
Maintenance of segment and appendage primordia by the Tribolium gene knödel
10
Maintenance of segment and appendage primordia by the Tribolium gene kno ¨del Hilde Wohlfrom a , Johannes Benno Schinko b , Martin Klingler a , Gregor Bucher b, * a Institut fu ¨r Biologie, Friedrich-Alexander-Universita ¨t Erlangen, Staudtstrasse 5, 91058 Erlangen, Germany b Johann-Friedrich-Blumenbach-Institut fu ¨r Zoologie und Anthropologie, Georg-August-Universita ¨t Go ¨ttingen, Abteilung Entwicklungsbiologie, Justus-von-Liebig-Weg 11, 37077 Go ¨ttingen, Germany Received 2 November 2005; received in revised form 11 April 2006; accepted 26 April 2006 Available online 5 May 2006 Abstract For homeotic and segment-polarity genes in Drosophila, a switch in gene regulation has been described that distinguishes patterning and maintenance phases. Maintenance of segment and organ primordia involves secondary patterning and differentiation steps, as well as survival factors regulating proliferation and organ size. In a screen for embryonic lethal mutations in the flour beetle Tribolium castaneum, we have recovered two alleles of the kno ¨del gene, which result in short, bag-like embryos. These embryos have severely reduced appendages and differentiate a cuticle that lacks most overt signs of segmentation. In addition, they lack bristles and display defects in the nervous system. Early patterning in kno ¨del mutant embryos is normal up to the extended germ band stage, as indicated by the formation of regular even-skipped (Tc’eve) and wingless (Tc’wg) stripes. Afterwards, however, these patterns degenerate. Similarly, proximo-distal growth and patterning of limbs are nearly normal initially, but limb primordia shrink, and proximo-distal patterns degenerate, during subsequent stages. kno ¨del could be a segment polarity gene required for segment border maintenance in both trunk and appendages. Alternatively, it may have a more general role in tissue or organ maintenance. q 2006 Elsevier Ireland Ltd. All rights reserved. Keywords: Tribolium; knodel; Naked cuticle; Segmentation; Segment polarity; Maintenance; leg development 1. Introduction The principles underlying the formation of Drosophila body segments are well understood (Pankratz and Jackle, 1990; St Johnston and Nusslein-Volhard, 1992). Equally important, but less understood, is the maintenance of established segment and organ primordia. During pattern formation the embryo is subdivided in ever smaller units as for instance seen by the expression domains of gap and pair rule genes. These subdivi- sions are transient in nature and are not directly transformed into morphological traits. Only the last level of the segmenta- tion hierarchy, the segment polarity pattern, provides direct cues for morphogenesis, i.e. compartment boundaries, cuticle differentiation or muscle attachment sites (Martinez Arias et al., 1988; Sanson, 2001; Larsen et al., 2003). Pattern maintenance, in contrast, requires stable cues that are able to maintain borders and regulate the size of morphological units. Only few of these cues are known. Mutual activation of wingless and engrailed expression is crucial for the maintenance of parasegmental boundaries during germ band extension. Afterwards, their expression domains become independent from each other (Heemskerk et al., 1991; Martinez Arias, 1993; DiNardo et al., 1994). Engrailed expression then comes under cell autono- mous control involving the action of polycomb group genes (Moazed and O’Farrell, 1992). In the case of the segment polarity gene gooseberry, different enhancer elements are responsible for early and late functions (Li et al., 1993). Adjustment of parasegment size involves cell death and delamination of miss-specified cells (Hughes and Krause, 2001). In wing imaginal discs, maintenance of parasegmental borders involves hedgehog regulated cell sorting (Dahmann and Basler, 2000). It has been assumed that differential cell adhesion may be involved but the molecules responsible for the hypothetical differential cell adhesion remains elusive despite specific screening efforts (Dahmann and Basler, 1999). All conclusions drawn from Drosophila research bear the drawback that this species develops through a highly derived Mechanisms of Development 123 (2006) 430–439 www.elsevier.com/locate/modo 0925-4773/$ - see front matter q 2006 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.mod.2006.04.003 * Corresponding author. Tel.: C49 551 395426; fax: C49 551 395416. E-mail address: [email protected] (G. Bucher).
Transcript of Maintenance of segment and appendage primordia by the Tribolium gene knödel

Maintenance of segment and appendage primordia
by the Tribolium gene knodel
Hilde Wohlfrom a, Johannes Benno Schinko b, Martin Klingler a, Gregor Bucher b,*
a Institut fur Biologie, Friedrich-Alexander-Universitat Erlangen, Staudtstrasse 5, 91058 Erlangen, Germanyb Johann-Friedrich-Blumenbach-Institut fur Zoologie und Anthropologie, Georg-August-Universitat Gottingen, Abteilung Entwicklungsbiologie,
Justus-von-Liebig-Weg 11, 37077 Gottingen, Germany
Received 2 November 2005; received in revised form 11 April 2006; accepted 26 April 2006
Available online 5 May 2006
Abstract
For homeotic and segment-polarity genes in Drosophila, a switch in gene regulation has been described that distinguishes patterning and
maintenance phases. Maintenance of segment and organ primordia involves secondary patterning and differentiation steps, as well as survival
factors regulating proliferation and organ size. In a screen for embryonic lethal mutations in the flour beetle Tribolium castaneum, we have
recovered two alleles of the knodel gene, which result in short, bag-like embryos. These embryos have severely reduced appendages and
differentiate a cuticle that lacks most overt signs of segmentation. In addition, they lack bristles and display defects in the nervous system. Early
patterning in knodel mutant embryos is normal up to the extended germ band stage, as indicated by the formation of regular even-skipped (Tc’eve)
and wingless (Tc’wg) stripes. Afterwards, however, these patterns degenerate. Similarly, proximo-distal growth and patterning of limbs are nearly
normal initially, but limb primordia shrink, and proximo-distal patterns degenerate, during subsequent stages. knodel could be a segment polarity
gene required for segment border maintenance in both trunk and appendages. Alternatively, it may have a more general role in tissue or organ
maintenance.
q 2006 Elsevier Ireland Ltd. All rights reserved.
Keywords: Tribolium; knodel; Naked cuticle; Segmentation; Segment polarity; Maintenance; leg development
1. Introduction
The principles underlying the formation of Drosophila body
segments are well understood (Pankratz and Jackle, 1990; St
Johnston and Nusslein-Volhard, 1992). Equally important, but
less understood, is the maintenance of established segment and
organ primordia. During pattern formation the embryo is
subdivided in ever smaller units as for instance seen by the
expression domains of gap and pair rule genes. These subdivi-
sions are transient in nature and are not directly transformed
into morphological traits. Only the last level of the segmenta-
tion hierarchy, the segment polarity pattern, provides direct
cues for morphogenesis, i.e. compartment boundaries, cuticle
differentiation or muscle attachment sites (Martinez Arias et al.,
1988; Sanson, 2001; Larsen et al., 2003). Pattern maintenance,
in contrast, requires stable cues that are able to maintain borders
and regulate the size of morphological units. Only few of these
0925-4773/$ - see front matter q 2006 Elsevier Ireland Ltd. All rights reserved.
doi:10.1016/j.mod.2006.04.003
* Corresponding author. Tel.: C49 551 395426; fax: C49 551 395416.
E-mail address: [email protected] (G. Bucher).
cues are known. Mutual activation of wingless and engrailed
expression is crucial for the maintenance of parasegmental
boundaries during germ band extension. Afterwards, their
expression domains become independent from each other
(Heemskerk et al., 1991; Martinez Arias, 1993; DiNardo et
al., 1994). Engrailed expression then comes under cell autono-
mous control involving the action of polycomb group genes
(Moazed and O’Farrell, 1992). In the case of the segment
polarity gene gooseberry, different enhancer elements are
responsible for early and late functions (Li et al., 1993).
Adjustment of parasegment size involves cell death and
delamination of miss-specified cells (Hughes and Krause,
2001). In wing imaginal discs, maintenance of parasegmental
borders involves hedgehog regulated cell sorting (Dahmann and
Basler, 2000). It has been assumed that differential cell
adhesion may be involved but the molecules responsible for
the hypothetical differential cell adhesion remains elusive
despite specific screening efforts (Dahmann and Basler, 1999).
All conclusions drawn from Drosophila research bear the
drawback that this species develops through a highly derived
Mechanisms of Development 123 (2006) 430–439
www.elsevier.com/locate/modo

H. Wohlfrom et al. / Mechanisms of Development 123 (2006) 430–439 431
mode of embryogenesis (long germ embryogenesis) that is not
typical for insects (Tautz et al., 1994; Davis and Patel, 2002).
Hence, statements on general properties of insect development
have to be based on studies of insects with more typical
features. We have chosen the red flour beetle Tribolium
castaneum as model system, because it is readily manipulated
by genetic, transgenic and RNAi approaches (Maderspacher et
al., 1998; Berghammer et al., 1999; Brown et al., 1999; Bucher
et al., 2002; Klingler, 2004). As in most insects, in Tribolium
only the anteriormost segments are patterned in the blastoderm
stage while all following segments are formed one by one from
a posterior growth zone (short germ embryogenesis). In
addition, Tribolium displays a fully developed larval head,
larval appendages and extraembryonic tissue. The morpho-
logical conservation suggests that also the genetic control
reflects the ancestral state more accurately than it does in
Drosophila. Indeed, it has been shown that early patterning
in Tribolium does not involve bicoid and that the abdominal
gap gene orthologs have drastically changed their function
(Brown et al., 2001; Bucher and Klingler, 2004; Cerny et al.,
2005). In contrast, on the level of segment polarity genes
studied so far, both expression and function appear to be
conserved to great extent (Brown et al., 1994; Nagy and
Carroll, 1994; Oppenheimer et al., 1999).
In a screen for embryonic lethal mutations in Tribolium, we
have recovered knodel (kno), a mutant lacking cuticular
segmentation. We show that segmentation itself is not compro-
mised but the maintenance of the pattern is impeded. We
speculate that kno may be a late acting segment polarity
gene, or a gene required for adhesion. The phenotype of kno
mutant embryos shows that also in short germ embryos, initial
patterning of the growing germ band is followed by a distinct
maintenance phase.
2. Experimental procedures
2.1. Identification of the knodel mutant strain
kno was identified independently in two genetic backgrounds. In a screen
for embryonic lethal mutations (Maderspacher et al., 1998), the allele 13h8 was
isolated in an EMS-treated SB wild-type background. Independently, we found
that our copy of a Tiw-1 wild-type strain harbored a large number of
chromosomes carrying a kno allele: 39 of approximately 400 inter se back-
crosses segregated embryos indistinguishable from 13h8. Complementation
analysis showed that the two mutants (kno13h8 and knoTiw1) are allelic. knodel
(dumpling) is a dish typical for southern Germany and consists of amorphous
balls made of potato flour or bread that resembles the knodel phenotype.
2.2. Complementation study
One knoTiw1 male was crossed to three kno13h8 females and the offspring
checked for mutant offspring (nZ8) and vice versa (nZ10). Mutant offspring
was detected in 3/8 and 7/10 of the crosses, respectively. Therefore, the
mutations do not complement and are alleles of the same locus.
2.3. Histology
Whole-mount in situ hybridizations were performed according to estab-
lished protocols (Tautz and Pfeifle, 1989). For initial double stainings,
fluorescein- and digoxigenin-labeled probes were detected using alkaline
phosphatase and beta-galaktosidase, the latter after signal enhancement via
biotin deposition (Prpic et al., 2001). For the remaining double in situs both
stainings were detected successively by alkaline phosphatase using NBT/BCip
and the fluorescent FastRed reaction (Sigma), respectively. The false color
pictures were produced by pasting the inversed bright field image into the red
and blue channels of the image of the fluorescent staining. Detailed protocols
are available from the authors. The stained embryos were dissected free from
the yolk and embedded in 50% glycerol. Cuticles were embedded in Hoyer’s
medium mixed with lactic acid (1:1) and analyzed using standard dark field
illumination. In addition, confocal microscopy of the auto fluorescent cuticle
was used (excitation: argon laser, 488 nm; emission filter: long pass 505 nm)
2.4. Embedding in low refraction medium
In glycerol, the yolk granules disturb optical observation. Therefore,
embryos usually have to be dissected from the yolk. In order to observe
embryos within the egg, we have developed an embedding method for making
yolk transparent for normal bright field microscopy. Embryos are dehydrated in
alcohol (ethanol or methanol, 25, 50, 75, 100%). They are then embedded in a
solution of high refractory index, ‘benz-mix’, a 4.3:1 mixture of benzyl
benzoate: benzyl alcohol for embryos dehydrated in ethanol (3:1 for embryos
dehydrated in methanol). This technique was applied to the whole-mount
Tc’even-skipped staining.
2.5. Scanning electron microscopy
Embryos were dehydrated by 10 min incubations in raising acetone
concentrations (70%, 80%, 2!98%, 2!100%). They were then immediately
transferred to HMDS (1,1,1,3,3,3 hexadimethyldisilazane; Merck-Suchardt,
Darmstadt) and incubated for 30 min. HMDS was then removed and embryos
transferred to an exsiccator loaded with silica gel balls (Merck-Suchardt,
Darmstadt). The exsiccator was evacuated in order to avoid contamination
with water (which causes shrinkage), and evaporation was performed over
night. Embryos were then transferred to an aluminium carrier (Plano, Wetzlar)
that had been prepared with a double-sided adherent disc (Plano, Wetzlar). The
embryo was then sputtered with gold (BIO-RAD SC 510, Munchen) at 200 V
for 120 s. Scans were performed with a Philips XS-20 scanning electron
microscope at 20 kV.
3. Results
3.1. knodel embryos lack segment boundaries and bristles
kno mutant larvae differentiate as nearly amorphous cuticu-
lar sacs that lack most bristles, proper appendages and any
sign of abdominal segmentation (Fig. 1A–C). The cuticle of
kno embryos appears to be shaped by the egg shell because
the cuticle closely lines the vitelline membrane. Accordingly,
the proportions of kno embryos differ strongly from wild-type:
they are about half as long, but approximately 30% more wide
than wild-type 1st instar (L1) larvae. The anterior posterior
axis is well established, however, as in all larvae the anterior
pole can be recognized. In the head, several rudimentary
appendages are usually formed, although it is often difficult
to assign their identity unequivocally. The labrum is
commonly discernable as are shortened antennae that lack
the flagellum. Mandibles, maxillae and labium are strongly
reduced to cuticular humps that sometimes carry distal
structures like reduced palps or sclerotized tips (mandible).
In most cases, remnants of most but not all gnathal segments
can be found. Legs are either missing or discernable as

Fig. 1. Cuticle phenotype of kno mutant embryos. White lines indicate the ventral midline, (B 0) and (C 0) are red–green representations of the embryos in (B) and (C),
respectively. (A) Wild-type larva displaying head, thorax with three pairs of legs, and eight discernible abdominal segments. (B) In weak kno phenotypes, the labrum
(open arrowhead), antenna (open arrow) and maxilla (star) are clearly visible. The labium is not present in this case, but in other specimens, the labium is present
while other gnathal structures may be missing. The only indication of trunk segmentation is provided by the remnants of thoracic legs (white arrowheads). Arrows
point to four nests of cuticular structures (those marked with D and E are shown in the respective panels). (C) Even in strong phenotypes, the labrum is usually visible
(open arrowhead) and indicates the anterior end of the embryo. Only few remnants of gnathal and thoracic appendages can be distinguished (open arrow: antenna,
white arrowheads: thorax). The lengths of wild-type and kno cuticles are to scale. The width of the cuticles cannot be compared, however, because only the wild-type
has been flattened in order to display all structures in one plane. Actually, kno cuticles are approximately 33% wider than cuticles of wild-type siblings. (D)–(E 0)
Close-up of ‘nests’ of cuticular structures of the cuticle shown in B. These structures presumably derive from malformed and fused bracts normally associated with
dorsal bristles. (D) This type of cuticular structure resembles an elevation with several sclerotized peaks pointing outwards. (E), (E 0) Two focal planes of another type
of bract nest. This structure is sunk below the surrounding cuticle and the blunt bracts point towards the center. An identical structure has been described for the
Tribolium cobbled mutant.
H. Wohlfrom et al. / Mechanisms of Development 123 (2006) 430–439432
cuticular bulges. Segmentation of gnathum and thorax is only
detectable from the reduced appendages since no segmental
folds are evident. Correspondingly, in the abdomen no signs
of segmentation whatsoever can be found. The posterior
terminal urogomphi are visible in a portion of kno mutant
cuticles, while the pygopods are never detected. Most differ-
entiated kno embryos lack spiracles—only in few cases a
single spiracle was found. Sometimes up to seven irregularly
distributed bristles are present, some of which split at the
base. In some embryos, however, one or two types of unusual
‘nests’ of cuticular structures can be found (Fig. 1D and E).
One type of nests has a distinct roundish circumference and
contains several bract-like structures that point to the center of
the nest (Fig. 1E). The other type consists of sclerotized tips
pointing upwards (Fig. 1D). These structures lack a clear
border and appear situated on cuticular elevations. In some
embryos we find 3–5 such nests while they are absent in
others. We have not found any regularity regarding their
position apart from the fact that they occur in the abdomen
only.

H. Wohlfrom et al. / Mechanisms of Development 123 (2006) 430–439 433
In order to test for penetrance of the mutation, we crossed a
single F1 male (probability for being a mutation carrier: 50%)
with three F1 females (cumulative probability that at least one
of them is a mutation carrier: 87.5%). Mutant phenotypes could
be detected in 21 of 48 such crosses (43.8%), which coin-
cidently, is exactly the expected proportion (50% * 87.5%Z43.8%). In order to test for heterozygous effects of the
mutation, we crossed mutation carrier males with wild-type
females and tested the offspring for cuticle defects. As no
defects could be found we conclude that the mutation is strictly
recessive.
In summary, the gene affected in kno is recessive lethal and
affects segmentation. The presence of rudimentary head and
thorax appendages indicates that segmentation is not totally
impeded and that some proximal-distal axis formation is
performed. In addition, the mutation also interferes with
bristle development. The kno gene may therefore also be
required for peripheral sensory organ development.
3.2. During germ band growth, even-skipped expression
is normal
To determine the moment when segmentation is disturbed in
kno mutant embryos, we stained the offspring of heterozygous
parents for the product of the pair-rule gene Tc’even-skipped
(Tc’eve). Like in Drosophila, this gene is initially expressed in
every other segment but resolves into a segmental pattern later
on. We did not detect any disturbance of the pair-rule pattern or
the subsequent segmental Tc’eve pattern in kno mutant
embryos during segmentation (not shown). We also find no
overt changes in the morphology of elongating embryos.
Together, these data indicate that growth and the segmentation
process are not affected in kno embryos.
After segmentation is completed, Tc’eve becomes expressed
in a segmentally reiterated pattern. Ventrally, Tc’eve
Fig. 2. Expression of Tc’eve wild-type (A,B) and kno embryos (C,D). The embryo
views with the anterior towards left (posterior parts are out of focus). (A,B) Tc’eve
clusters most likely mark mesodermal cells that contribute to the larval heart and dor
(A). (C) A kno mutant embryo at approximately the same age as that in (A). Staining
the latter comprise more cells and come into close proximity. (D) In older mutant
number of expressing cells. Some clusters have fused reducing the overall number o
(see text). Note that we do neither find changes in Tc’eve expression in younger st
expression presumably marks a set of neural precursor cells
(Fig. 2A and B). This pattern is conserved within insects and
crustaceans (Duman-Scheel and Patel, 1999). In kno mutant
embryos, also the neural Tc’eve expression initiates normally
but becomes increasingly disorganized during retraction stages
(Fig. 2C and D). Eventually, the pattern is strongly disturbed in
embryos that have finished retraction. In Drosophila, eve-
expressing cell clusters arise segmentally also in the dorsal
mesoderm and contribute to the larval heart and dorsal somatic
muscles. Among the regulators necessary for proper activation
of these cell clusters are several segmentation genes: wingless,
hedgehog and sloppy paired (Lawrence et al., 1995; Riech-
mann et al., 1997; Liu et al., 2006). In Tribolium, we find
Tc’eve positive cell clusters that resemble those in Drosophila
with respect to the onset of expression, their number and dorsal
position (Fig. 2A, black arrowhead). We therefore assume that
these clusters are heart and dorsal somatic muscle precursors.
In kno mutant embryos, the staining in these dorsal cell clusters
becomes more prominent than in wild-type (Fig. 2C, black
arrowhead). This is partly due to the greater number of cells in
each cluster but might in addition reflect Tc’eve up-regulation.
In addition, some clusters appear to fuse with their segmental
neighbors. Accordingly, the number of clusters is reduced in
kno mutant embryos as compared to wild-type. In the stages
examined, the number is not reduced to half as would be
expected for a pair rule phenotype. Intriguingly, this pattern is
almost identical to what happens in Drosophila when wingless
is evenly expressed in embryos (Lawrence et al., 1995). This
raises the possibility that knodel may be a component of the
wingless signaling pathway.
Together, the Tc’eve staining suggests that segmentation
and early neurogenesis are not affected while development
becomes disturbed during retraction. This indicates that kno
probably is involved in maintenance of segment primordia
rather than their establishment.
s are embedded in low refraction medium without removing the yolk. Ventral
is expressed in a specific and segmentally repeated set of neurons. Dorsal cell
sal somatic muscles (black arrowhead). The embryo in B is older than the one in
in ventral cell clusters fades while it intensifies in the dorsal clusters. In addition,
embryos, the lateral clusters have further gained in strength of expression and
f clusters. This phenotype is similar to wingless over expression in Drosophila
ages undergoing segmentation nor hints for a pair rule defect.

H. Wohlfrom et al. / Mechanisms of Development 123 (2006) 430–439434
3.3. Segment polarity gene patterns degrades with
developmental time
In order to get more insight into the nature of the defects, we
analyzed segmental markers in kno mutant embryos. The
boundary between the expression domains of the segment
polarity genes engrailed and wingless defines the paraseg-
mental boundaries of all arthropods investigated so far
(Damen, 2002). We investigated the patterns of Tc’wingless
(Tc’wg) and Tc’engrailed (Tc’en) and did not find any
alteration in early stages up to the extended germ band stage
(not shown). Thereafter, the pattern begins to degrade
(Fig. 3D–I, M–Q). The effect of the kno mutation on the
Tc’wg pattern is particularly severe. In wild-type embryos,
Tc’wg staining evolves into a complex but regular pattern of
Tc’wg positive cells (Nagy and Carroll, 1994). In kno mutant
embryos this pattern is initially modified by under- and hyper-
regulation of certain expression spots (Fig. 3E and F compare
medial and lateral spots (red and blue arrowheads, respect-
ively) with wild-type in B,C). Later, the wild-type pattern is
lost almost completely and only a few irregularly distributed
Tc’wg positive cell clusters are maintained (Fig. 3G and I).
Similar to the Tc’eve staining, the Tc’wg expression in kno
mutant embryos becomes stronger in the lateral parts than in
median parts of the germ band. These lateral parts of the germ
band represent future dorsal tissues. Apparently, the require-
ment for kno is not equally distributed along the dorsal-ventral
axes. The changes in the Tc’en patterns are less severe
(Fig. 3M–Q, compare to wild-type J–L). In retracting
embryos, the pattern begins to fade along the ventral midline
while the stripes remain almost normal laterally (Fig. 3M and
M 0, compare to J). In older embryos, the stripes become
discontinuous and irregular laterally while the ventral tissue
remains free of Tc’en staining (Fig. 3N,O,Q, compare to K,L).
These data demonstrate that the process of segment boundary
formation is intact in kno mutant embryos but that the
maintenance of the segmental pattern is impeded.
3.4. Appendage primordia are initiated but shrink subsequently
We wondered whether the reduction of appendages is a
secondary consequence of the segment degeneration pheno-
type or whether appendages are independently affected by lack
of kno function. We examined wild-type and mutant embryos
by scanning electron microscopy (SEM) (Fig. 4). We examined
stages where segmentation is completed and appendages are
growing out (32–48 h of development at 32 8C). We find that at
these stages all appendages are present in mutants but are
shorter than in wild-type embryos at a similar stage. However,
these early appendage defects are much less severe than those
in 1st instar larvae (Fig. 1B and C). To confirm our impression
that the initiation of leg patterning is normal in kno mutants, we
stained for Tc’Distal-less (Tc’Dll) as distal marker, and
Tc’dachshund (Tc’dac) as marker for intermediate proximo-
distal positions (Beermann et al., 2001; Prpic et al., 2001).
Indeed, prior to germband retraction stages, no pronounced
difference from the wild-type expression patterns could be
observed (not shown). Even in advanced embryonic stages,
where body segmentation of mutant embryos is already visibly
disturbed, the expression of these leg genes is still strikingly
normal (Fig. 5). The almost complete deletion of leg structures
in mature embryos therefore must involve secondary degener-
ation of leg primordia, which developed quite normally during
earlier embryo stages. Similarly as in anterior–posterior
patterning, this suggests that kno mutant embryos have a
problem in maintaining proximo-distal structures rather than
in establishing them.
4. Discussion
First instar cuticles of larvae mutant for kno lack overt
abdominal segmentation and have severely reduced appen-
dages. In addition, they lack almost all bristles and display
disorganized patterns of neural precursor cells. It should be
pointed out that segmentation and appendage defects are
usually not linked in Tribolium. Segmentation mutants like
itchy, scratchy, krusty, kraken and others affect segmentation
but not proximo-distal patterning of the appendages (Sulston
and Anderson, 1996; Maderspacher et al., 1998). In godzilla
and bollig, abnormally patterned legs are observed, however,
these appendage phenotypes appear to be secondary effects of
early trunk segmentation defects. Our results indicate that the
kno phenotypes result from degeneration of well initiated
segmental and appendage primordia.
4.1. knodel shares phenotypic traits with the Tribolium
cobbled mutation
A similar combination of sac like appearance and nests of
bracts has previously been described for the mutation cobbled
(Sulston and Anderson, 1996). In contrast to kno, however, the
appendages of cobbled embryos are much better developed,
and all body segment boundaries can still be identified. Also
cobbled mutant embryos lack bristles. In contrast to kno, the
nests of bracts occur in almost all abdominal segments at the
approximate dorso-ventral position of wild-type bristle-bracts.
The combination of defects suggests that cobbled could be a
weak allele at the kno locus. Alternatively, cobbled could be a
gene involved in the same pathway as kno. Unfortunately, the
cobbled mutation has been lost, i.e. we were unable to test kno
and cobbled for complementation.
4.2. Is knodel involved in cell adhesion?
Degeneration of tissues is a hallmark of mutations that affect
epithelial cell polarity and adhesion. Drosophila embryos with
reduced armadillo/beta-catenin function for instance form a
normal blastodermal epithelium but subsequent morphogenetic
movements like germ band extension do not occur properly or
not at all. The epithelium loses its apical basal polarity and
subsequently degenerates into a disordered multilayered
mesenchyme. A contiguous cuticle is not secreted in such
degenerating embryos leading to only some residual cuticle
scrabs (Amin et al., 1999). Such ‘crumbs-like’ phenotypes are
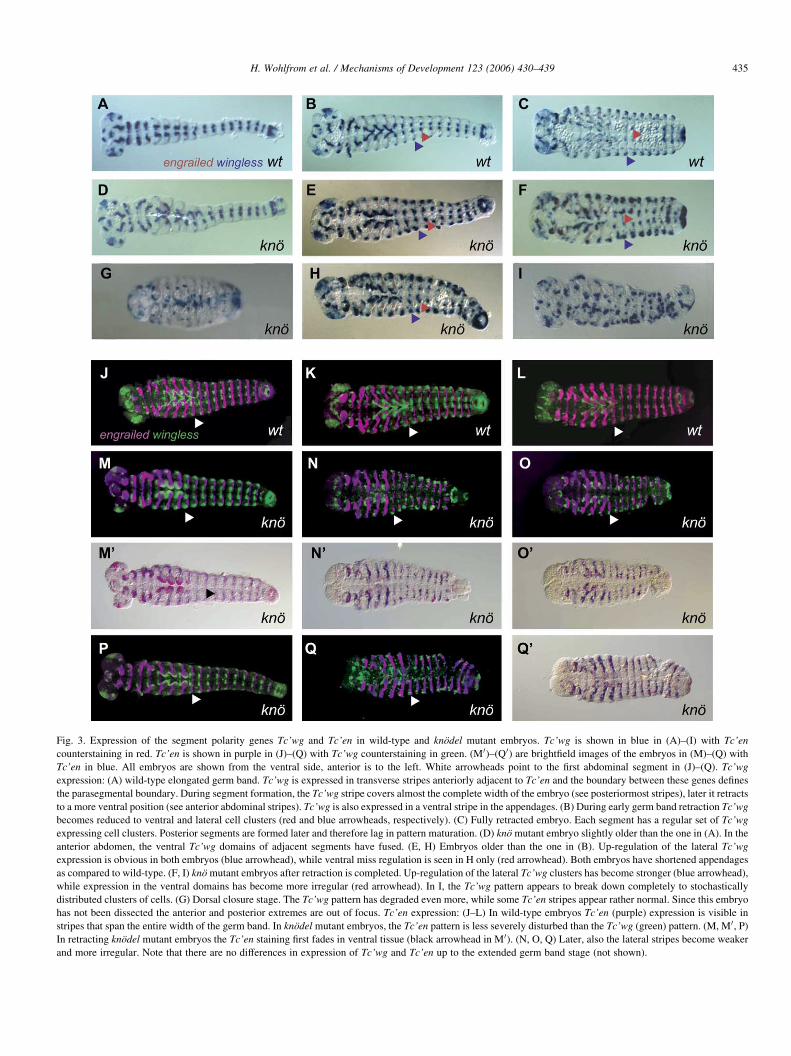
Fig. 3. Expression of the segment polarity genes Tc’wg and Tc’en in wild-type and knodel mutant embryos. Tc’wg is shown in blue in (A)–(I) with Tc’en
counterstaining in red. Tc’en is shown in purple in (J)–(Q) with Tc’wg counterstaining in green. (M 0)–(Q 0) are brightfield images of the embryos in (M)–(Q) with
Tc’en in blue. All embryos are shown from the ventral side, anterior is to the left. White arrowheads point to the first abdominal segment in (J)–(Q). Tc’wg
expression: (A) wild-type elongated germ band. Tc’wg is expressed in transverse stripes anteriorly adjacent to Tc’en and the boundary between these genes defines
the parasegmental boundary. During segment formation, the Tc’wg stripe covers almost the complete width of the embryo (see posteriormost stripes), later it retracts
to a more ventral position (see anterior abdominal stripes). Tc’wg is also expressed in a ventral stripe in the appendages. (B) During early germ band retraction Tc’wg
becomes reduced to ventral and lateral cell clusters (red and blue arrowheads, respectively). (C) Fully retracted embryo. Each segment has a regular set of Tc’wg
expressing cell clusters. Posterior segments are formed later and therefore lag in pattern maturation. (D) kno mutant embryo slightly older than the one in (A). In the
anterior abdomen, the ventral Tc’wg domains of adjacent segments have fused. (E, H) Embryos older than the one in (B). Up-regulation of the lateral Tc’wg
expression is obvious in both embryos (blue arrowhead), while ventral miss regulation is seen in H only (red arrowhead). Both embryos have shortened appendages
as compared to wild-type. (F, I) kno mutant embryos after retraction is completed. Up-regulation of the lateral Tc’wg clusters has become stronger (blue arrowhead),
while expression in the ventral domains has become more irregular (red arrowhead). In I, the Tc’wg pattern appears to break down completely to stochastically
distributed clusters of cells. (G) Dorsal closure stage. The Tc’wg pattern has degraded even more, while some Tc’en stripes appear rather normal. Since this embryo
has not been dissected the anterior and posterior extremes are out of focus. Tc’en expression: (J–L) In wild-type embryos Tc’en (purple) expression is visible in
stripes that span the entire width of the germ band. In knodel mutant embryos, the Tc’en pattern is less severely disturbed than the Tc’wg (green) pattern. (M, M 0, P)
In retracting knodel mutant embryos the Tc’en staining first fades in ventral tissue (black arrowhead in M 0). (N, O, Q) Later, also the lateral stripes become weaker
and more irregular. Note that there are no differences in expression of Tc’wg and Tc’en up to the extended germ band stage (not shown).
H. Wohlfrom et al. / Mechanisms of Development 123 (2006) 430–439 435
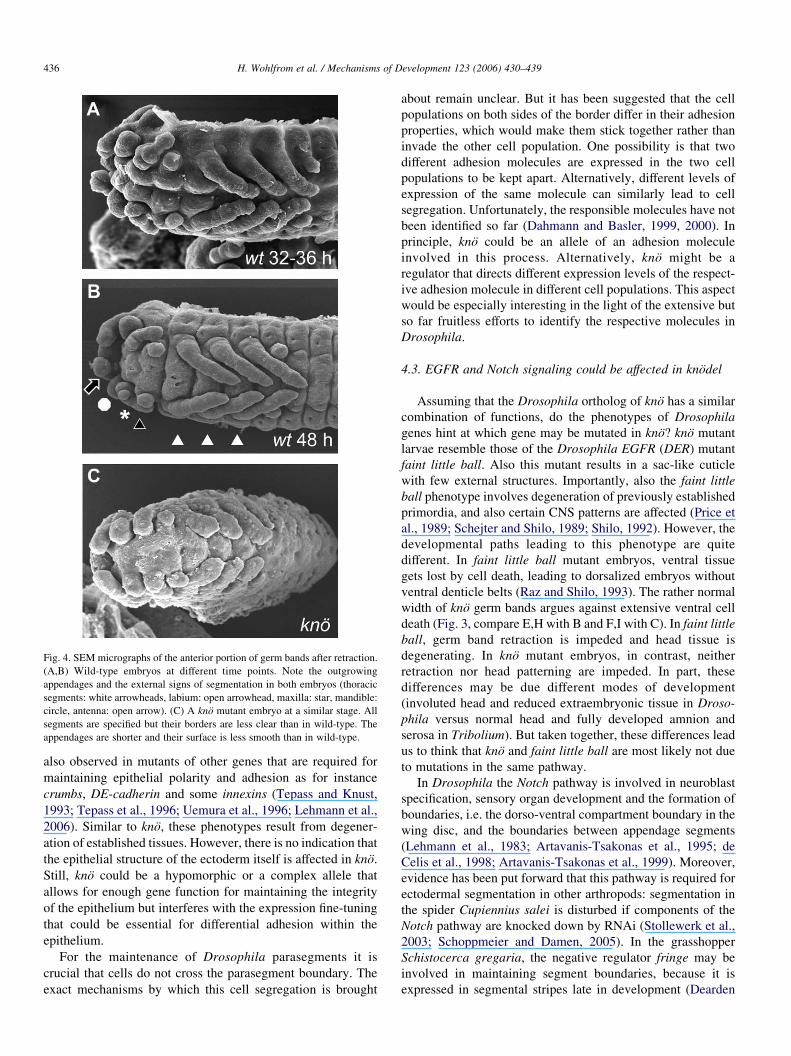
Fig. 4. SEM micrographs of the anterior portion of germ bands after retraction.
(A,B) Wild-type embryos at different time points. Note the outgrowing
appendages and the external signs of segmentation in both embryos (thoracic
segments: white arrowheads, labium: open arrowhead, maxilla: star, mandible:
circle, antenna: open arrow). (C) A kno mutant embryo at a similar stage. All
segments are specified but their borders are less clear than in wild-type. The
appendages are shorter and their surface is less smooth than in wild-type.
H. Wohlfrom et al. / Mechanisms of Development 123 (2006) 430–439436
also observed in mutants of other genes that are required for
maintaining epithelial polarity and adhesion as for instance
crumbs, DE-cadherin and some innexins (Tepass and Knust,
1993; Tepass et al., 1996; Uemura et al., 1996; Lehmann et al.,
2006). Similar to kno, these phenotypes result from degener-
ation of established tissues. However, there is no indication that
the epithelial structure of the ectoderm itself is affected in kno.
Still, kno could be a hypomorphic or a complex allele that
allows for enough gene function for maintaining the integrity
of the epithelium but interferes with the expression fine-tuning
that could be essential for differential adhesion within the
epithelium.
For the maintenance of Drosophila parasegments it is
crucial that cells do not cross the parasegment boundary. The
exact mechanisms by which this cell segregation is brought
about remain unclear. But it has been suggested that the cell
populations on both sides of the border differ in their adhesion
properties, which would make them stick together rather than
invade the other cell population. One possibility is that two
different adhesion molecules are expressed in the two cell
populations to be kept apart. Alternatively, different levels of
expression of the same molecule can similarly lead to cell
segregation. Unfortunately, the responsible molecules have not
been identified so far (Dahmann and Basler, 1999, 2000). In
principle, kno could be an allele of an adhesion molecule
involved in this process. Alternatively, kno might be a
regulator that directs different expression levels of the respect-
ive adhesion molecule in different cell populations. This aspect
would be especially interesting in the light of the extensive but
so far fruitless efforts to identify the respective molecules in
Drosophila.
4.3. EGFR and Notch signaling could be affected in knodel
Assuming that the Drosophila ortholog of kno has a similar
combination of functions, do the phenotypes of Drosophila
genes hint at which gene may be mutated in kno? kno mutant
larvae resemble those of the Drosophila EGFR (DER) mutant
faint little ball. Also this mutant results in a sac-like cuticle
with few external structures. Importantly, also the faint little
ball phenotype involves degeneration of previously established
primordia, and also certain CNS patterns are affected (Price et
al., 1989; Schejter and Shilo, 1989; Shilo, 1992). However, the
developmental paths leading to this phenotype are quite
different. In faint little ball mutant embryos, ventral tissue
gets lost by cell death, leading to dorsalized embryos without
ventral denticle belts (Raz and Shilo, 1993). The rather normal
width of kno germ bands argues against extensive ventral cell
death (Fig. 3, compare E,H with B and F,I with C). In faint little
ball, germ band retraction is impeded and head tissue is
degenerating. In kno mutant embryos, in contrast, neither
retraction nor head patterning are impeded. In part, these
differences may be due different modes of development
(involuted head and reduced extraembryonic tissue in Droso-
phila versus normal head and fully developed amnion and
serosa in Tribolium). But taken together, these differences lead
us to think that kno and faint little ball are most likely not due
to mutations in the same pathway.
In Drosophila the Notch pathway is involved in neuroblast
specification, sensory organ development and the formation of
boundaries, i.e. the dorso-ventral compartment boundary in the
wing disc, and the boundaries between appendage segments
(Lehmann et al., 1983; Artavanis-Tsakonas et al., 1995; de
Celis et al., 1998; Artavanis-Tsakonas et al., 1999). Moreover,
evidence has been put forward that this pathway is required for
ectodermal segmentation in other arthropods: segmentation in
the spider Cupiennius salei is disturbed if components of the
Notch pathway are knocked down by RNAi (Stollewerk et al.,
2003; Schoppmeier and Damen, 2005). In the grasshopper
Schistocerca gregaria, the negative regulator fringe may be
involved in maintaining segment boundaries, because it is
expressed in segmental stripes late in development (Dearden

Fig. 5. Expression of the appendage genes Tc’Distal-less (Tc’Dll) and Tc’dachshund (Tc’dac) suggest that proximo-distal pattern formation is not impeded in kno
mutant embryos. (A, B) In wild-type, Tc’Dll is expressed in the distal portion of every appendage except the mandibles. At later stages a ring appears in a more
proximal position. (D, E) In kno mutant embryos, leg morphology and Tc’Dll expression are not severely altered, even though the trunk segment pattern already has
degenerated. (C,F) Tc’dac is expressed in medial portions of the leg in both wild-type (C) and kno mutant embryos (F).
H. Wohlfrom et al. / Mechanisms of Development 123 (2006) 430–439 437
and Akam, 2000). In Drosophila, Notch is not involved in
segmentation of the ectoderm but plays a role in the segmental
patterning of the mesoderm (Tapanes-Castillo and Baylies,
2004). It is conceivable that Notch may have a role in body
segment boundary formation in Tribolium, which has been lost
in Drosophila. An involvement of kno in Notch signaling could
also explain leg phenotypes because this pathway is required
for establishing the joints of appendages in Drosophila
imaginal discs. However, the degeneration of the entire
appendages in kno argues for a function that is not restricted
to podomer boundaries. The nests of bracts found in kno could
represent a weak neurogenic phenotype. To explain the strong
late defects (i.e. PNS, segmentation and limb development) in
combination with weak early (neurogenesis) phenotypes one
might have to consider maternal contribution of the gene
product. This could specifically rescue early embryonic
requirements (i.e. neurogenesis in the CNS). This scenario is
unlikely, however, since cell proliferation in the growth zone of
short germ embryos should rapidly dilute maternal mRNAs
such that at least neurogenesis in the posterior abdomen likely
has to rely on zygotic functions.
4.4. knodel could be involved in wingless signaling
We prefer the hypothesis that knodel is a segment polarity
gene, probably involved in the wingless pathway. Indeed,
degeneration of a well-established segment pattern is a hall-
mark of segment-polarity phenotypes in Drosophila. For
example, the phenotype of the wingless antagonist naked
cuticle (nkd) (Zeng et al., 2000) resembles that of kno.
Drosophila nkd mutant larvae are much shorter than wild-
type, show no overt sign of segmentation and are devoid of
denticles. nkd is also involved in neurogenesis (Deshpande
et al., 2001). Similar to kno mutant embryos, engrailed and
wingless stripes are initiated normally in nkd mutant embryos,
but the pattern degrades after the elongated germ band stage is
reached, leading to ectopic wingless stripes. This is reminiscent
of the miss regulation of wingless in kno mutants. However, the
response of engrailed to the Drosophila nkd mutation differs
from our findings in Tribolium in that the engrailed stripes
become broader, which we have not observed in kno mutant
embryos. However, the kno mutants may not represent the
complete knockout of the gene, or the regulatory interactions in
Tribolium may be subtly different from Drosophila. Further
support for kno being involved in wingless signaling comes
from the effects of kno on the dorsal mesoderm Tc’eve positive
cell clusters: They become enlarged and partially fuse with
their neighbors (Fig. 2C and D). Exactly the same is observed
when wingless is experimentally evenly distributed in Droso-
phila embryos (Lawrence et al., 1995).
If kno represents a segment polarity gene, its appendage
defects would be a secondary consequence of the degeneration
of segmental patterns because the parasegment boundary runs
through the appendage. Alternatively, a segment polarity gene
could be mutated in kno that plays direct roles in both body
segmentation and leg patterning, as is the case with odd-
skipped in Drosophila (Hao et al., 2003). Mapping of the
mutant and comparison with the Tribolium genome sequence

H. Wohlfrom et al. / Mechanisms of Development 123 (2006) 430–439438
should clarify if this locus represents the homolog of a gene
that already has been studied in Drosophila. Alternatively, this
locus may represent a new gene only required in Tribolium, or
whose function has escaped attention in Drosophila.
kno is clearly a gene not required for the formation of
segment boundaries but for maintaining their integrity. This
shows that pattern formation and maintenance rely at least in
part on the action of different genes and/or on different
regulation of the same genes. A switch in regulation has for
instance been described for engrailed expression. Initially it is
wingless-dependent (Martinez Arias et al., 1988; Heemskerk
et al., 1991) while later maintenance of the striped pattern is
wingless independent but requires polycomb group genes
(DiNardo et al., 1988; Moazed and O’Farrell, 1992). In the
case of gooseberry, such different modes of regulation could be
assigned to separate ‘early’ and ‘late’ enhancer elements
(Li et al., 1993). By classical forward genetics, the mainten-
ance process is difficult to study because many genes are
involved also in preceding pattern formation. Their late
function is thus obscured by the early one. The availability of
the genomic sequence and the ease of reverse genetics (Brown
et al., 1999; Bucher et al., 2002; Klingler, 2004) make
Tribolium an ideal model system for studying the genes and
processes necessary for late development that are required for
maintenance of once established patterns.
References
Amin, A., Li, Y., Finkelstein, R., 1999. Hedgehog activates the EGF receptor
pathway during Drosophila head development. Development 126, 2623–
2630.
Artavanis-Tsakonas, S., Matsuno, K., Fortini, M.E., 1995. Notch signaling.
Science 268, 225–232.
Artavanis-Tsakonas, S., Rand, M.D., Lake, R.J., 1999. Notch signaling: cell
fate control and signal integration in development. Science 284, 770–776.
Beermann, A., Jay, D.G., Beeman, R.W., Hulskamp, M., Tautz, D., Jurgens, G.,
2001. The Short antennae gene of Tribolium is required for limb develop-
ment and encodes the orthologue of the Drosophila Distal-less protein.
Development 128, 287–297.
Berghammer, A.J., Klingler, M., Wimmer, E.A., 1999. A universal marker for
transgenic insects. Nature 402, 370–371.
Brown, S.J., Patel, N.H., Denell, R.E., 1994. Embryonic expression of the
single Tribolium engrailed homolog. Dev. Genet. 15, 7–18.
Brown, S.J., Mahaffey, J.P., Lorenzen, M.D., Denell, R.E., Mahaffey, J.W.,
1999. Using RNAi to investigate orthologous homeotic gene function
during development of distantly related insects. Evol. Dev. 1, 11–15.
Brown, S., Fellers, J., Shippy, T., Denell, R., Stauber, M., Schmidt-Ott, U.,
2001. A strategy for mapping bicoid on the phylogenetic tree. Curr. Biol.
11, R43–R44.
Bucher, G., Klingler, M., 2004. Divergent segmentation mechanism in the short
germ insect Tribolium revealed by giant expression and function. Develop-
ment 131, 1729–1740.
Bucher, G., Scholten, J., Klingler, M., 2002. Parental RNAi in Tribolium
(Coleoptera). Curr. Biol. 12, R85–R86.
Cerny, A.C., Bucher, G., Schroder, R., Klingler, M., 2005. Breakdown of
abdominal patterning in the Tribolium Kruppel mutant jaws. Development
132, 5353–63.
Dahmann, C., Basler, K., 1999. Compartment boundaries: at the edge of
development. Trends Genet. 15, 320–326.
Dahmann, C., Basler, K., 2000. Opposing transcriptional outputs of Hedgehog
signaling and engrailed control compartmental cell sorting at the Droso-
phila A/P boundary. Cell 100, 411–422.
Damen, W.G., 2002. Parasegmental organization of the spider embryo implies
that the parasegment is an evolutionary conserved entity in arthropod
embryogenesis. Development 129, 1239–1250.
Davis, G.K., Patel, N.H., 2002. Short, long, and beyond: molecular and
embryological approaches to insect segmentation. Annu. Rev. Entomol.
47, 669–699.
Dearden, P., Akam, M., 2000. A role for Fringe in segment morphogenesis but
not segment formation in the grasshopper, Schistocerca gregaria. Dev.
Genes Evol. 210, 329–336.
de Celis, J.F., Tyler, D.M., de Celis, J., Bray, S.J., 1998. Notch signalling
mediates segmentation of the Drosophila leg. Development 125, 4617–4626.
Deshpande, N., Dittrich, R., Technau, G.M., Urban, J., 2001. Successive
specification of Drosophila neuroblasts NB 6-4 and NB 7-3 depends on
interaction of the segment polarity genes wingless, gooseberry and naked
cuticle. Development 128, 3253–3261.
DiNardo, S., Sher, E., Heemskerk-Jongens, J., Kassis, J.A., O’Farrell, P.H.,
1988. Two-tiered regulation of spatially patterned engrailed gene
expression during Drosophila embryogenesis. Nature 332, 604–609.
DiNardo, S., Heemskerk, J., Dougan, S., O’Farrell, P.H., 1994. The making of a
maggot: patterning the Drosophila embryonic epidermis. Curr. Opin.
Genet. Dev. 4, 529–534.
Duman-Scheel, M., Patel, N.H., 1999. Analysis of molecular marker expression
reveals neuronal homology in distantly related arthropods. Development
126, 2327–2334.
Hao, I., Green, R.B., Dunaevsky, O., Lengyel, J.A., Rauskolb, C., 2003. The
odd-skipped family of zinc finger genes promotes Drosophila leg segmen-
tation. Dev. Biol. 263, 282–295.
Heemskerk, J., DiNardo, S., Kostriken, R., O’Farrell, P.H., 1991. Multiple
modes of engrailed regulation in the progression towards cell fate
determination. Nature 352, 404–410.
Hughes, S.C., Krause, H.M., 2001. Establishment and maintenance of para-
segmental compartments. Development 128, 1109–1118.
Klingler, M., 2004. Tribolium. Curr. Biol. 14, R639–R640.
Larsen, C.W., Hirst, E., Alexandre, C., Vincent, J.P., 2003. Segment boundary
formation in Drosophila embryos. Development 130, 5625–5635.
Lawrence, P.A., Bodmer, R., Vincent, J.P., 1995. Segmental patterning of heart
precursors in Drosophila. Development 121, 4303–4308.
Lehmann, R., Jimenez, F., Dietrich, U., Campos-Ortega, J.A., 1983. On the
phenotype and development of mutants of early neurogenesis in Drosophila
melanogaster. Wilhelm Roux Arch. Dev. Biol. 192, 62–74.
Lehmann, C., Lechner, H., Loer, B., Knieps, M., Herrmann, S., Famulok, M.,
et al., 2006. Heteromerization of innexin gap junction proteins regulates
epithelial tissue organization in Drosophila. Mol. Biol. Cell.
Li, X., Gutjahr, T., Noll, M., 1993. Separable regulatory elements mediate the
establishment and maintenance of cell states by the Drosophila segment-
polarity gene gooseberry. Eur. Mol. Biol. Org. J. 12, 1427–1436.
Liu, J., Qian, L., Wessells, R.J., Bidet, Y., Jagla, K., Bodmer, R., 2006.
Hedgehog and RAS pathways cooperate in the anterior–posterior specifi-
cation and positioning of cardiac progenitor cells. Dev. Biol. 290, 373–385.
Maderspacher, F., Bucher, G., Klingler, M., 1998. Pair-rule and gap gene
mutants in the flour beetle Tribolium castaneum. Dev. Genes Evol. 208,
558–568.
Martinez Arias, A., 1993. The development of Drosophila melanogaster. In:
Bate, M., Martinez Arias, A. (Eds.), Development and patterning of the
larval epidermis of Drosophila, vol. 1. Cold Spring Harbour Press, Cold
Spring Harbour, NY, pp. 517–608.
Martinez Arias, A., Baker, N.E., Ingham, P.W., 1988. Role of segment polarity
genes in the definition and maintenance of cell states in the Drosophila
embryo. Development 103, 157–170.
Moazed, D., O’Farrell, P.H., 1992. Maintenance of the engrailed expression
pattern by Polycomb group genes in Drosophila. Development 116, 805–
810.
Nagy, L.M., Carroll, S., 1994. Conservation of wingless patterning functions in
the short-germ embryos of Tribolium castaneum. Nature 367, 460–463.
Oppenheimer, D.I., MacNicol, A.M., Patel, N.H., 1999. Functional conserva-
tion of the wingless-engrailed interaction as shown by a widely applicable
baculovirus misexpression system. Curr. Biol. 9, 1288–1296.

H. Wohlfrom et al. / Mechanisms of Development 123 (2006) 430–439 439
Pankratz, M.J., Jackle, H., 1990. Making stripes in the Drosophila embryo.
Trends Genet. 6, 287–292.
Price, J.V., Clifford, R.J., Schupbach, T., 1989. The maternal ventralizing locus
torpedo is allelic to faint little ball, an embryonic lethal, and encodes the
Drosophila EGF receptor homolog. Cell 56, 1085–1092.
Prpic, N.M., Wigand, B., Damen, W.G., Klingler, M., 2001. Expression of
dachshund in wild-type and Distal-less mutant Tribolium corroborates
serial homologies in insect appendages. Dev. Genes Evol. 211, 467–477.
Raz, E., Shilo, B.Z., 1993. Establishment of ventral cell fates in the Drosophila
embryonic ectoderm requires DER, the EGF receptor homolog. Genes Dev.
7, 1937–1948.
Riechmann, V., Irion, U., Wilson, R., Grosskortenhaus, R., Leptin, M., 1997.
Control of cell fates and segmentation in the Drosophila mesoderm.
Development 124, 2915–2922.
Sanson, B., 2001. Generating patterns from fields of cells. Examples from
Drosophila segmentation. Eur. Mol. Biol. Org. Rep. 2, 1083–1088.
Schejter, E.D., Shilo, B.Z., 1989. The Drosophila EGF receptor homolog
(DER) gene is allelic to faint little ball, a locus essential for embryonic
development. Cell 56, 1093–1104.
Schoppmeier, M., Damen, W.G., 2005. Suppressor of Hairless and Presenilin
phenotypes imply involvement of canonical Notch-signalling in segmenta-
tion of the spider Cupiennius salei. Dev. Biol. 280, 211–224.
Shilo, B.Z., 1992. Roles of receptor tyrosine kinases in Drosophila develop-
ment. Fed. Am. Soc. Exp. Biol. J. 6, 2915–2922.
St Johnston, D., Nusslein-Volhard, C., 1992. The origin of pattern and polarity
in the Drosophila embryo. Cell 68, 201–219.
Stollewerk, A., Schoppmeier, M., Damen, W.G., 2003. Involvement of Notch
and Delta genes in spider segmentation. Nature 423, 863–865.
Sulston, I.A., Anderson, K.V., 1996. Embryonic patterning mutants of
Tribolium castaneum. Development 122, 805–814.
Tapanes-Castillo, A., Baylies, M.K., 2004. Notch signaling patterns Droso-
phila mesodermal segments by regulating the bHLH transcription factor
twist. Development 131, 2359–2372.
Tautz, D., Pfeifle, C., 1989. A non-radioactive in situ hybridization method for
the localization of specific RNAs in Drosophila embryos reveals trans-
lational control of the segmentation gene hunchback. Chromosoma 98,
81–85.
Tautz, D., Friedrich, M., Schroder, R., 1994. Insect embryogenesis—what is
ancestral and what is derived? Dev Suppl, 193–199.
Tepass, U., Knust, E., 1993. Crumbs and stardust act in a genetic pathway that
controls the organization of epithelia in Drosophila melanogaster. Dev.
Biol. 159, 311–326.
Tepass, U., Gruszynski-DeFeo, E., Haag, T.A., Omatyar, L., Torok, T.,
Hartenstein, V., 1996. Shotgun encodes Drosophila E-cadherin and is
preferentially required during cell rearrangement in the neurectoderm and
other morphogenetically active epithelia. Genes Dev. 10, 672–685.
Uemura, T., Oda, H., Kraut, R., Hayashi, S., Kotaoka, Y., Takeichi, M., 1996.
Zygotic Drosophila E-cadherin expression is required for processes of
dynamic epithelial cell rearrangement in the Drosophila embryo. Genes
Dev. 10, 659–671.
Zeng, W., Wharton Jr., K.A., Mack, J.A., Wang, K., Gadbaw, M., Suyama, K.,
et al., 2000. Naked cuticle encodes an inducible antagonist of Wnt
signalling. Nature 403, 789–795.




















