
Culture, Values, And The Environment In Central Africa - DUNE

*v: v
IF,
Ecology of Desert Environments, pp. 281 -291, 2001Editor: Ishwar PrakashScientific Publishers (India), P.O.Box 91,Jodhpurhttp://www.scientificpub.com
Thermal ecology and seasonal activity patterns ofErodius sauditus, an Arabian dune beetle
(Tenebrionidae: Erodinae)
Michael D: Robinson and David A. Clayton
Department ofBiology, Sultan Qaboos University, POBox 36,Al Khoud P.O. 123, Muscat, Oman
The dune beetle Erodius sauditus was the first crepuscular species to beinvestigated in an actograph. Data from field studies is presented oftheirbody Mil environmental temperatures and seasonal surface activitypatterns. While the mechanism of entrainment of the Orcadian clockrenins problematical, the data confirm that the activity is usuallybimodal butismodified by seasonal and daily changes in temperature.Keywords: circadian clocks; desert beetles; activity patterns —*•
Introduction vrs«-r-rr--
The influence of John Cloudsley-Thompson in the ^l^^J^^ZThompson and Chadwick, 1964; Clpudsley/rhompson,196$;L984, 1991) and tnechroSiology of the organisms that inhabit them (Cloudsley-Thompson 1960; 1961,1^3 1970 ^980 Applki et al., 1987) has been considerable and spanned most major^^^^^^T!^^^ 19?l; 1989a; 1991) ^oughout ^oridtravels Cloudsley-Thompsoh has been an enthusiastic ambassador of these subjects,eToS^gmahy researchers, including the present authors in their «£"™>"either dSectiy or through encouraging scholarship by to*******^^fserlArid Environments and the Springer Verlag book series, Adaptations of DesertOrganisms.
The timing of surface activity is recognised as af^^^^^^desert livine (Cloudsley-Thompson, 1991) and a number of species have been studiedLdetuXSdsley-Thompson (1956); Costa (1995) and S0mme (1995) and which
X

ECOLOGY OF DESERTENVIRONMENTS
(AFestschrift forProf. J.L.Cloudsley-Thompson on his 80thBirthday)
CONTR/BUTOR'SCopy
4»
•cd!H%WTfiHU3UPW3grii •CompeleassBt'gry .Copy
Editor
ISHWAR PRAKASHPh.il, D.Sc.
Desert Regional Station,Zoological Survey of India,
Jodhpur - 342 005 (INDIA).
SCIENTIFIC PUBLISHERS (INDIA)P.O. BOX 91 • JODHPUR
X

282 Robinson & Clayton
are described as either diurnal, nocturnal or crepuscular. Ignoring seasonal changesin abundance (El Borollosy & Awadallah, 1973; Richman et al., 1982), some of thelonger lived species also exhibit a seasonal change of activity (Alicata et al., 1979;Kramm & Kramm, 1972; Hafez & Makky, 1959; Holm & Edney, 1973).
It is also recognised that there is an underlying circadian rhythm to this activitythat is entrained by environmental 'zeitgebers' and a number of studies havedemonstrated this in species described as nocturnal (Constaninou & Cloudsley-Thompson, 1982) or diurnal (Constaninou & Cloudsley-Thompson, 1980) or bimodallyactive (Constaninou & Cloudsley-Thompson, 1985, Erbeling & Paarmann, 1985)where the entraining signal is either light or light and temperature.
Tenebrionid beetles are ubiquitous to many of the world's deserts (Crawford,1981), including Arabia (Abushama & Cloudsley-Thompson, 1978; Kasab, 1981;Aldryhim et al., 1992; Faragella, 1998). One of the eight species of the genus found inArabia, Erodius sauditus is a small (8-17 mm elytral length), short-lived,psammophilous tenebrionid beetle. In Kuwait this beetle is found in the accumulatedsand drifts formed on the southern edge of the Jal Az Zor escarpment where its adultlife cycle is played out for a little^bver 3 months, between late February and earlyJune each year. In 1989 its assortative mating system was investigated (Clayton,1991). Using beetles collected from Abdally in northern Kuwait (29 March 1983),Cloudsley-Thompson & Constantiriou (1985) described the endogenous circadianrhythm of Erodius octocostatus, which wajs later identified as E. sauditus (Al Houty,1989). It was therefore appropriate to investigate the natural free-running activitycycle of this species in its natural environment.
Methods
The study site was in one of the longer gulhes of the Mdairrah region along theJal Az Zor escarpment (29° 34' 20 N, 47r52' 40 E). Between 15February and 6 June1990 24 visits to this and two adjacent dune systems were made. At the main site a100m long, 20 m high sand dune slip face at the head of the gully was marked out in5m sections and hourly counts of beetles in each section were made. During each visitseveral environmental variables were recorded. A YSI Tele-thermometer, Model44TD equipped with Model 401 thermister probes was used to measure thetemperature of the air at a height of 5cm, the sand "surface in the sun and shade andin the sand at depths of 0.5, 5,10, 15, 20 and 25cm.
Additionailly, wind lipeed (m/sec) was measured with an anemometer (Casella,London) placed such that the revolving cups were at surface level and the relativehumidity with a twirling hydrometer. More specific temperatures of individualbeetles and the sand where they were found v/as also taken using a YSIsyringe probeModel 524 which was calibrated by a conversion chart supplied by the manufacturer.While most visits lasted 2-3 hrs, on three occasions beetle counts and temperatureswere recorded for several hrs and on a further five occasions data was collected over a24 hr period.
Results
The beetles begin to appear in late February and increase in numbers duringMarch; reaching a peak in early April (Figure 1), possiblybecause of the appearance
o
3Cft
co
>
*5
18
Seasonalactivity patterns ofanArabian dune beetle
I I I I I n i r i i i i i i i
10 MarchN=179
i i i i i m
:i rrm i n i h h rfrrri ii rffrt i-20 R8arch 24 RfiarchN-S42 N=905
I I .I I I I27 MarchN-702
-H~f
11 m 11 ii 11 in 1111111 ri nun^ . 6 ApriS
N=1021
12 15 18 21 24 3 € 9Time [hrs)
-4©
&
gc
5.
283
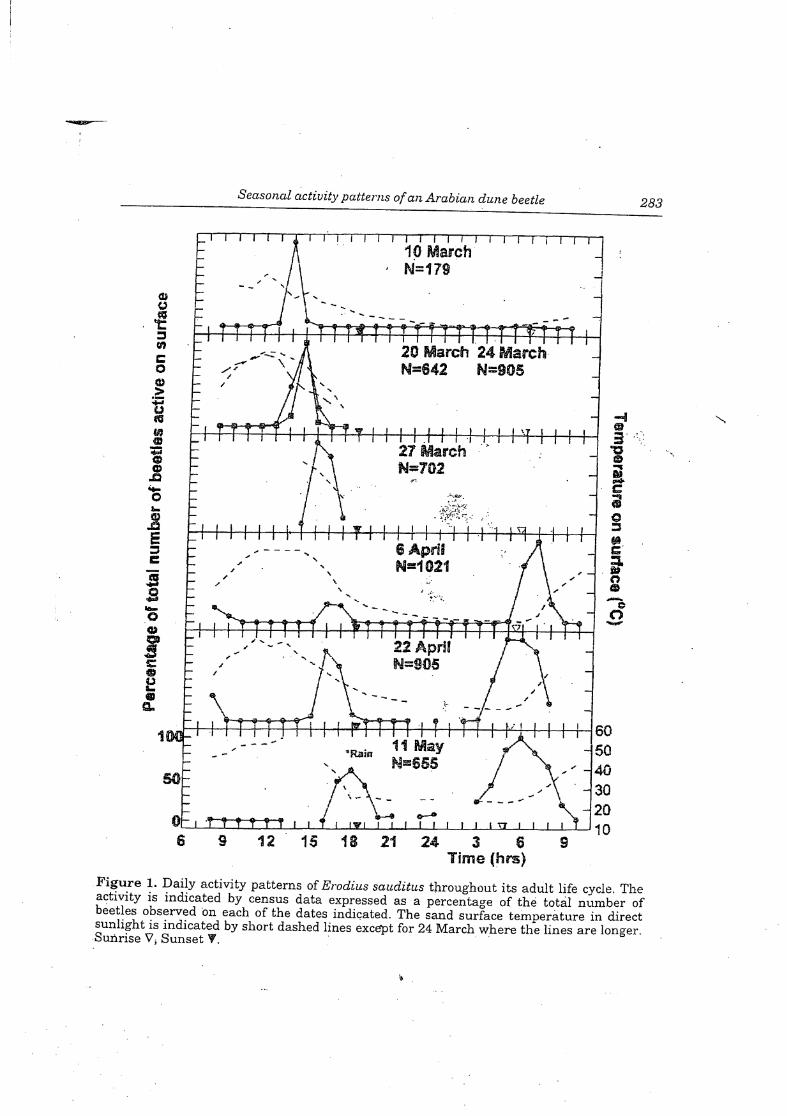
Figure 1. Daily activity patterns of Erodius sauditus throughout its adult life cycle. Theactivity is indicated by census data expressed as a percentage of the total number ofbeetles observed on each of the dates indicated. The sand surface temperature in directsunlight is indicated by short dashed lines except for 24 March where the lines are longerSunrise V; Sunset W.
N
V
284 Robinson & Clayton
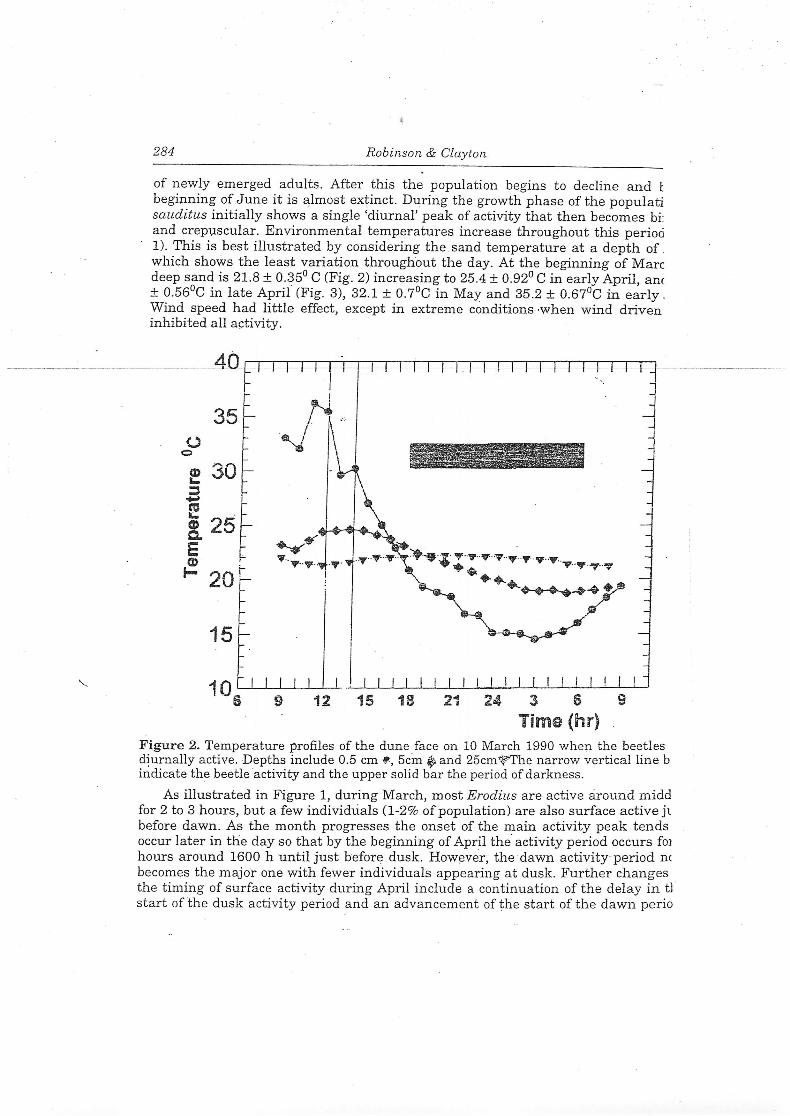
of newly emerged adults. After this the population begins to decline and Ibeginning ofJune it is almost extinct. During the growth phase of the populatisauditus initially shows a single 'diurnal' peak of activity that then becomes bi:and crepuscular. Environmental temperatures increase throughout this period1). This is best illustrated by considering the sand temperature at a depth of.which shows the least variation throughout the day. At the beginning of Marcdeep sand is 21.8 ± 0.35° C (Fig. 2) increasing to 25.4 ± 0.92° Cin early April, ant± 0.56°C in late April' (Fig. 3), 32.1 ± 0.7°C in May and 35.2 ± 0.67°C in early .Wind speed had little effect, except in extreme conditions when wind driveninhibited all activity.
Figure 2. Temperature profiles of the dune face on 10 March 1990 when the beetlesdiurnally active. Depths include 0.5 cm 9-, 5cm ^ and 25cm^-The narrow vertical line bindicate the beetle activity and the upper solid bar the period of darkness.
As illustrated in Figure 1, during March, most Erodius are active around middfor 2 to 3 hours, but a few individxials (1-2% of population) are also surface active jibefore dawn. As the month progresses the onset of the main activity peak tendsoccur later in the day so that by the beginning of April the activity period occurs foihours around 1600 h until just before dusk. However, the dawn activity period ncbecomes the major one with fewer individuals appearing at dusk. Further changesthe timing of surface activity during April include a continuation of the delay in tlstart of the dusk activity period and an advancement of the start of the dawn perio
Seasonal activity patterns ofanArabian dune beetle
12 15 18 21 24 3 6 9
Time (hr)
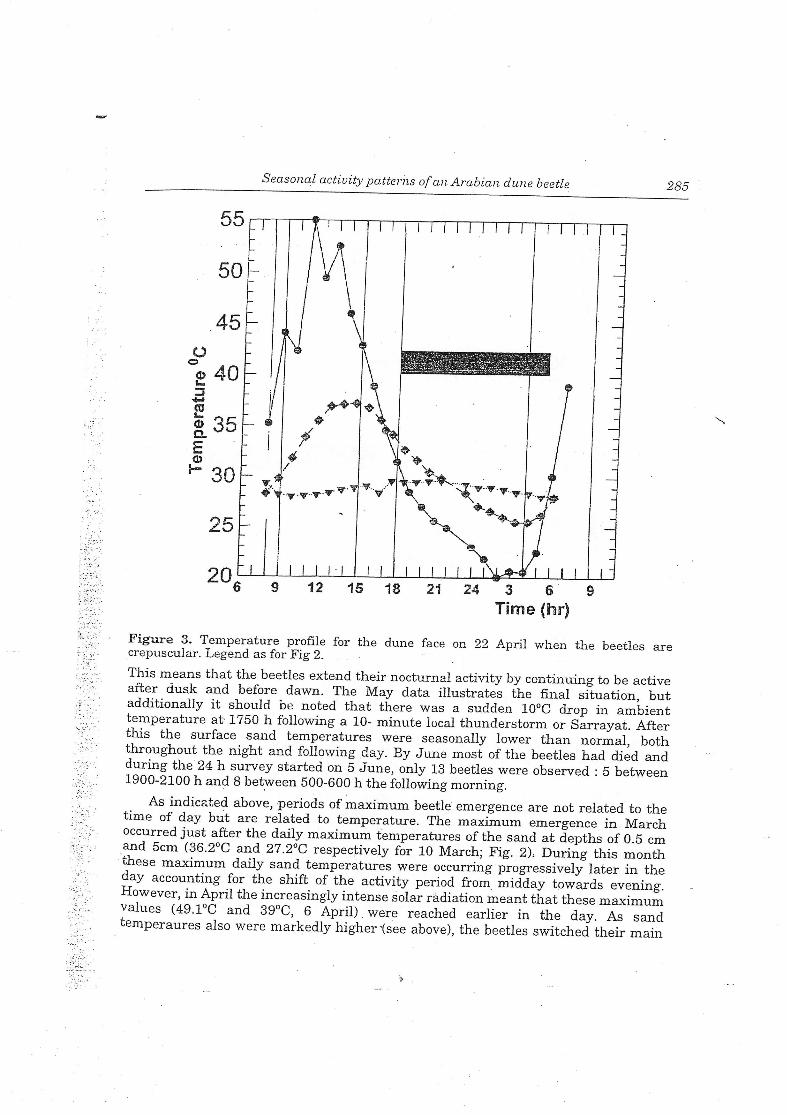
Figure 3. Temperature profile for the dune face on 22 April when the beetles arecrepuscular. Legend as for Fig 2. .
This means that the beetles extend their nocturnal activity bv continuing to be activealter dusk and before dawn. The May data illustrates the final situation butadditionally it should be noted that there was a sudden 10°C drop in am'bienttemperature at 1750 h following a 10- minute local thunderstorm or Sarrayat. Afterthis the surface sand temperatures were seasonally lower than normal boththroughout the night and following day. By June most ofthe beetles had died andduring the 24 h survey started on 5 June, only 13 beetles were observed •5 between1900-2100 h and8 between 500-600 h thefollowing morning.
As indicated above, periods ofmaximum beetle emergence are not related to thetime of day but are related to temperature. The maximum emergence in Marchoccurred just after the daily maximum temperatures ofthe sand at depths of0 5 cmand 5cm (36.2°C and 27.2°C respectively for 10 March; Fig. 2), During this monthtnese maximum daily sand temperatures were occurring progressively later in theday accounting for the shift of the activity period from midday towards eveningHowever, mApril the increasingly intense solar radiation meant that these maximumvalues (49.1°C and 39°C, 6 April) were reached earlier in the day As sandtemperaures also were markedly higher "(see above), the beetles switched their main
285
N
286 Robinson & Clayton
activity peak to the dawn period where the 5cm depth temperature was close to thelowest value in the 24 h period (Fig 3). Subsequently in the latter part of the beetle'slife cycle the emergence peaks coincided with the daily minimum (23.2 °C, 22 Apriland 26.7 °C, 11 May). While only a few data are available, on 20 March beetles werefound buried to a depth of 5-8 cm (n=5) while on 14 April they were at 10.5-21cm(n=ll).
Beetle temperatures at the morning emergence vary from 14°C in April to about24° C in March and May. Beetles remain active during the morning heating phaseuntil body temperatures reach 42 to 44°C (Table 1). In the afternoon they emerge atbody temperatures nlartheir voluntary maximum (43.24 ± 0.92 °C, n=9), and slowlydecrease to temperatures similar to the sand surface at the time of burial (24 to 37°C).
Table 1. Summary ofbody temperatures CO of Erodius sauditus during variousactivity periods
Date Time " Minimum Maximum Notes
March 27 13 : 50-17 :01 24.5 41.7 Estimated from
sand temperatureApril 7 05 : 26-08 :52 14.5 43.2 males
14.7 42.9 females
15 :45-18 :14 31.7 44.3 males30.7 44.8 "females
May 11 15 : 57-17 ::05 36.9 43.0 males
37.1 42.9 females
17: 30-19 : 04 26.9 30.7 males - after rain26.9 30.0 females - after ain
20 : 00-03 : 18 26.9 30.0 males
26.8 30.5 females
May 12 04:40-09 : 31 24.5 43.7 males
24.7 42.7 females
Discussion
Rhythmic locomotory activity has been described in various desert arthropods,and tenebrionid beetles have been a favourite organism of field and laboratoryresearchers alike (Cloudsley-Thompson, 1961; Cloudsley-Thompson 1989b; Cloudsley-Thompson, 1991; Cloudsley-Thompson & Constantinou, 1981; 1985; Crawford, 1981;Applin et al., 1987; Alpatov et al., 1994). Internal circadian clocks have beenpostulated to be the primary regulatory mechanism of activity and would appear to beespecially important for burrowing species which live in relatively stableenvironments that lack usual external.stimuli. Alpatov et al., (1999) conclude thatcircadian rhythms are the product of endogenous regulation by a circadianpacemaker, which is probably two coupled oscillators (Alpatov et al., 1994), and byexogeneous 'masking3 of the rhythm by environmental stimuli that allow fine tuningto changing local conditions (Rietveld et al., 1993).
Erodius initially appears to have one activity peak during the day, but thenswitches to being crepuscular. It is appropriate to consider these phases separately
X

V
Seasonal activity patternsofanArabian dunebeetle 287
and to begin with the latter as this period ofErodius' life cycle is closely matched tothe locomotory activity in an actograph (Cloudsley-Thompson &Constantinou, 1985).
In late April and May emergence ofErodius occurred before dawn, and in Maythe afternoon activity extended into the night. In the actograph at a constanttemperature of 25 °C, most activity occurred from about 1 h before the onset of thelight perioduntil just before the onset of the dark period. Further, activity was muchreduced in the middle of the light period as would be the case for a crepuscularspecies. Nevertheless, because the start ofactivity was delayed and the free runningperiod was lengthened in constant light, Cloudsley-Thompson & Constantinou (1985)concluded that the circadian rhythm of this species was physiologically more like thatof a nocturnal than a diurnal species. Entrainment of the beetles' circadian rhythmwas by the onset of dark.
In the laboratory, individual Trigonoscelis gigas exhibit a wide range offree-running circadian patterns, but have a tightly maintained crepuscularperiod ofsurface activity in the field (Alpatov et aL, 1994): Beetles held in buried activitymeters in their natural habitat in the Karakum desert showed an entrained circadianrhythm when at a depth (5 cm) where"the temperature showed a daily cyclicalvariation and a free-running rhythm when buried deeper (25 cm) where thetemperature was constant (Alpatov et aL, 1999). During inactivity the beetles werenormally found at the greater depth and Alpotov et al., (1999) suggested that either atemperature or light zeitgeber operate during the short activity period.
In Kuwait sand temperatures at 25 cm also remain relatively constant, but asErodius only buries shallowly, the zeitgebers could be either the daily high or lowtemperature values beneath the sand or the onset of darkness as suggested from thelaboratory study (Cloudsley-Thompson & Constantinou, 1985).
When the switch from the single to the double period of activity occurred at thebeginning ofApril and thereafter, the population's morning emergence and afternoonburial times coincided with sunrise and sunset and not any particular sand or bodytemperature. At this time light intensity could be the environmental signal for timingthe cycles, but in the first, day-activepart of the season, it is more likely that the cuewas temperature. Additional actograph studies which manipulate the ambienttemperature as Constantinou & Cloudsley-Thompson (1980) performed on Adesmiacancellata may resolve this issue. What is clear however, is that the beetles use theambient temperature to time their activity. In the early part of their season it is toocold for dawn activity, in May the beetles extended their activity after a sudden rain
.shower when the sand temperature was 3 degrees less than normal. The presence ofmoisture in the habitat, however, mayhave contributedto the extendedactivity.
On 7and 8 April, beetles emerged at dawn but environmental and bodytemperatures were 10°C lower than in March or May (Table 1), However, thesubnormal surface temperatures didnot deter the beetles' emergence, although theirmovements were greatly impaired and predators could easily have taken them. Atsimilar low body temperatures some diurnal Namib Desert dune tenebrionidsoccasionally emerge at night into the incoming moist fog. Here the reason seemsclear; dehydration drives the beetles to abort normal activity rhythms and riskpredation (Seely, 1979 ).

ooo
Robinson &Clayton
available to the organisms after their arousalbyT^rcJl clock ^ ^^ ^Although the identity and mechanism of such signals have not been clarified «,„
temperature and gaining time to maximize food intake snd^L^e mats ^Several of our observations ofErodius sauditus support these concents Th* *Wbeetles to emerge on the'morning of T.April were predomlanS^malesXm^S*00to
S^aturelr.l^cSelr'3' " ^ 1!mited fr°m W~J6 JSOn several occasions not restricted to that following rain, females extended th.ir
afternoon acfavity beyond that ofmost males. During the last30 to4? ml3« £.
hv J^ ^ When STmer SUrface temPeratures are suitable (20-45°C) for activitv
rriess^ilS ' y^ seemin^ ^appropriate conditionsTdesert dules SUrVWal m^^^cted"environments such
Kdd observations on the activity pattern ofJ?, sauditus generally corroborate theactograph studies of Cloudsley-Thompson &Constantinou(19^7beetieIs ~normally crespuscular, and the circadian clock appears to be entrained by the oSet ofdarkness However, field data suggest that sand temperaturTiTan h^tLtSSTS tWUeSc1 *? T "ri68 t°1time ^ ^seasonal"urTaceTcS^according to their ecological and physiological needs. Alone, either type of studvprovides an incomp ete picture of the rhythmic acitivity of desert^AoSstudSsTeSeS* °f Pr°feSSOr ^^-^-P-n, both field ana SSoryti^a^ understanding of chronobiology and"
References
Abusliama, F.T. &Cloudsley Thompson, J.L. (1978). Desert arthropods of Kuwait and theirdistribution. Entomologist's Monthly Magazine, 114:149-151. Kuwait and tneir
N

Seasonal activity patterns ofan Arabian dune beetle 289
Aldryhim, Y.N., Mills III, C.W. & Alawood, AS. (1992). Ecological distribution and seasonalityof darkling beetles (Coleoptera: Tenebrionidae) in the central region of Saudi Arabia.Journal ofArid Environments, 23: 415-422.
Al Houty, W. (1989). Insects of Kuwait. University of Kuwait, Al Marzouk Press, Kuwait, pp187.
Alicata, P., Caruso, D., Costa, G, Marcellirio, I, Motta, S., Petralia, A. (1979). Ricercheeco-etologiche sulla fauna delle dune costiere di Portopala (Siracusa). II Comportamento,distribuzione e ritmi di attivata di Pimilia grossa Fabr (Coleoptera, Tenebrionidae).Animalia: 6, 33-48.
Alpatov, A.M., Rietveld, W.J., Oryntaeva, L.B. & Putilov, A.A. (1994). Properties of thetwo-peak free running circadian rhythm of locomotor activity of the sand-deserttenebrionid beetle Trigonoscelis gigas Reitter. Biological Rhythm Research, 25:153-167.
Alpatov, A.M.. Zotov, V.A., Tshernyshev, W.B. & Rietveld, W.J. (1999). Endogenous circadianrhythm is a crucial tool for survival of the sand-desert tenebrionid beetle Trigonoscelisgigas Reitter. Biological Rhythm Research, 30:104-109. ^
Applin, D.G., Cloudsley-Thompson, J.L & Constantinou, C. (1987). Molecular and physiologicalmechanisms in chronobiology —their manifestations in the desert ecosystem. Journal ofAi'id Environments, 13:187-198.
Clayton, D.A. (1991). Assortative mating in the dune beetle Erodius sauditus (Tenebrionidae:Erodiini) in Kuwait. Journal ofArid Environments, 21: 81-89.
Cloudsley Thompson, J.L. (1956). Studies in diurnal rhythms-VI Bioclimatic observations inTunisia and their significance in relation to the physiology of the fauna, especiallywoodlice, centipedes, scorpoins and beetles. Annual Magazine of Natural history, 12:305-329.
Cloudsley Thompson, J.L. (1960). Adaptive functions of circadian rhythms. Cold SpringsHarbor Symposium on Quantitative Biology. 25, 345-355.
Cloudsley Thompson, J. L. (1961). Rhythmic activity in animal physiology and behaviour.London: Academic Press.
Cloudsley Thompson, J.L. (1963). Light responses and diurnal rhythms in desert tenebrionidae.EntomologiaExpenmentalisetApplicata, 6:75-78.
Cloudsley Thompson, J.L. (1968). The Merkhiyat Jebels: A desert community. In: Brown, G.W.(Ed.) Desert Biology Volume 1, pp. 1-20. London: Academic Press.
Cloudsley Thompson, J.L. (1970). Recent work on the adaptive function of circadian andseasonal rhythms in Invertebrates. Journal ofInterdisciplinary Cycle Reseach, 1: 5-19.
Cloudsley Thompson, J.L. (1980). Biological clocks:. Their fuctions in nature. London,Wiedenfeld & Nicolson. 138pp
Cloudsley Thompson, J.L. (1981). A comparison of rhythmic locomotory activity in tropicalforest Artropods with that in desert species. Journal ofArid Environments, 4: 327-334.
Cloudsley Thompson, J.L. (1984). Terrestrial animals in dry heat: Arthropods. In: Dill, D.B(Ed.) Adaptation to the environment. Handbook ofPhysiology, 4:. 451-465.
Cloudsley Thompson, J.L. (1989a). Temperature and the activity of ants and other insects., incertral Australia. Journal ofArid Environments, 16:185-192!
Cloudsley Thompson, J.L. (1989b). Behavioural adaptations of desert arthropods. Both Acad.Gioenia Sci. Nat. 87: 20 (332): 282-293.
Cloudsley Thompson, J.L. (1991,). Ecophysiology of desert arthropods and reptiles. Berlin:Springer-Verlag.

290 Robinson &Clayton
Cloudsley Thompson, J.L. &Chadwick, M.J. (1964). Life in deserts. London, Foulis. 205 pp.Cloudsley Thompson, J.L. &Constantinou, C. (1981). Effects of light and temperature on t]
circadian rhythmic of locomotory activity of Pimilia grandis Klug (Coleootertenebnomdae) from Sudan. Journal ofAridEnvironments, 4" 131-163.
Cloudsley-Thompson, J.L. & Constantinou, C. (1985). Biological clocks in desert beeth(Tenebnomdae) with special reference to Erodius octocostatus Peyerinhof in KuwaiJournal ofthe University ofKuwait, 12:237-242.
Constantinou, C. & Cloudsley-Thompson, J.L. (1980). Circadian rhythmicity in Adesmicancellata L. (Coleoptera :Tenebrionidae) from Kuwait. Journal ofArid Environments, I
Constantinou C. &Cloudsley-Thompson, J.L. (1982). Circadian rhythmicity in Trachyderrrux£ci%nForsk) (Cole°Ptera :Tenebrionidae) from Sudan. Journal ofArid EnvironmenU5: 369-374.
Co'nstantinou, C. &Cloudsley-Thompson, J. L. (1985). Interactions between light and temperature m synchronising the circadian rhythm of locomotary activity in Thermophilunduodeamguttatum (Coleoptera : Carabidae) from Kuwait. Journal ofArid Environments8:157-164.
Costa, G. 1995. Behavioural adaptations ofdesert animals. Berlin: Springer-Verlag.Crawford, C.S. (1981). Biology ofdesert Invertebrates. Berlin: Springer Verlag. 314pp.El Borollosy, F.M. &Awadallah, K.T. (1973).'On the seasonalabundance ofthe carabid beetle
Calosoma chlorostictum chlorostictum DEG. Bulletin of the Society of Entomologists ofEgypt, 57j 247-250. s J
Erbeling, L. & Paarmann, W. (1985). Diel activity patterns of the desert carabid beetleThermophilum (=AnthiaJ sexmaculatum F. (Coleoptera: Carabidae). Journal of AridEnvironments, 8: 141-156.
Faragella, A.A. (1998). Seasonal occurrence and habitat distribution of tenebrionid beeltes(Coleoptera : Tenebrionidae) in Khulais Valley, Western Saudi Arabai. Elytron 12105-112. J
Hafez, M. & Makky A.M.M. (1959). Studies ondesert insects in Egypt III. On the bionomics ofAdesmia bicannata Klug (Coleoptera : Tenebrionidae). Bulletin of the Society ofEntomologists ofEgypt, 43: 89-113.
Holm, E. &Edney, E.B. (1973). Daily activity ofNamib desert arthropods in relation to climate.Ecology, 5.4: 45-56. -—- ~
Kasab, Z. 1981. Insects of Saudi Arabia. Coleoptera: Family Tenebrionidae (Part 2). In:Wittmer, W. &Buttiker, W. (eds) Fauna ofSaudiArabia,3, 276-401.
Kramm, R.A. & Kramm, KR., (1972). Activities of certain species of Eleodes in relation toseason, temperature and time of day at Joshua Tree National Monument (Coleoptera :Tenebrionidae). TheSouthwestern Naturalist, 16:341-355.
Marden, J.H. (1987). In pursuit of females : following and contest berhaviour by males of aNamib Desert tenebrionid beetle, Physadesmiaglobosa. Ethology, 75:15-24.
Richman, D.B., Huddleston, E.W. &Ortiz, M. (1982). Seasonal activity oftenebrionid beetles inNew Mexico mesquite dunes. The Southwestern Naturalist, 27:305-308.
Rietveld, W.J., Minors, D.S., Waterhouse, J. M. (1993). Circardian rhythms and masking : anoverview. Chronobiology International, 18: 306-312.
Roberts, C.S., Seely, M.K., Ward, D., Mitchell, D., &Campbell, J.D. (1991). Body temperaturesof Namib Desert tenebrionid beetles: their relationship in laboratory and field.Physiological Entomology, 16: 463-475.
N

Seasonal activity patterns ofan Arabian dune beetle 291
Robinson M.D. & Seely, M.K. (1980). Physical and biotic environments of the southern Namibdune ecosystem. Journal ofArid Environments, 3: 183-203.
Seely, M.K. (1979). Irregular fog as a water ^source for desert dune beetles. Oecologia, 42:213-227.
Slobodchikoff, C.N. (1983). Water balance and temperature preferences, and their role inregulating activity times of tenebrionid beetles. Oikos: 40 :113-119.
S0mme, L. (1995). Invertebrates in hotand coldarid environments. Berlin: Springer-Verlag,Ward, D., & Seely, M.K. (1996).Behavioural thermoregulation in six Namib Desert tenebrionid
beetle species.(Coleoptera). Annals Entomological SocietyofAmerica, 89:442-451.
X
Copyright © 2022 FDOKUMEN




















