigning showed a clear directresponse to selection and a steady divergence between thetwo selection regimes in both selection replications(Fig. 1). After 16 generations of selection, S strain beetlesfeigned death for 0.1 s, while L strain beetles showedmore than 2 min of death-feigning on average.
The frequency of death-feigning behaviour also showeda clear correlated response to selection for the duration of
Generations
Du
rati
on o
f d
eath
-fei
gnin
g (s
)
(c)
0.01
0.1
1
10
100
1000
10000(a)
0 4 8 12 16
(d)
0.01
0.1
1
10
100
1000
0 4 8 12 16
(b)
Figure 1. Direct response to selection in (a) males of replicate 1, (b) males of replicate 2, (c) females of replicate 1 and (d) females of replicate
2 in T. castaneum on short (S-1) (filled triangles), short (S-2) (opened diamonds), long (L-1) (filled circles) and long (L-2) (opened squares)duration of death-feigning. Means are shown with SD as vertical bars on each symbol. During generations 11 and 13 for S-1 and L-1 lines,
and generations 11 and 12 for S-2 and L-2 lines, the trait could not be selected and thus the selection was relaxed.

MIYATAKE ET AL.: PLEIOTROPIC ANTIPREDATOR STRATEGIES 117
death-feigning (Fig. 2). After 10 generations, almost all Lstrain beetles feigned death, whereas few S strain beetlesdid in both selection replicates (Fig. 2).
Adult Locomotor Activity
Walking distances of adult male beetles derived fromShort and Long strains are shown in Fig. 3. Adult travelled
S1(56)
L1(61)
S2(55)
L2(56)
Selection lines
Wal
kin
g d
ista
nce
s (m
)
0.6
0.8
1
1.2
1.4
Figure 3. Walking distances of T. castaneum male adults of short (,,S-1 and S-2), long (-, L-1 and L-2) lines at generation 12. The num-
ber in parentheses show sample size, and the means are shown � SE.
Freq
uen
cy o
f d
eath
fei
gnin
g
0
0.2
0.4
0.6
0.8
1
0
0.2
0.4
0.6
0.8
1
0 4 8 12 16
0 4 8 12 16
(a)
(b)
Generations
Figure 2. Relationship between the frequency of death-feigning in
(a) male and (b) female T. castaneum (correlated response to selec-
tion) and the generations for disrupted selection for duration ofdeath-feigning. Triangles and diamonds show S-1 and S-2 lines,
and circles and squares show L-1 and L-2 lines, respectively. During
generations 11 and 13 for S-1 and L-1 lines, and generations 11 and
12 for S-2 and L-2 lines, the correlated response was not measured.
distance was significantly shorter in beetles from L strainsthan S strains in males (Fig. 3). ManneWhitney U testsshowed highly significant differences in walking distancesbetween S-1 and L-1 (Z ¼ 5.32, N1 ¼ 56, N2 ¼ 61, P <0.0001) and between S-2 and L-2 (Z ¼ 2.55, N1 ¼ 55,N2 ¼ 56, P ¼ 0.01): significant differences were found be-tween replicates within the selection regimes (S-1 versusS-2: Z ¼ 4.19, N1 ¼ 56, N2 ¼ 55, P < 0.0001, L-1 versusL-2: Z ¼ 1.94, N1 ¼ 61, N2 ¼ 56, P ¼ 0.05).
Duration of Death-Feigning and LocomotorActivity in Crosses
Significant differences were found in the duration ofdeath-feigning (H3 ¼ 57.4, P < 0.0001) and in walking dis-tance (H3 ¼ 37.7, P < 0.0001) among crossed populations(Table 1). The adults derived from the reciprocal crosses,that is, S-1 females � L-1 males and L-1 females � S-1males, showed statistically the same duration of death-feigning and the same walking distances as the adultsderived from the cross of S-1 females � S-1 males. The adultsfrom the cross of L-1 females � L-1 males had significantlylonger duration of death-feigning and shorter walking dis-tance compared with the cross within S-strains and the re-ciprocal crosses between S and L strains (Table 1). Thissuggests the dominance in direction of short strains forthe duration of death-feigning and the locomotor activity.
Response of Adults to Artificial Threat
All adults derived from L strains (N ¼ 20 for L-1 and L-2strains) feigned death by artificial threat, while almost alladults from S strains fled by the threat and went out of thefilter paper within 1 min (19 of 20 adults in S-1 strain, and20 of 20 adults in S-2 strain). Logistic analysis showeda significant effect of selection regimen (c2
1 ¼ 27:37,P < 0.0001), but nonsignificant effect of replicates(c2
1 ¼ 0:19, P ¼ 0.66) and the interaction of selection regi-men and replicates (c2
1 ¼ 0:19, P ¼ 0.66).
Tonic Immobility of Larvae
The percentages of larvae expressing tonic immobility(TI) were significantly higher in L than in S strains (Fig. 4).
Table 1. Duration of death-feigning and walking distance of adultsderived from crosses between S-1 and L-1 lines
Cross female�male
Duration of
death-feigning (s)
Walking
distance (m)
N Mean�SD N Mean�SD
Short�short 69 1.17�6.39a 64 0.69�0.36a
Short�long 69 18.25�29.62a 61 0.74�0.24a
Long�short 70 42.14�71.71a 58 0.80�0.30a
Long�long 66 311.23�385.51b 62 0.52�0.21b
Same superscript letter indicates no significant difference in durationor distance by Holm’s sequential Bonferroni method (a ¼ 0.01; Rice1989).
ANIMAL BEHAVIOUR, 75, 1118
Logistic analysis showed a significant effect of selectionregimen (c2
1 ¼ 86:74, P < 0.0001), but nonsignificant ef-fect of replicates (c2
1 ¼ 0:13, P ¼ 0.72) and the interactionof selection regimen and replicates (c2
1 ¼ 0:13, P ¼ 0.72).Larvae from L strains had significantly longer duration
of tonic immobility compared to those of S strains (Fig. 4).ManneWhitney U tests showed highly significant differ-ences in the duration of tonic immobility between S-1and L-1 (Z ¼ 6.46, N1 ¼ N2 ¼ 50, P < 0.0001) and betweenS-2 and L-2 (Z ¼ 6.97, N1 ¼ N2 ¼ 50, P < 0.0001): signifi-cant differences were found between replicates onlywithin L-regimes (L-1 versus L-2: Z ¼ 2.37, N1 ¼ N2 ¼ 50,P ¼ 0.02, S-1 versus S-2: Z ¼ 0.13, N1 ¼ N2 ¼ 50, P ¼ 0.89).
Expression of Brain Dopamine
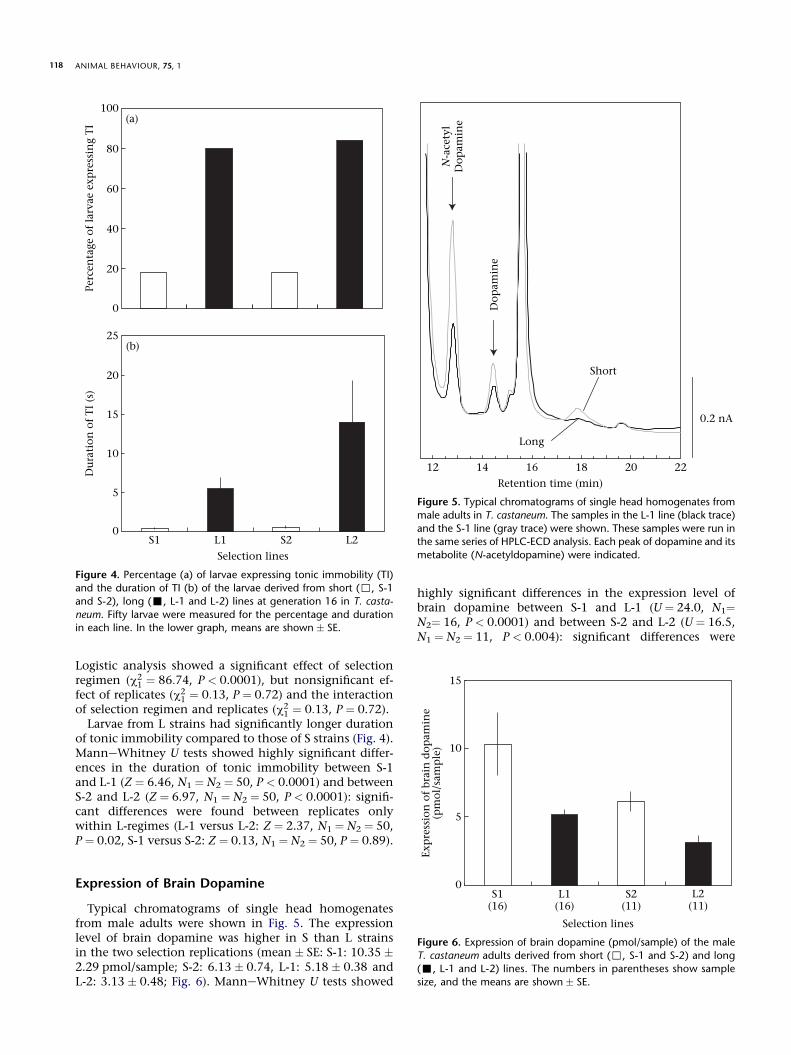
Typical chromatograms of single head homogenatesfrom male adults were shown in Fig. 5. The expressionlevel of brain dopamine was higher in S than L strainsin the two selection replications (mean � SE: S-1: 10.35 �2.29 pmol/sample; S-2: 6.13 � 0.74, L-1: 5.18 � 0.38 andL-2: 3.13 � 0.48; Fig. 6). ManneWhitney U tests showed
0
20
40
60
80
100(a)
Perc
enta
ge o
f la
rvae
exp
ress
ing
TI
0
5
10
15
20
25
S1 S2L1 L2
(b)
Selection lines
Du
rati
on o
f T
I (s
)
Figure 4. Percentage (a) of larvae expressing tonic immobility (TI)
and the duration of TI (b) of the larvae derived from short (,, S-1
and S-2), long (-, L-1 and L-2) lines at generation 16 in T. casta-
neum. Fifty larvae were measured for the percentage and durationin each line. In the lower graph, means are shown � SE.
highly significant differences in the expression level ofbrain dopamine between S-1 and L-1 (U ¼ 24.0, N1¼N2¼ 16, P < 0.0001) and between S-2 and L-2 (U ¼ 16.5,N1 ¼ N2 ¼ 11, P < 0.004): significant differences were
Retention time (min)
12 14 16 18 20 22
0.2 nA
Dop
amin
e
Short
LongN
-ace
tyl
Dop
amin
e
Figure 5. Typical chromatograms of single head homogenates from
male adults in T. castaneum. The samples in the L-1 line (black trace)
and the S-1 line (gray trace) were shown. These samples were run inthe same series of HPLC-ECD analysis. Each peak of dopamine and its
metabolite (N-acetyldopamine) were indicated.
0
5
10
15
S1(16)
S2(11)
L1(16)
L2(11)
Selection lines
Exp
ress
ion
of
brai
n d
opam
ine
(pm
ol/s
amp
le)
Figure 6. Expression of brain dopamine (pmol/sample) of the male
T. castaneum adults derived from short (,, S-1 and S-2) and long(-, L-1 and L-2) lines. The numbers in parentheses show sample
size, and the means are shown � SE.

MIYATAKE ET AL.: PLEIOTROPIC ANTIPREDATOR STRATEGIES 119
found between replicates within the selection regimes (S-1versus S-2: U ¼ 40.0, N1 ¼ 16, N2 ¼ 11, P ¼ 0.02, L-1 versusL-2: U ¼ 31.0, N1 ¼ 16, N2 ¼ 11, P ¼ 0.005).
DISCUSSION
A genetic correlation was found between death-feigningbehaviour and the level of running with locomotoractivity, which is associated with fleeing in T. castaneum.Beetles derived from strains having less and more death-feigning had longer and shorter travelling distances, re-spectively. Predator avoidance rates may depend ona prey’s travel speed when the prey escapes by fleeing.On the other hand, death-feigning behaviour is adaptiveto survival when T. castaneum adults encounter a modelpredator (Miyatake et al. 2004). Therefore, having alterna-tives, immobilizing or fleeing, may be important to sur-vival for a prey when the prey encounters predators.Alternative behavioural options to escape from predatorsare known for various prey behaviours in a wide rangeof taxonomic groups, for example, swimming away orhiding in a refuge for fishes (Lehtiniemi 2005), runningor hiding for lizards (Martin & Lopez 2000), fleeing orfighting for crabs (Wasson & Lyon 2005) as phenotypictactics, and flying away or feigning death for another bee-tle species as genetic strategies (Ohno & Miyatake 2007).
To date, several negative relationships between death-feigning and activity levels have been reported at a phys-iological level in a parasitoid wasp (King & Leaich 2006)and sweetpotato beetle (Miyatake 2001a). Among beetles,starved adults had a lower frequency of death-feigning(Acheampong & Mitchell 1997; Miyatake 2001b). Hightemperatures, at which insects are usually activated, sup-pressed death-feigning behaviour in a water bug (Holmes1906). These studies suggest the existence of two behav-ioural modes, active and resting. The present results sug-gest that the physiological relationship on the twomodes has a genetic base.
Crossing experiments showed that beetles with highactivity and a low frequency of death-feigning weredominant compared to beetles with low activity anda high frequency of death-feigning in T. castaneum. Thissuggests the dominance in direction of short strains forthe duration of death-feigning and the locomotor activity,implying that death-feigning intensity and locomotoractivity may have the same genetic background. This sup-ports the hypothesis that the genetic factor controlling ac-tivity and death-feigning is the same.
L-strain adults feigned death, but S-strain adults fled,when they encountered artificial threat. This suggestsa negative pleiotropic relationship between fleeing andfeigning death in T. castaneum correlated with a geneticfactor. The tonic immobility of larvae was also affectedby artificial selection for the duration of adult death-feign-ing as a correlated response. The frequency of larvae ex-pressing tonic immobility was higher and the durationof immobility was longer in L than S strains selected forthe duration of adult death-feigning, respectively. The cor-related response suggests that chemical componentsmaintained in the body through metamorphosis controltonic immobility and activity. An adaptive significance
of death-feigning behaviours of T. castaneum adults wassuggested in Miyatake et al. (2004), in which moredeath-feigning adults survived than beetles that did notfeign death when they encountered a model predator,a jumping spider. On the other hand, the effects of thesepredators on larval tonic immobility in T. castaneumhave not been studied, although a few anthocoridbugs have been reported as potential predators againstT. castaneum young larvae (Parajulee & Phillips 1993;Parajulee et al. 1994; Donnelly & Phillips 2001). Thus,ecological significance of larval tonic immobility isunknown.
Generally, neuromodulators, or biogenic amines includ-ing octopamine, serotonin, tyramine, and dopamine,regulate insect behaviours (e.g. Libersat & Pfluger 2004;Fahrbach & Mesce 2005; Roeder 2005). Octopamine, aninvertebrate counterpart of the vertebrate adrenergictransmitter (Hoyle 1975; Hoyle & Barker 1975), affects in-sect activity. In crickets, fighting behaviours are regulatedby octopamine (Stevenson et al. 2005), and dopamine andoctopamine are required for establishing a level of excit-ability sufficient for aggressive behaviour (Stevensonet al. 2000). High haemolymph octopamine titres are cor-related with aggressive behaviour and dominance in malecrickets (Adamo et al. 1995). Dopamine and octopamineenhances the escape response in a cockroach species(Goldstein & Camhi 1991; Casagrand & Ritzmann1992), and dopamine also affects the motor activity ofhoneybees (Bozic & Woodring 1998; Menzel et al. 1999).
In the present result, we thus compared the expressionof brain dopamine in adults of selected strains to test thehypothesis that the biogenic amines controlling insectactivity also affect antipredator behaviour, includingfeigning death and fleeing. As a result, clear evidence fora genetic relationship between dopamine and locomotionor feigning death behaviour was obtained. That is,a significantly higher amount of dopamine was expressedin the adult brain of S than of L strains. The two replicatesof the artificial selection showed the similar dopamineexpression results, indicating that the selection, notgenetic drift, regulates the expression of brain dopamine.This suggests that the heritable difference in brain dopa-mine levels is associated with death-feigning and fleeingor locomotor activity.
Actual amount of brain dopamine has been measured inseveral studies: 0.5 pmol/brain in Drosophila melanogaster(McClung & Hirsh 1999), 2e2.5 pmol/brain in the blowfly Phormia regina (Brookhart et al. 1987), 6e25 pmol/brain in the worker (Harris & Woodring 1992; Tayloret al. 1992) and 50e86 pmol/brain in the queen (Brandeset al. 1990; Harano et al. 2005) of Apis mellifera, and14e40 pmol/brain in the cockroach (Sloley & Owen1982; Barreteau et al. 1993). In the present study, dopa-mine levels in brain and the attached tissues of T. castaneumwere about 6e10 pmol and about 3e5 pmol in short andlong strains, respectively. We measured the amount ofdopamine per contents including brain, suboesophagealganglion, frontal ganglion and mandibular muscle, becausethe beetle’s size was too small to dissect only brain, and thusthe amounts may be higher compared to the other insects ifthe beetle’s size is considered.

ANIMAL BEHAVIOUR, 75, 1120
Starved adults had a lower frequency of death-feigning inthe Colorado potato beetle (Acheampong & Mitchell 1997)and the sweetpotato weevil (Miyatake 2001b). Some studiesshowed that starvation affects the level of biogenic aminesin insect body. For example, serotonin and dopamine levelsin the cockroach’s brain were higher in the starved malesthan in the fed males (Barreteau et al. 1993). In the locustSchistocera gregaria, depriving of food caused an increase inactivity, speed of movement and the level of haemolymphoctopamine (Davenport & Evans 1984). Inactive adultsshowed higher dopamine levels under stress conditions inDrosophila melanogaster (Chentsova et al. 2002). These stud-ies suggest a negative relationship between starving, that is,stressful condition, and feigning death pleiotropically con-trolled by biogenic amines in insects.
Natural selection may act on the level of a neuroactivesubstance pleiotropically controlling immobility and es-cape by running. Many mono amines provide orchestraleffects on insect behaviours (Libersat & Pfluger 2004). Forexample, in the cockroach, dopamine and octopamineenhance, but serotonin decreases the escape response(Goldstein & Camhi 1991; Casagrand & Ritzmann1992). The present study implied that feigning deathand fleeing, possible alternatives as antipredator strategies,are pleiotropically regulated by dopamine. It will be inter-esting to examine the effect of the other biogenic amineson death-feigning behaviour in relation to dopamine,which enhances activity.
Acknowledgments
The authors thank Dr Mike Speed and two anonymousreferees for valuable comments on the manuscript andYukari Takeda, Atsushi Sugita and Yusuke Nishi for insectrearing, and Dr Akira Matsumoto and Prof. Masaki Sakaifor useful advice on neuroactive substances. This work wassupported by KAKENHI 19370011 and 19657026, Grant-in-Aid for Scientific Research, JSPS and MEXT to T.M.
References
Acheampong, S. & Mitchell, B. K. 1997. Quiescence in the Colo-
rado potato beetle, Leptinotarsa decemlineata. Entomologia Experi-
mentalis et Applicata, 82, 83e89.
Adamo,S.A., Linn, C.E.& Hoy, R.R.1995. The role ofneurohormonal
octopamine during ‘‘fight or flight’’ behavior in the field cricket Gryl-lus bimaculatus. Journal of Experimental Biology, 198, 1691e1700.
Agresti, A. 1990. Categorical Data Analysis. New York: J. Wiley.
Anton, A. H. & Sayre, D. F. 1962. A study of the factors affecting the
aluminum oxide-trihydroxyindole procedure for the analysis ofcatecholamines. Journal of Pharmacology and Experimental Thera-
peutics, 138, 360e374.
Barreteau, H., Goudey-Perriere, F., Perriere, C., Jacquot, C., Gayral,P., Grosclaude, J. M. & Brousse-Gaury, P. 1993. Influence on fast-
ing and isolation on biogenic amine levels in the nervous system of
the male cockroach Blaberus craniifer Burm. (Dictyoptera, Blaberi-dae). Comparative Biochemistry and Physiology, 105, 11e16.
Bicker, G. & Menzel, R. 1989. Chemical codes for the control ofbehaviour in arthropods. Nature, 337, 33e39.
Bozic, J. & Woodring, J. 1998. Variations of brain biogenic aminesin mature honeybees and induction of recruitment behavior. Com-
parative Biochemistry and Physiology A, 120, 737e744.
Brandes, C., Sugawa, M. & Menzel, R. 1990. High-performance
liquid chromatography (HPLC) measurement of catecholamines
in single honeybee brains reveals caste-specific differencesbetween worker bees and queens in Apis mellifera. Comparative
Biochemistry and Physiology, 97, 53e57.
Brookhart, G. L., Edgecomb, R. S. & Murdock, L. L. 1987. Amphet-
amine and reserpine deplete brain biogenic amines and alter blow
fly feeding behavior. Journal of Neurochemistry, 48, 1307e1315.
Casagrand, J. L. & Ritzmann, R. E. 1992. Biogenic amines modulate
synaptic transmission between identified giant interneurons and
thoracic interneurons in the escape system of the cockroach.Journal of Neurobiology, 23, 644e655.
Chentsova, N. A., Gruntenko, N. E., Bogomolova, E. V.,Adonyeva, N. V., Karpova, E. K. & Rauschenbach, I. Y. 2002.
Stress response in Drosophila melanogaster strain inactive with
decreased tyramine and octopamine contents. Journal of Compar-
ative Physiology B, 172, 643e650.
Davenport, A. P. & Evans, P. D. 1984. Changes in haemolymph
octopamine levels associated with food deprivation in the locust,Schistocerca gregaria. Physiological Entomology, 9, 269e274.
Donnelly, B. E. & Phillips, T. W. 2001. Functional response ofXylocoris flavipes (Hemiptera: Anthocoridae): effects of prey spe-
cies and habitat. Environmental Entomology, 30, 617e624.
Ebermann, E. 1991. Thanatosis or feigning death in mites of the
family Scutacaridae. In: Acari: Reproduction, Development and
Life-history Strategies (Ed. by R. Schuster & P. W. Murphy), pp.
399e401. London: Chapman & Hall.
Edmunds, M. 1974. Defense in Animals. London: Longman.
Evans, P. D. 1980. Biogenic amines in the insect nervous system.
Advances in Insect Physiology, 15, 317e473.
Fabre, J. H. 1900. Souvenirs Entomologiques. Paris: Delgrave 7 eme
Serie.
Fahrbach, S. E. & Mesce, K. A. 2005. ‘‘Neuroethoendocrinology’’:
integration of field and laboratory studies in insect neuroendocri-
nology. Hormones and Behavior, 48, 352e359.
Fedotov, V. P., Kholodkevitch, S. V. & Udalova, G. P. 2006.
Cardiac activity of freshwater crayfish at wakefulness, rest, and‘‘animal hypnosis’’. Journal of Evolutionary Biochemistry and Physiol-
ogy, 42, 49e59.
Franq, E. N. 1969. Behavioral aspects of feigned death in the opossum
Didelphis marsupialis. American Midland Naturalist, 81, 556e567.
Gargaglioni, L. H., Pereira, A. S. F. & Hoffmann, A. 2001. Basalmidbrain modulation of tonic immobility in the toad Bufo paracne-
mis. Physiology and Behavior, 72, 297e303.
Gart, J. J., Pettigrew, H. M. & Thomas, D. G. 1985. The effect of
bias, variance estimation, skewness and kurtosis of the empirical
logit on weighed least squares analyses. Biometrika, 72, 179e190.
Goldstein, R. S. & Camhi, J. M. 1991. Different effects of the
biogenic amines, dopamine, serotonin and octopamine on thethoracic and abdominal portions of the escape circuit in the cock-
roach. Journal of Comparative Physiology A, 168, 103e112.
Haldane, J. B. S. 1955. The estimation and significance of the loga-rithm of a ratio of frequencies. Annals of Human Genetics, 20,
309e311.
Harano, K., Sasaki, K. & Nagao, T. 2005. Depression of brain dopa-
mine and its metabolite after mating in European honeybee (Apis
mellifera) queens. Naturwissenchaften, 92, 310e313.
Harris, J. W. & Woodring, J. 1992. Effects of stress, age, season, and
source colony on levels of octopamine, dopamine and serotonin in
the honey bee (Apis mellifera L.) brain. Journal of Insect Physiology,38, 29e35.
Hofmann, H. A. & Stevenson, P. A. 2000. Flight restores fight incrickets. Nature, 403, 613.
Holmes, J. S. 1906. Death-feigning in Ranatra. Journal of Compara-tive Neurology and Psychology, 16, 200e216.

MIYATAKE ET AL.: PLEIOTROPIC ANTIPREDATOR STRATEGIES 121
Honma, A., Oku, S. & Nishida, T. 2006. Adaptive significance of
death feigning posture as a specialized inducible defense against
gape-limited predators. Proceedings of the Royal Society of London,Series B, 273, 1631e1636.
Howe, J. C. 1991. Field observations of death feigning in the convicttang, Acanthrus triostegus (Linnaeus), with comments on the noc-
turnal color pattern in juvenile specimens. Journal of Aquariculture
and Aquatic Sciences, 4, 13e15.
Hoyle, G. 1975. Evidence that insect dorsal unpaired median (DUM)
neurons are octopaminergic. Journal of Experimental Zoology, 193,
425e431.
Hoyle, G. & Barker, D. L. 1975. Synthesis of octopamine by insect
dorsal median unpaired neurons. Journal of Experimental Zoology,193, 433e439.
Hozumi, N. & Miyatake, T. 2005. Body-size dependent difference indeath-feigning behavior of adult Callosobruchus chinensis. Journal
of Insect Behavior, 18, 557e566.
King, B. H. & Leaich, H. R. 2006. Variation in propensity to exhibit
thanatosis in Nasonia vitripennis (Hymenoptera: Pteromalidae).
Journal of Insect Behavior, 19, 241e249.
Kume, K., Kume, S., Park, S. K., Hirsh, J. & Jackson, F. R. 2005.
Dopamine is a regulator of arousal in the fruit fly. Journal of
Neuroscience, 25, 7377e7384.
Lehtiniemi, M. 2005. Swin or hide: predator cues cause species
specific reactions in young fish larvae. Journal of Fish Biology, 66,1285e1299.
Libersat, F. & Pfluger, H. J. 2004. Monoamines and the orchestra-tion of behavior. BioScience, 54, 17e25.
Lima, S. L. 1998. Stress and decision making under the risk of preda-tion: recent developments from behavioral, reproductive, and eco-
logical perspectives. Advances in Studies of Behavior, 27, 215e290.
Lima, S. L. & Dill, L. M. 1990. Behavioral decisions made under the
risk of predation: a review and prospectus. Canadian Journal of
Zoology, 68, 619e640.
McCallum, M. L. 1999. Acris crepitans (northern cricket frog) death
feigning. Herpetological Review, 30, 90.
McClung, C. & Hirsh, J. 1999. The trace amine tyramine is essential for
sensitization to cocaine in Drosophila. Current Biology, 9, 853e860.
Martin, J. & Lopez, P. 2000. Costs of refuge use affect escape decisions
of Iberian rock lizards Lacerta monticola. Ethology, 106, 483e492.
Menzel, R., Heyne, A., Kinzel, C., Gerber, B. & Fiala, A. 1999.
Pharmacological dissociation between the reinforcing, sensitizing,
and response-releasing functions of reward in honeybee classical
conditioning. Behavioral Neuroscience, 113, 744e754.
Miyatake, T. 2001a. Diurnal periodicity of death-feigning in Cylas
formicarius (Coleoptera: Brentidae). Journal of Insect Behavior, 14,421e432.
Miyatake, T. 2001b. Effects of starvation on death-feigning in adultsof Cylas formicarius (Coleoptera: Brentidae). Annals of the Entomo-
logical Society of America, 94, 612e616.
Miyatake, T., Katayama, K., Takeda, Y., Nakashima, A., Sugita,A. & Mizumoto, M. 2004. Is death-feigning adaptive? Heritable
variation in fitness difference of death-feigning behaviour. Proceed-
ings of the Royal Society of London, Series B, 271, 2293e2296.
Ohno, T. & Miyatake, T. 2007. Drop or fly? Negative genetic corre-
lation between death-feigning intensity and flying ability as alter-native anti-predator strategies. Proceedings of the Royal Society of
London, Series B, 274, 555e560.
Parajulee, M. N. & Phillips, T. W. 1993. Effect of prey species on
development and reproduction of the predator Lyctocoris campest-
ris (Heteroptera: Anthocoridae). Environmental Entomology, 22,
1035e1042.
Parajulee, M. N., Phillips, T. W. & Hogg, D. B. 1994. Functional
response of Lyctocoris campestris (F.) adults: effects of predator
sex, prey species, and experimental habitat. Biological Control, 4,
80e87.
Prohammer, L. A. & Wade, M. J. 1981. Geographic and genetic
variation in death-feigning behavior in the flour beetle, Tribolium
castaneum. Behavior Genetics, 11, 395e401.
Refshauge, C., Kissinger, P. T., Dreiling, R., Blank, L., Freeman,R. & Adams, R. N. 1974. New high performance liquid chromato-graphic analysis of brain catecholamines. Life Sciences, 14,
311e322.
Rice, W. R. 1989. Analyzing tables of statistical tests. Evolution, 43,
223e225.
Roeder, T. 2005. Tyramine and octopamine: ruling behavior and
metabolism. Annual Review of Entomology, 50, 477.
Ruxton, G. 2006. Grasshoppers don’t play possum. Nature, 440,
880.
Ruxton, G. D., Sherratt, T. N. & Speed, M. P. 2004. Avoiding At-
tack: The Evolutionary Ecology of Crypsis, Warning Signals, and
Mimicry. Oxford: Oxford University Press.
Sargeant, A. B. & Eberhardt, L. E. 1975. Death feigning by ducks in
response to predation by red foxes (Vulpes fulva). American Mid-
land Naturalist, 94, 108e119.
Sasaki, K. & Nagao, T. 2001. Distribution and levels of dopamine
and its metabolites in brains of reproductive workers in honey-bees. Journal of Insect Physiology, 47, 1205e1216.
Sasaki, K. & Nagao, T. 2002. Brain tyramine and reproductive states
of workers in honeybees. Journal of Insect Physiology, 48,1075e1085.
SAS Institute. 2005. JMP6 Release 6.0.3. Cary, North Carolina: SASInstitute Inc.
Scheiner, R., Pluckhahn, S., Oney, B., Blenau, W. & Erber, J. 2002.Behavioural pharmacology of octopamine, tyramine and dopa-
mine in honey bees. Behavioural Brain Research, 136, 545e553.
Scheiner, R., Baumann, A. & Blenau, W. 2006. Aminergic control
and modulation of honeybee behaviour. Current Neuropharmacol-
ogy, 4, 259e276.
Sih, A. 1992. Prey uncertainty and the balancing of antipredator and
foraging needs. American Naturalist, 139, 1052e1069.
Sloley, B. D. & Owen, M. D. 1982. The effects of reserpine on amine
concentrations in the nervous system of the cockroach (Periplanta
americana). Insect Biochemistry, 12, 469e476.
Stevenson, P. A., Hofmann, H. A., Schoch, K. & Schildberger, K.2000. The fight and flight responses of crickets depleted ofbiogenic amines. Journal of Neurobiology, 43, 107e120.
Stevenson, P. A., Dyakonova, V., Rillich, J. & Schldberger, K.2005. Octopamine and experience-dependent modulation of
aggression in crickets. Journal of Neuroscience, 25, 1431e1441.
Tabuchi, K., Moriya, S. & Ishizaki, M. 2007. Locomotion ability
variation among instars of the bean bug, Riptortus pedestris
(Fabricius) (Heteroptera: Alydidae) nymphs. Environmental Ento-
mology, 36, 871e877.
Taylor, D. J., Robinson, G. E., Logan, B. J., Laverty, R. & Mercer,A. R. 1992. Changes in brain amine levels associated with themorphological and behavioural development of the worker
honeybee. Journal of Comparative Physiology A, 170, 715e721.
Unoki, S., Matsumoto, Y. & Mizunami, M. 2005. Participation of
octopaminergic reward system and dopaminergic punishment
system in insect olfactory learning revealed by pharmacological
study. European Journal of Neuroscience, 122, 1409e1416.
Unoki, S., Matsumoto, Y. & Mizunami, M. 2006. Roles of octopa-
minergic and dopaminergic neurons in mediating reward andpunishment signals in insect visual learning. European Journal of
Neuroscience, 24, 2031e2038.
Wasson, K. & Lyon, B. E. 2005. Flight or fight: flexible antipredatory
strategies in porcelain crabs. Behavioral Ecology, 16, 1037e1041.
Copyright © 2022 FDOKUMEN










