
Satellite DNA and heterochromatin of the flour beetle Tribolium confusum
Upload
independentCategory
view
2download
0
www.elsevier.com/locate/ypest
Pesticide Biochemistry and Physiology 85 (2006) 84–90
Flufenoxuron, an acylurea insect growth regulator,alters development of Tribolium castaneum (Herbst)(Coleoptera: Tenebrionidae) by modulating levels of
chitin, soluble protein content, and HSP70 and p34cdc2
in the larval tissues
Shaila Salokhe a,b, Angshuman Sarkar a, Abhijeet Kulkarni a,Samindranath Mukherjee b, Jayanta K. Pal a,*
a Department of Biotechnology, University of Pune, Pune 411007, Indiab Entomology Laboratory, National Chemical Laboratory (CSIR), Pune 411008, India
Received 9 August 2005; accepted 10 October 2005Available online 28 November 2005
Abstract
The effect of sublethal concentrations, 0.00141% (LC20), 0.00251% (LC30), and 0.0036% (LC40) of a dispersible formulation of anacylurea insect growth regulator, flufenoxuron (Cascade) on certain biochemical parameters in the larvae of Tribolium castaneum wasinvestigated. When neonates were fed on diet treated with sublethal concentrations for 24 h, it was observed that at all concentrationstested, there was a significant reduction in chitin content on the 15th day of development. Total soluble protein content at LC20 and LC30
decreased with increasing age of the larvae. At LC20 and LC40 concentrations there was a progressive increase in the protein: chitin ratioas a function of increase in age of the larvae. SDS–PAGE analysis of the larval tissue extracts indicated gross quantitative changes insome of the protein bands (MW 50–97 kDa). Western blot analysis revealed significant increase in the level of HSP70 in the extractsof larvae fed on LC30 treated diet, on the 7th and 10th day of development in the decreasing order. Conversely, a significant decreasein the hyper-phosphorylated form of p34cdc2 kinase due to flufenoxuron treatment indicating modulation of cell cycle regulation wasobserved. Thus, sublethal concentrations of flufenoxuron alter expression of developmentally regulated proteins, HSP70 and p34cdc2
and chitin formation in a stage-specific manner thereby resulting developmental abnormalities in T. castaneum.� 2005 Elsevier Inc. All rights reserved.
Keywords: Tribolium castaneum; Flufenoxuron; Sublethal effects; Development; Total soluble proteins; Chitin; HSP70; p34cdc2; Phosphotyrosine
1. Introduction
Insect growth regulators (IGRs) include compoundsthat affect moulting and metamorphosis by mimickingjuvenile hormone (JH agonists) or usually antagonizingJH activity (ecdysteroid agonists) or by interfering withcuticle formation (chitin synthesis inhibitors) [1–3].
0048-3575/$ - see front matter � 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.pestbp.2005.10.006
* Corresponding author. Fax: +91 20 25691821.E-mail addresses: [email protected], [email protected] (J.K.
Pal).
Although efficacy and toxicological effects of chitin synthe-sis inhibitors have been extensively investigated [3] fewstudies have dealt with their sublethal effects. Radwanet al. [4] studied the effect of sublethal doses of Dimilinon the reproductive performance of Spodoptera littoralis
Boisduval for three consecutive generations by treatingthe fourth instars of each generation. Biddinger and Hull[5] reported on the sublethal effects of several classes ofIGRs on the tufted apple bud moth Platynota idaeusalis.The effects of sublethal concentrations of flufenoxuron onvarious stages in the life cycle and reproductive end points

S. Salokhe et al. / Pesticide Biochemistry and Physiology 85 (2006) 84–90 85
(viz. time to pupation, time to adult emergence, % adultemergence, fecundity, fertilization and hatching success,and larval viability) were reported for the first time inTribolium castaneum by us [6]. Studies on such effects areimportant for the assessment of overall ecological impactof IGRs or pesticides in general since non-target speciesin the periphery of the treated area often receive sublethaldoses. Besides, it would also be interesting to know theresponse of the organism at the molecular level under suchstressful conditions.
Chitin synthesis inhibitors act by interrupting the syn-thesis and transport of specific proteins that are requiredfor the assembly of N-acetyl-D-glucosamine (Glc-NAC)monomers into polymeric chitin [7]. It is also a well-estab-lished fact that living organisms respond at the cellularlevel to unfavorable conditions such as heat or other stress-ful situations including exposure to xenobiotics, UV, heavymetals, oxidizing agents, mutagens, carcinogens, insecti-cides, and gene expression inhibitors, by expression of spe-cific sets of proteins called the heat shock/stress proteins(HSPs) [8–12]. Recent studies indicate that stress proteinsplay a role in toxicity since they are induced as a resultof damages caused to the cell by the toxicant [13,14]. Sim-ilarly, it is also known that precise activation and inactiva-tion of the cyclin-dependent kinases are necessary fornormal cellular proliferation since they play a major rolein controlling the activities of various proteins during thecell cycle by phosphorylating them. Further, similar com-pounds affecting chitin biosynthesis by modulating cAMPdependent protein kinase has also been reported [15].
A review of literature indicates that there is paucity ofinformation on IGR-induced stress and their effect on var-ious biochemical parameters in insects. We have recentlyreported the effect of sublethal concentrations of flufenoxu-ron on fecundity and morphological abnormalities duringdevelopment of T. castaneum [6]. The present study is acontinuation of the earlier one [6] with an emphasis onthe biochemical and molecular changes as an effect of sub-lethal concentrations of flufenoxuron (Cascade) on the lar-val tissues of T. castaneum (Herbst) (Coleoptera:Tenebrionidae). We have determined the effects on chitin,total soluble protein as well as a stress protein (HSP70)and a cell cycle regulatory protein (p34cdc2), since theseare considered to be general indicators of sublethal cellularprotein damage [16].
2. Materials and methods
A stock culture of T. castaneum was maintained on adiet containing wheat flour and 5% Brewers yeast, at29 ± 1 �C and 60% relative humidity. Eggs were collectedby sieving (sieve number 40) diet infested with adults. Adispersible concentrate formulation of flufenoxuron (Cas-cade) belonging to the benzoylphenylurea family wasmixed at various sublethal concentrations (LC20, LC30,and LC40) in diet using acetone as a carrier solvent. Ace-tone treated diet was used as control.
All reagents were either purchased from Sigma Chemi-cals, USA or Life Technologies (Gibco-BRL, USA). Thesources of specific chemicals are as follows: p-dimethylami-nobenzaldehyde (DMAB) was purchased from Qualigensfine chemicals. Potassium tetraborate was purchasedfrom Sigma Chemicals, Coomassie brilliant blue G(CBB), and bovine serum albumin (BSA) were purchasedfrom Sigma Chemicals, USA. Protein molecular weightmarkers were purchased from Bangalore Genei, India.Anti-HSP70, anti-p34cdc2 and anti-phosphotyrosine mono-clonal antibodies, and the secondary antibodies (anti-mouse IgG-alkaline phosphatase) were purchased fromSigma Chemicals, USA. Nitrocellulose membrane waspurchased from Life Technologies (Gibco-BRL), India.
2.1. Chitin extraction and estimation
The effects of sublethal concentrations of flufenoxuronon chitin content of the larvae were determined by releas-ing the neonates in the treated diet for 24 h. Subsequently,the larvae were transferred to normal diet. Diet mixed withappropriate quantity of acetone was used as control. Fivereplicates of 250 larvae each were prepared. After the treat-ment, 200 larvae on 7th day, 150 larvae on 10th day and100 larvae on 15th day were weighed and homogenizedin homogenizing buffer (100 mg/ml), containing Tris(5 mM), glycine (38 mM), pH 8.4. The homogenate wascentrifuged at 10,000 rpm for 15 min and the precipitatewas dissolved in 6 N HCl. The dissolved precipitate wastransferred into hydrolysis tubes which were sealed undervacuum and the material was hydrolyzed at 100 �C for16 h in a temperature controlled heating block. On comple-tion of hydrolysis, the solution was centrifuged at10,000 rpm for 10 min. The supernatant was collectedand neutralized with 30% NaOH. N-Acetylglucosaminecontent was estimated according to the method of Ressiget al. [17]. The entire experiment was repeated thrice ondifferent occasions.
Glucosamine 0.1–1lmole was taken in a capped testtube. To this was added 1.5% acetic anhydride (v/v) pre-pared in acetone and 1 ml potassium tetraborate(100 mM), pH 8.5. The tube was sealed tightly, incubatedin boiling water bath for 3 min and cooled immediatelyin an ice bath. Subsequently 3 ml of DMAB [10% DMABprepared in acetic acid containing 12.5% 10 N HCl (v/v)]diluted with acetic acid (1:9) was added to the solution inthe tube. The tube was incubated at 37 �C for 20 min.Absorbance of the mixture was measured at 585 nm in aJasco V-550 UV–visible spectrophotometer.
2.2. Protein extraction and estimation
Neonates of T. castaneum were fed on control- andtreated-diet (with sublethal concentrations of flufenoxuron,LC20, LC30, and LC40) for 24 h and then transferred tonormal diet. Five replications of 250 larvae each were madefor each concentration. After the treatment, 200 larvae on
A Quantitative analysis of chitin
00.5
11.5
22.5
33.5
7th 10th 15th
7th 10th 15th
Days
dy
oblavral
gu/
nitihc
gn
th
giew
control
LC-20
LC-30
LC-40
B Quantitative analysis of protein
0
5
10
15
20
25
Daysd
yo
blavralg
u/niet
orp
gn
th
giew
control
LC-20
LC-30
LC-40
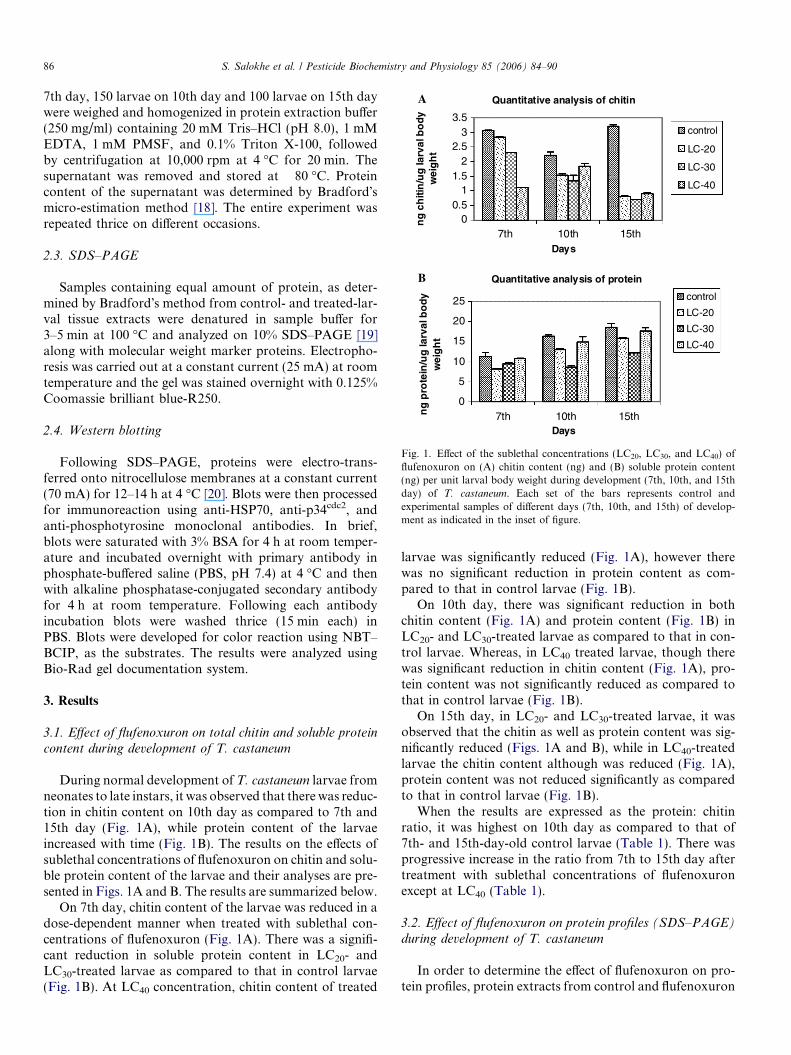
Fig. 1. Effect of the sublethal concentrations (LC20, LC30, and LC40) offlufenoxuron on (A) chitin content (ng) and (B) soluble protein content(ng) per unit larval body weight during development (7th, 10th, and 15thday) of T. castaneum. Each set of the bars represents control andexperimental samples of different days (7th, 10th, and 15th) of develop-ment as indicated in the inset of figure.
86 S. Salokhe et al. / Pesticide Biochemistry and Physiology 85 (2006) 84–90
7th day, 150 larvae on 10th day and 100 larvae on 15th daywere weighed and homogenized in protein extraction buffer(250 mg/ml) containing 20 mM Tris–HCl (pH 8.0), 1 mMEDTA, 1 mM PMSF, and 0.1% Triton X-100, followedby centrifugation at 10,000 rpm at 4 �C for 20 min. Thesupernatant was removed and stored at �80 �C. Proteincontent of the supernatant was determined by Bradford�smicro-estimation method [18]. The entire experiment wasrepeated thrice on different occasions.
2.3. SDS–PAGE
Samples containing equal amount of protein, as deter-mined by Bradford�s method from control- and treated-lar-val tissue extracts were denatured in sample buffer for3–5 min at 100 �C and analyzed on 10% SDS–PAGE [19]along with molecular weight marker proteins. Electropho-resis was carried out at a constant current (25 mA) at roomtemperature and the gel was stained overnight with 0.125%Coomassie brilliant blue-R250.
2.4. Western blotting
Following SDS–PAGE, proteins were electro-trans-ferred onto nitrocellulose membranes at a constant current(70 mA) for 12–14 h at 4 �C [20]. Blots were then processedfor immunoreaction using anti-HSP70, anti-p34cdc2, andanti-phosphotyrosine monoclonal antibodies. In brief,blots were saturated with 3% BSA for 4 h at room temper-ature and incubated overnight with primary antibody inphosphate-buffered saline (PBS, pH 7.4) at 4 �C and thenwith alkaline phosphatase-conjugated secondary antibodyfor 4 h at room temperature. Following each antibodyincubation blots were washed thrice (15 min each) inPBS. Blots were developed for color reaction using NBT–BCIP, as the substrates. The results were analyzed usingBio-Rad gel documentation system.
3. Results
3.1. Effect of flufenoxuron on total chitin and soluble protein
content during development of T. castaneum
During normal development of T. castaneum larvae fromneonates to late instars, it was observed that there was reduc-tion in chitin content on 10th day as compared to 7th and15th day (Fig. 1A), while protein content of the larvaeincreased with time (Fig. 1B). The results on the effects ofsublethal concentrations of flufenoxuron on chitin and solu-ble protein content of the larvae and their analyses are pre-sented in Figs. 1A and B. The results are summarized below.
On 7th day, chitin content of the larvae was reduced in adose-dependent manner when treated with sublethal con-centrations of flufenoxuron (Fig. 1A). There was a signifi-cant reduction in soluble protein content in LC20- andLC30-treated larvae as compared to that in control larvae(Fig. 1B). At LC40 concentration, chitin content of treated
larvae was significantly reduced (Fig. 1A), however therewas no significant reduction in protein content as com-pared to that in control larvae (Fig. 1B).
On 10th day, there was significant reduction in bothchitin content (Fig. 1A) and protein content (Fig. 1B) inLC20- and LC30-treated larvae as compared to that in con-trol larvae. Whereas, in LC40 treated larvae, though therewas significant reduction in chitin content (Fig. 1A), pro-tein content was not significantly reduced as compared tothat in control larvae (Fig. 1B).
On 15th day, in LC20- and LC30-treated larvae, it wasobserved that the chitin as well as protein content was sig-nificantly reduced (Figs. 1A and B), while in LC40-treatedlarvae the chitin content although was reduced (Fig. 1A),protein content was not reduced significantly as comparedto that in control larvae (Fig. 1B).
When the results are expressed as the protein: chitinratio, it was highest on 10th day as compared to that of7th- and 15th-day-old control larvae (Table 1). There wasprogressive increase in the ratio from 7th to 15th day aftertreatment with sublethal concentrations of flufenoxuronexcept at LC40 (Table 1).
3.2. Effect of flufenoxuron on protein profiles (SDS–PAGE)
during development of T. castaneum
In order to determine the effect of flufenoxuron on pro-tein profiles, protein extracts from control and flufenoxuron
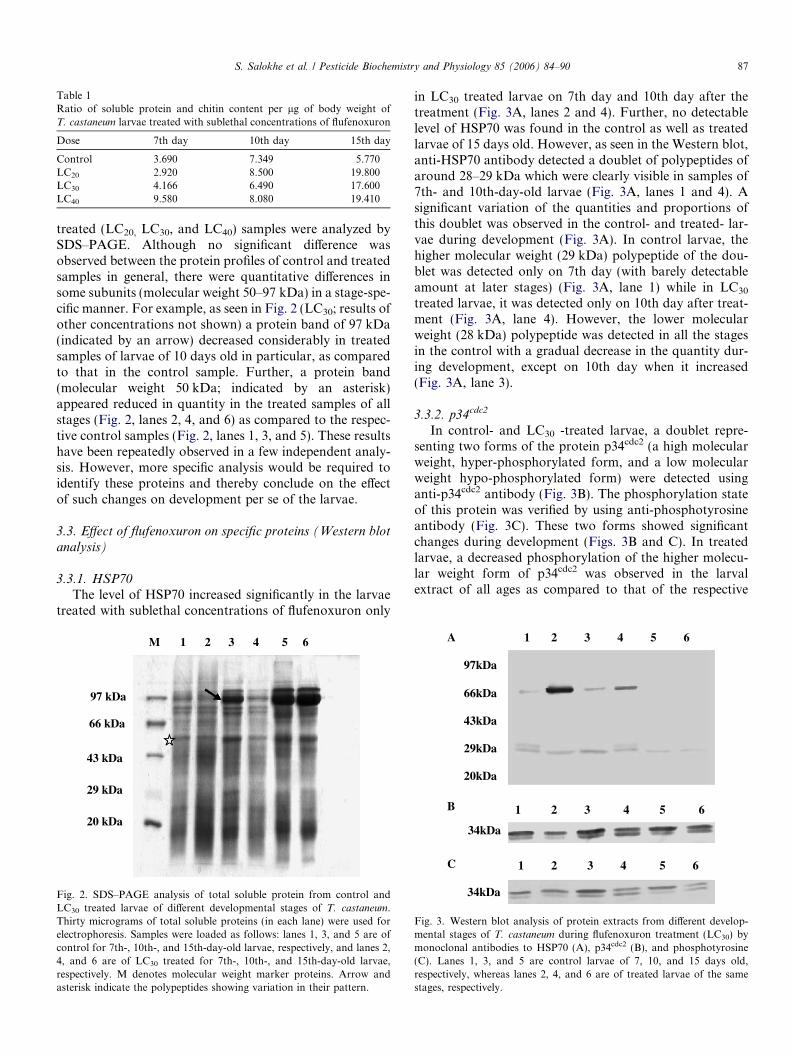
Table 1Ratio of soluble protein and chitin content per lg of body weight ofT. castaneum larvae treated with sublethal concentrations of flufenoxuron
Dose 7th day 10th day 15th day
Control 3.690 7.349 5.770LC20 2.920 8.500 19.800LC30 4.166 6.490 17.600LC40 9.580 8.080 19.410
S. Salokhe et al. / Pesticide Biochemistry and Physiology 85 (2006) 84–90 87
treated (LC20, LC30, and LC40) samples were analyzed bySDS–PAGE. Although no significant difference wasobserved between the protein profiles of control and treatedsamples in general, there were quantitative differences insome subunits (molecular weight 50–97 kDa) in a stage-spe-cific manner. For example, as seen in Fig. 2 (LC30; results ofother concentrations not shown) a protein band of 97 kDa(indicated by an arrow) decreased considerably in treatedsamples of larvae of 10 days old in particular, as comparedto that in the control sample. Further, a protein band(molecular weight 50 kDa; indicated by an asterisk)appeared reduced in quantity in the treated samples of allstages (Fig. 2, lanes 2, 4, and 6) as compared to the respec-tive control samples (Fig. 2, lanes 1, 3, and 5). These resultshave been repeatedly observed in a few independent analy-sis. However, more specific analysis would be required toidentify these proteins and thereby conclude on the effectof such changes on development per se of the larvae.
3.3. Effect of flufenoxuron on specific proteins (Western blot
analysis)
3.3.1. HSP70The level of HSP70 increased significantly in the larvae
treated with sublethal concentrations of flufenoxuron only
M 1 2 3 4 5 6
97 kDa
66 kDa
43 kDa
29 kDa
20 kDa
Fig. 2. SDS–PAGE analysis of total soluble protein from control andLC30 treated larvae of different developmental stages of T. castaneum.Thirty micrograms of total soluble proteins (in each lane) were used forelectrophoresis. Samples were loaded as follows: lanes 1, 3, and 5 are ofcontrol for 7th-, 10th-, and 15th-day-old larvae, respectively, and lanes 2,4, and 6 are of LC30 treated for 7th-, 10th-, and 15th-day-old larvae,respectively. M denotes molecular weight marker proteins. Arrow andasterisk indicate the polypeptides showing variation in their pattern.
in LC30 treated larvae on 7th day and 10th day after thetreatment (Fig. 3A, lanes 2 and 4). Further, no detectablelevel of HSP70 was found in the control as well as treatedlarvae of 15 days old. However, as seen in the Western blot,anti-HSP70 antibody detected a doublet of polypeptides ofaround 28–29 kDa which were clearly visible in samples of7th- and 10th-day-old larvae (Fig. 3A, lanes 1 and 4). Asignificant variation of the quantities and proportions ofthis doublet was observed in the control- and treated- lar-vae during development (Fig. 3A). In control larvae, thehigher molecular weight (29 kDa) polypeptide of the dou-blet was detected only on 7th day (with barely detectableamount at later stages) (Fig. 3A, lane 1) while in LC30
treated larvae, it was detected only on 10th day after treat-ment (Fig. 3A, lane 4). However, the lower molecularweight (28 kDa) polypeptide was detected in all the stagesin the control with a gradual decrease in the quantity dur-ing development, except on 10th day when it increased(Fig. 3A, lane 3).
3.3.2. p34cdc2
In control- and LC30 -treated larvae, a doublet repre-senting two forms of the protein p34cdc2 (a high molecularweight, hyper-phosphorylated form, and a low molecularweight hypo-phosphorylated form) were detected usinganti-p34cdc2 antibody (Fig. 3B). The phosphorylation stateof this protein was verified by using anti-phosphotyrosineantibody (Fig. 3C). These two forms showed significantchanges during development (Figs. 3B and C). In treatedlarvae, a decreased phosphorylation of the higher molecu-lar weight form of p34cdc2 was observed in the larvalextract of all ages as compared to that of the respective
Fig. 3. Western blot analysis of protein extracts from different develop-mental stages of T. castaneum during flufenoxuron treatment (LC30) bymonoclonal antibodies to HSP70 (A), p34cdc2 (B), and phosphotyrosine(C). Lanes 1, 3, and 5 are control larvae of 7, 10, and 15 days old,respectively, whereas lanes 2, 4, and 6 are of treated larvae of the samestages, respectively.

88 S. Salokhe et al. / Pesticide Biochemistry and Physiology 85 (2006) 84–90
controls (Figs. 3B and C). Further, it was observed thatthere was a substantial decrease in the overall basal levelof p34cdc2 during flufenoxuron treatment (Figs. 3B and C).
4. Discussion
Tribolium castaneum neonates fed on diet treated withvarious sublethal concentrations of flufenoxuron for 24 hshowed overall reduction in the total soluble protein con-tent at all concentrations except at LC40 during their devel-opment. SDS–PAGE protein analysis revealed quantitativevariation in some protein subunits (50–97 kDa) in responseto oral administration of sublethal concentrations of flufe-noxuron through diet. In general, the soluble protein con-tent/unit body weight decreased with an increase inconcentration of flufenoxuron. Similar results were report-ed by Kulkarni and Mehrotra [21] in Schistocerca gregaria
due to Sumithion treatment. However, it is to be noted thatthe mechanisms of action of these two chemicals appear tobe different. Proteins of 50 kDa were expressed more incontrol- than in treated-larvae. Such proteins are presumedto be associated with enzymatic activities related with thecuticle (sclerotization, melanin formation, and catechol-amine metabolism). Deficiency of such proteins interfereswith the digestion of cuticle during moulting as observedin Drosophila melanogaster and Manduca sexta by Marcuand Locke [22]. The quantitative reduction of these pro-teins in treated larvae in the present study possibly resultedin larval–pupal and pupal–adult intermediates in T. casta-
neum as reported earlier [6].Observations on total chitin content of the larvae treat-
ed with sublethal concentrations of flufenoxuron revealedthat reduction in chitin content was more significant on15th day after the treatment. Further, protein: chitin ratiowas higher on 10th day as compared to 7th and 15th day incontrol larvae. Whereas in treated larvae, the ratioincreased with an increase in concentration of flufenoxuronfrom 7th to 15th day, except at LC40 in which there wasdecrease in the ratio on 10th day. Our results thus, are inagreement with those observed by Ishaaya and Casida[23] and Clarke and Jewess [24]. Similar dose-dependenteffects of flufenoxuron on protein: chitin ratio with timeelapsing after flufenoxuron treatment was recently reportedby Sammour and EL-Ansary [25] in case of S. littoralis lar-vae. Cross-linking of chitin and protein to form cuticleplays a crucial role in the development of insect. Variationsin protein: chitin ratio due to treatment with sublethal con-centrations of flufenoxuron affected growth and develop-ment of T. castaneum as reflected by decrease in larvalweight, delay in the development of pupal and adult stagesas well as formation of abnormal adults, as reportedearlier [6].
It was interesting to note that at higher concentration offlufenoxuron (LC40), the total soluble protein and chitincontent of the treated larvae were not much affected. Thiscould possibly be due to the effect of higher doses whichact as a trigger for activation of detoxifying enzymes as
has been shown in case of Triatoma infestans [26] and alsoin case of T. castaneum in response to azadirachtin [27].Further, involvement of some low molecular weight(23 kDa) proteins in vitellogenesis and embryogenesis hasbeen reported in case of Dipteran larva by Robert andBrock [28]. Here, we report similar observations withrespect to the effect of LC40 on T. castaneum larvae,wherein quantitative changes in proteins with similarmolecular size were found (data not shown). In our earlierstudy [6], it was demonstrated that larvae fed on LC40 afterreaching adulthood exhibited reduction in fecundity. Thelow molecular weight proteins reported herein may haveaffected fecundity in adults as reported earlier [6], a possi-bility which cannot be ruled out.
Increased expression of one of the major stress proteins,HSP70 in the larvae treated with sublethal concentration offlufenoxuron was significant in LC30 treated larvae on 7thand 10th day after the treatment. Further, it was repeatedlyobserved that there was cross reactivity of HSP70 withsome low molecular weight (28 and 29 kDa) proteins,which seemed to resolve in the gel near the mobility ofthe small heat shock proteins. Alternatively, these lowmolecular weight proteins may possibly be the proteolyticfragments of HSP70 as observed by Rybczynski and Gil-bert [29] in M. sexta. These low molecular weight proteinswere found in the early developmental period (7-day-oldlarvae) and their expression reduced gradually during sub-sequent stages of development. Proteins of low molecularweight (29 kDa) were found to be the major haemolymphproteins (MHP) in Bombyx mori by Bosquet and Clavez[30]. Further, Plantevin et al. [31] found that the rates ofsynthesis of these MHP showed quantitative changes dur-ing development in B. mori and their synthesis was regulat-ed by juvenile hormone (JH) titer. In LC30 treated larvae,the induction of 29 kDa protein expression on 10th day(Fig. 2, lane 4) coincides with the prolongation of the larvalperiod. However, it is not known as to how flufenoxuroninfluences JH titer, which in turn modulates expression ofsuch proteins. There appears to be some correlationbetween the expression of these low molecular weight pro-teins (small HSPs of 28 and 29 kDa) and the phosphoryla-tion state of p34cdc2. However, further investigations arerequired to explain the significance of such observations.Absence of expression of HSP70 (results not shown) athigher concentration (LC40) is more likely to be relatedto collapse of the cellular machinery, rather than its inabil-ity to induce HSP70 as observed by Bierkens et al. [32] inRaphidocelis subcapitula. Thus, it appears that inducedexpression of HSP70 on 7th and 10th day provides themain protection against insect growth regulatory stress atLC30 concentration of flufenoxuron.
A key step in regulating the entry of eukaryotic cells intomitosis is the activation of protein kinase p34cdc2 by changein its phosphorylation state. Therefore, it was interestingto observe the changes in the phosphorylation statusof p34cdc2 during the developmental process of T. castane-
um larvae treated with sublethal concentrations of

S. Salokhe et al. / Pesticide Biochemistry and Physiology 85 (2006) 84–90 89
flufenoxuron. p34cdc2 from treated- and control-tissue wasobserved as two bands by immunoblotting. Tyr15 phos-phorylation normally functions to delay M phase by pre-venting p34cdc2 activation [33]. In the present study,tyrosine was found to be less phosphorylated in LC30 treat-ed larvae than that of the control of the same stage (Figs.3B and C). Thus, these data suggest that p34cdc2 is under-going activation through partial dephosphorylation (tyro-sine residue) during the treatment. Similar findings on theselective activation of PKA [15] and phosphorylation of aprotein of 39 kDa [34] for diflubenzuron treated Americancockroach, Periplaneta americana have been reported.Activation of p34cdc2 might cause developing larval cellsto enter mitosis prematurely which is reflected by the effectsproduced on life cycle as well as reproductive end points,reported by us earlier [6].
Flufenoxuron at sublethal concentrations was found toaffect larval weight, percent pupation and percent adultemergence, as well as time taken for adult emergence in adose-dependent manner [6]. A small proportion of larval–pupal as well as pupal–adult intermediates were observedat all concentrations. The fecundity of adults emergingfrom treated larvae was greatly reduced and at higher con-centrations, deformed adults with short life span wereobserved [6]. In the present study, we found that at themolecular level the two important components, chitinand total soluble proteins, which are involved in the growthand developmental process of the insect were significantlyaffected due to treatment with sublethal concentrations offlufenoxuron, which coincided with developmental as wellas reproductive impairments. The cellular response in theform of induction of HSP70 was observed on 7th and10th day after treatment in case of larvae fed on LC30 treat-ed diet along with the phosphorylation state and activationof cell cycle regulatory protein, p34cdc2. This is furtherestablished by the absence of expression of p34cdc2 in caseof larvae fed on LC40 concentration of flufenoxuron andalso reflected by the effects produced on life cycle as wellas reproductive endpoints, reported by us earlier [6].
In conclusion, these biochemical data suggest that theeffect of flufenoxuron on fecundity, early embryonic abnor-malities and later processes such as moulting, formation ofpupae and adults of T. casteneum may be mediatedthrough perturbation of cell cycle as well as altered expres-sion of developmentally regulated chitin and proteinsinvolved in cuticle formation in a stage-specific manner.
Acknowledgments
Financial help in the form of a research grant fromCouncil of Scientific and Industrial Research (CSIR) [Pro-ject No. 9/137 (377)/2001-EMR-I] to JKP is duly acknowl-edged. A.S. was supported by a fellowship from CSIR(SRF). A.K. is supported by a fellowship from UGC(JRF). We thank Dr. Anita Kar for permitting us to useBio-Rad Gel documentation system for protein analysis.
References
[1] H. Smet, M. Rans, A. DeLoof, Comparative effectiveness of insectgrowth regulators with juvenile hormone, anti-juvenile hormone andchitin synthesis inhibiting activity against several stored food insectpests, In: Fleurat-Lessard, F., Ducon, P. (Eds.). Proceedings 5thInternational Working Conference on Stored Product Protection,France, 1990, pp. 649–657.
[2] H. Oberlander, D.L. Silhacek, E. Leach, I. Ishaaya, E. Shaaya,Benzoylphenyl urea inhibits chitin synthesis without interfering withamino sugar uptake in imaginal wing discs of Plodia interpuntella,Arch. Insect Biochem. Physiol. 18 (1991) 219–227.
[3] H. Oberlander, D.L. Silhacek, E. Shaaya, I. Ishaaya, Current statusand future perspectives of the use of insect growth regulators for thecontrol of stored product insects, J. Stored Prod. Res. 33 (1997) 1–6.
[4] H.S. Radwan, A.M.R. Abo-Elghar, I.M.A. Ammar, Reproductiveperformance of Spodoptera littoralis (Boisd.) treated topically withsub lethal doses of an anti-moulting IGR (Dimilin), Z. Angew.Entomol. 86 (1978) 414–419.
[5] D.J. Biddinger, L.A. Hull, Sub lethal effects of selected insecticides ongrowth and reproduction of a laboratory susceptible strain of tuftedapple bud moth (Lepidoptera: Tortricidae), J. Econ. Entomol. 92(1999) 314–323.
[6] S.G. Salokhe, J.K. Pal, S.N. Mukherjee, Effect of sub lethalconcentrations of flufenoxuron on growth, development, and repro-ductive performance of Tribolium castaneum (Herbst.), Invertebr.Reprod. Dev. 43 (2003) 141–150.
[7] H. Oberlander, D.L. Silhacek, New perspectives on the modes ofaction of Benzoylphenyl urea insecticides, in: I. Ishaaya, D. Degheele(Eds.), Insecticides with Novel Modes of Action, Narosa PublishingHouse, New Delhi, 1998.
[8] S. Lindquist, The heat shock response, Annu. Rev. Biochem. 55(1986) 1151–1191.
[9] L. Nover, The Heat Shock Response, CRC Press, Boca Raton, 1991.[10] M.E. Feeder, Ecological and evolutionary physiology of stress
proteins and the stress response: the Drosophila melanogaster model,in: I.A. Johnston (Ed.), Animals and Temperature: Phenotypic andEvolutionary Adaptation, Cambridge University Press, Cambridge,1996, pp. 79–102.
[11] U. Fiege, R.I. Morimoto, B.S. Polla, Stress Inducible CellularResponses, Birkhauser Verlag, Basel, Switzerland, 1996.
[12] D.L. Delinger, G.D. Yocum, Physiology of heat sensitivity, in: G.J.Hallman, D.L. Delinger (Eds.), Temperature Sensitivity in Insectsand Application in Integrated Pest Management, Westviews Press,Oxford, 1998, pp. 7–53.
[13] L.E. Hightower, Heat shock, stress proteins, chaperones, andproteotoxicity, Cell 66 (1991) 191–197.
[14] B.M. Sanders, Stress proteins in aquatic organisms: an environmentalperspective, Crit. Rev. Toxicol. 23 (1993) 49–75.
[15] S. Ishii, F. Matsumura, Diflubenzuron-induced changes in activitiesof the cAMP dependent protein kinase in the newly moltedintegument of American cockroach in situ and in cell free conditions,Insect Biochem. Mol. Biol. 34 (2004) 743–752.
[16] I. Werner, J. Geist, M. Okihiro, P. Roisenkranz, D. Hinton, Effect ofdietary exposure to the pyrethroid pesticide esfenvalerate on medaka(Oryzias latipes), Marine Environ. Res. 54 (2002) 609–614.
[17] J.L. Ressig, J.L. Strominger, L.F. Lefloir, Modified colorimetricmethod for the estimation of N-acetyl amino sugars, J. Biol. Chem.217 (1955) 959–966.
[18] M.M. Bradford, A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein–dyebinding, Anal. Biochem. 72 (1976) 248–254.
[19] U.K. Laemmli, Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4, Nature (London) 227 (1970) 680–685.
[20] H. Towbin, T. Staehelin, J. Gordon, Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose: procedure andsome applications, Proc. Natl. Acad. Sci. USA 83 (1979) 4849–4853.

90 S. Salokhe et al. / Pesticide Biochemistry and Physiology 85 (2006) 84–90
[21] A.P. Kulkarni, K.N. Mehrotra, Effects of dieldrin and sumithion onamino acid nitrogen and protein in haemolymph of desert locust,Schistocerca gregaria, Pestic. Biochem. Physiol. 3 (1975) 420–431.
[22] O. Marcu, M. Locke, A cuticular protein from the moulting stages ofan insect, Insect Biochem. Mol. Biol. 28 (1998) 659–669.
[23] I. Ishaaya, J.E. Casida, Dietary TH6040 alters composition andenzyme activity of housefly larval cuticle, Pestic. Biochem. Physiol. 4(1974) 484–490.
[24] B.S. Clarke, P.J. Jewess, The inhibition of chitin synthesis inSpodoptera littoralis larvae by flufenoxuron, teflubenzuron, anddiflubenzuron, Pestic. Sci. 28 (1990) 377–388.
[25] E.A. Sammour, A. EL-Ansary, Mechanism of action of acylureaflufenoxuron on larvae of the Egyptian leaf worm Spodoptera
littoralis, J. Egypt. Ger. Soc. Zool. 21 (A) (1996) 27–47.[26] E. Wood, N. Casebe, F. Melgar, E. Zerba, Distribution and
properties of glutathione-S-transferase from Triatoma infestans,Comp. Biochem. Physiol. 84 (1986) 607–618.
[27] S.N. Mukherjee, S.K. Rawal, S.S. Ghumare, R.N. Sharma, Hormeticconcentrations of azadirachtin and isoesterase profiles in T. castaneum
(Herbst) (Coleoptera: Tenebrionidae), Experientia 49 (1993) 557–560.[28] D.B. Robert, H.W. Brock, The major serum proteins of Diptera
larvae, Experientia 37 (1981) 103–110.
[29] R. Rybczynski, L.I. Gilbert, cDNA cloning and expression of ahormone regulated heat shock protein (hsc 70) from the prothoracicgland of Manduca Sexta, Insect Biochem. Mol. Biol. 30 (2000)579–589.
[30] G. Bosquet, B. Clavez, Juvenile hormone modifications of geneexpression in fat body and posterior silk glands of Bombyx mori, J.Insect Physiol. 31 (1985) 603–610.
[31] G. Plantevin, G. Bosquet, B. Calvez, C. Nardon, Relationshipsbetween juvenile hormone levels and synthesis of major haemolymphproteins in Bombyx mori larvae, Comp. Biochem. Physiol. 86B (1987)501–507.
[32] J. Bierkens, J. Maes, F. Vander Plaetse, Dose-dependent induction ofheat shock protein Hsp 70 synthesis in Raphidocelis subcapitata
following exposure to different classes of environmental pollutants,Environ. Pollution 101 (1998) 91–97.
[33] K.L. Gould, P. Nurse, Tyrosine phosphorylation of the fission yeastcdc2+ protein kinase regulates entry into mitosis, Nature 342 (1989)39–45.
[34] Y. Nakagawa, S. Ishii, F. Matsumura, Diflubenzuron stimulatesphosphorylation of a 39 kDa protein from newly molted Americancockroach (Periplaneta americana), Insect Biochem. Mol. Biol. 26(1996) 891–898.
Copyright © 2022 FDOKUMEN








