
Alternate Reproductive Tactics in an African Dung Beetle, Circellium bacchus (Scarabeidae
13
Alternate Reproductive Tactics in an African Dung Beetle, Circellium bacchus (Scarabeidae) Chevonne Reynolds & Marcus J. Byrne Revised: 24 October 2012 / Accepted: 25 October 2012 # Springer Science+Business Media New York 2012 Abstract Male-male competition is recognized as a potent force of sexual selection. When intra-sexual competition is strong then selection theory predicts that alternative male phenotypes will evolve. Circellium bacchus is a large, hornless, ball-rolling dung beetle with extensive variation in size and subject to intense male-male contest competition. We proposed that small male C. bacchus, which are unlikely to be successful in male contest competition, would adopt the alternative reproductive tactic of sneaking copulations. This alternative tactic is likely to influence not only behavior, but morphology with an expectation that sneaking males would invest more resources in testes development. Investigation of testis allometry revealed that smaller male C. bacchus beetles had relatively larger testes than their bigger conspecifics. Furthermore, as resources may be limited during larval development, differential investment in testes development should result in adult male beetles either competing for fertilizations or for access to mates. This is seen in C. bacchus as two alternate male phenotypes; smaller beetles with a relatively low body mass invest proportion- ately more in testes development compared to larger, heavier form in which testes size does not scale with condition. To our knowledge this study provides the first investigation of alternative phenotypes in the reproductive tactics of a ball-rolling dung beetle species. Keywords Alternative reproductive tactics . male-male competition . sperm competition . operational sex ratio . Circellium bacchus . testis . sneaks J Insect Behav DOI 10.1007/s10905-012-9365-1 C. Reynolds : M. J. Byrne School of Animal, Plant and Environmental Sciences, University of the Witwatersrand, Private Bag 3, Wits 2050, South Africa C. Reynolds Organization for Tropical Studies, South Africa Program, P.O. Box 33, Skukuza 1350, South Africa C. Reynolds (*) Percy FitzPatrick Institute of African Ornithology, DST/NRF Centre of Excellence, University of Cape Town, Rondebosch, Cape Town 7701, South Africa e-mail: [email protected]
Transcript of Alternate Reproductive Tactics in an African Dung Beetle, Circellium bacchus (Scarabeidae

Alternate Reproductive Tactics in an African Dung Beetle,Circellium bacchus (Scarabeidae)
Chevonne Reynolds & Marcus J. Byrne
Revised: 24 October 2012 /Accepted: 25 October 2012# Springer Science+Business Media New York 2012
Abstract Male-male competition is recognized as a potent force of sexual selection.When intra-sexual competition is strong then selection theory predicts that alternativemale phenotypes will evolve. Circellium bacchus is a large, hornless, ball-rollingdung beetle with extensive variation in size and subject to intense male-male contestcompetition. We proposed that small male C. bacchus, which are unlikely to besuccessful in male contest competition, would adopt the alternative reproductivetactic of sneaking copulations. This alternative tactic is likely to influence not onlybehavior, but morphology with an expectation that sneaking males would invest moreresources in testes development. Investigation of testis allometry revealed that smallermale C. bacchus beetles had relatively larger testes than their bigger conspecifics.Furthermore, as resources may be limited during larval development, differentialinvestment in testes development should result in adult male beetles either competingfor fertilizations or for access to mates. This is seen in C. bacchus as two alternatemale phenotypes; smaller beetles with a relatively low body mass invest proportion-ately more in testes development compared to larger, heavier form in which testessize does not scale with condition. To our knowledge this study provides the firstinvestigation of alternative phenotypes in the reproductive tactics of a ball-rollingdung beetle species.
Keywords Alternative reproductive tactics . male-male competition . spermcompetition . operational sex ratio .Circellium bacchus . testis . sneaks
J Insect BehavDOI 10.1007/s10905-012-9365-1
C. Reynolds :M. J. ByrneSchool of Animal, Plant and Environmental Sciences, University of the Witwatersrand, Private Bag 3,Wits 2050, South Africa
C. ReynoldsOrganization for Tropical Studies, South Africa Program, P.O. Box 33, Skukuza 1350, South Africa
C. Reynolds (*)Percy FitzPatrick Institute of African Ornithology, DST/NRF Centre of Excellence,University of Cape Town, Rondebosch, Cape Town 7701, South Africae-mail: [email protected]

Introduction
Within a species, individuals vary in their ability to compete for mates, thus affordingsome members a reproductive advantage over others of the same sex (Darwin 1871).Competition for mates by males has long been recognized as important in shaping theevolution of a variety of exaggerated secondary sexual characteristics (Andersson1994; Emlen 2008). These include the bright, dramatic plumage seen in birds-of-paradise (Gilliard 1969; Diamond 1986), the color diversification of Malawiancichlids (Turner 1994; Deutsch 1997; van Oppen et al. 1998) and weapons ofmale-male competition, such as the antlers of deer or the horns of dung beetles(Kruuk et al. 2002; Emlen et al. 2005; Emlen 2008). Consequently, males whichare less attractive to females or are unsuccessful in male-male competition must adoptalternative tactics to achieve reproductive success (Simmons et al. 2007). So, underthese circumstances how does a less well-endowed individual gain the upper hand?
Male dung beetles compete to gain access to a female, with the outcome of thesecontests usually predicted by body size (Rasmussen 1994) and/or extent of weaponry(Knell 2011). This suggests that successful reproduction will become skewed towardsmales with the most prominent traits (Simmons 2011). However, Emlen and Oring(1977) propose that in species where individuals experience strong intra-sexualcompetition, then selection theory predicts the evolution of alternative male behaviorsin order to gain access to mates. In many tunneling dung beetle species, smaller malesthat are usually unsuccessful in male-male contests, attempt to gain matings bysneaking copulations with receptive females by digging side tunnels into the nest,whilst their competitive superiors are preoccupied with mate-guarding behavior at themain entrance to the nest (Simmons et al. 2007). This sneaking ploy has given rise tosignificant evolutionary consequences for male behavior, morphology and physiolo-gy (Birkhead 1998; Byrne et al. 2002; Longair 2004; Simmons and Emlen 2006;Simmons et al. 2007). These include a developmental decision between alternativephenotypes, often based on male body size. This is best exemplified by larger malesin the dung beetle genus Onthophagus (Scarabaeidae: Scarabaeinae) bearing horns,which they use to guard access to females, while their smaller, sneakier counterpartsare hornless (Emlen 1997).
Females of many taxa often mate with more than one male (Parker 1970; Birkheadet al. 1987; Parker 1990). This generates sperm competition where the sperm ofdifferent males contest, after copulation, to fertilize the available ova (Parker 1970).Sperm competition theory predicts that i) sperm compete in a situation analogous to araffle whereby fertilization probability increases with the proportion of a given male’ssperm in the female’s reproductive tract and ii) an increased risk of sperm competitionwill require increased expenditure on ejaculate by competing males (Parker 1990). Byvirtue of their mating tactics, sneaking males will always be subject to spermcompetition (Simmons et al. 2007). Sperm competition thus favors traits such aslarger testes size, which produce greater numbers of sperm required for largerejaculates (Parker 1998) and should improve the male’s fertilization success(Simmons and García-Conzález 2008). Supporting empirical evidence shows thatmales which are subject to intense sperm competition invest more resources inqualities such as larger testes and increased ejaculate volume (Møller 1991; Gage1995; Harcourt et al. 1995; Byrne et al. 2002; Pitcher et al. 2005).
J Insect Behav

Promotion of sperm competition by a female can enhance her reproductive fitness(Simmons 2011), especially in the case of dung beetles, where male-male competitioninfluenced by body size is not necessarily a good indicator of male fitness. This is aconsequence of a low heritability of body size, which is instead primarily dictated bythe size of the brood ball in which the individual developed. Larvae with access tomore food resources can thus grow larger and consequently molt into larger pupaeand adults (Emlen 1997; Moczek 2011). This can result in extensive variation in sizewithin a single beetle species and provides a useful framework to study the evolutionof alternative breeding tactics where selection for different phenotypes with differentmating tactics is predicted (Emlen 1997; Watson and Simmons 2010).
These alternate mating tactics have been well studied in tunneling dung beetlespecies where large “major” males have horns and smaller “minor” males are usuallyhornless sneakers (see reviews in Simmons and Ridsill-Smith 2011). However, fewstudies have investigated the evolution of alternate breeding tactics in hornless ball-rolling dung beetles, which do not build nests with foraging tunnels that can guardedby the “major” male. Sato and Hiramatsu (1993) examined sexual selection in theball-rolling Khepher platynotus, Bates (Scarabaeidae: Scarabaeinae) and Le Roux etal. (2008) made behavioral and allometric observations on mate selection in theflightless ball-roller Circellium bacchus Fabricius (Scarabaeinae: Canthonini). Bothspecies show increasingly male skewed sex ratios during the breeding season, whichwould be predicted to lead to male-male competition and to sperm competition.
Circellium bacchus is a large, hornless, diurnal dung beetle species which occursin the Eastern Cape and Western Cape provinces of South Africa. Sexual dimorphismis only apparent in a tiny tuft of hairs seen on the hind legs of males. Although bothsexes roll food balls, only female C. bacchus make brood balls (Kryger et al. 2006).Male C. bacchus typically compete for access to ball rolling females, with largermales winning significantly more fights (Kryger et al. 2006). This male then followsclosely behind the female as she rolls the ball away in search of a suitable burial site.The victorious male perches on top of the brood ball as it is buried by the female andsubsequently copulates with her underground (Le Roux et al. 2008). The male re-emerges after 5–7 days and continues in search of additional mates at dung piles,whilst the female remains underground for the duration of larval development, whichmay take up to 6 months (Kryger et al. 2006). Small sneaking males have been seento circumvent contesting larger males to get access to the female, and successfullycopulate with her (Le Roux et al. 2008). The differing amounts of time and energyinvested in reproduction by each sex results in a male-biased operational sex ratio andsuggest that there will be increasing competition amongst males (Emlen and Oring1977), as more females are occupied in brood care as the season progresses.
Both pre- and post-copulatory mate selection mechanisms have been investigatedin C. bacchus (Le Roux et al. 2008). Although the species lacks any obvioussecondary sexual characteristics, males are very aggressive towards each other whencompeting for females and male-male contest competition probably plays a centralrole in pre-copulatory mate selection. The allometry which underlies this behavior isinteresting and sometimes surprising. For instance the “one size fits all” hypothesis(Eberhard et al. 1998) was proposed to explain the negative allometric relationshipbetween aedeagus size and body size in male C. bacchus. This post-copulatory mateselection mechanism assumes that genitalia which fit average-sized females allow
J Insect Behav

males to improve fertilization success by not being restricted to similar sized females (LeRoux et al. 2008). Both these mechanisms have merit. However, given the large sizerange seen in C. bacchus (Body length; range 22–47 mm, males x037.5±5.3 mm,females x035.1±4.1 mm; Kryger et al. 2006), we predict that testes size should alsoscale negatively with body size, but driven by male-male competition not femalechoice.
Smaller male C. bacchus, which are unlikely to be successful in contest compe-tition, should adopt the alternative reproductive tactic of sneaking copulations andincrease their investment in sperm production (Vahed and Parker 2011). Dung beetletestes develop during the larval stage and their size is dependent on the resourcesinvested in them at that stage (Simmons et al. 2007). However, Carsten-Conner et al.(2010) found that larger testes and larger ejaculates also required resources for theirmaintenance in walnut flies, suggesting that a resource allocation trade-off duringdevelopment will result in dung beetle males either competing for fertilizations orcompeting for access to mates as an adult (Simmons and Emlen 2006). Because largertestes and ejaculates are also costly to maintain, a further resource partitioning mayoccur after eclosion (Simmons and Kotiaho 2007), where adult resources could eitherbe directed towards building muscle mass or diverted to testes maintenance.
In this study we investigated the relationship between body size and testes size inC. bacchus. We predicted that i) smaller male beetles should have larger testesrelative to their body size and ii) investment in testes would be traded-off againstbody condition. The study provides the first examination of alternative phenotypes inthe reproductive tactics of a ball-rolling beetle.
Methods and Materials
Study Species
Circellium bacchus is a large and flightless dung beetle (Fabricius 1781). Its distri-bution is restricted to a few habitat fragments in the Western and Eastern CapeProvinces of South Africa (Chown et al. 1995), which qualifies the species asVulnerable according to the IUCN. They are unusual in that only female beetlesinitiate, form and roll brood balls, an exercise which is usually carried out by malesalone or in cooperation with the female in other species (Scholtz 2009). Furthermore,fecundity is among the lowest recorded for any dung beetle with an average of onlyone offspring produced a year. Female C. bacchus exhibit remarkable maternal care,remaining underground with the brood for a minimum of 120 days until the new adultemerges (Kryger et al. 2006).
Study Site
De Hoop Nature Reserve in the Western Cape Province of South Africa (34° 26′ S20° 32′ E) covers an area of 34 000 hectares and ranges in altitude from 0 to 290 m.Mean temperatures range between 12 and 22 °C and average rainfall is 350–600 mmper year, with 30 % falling in the winter months. The vegetation is classified asLimestone Fynbos and is characterized by Limestone Sugarbush (Protea obtusifolia),
J Insect Behav

Limestone Conebush (Leucadendron meridianum) and Limestone Pincushion(Leucospermum truncatum). However, huge stands of the exotic invasive, Rooikrans(Acacia cyclops) are also common in this vegetation type (Rebelo 1998).
Population Characteristics and Allometry
Data collection was completed from the 29th–31st October 2011, during the summerbreeding season which usually falls in October/November (Kryger et al. 2006). Atotal of 110 C. bacchus specimens were collected by both active searching and pitfalltrapping from one area in the reserve: an open, undisturbed, grassy patch adjacent toan Acacia cyclops stand.
All beetles were sexed using the single row of short stiff bristles on the tibia of thehind leg of the males. The following measurements were taken on all beetlescaptured; body mass (Electronic Balance -Shimadzu Corporation Bl-320H;±0.001 g); pronotum width; anterior-right tibia length and the distal tibial toothlength (Digital calipers -Toolquip and Allied EV150±0.01 mm). Body mass dividedby pronotum width was used as an index of condition, and the tibial tooth lengthdivided by tibia length was used as a proxy for age (Tynedale-Biscoe 1978). Beetlesnot used for the rest of the study were released were they were collected.
Thirty randomly selected male beetles (approximately half of the male specimenscollected) were killed by immersion in 75 % alcohol for 30 min, and then dissectedimmediately from the dorsal surface. The bright orange testes were removed and mountedin a drop of water on a microscope slide. A digital micrograph was taken at × 10magnification (Wild M8 microscope; ScopeImage camera) and the surface area of thelarger testis was measured using image analysis software (ImageJ). The surface area of thetestis was used as a proxy for testis size. To correct for body size, relative testis size wasderived from the testis area divided by the body mass.
Data Analyses
Statistical analyses were performed in STATSISTICA 8.0 (Statsoft 2006). Datawere tested for normality and transformed as required by the assumptions of thestatistical tests used.
A chi-squared test (with Yates’ Correction; Flower et al. 1998) was used todetermine if the ratio of male to female beetles at the study site was significantlydifferent from one. As the breeding season progresses, the ratio of male to femalebeetles becomes largely male-biased because brooding females remain underground,and is indicative of the intense competition experience by male beetles (Kryger et al.2006). Two-tailed t-tests were used to determine if differences between the sexes insize, weight, condition index and age were statistically significant.
Linear ordinary least squares (OLS) regression was used to determine whether arelationship existed between i) body size and relative testis size; ii) body size and thecondition index; and iii) the condition index and relative testis size (Eberhard et al1998). There has been some debate in the literature over which regression method ismore suitable for these kinds of analyses (Eberhard et al. 1999; Green 1999). SimpleOLS regression is a Model I regression and assumes a functional or causal linearrelationship between a dependent y variable and an independent x variable. It also
J Insect Behav
assumes that the x variable has been measured without error (Flower et al. 1998).Green (1999) however, argues that both variables will have been measured with errorand that a Model II reduced major axis (RMA) regression is better suited for dealingwith the inherent variability. RMA regression was therefore applied to these data, butas the results were not different to the OLS regression the RMA results are notreported further.
Results
Population Characteristics
A total of 110 beetles were caught over the sampling period; 77 males and 33females, resulting in a significantly male-biased sex ratio (M:F 1:0.43; Χ2016.81;P<0.001).
The size of the beetles varied across a wide range of pronotum widths (range17.98–29.65 mm; n0110), with males (range 17.98–29.65 mm; x025.10±2.50 mm,n077) being significantly larger on average than females (range 19.47–28.45; x023.24±2.36, n033) (t10803.61; P<0.001) (Fig. 1). Male beetles were also signifi-cantly heavier and had a significantly higher condition index (x mass05.62±1.52 g; xCI00.221±0.043) than females (x mass04.63±1.50 g; x CI00.196±0.047) (t10803.17; P00.002 and t10802.80; P00.006 respectively). Female beetles were signifi-cantly younger than males (t1080−2.27; P00.025) (Fig. 2).
Allometry
The random subset of 30 male beetles selected to explore alternative reproductivetactics exhibited a normal size distribution (Χ201.65; P00.200), and pronotum width(range 19.38–29.65 mm; x025.30±2.55 mm); which was not significantly differentto the entire sampled male population (t10500.38; P00.703).
16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
Pronotum Width (mm)
0
2
4
6
8
10
12
14
16
Fre
qu
ency
MaleFemale
Fig. 1 Size dimorphism of males and females in a population of Circellium bacchus dung beetles. Malesare significantly larger than females. (t10803.61; P<0.001). n077 males; 33 females
J Insect Behav

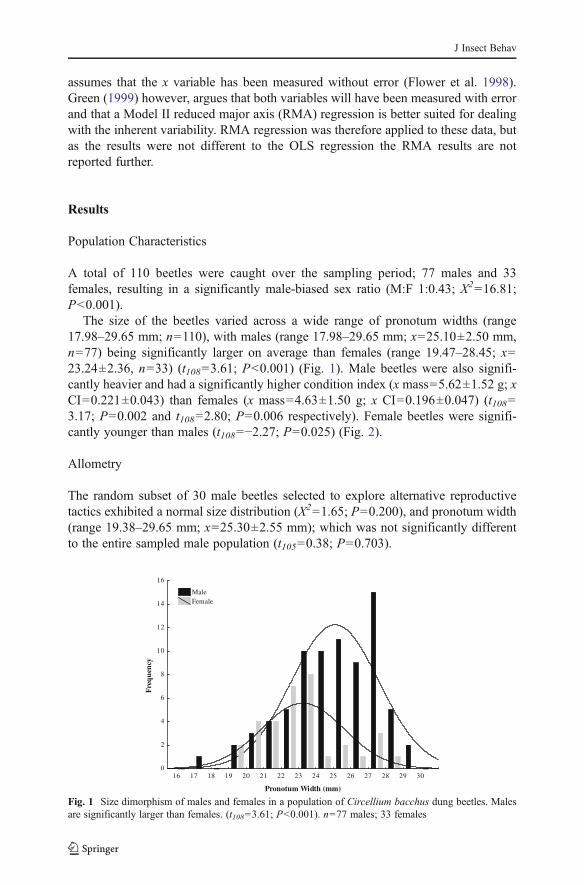
A negative allometric relationship was found between relative testis size and bodysize, indicating that smaller beetles have relatively larger testes compared to theirlarger conspecifics (F1, 28042.22; P<0.000) (Fig. 3). By plotting body size againstcondition index, smaller males, with a pronotum width less than 25.3 mm, werefound to have a significantly shallower slope of condition index (F1, 28022.972; P<0.000) than larger males (>25.3 mm pronotum width), creating a break-point in theregression around 25 mm pronotum width (Fig. 4). This break-point was randomlychosen as the mean pronotum width of the sample; however similar results were seenwhen the median or mode were used as break-points. This break-point was also seenwhen condition index was regressed against relative testis size, where lower condition(smaller) males showed a significant negative relationship between condition index
elameFelaM
Sex
0.17
0.18
0.19
0.20
0.21
0.22
0.23
0.24
Con
diti
on I
ndex
(M
ass/
Pro
notu
m W
idth
)
0.380
0.385
0.390
0.395
0.400
0.405
0.410
0.415
0.420
Rel
ativ
e A
ge (
Tib
ial T
ooth
/Tib
ial L
engt
h)
Condition Index
Relative Age
Fig. 2 Condition index and relative age of males and females in a population of Circellium bacchus dungbeetles. The condition index was derived from pronotum width/body mass; and relative age from distaltibial tooth length/tibial length. Note the relative age scale runs in reverse to place younger individualsnearer the origin. Males were significantly older and in better condition than females. n077 males; 33females. white square 0 mean; box 0 mean ± std error; and whiskers 0 mean ± 1.96*std. error
Fig. 3 The relationship between relative testis size (testis area/body mass) and body size (pronotum width)in male Circellium bacchus dung beetles. Small males have relatively larger testes than large males. n030males
J Insect Behav
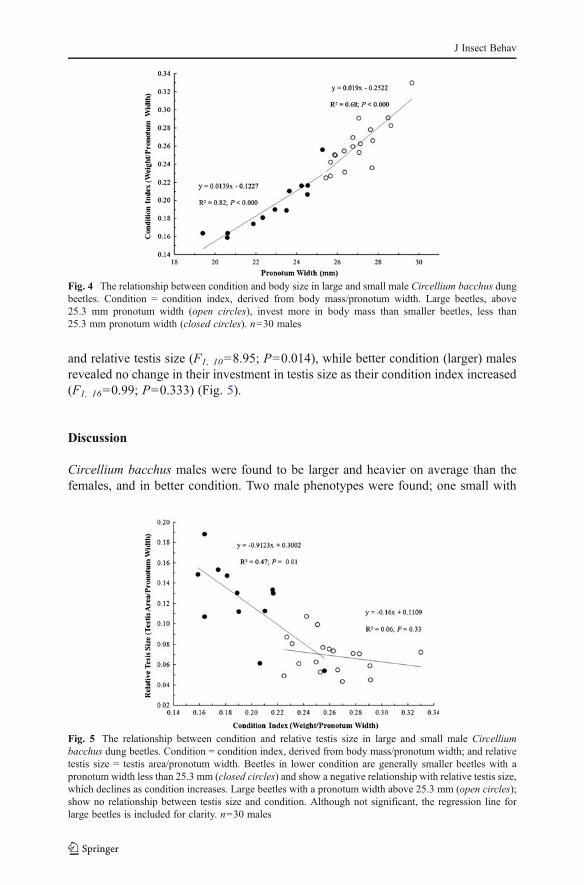
and relative testis size (F1, 1008.95; P00.014), while better condition (larger) malesrevealed no change in their investment in testis size as their condition index increased(F1, 1600.99; P00.333) (Fig. 5).
Discussion
Circellium bacchus males were found to be larger and heavier on average than thefemales, and in better condition. Two male phenotypes were found; one small with
Fig. 4 The relationship between condition and body size in large and small male Circellium bacchus dungbeetles. Condition 0 condition index, derived from body mass/pronotum width. Large beetles, above25.3 mm pronotum width (open circles), invest more in body mass than smaller beetles, less than25.3 mm pronotum width (closed circles). n030 males
Fig. 5 The relationship between condition and relative testis size in large and small male Circelliumbacchus dung beetles. Condition 0 condition index, derived from body mass/pronotum width; and relativetestis size 0 testis area/pronotum width. Beetles in lower condition are generally smaller beetles with apronotum width less than 25.3 mm (closed circles) and show a negative relationship with relative testis size,which declines as condition increases. Large beetles with a pronotum width above 25.3 mm (open circles);show no relationship between testis size and condition. Although not significant, the regression line forlarge beetles is included for clarity. n030 males
J Insect Behav

relatively large testis, and one large which instead had a higher condition index.These body mass results contrast with those of Kryger et al. (2006) where femalebeetles were recorded as being slightly heavier than their male conspecifics, suggest-ing that the population sampled in the current study contained immature femalesundergoing maturation feeding. This conclusion is supported by females also beingsignificantly younger and in poorer condition than their male counterparts.
The male-biased sex ratio seen in this C. bacchus population implies that compe-tition between males for females will be high, especially if most mature females areunderground performing brood care. Kryger et al. (2006) reported an equally oppositefemale-biased sex ratio, which appears to have been a transcription error, as both LeRoux et al. (2008) and Scholtz (2009) discuss the Kryger results as an example of amale-biased population. A male bias is expected to increase as the breeding seasonprogresses, when increasing numbers of females will be confined underground withtheir brood. October, when this study was conducted, is the period of peak activity forC. bacchus, particularly after periods of rain (Kryger et al. 2006).
Circellium bacchus are sexually dimorphic in that male beetles are significantlylarger on average than females (Kryger et al. 2006). Le Roux et al. (2008) suggestedthat size might be used by females to select mates. Male-biased sexual size dimor-phism is indicative of breeding systems mediated by male-male contest competition(Trivers 1972; Andersson 1994). These systems are characterized by aggressiveencounters between males, with body size reliably predicting the outcome of suchcontests and larger beetles winning significantly more fights (Lawrence 1986; LeRoux et al. 2008). Moreover, the operational sex ratio (the ratio of sexually activemales to receptive females) is likely to further increase competition for mates, assexually mature females become rarer within the population.
Emlen and Oring (1977) proposed that the operational sex ratio determines theintensity of competition for mates. Given that the male De Hoop C. bacchuspopulation was found to be more than double that of the female population (in thisstudy and two prior studies [Kryger et al. 2006 and Le Roux et al. 2008]) and thatfemale beetles were significantly younger and in poorer condition than their maleconspecifics, access to receptive females will be substantially lower than what wasmeasured and would consequently further intensify the extent of male-male compe-tition. No breeding pairs were noted in the limited observations that were made.
The likelihood of strong pre-copulatory competition for mates between C. bacchusmales predicts evolution of an alternative breeding tactics that would allow smaller,less physically competitive males to gain access to females (Emlen and Oring 1977).The break-point at a 25.3 mm pronotum width suggests that smaller C. bacchus“minor” males should adopt the alternate behavior of sneaking copulations. Thisbehavior was observed once by Le Roux et al. (2008), where a smaller male C.bacchus was buried with the ball by the female and successfully copulated with her,whilst larger males were fighting each other. Sneaks, however, are almost alwayssubject to post-copulatory sperm competition (Simmons et al. 2007). In response,Parker (1970) showed that the probability of fertilization success should increase withthe quantity of sperm a male could deposit into the female’s reproductive tract.Consequently, such sperm competition ought to favor increased sperm production,presumably from relatively larger testes (Simmons and García-Conzález 2008). Theseresults show smaller C. bacchus males have increased investment in testes size
J Insect Behav

relative to larger males, presumably to improve the possibility of gainingfertilization post-copulation using an alternative reproductive tactic. Largermales invest resources in body weight (presumably muscle), increasing theircondition index score, without diverting any of those resources to testes size,which appears to remain static even as condition improves. Nevertheless,manipulative experiments are required to test this conclusion. Knell (2011)reports that weight gain correlates with horn size in Euoniticellus intermedius(Reiche) (Scarabaeidae: Scarabaeinae), a typical tunneling dung beetle which exhibitstwo male phenotypes, horned and hornless, which is probably dependent on the sizeof the third instar larva (Pomfret and Knell 2006; Emlen and Nijhout 2001). If large(>25.3 mm pronotum width) C. bacchus males are equivalent to the large majormales of horned species, then weight gain for larger males would be expected as bodymass is the best predictor of victory in male fights (Le Roux et al. 2008). On the otherhand, C. bacchus males in the poorest condition had the largest testes relative to theirbody weight. If these minor males do employ sneak mating tactics they areunlikely to require strength for mating success, whereas the outcome of contestsbetween major males are probably heavily influenced by strength. Such “con-dition dependent” trait expression has been found in other horned beetles (Knelland Simmons 2010).
Testes size is determined by the proportion of resources allocated to this tissueduring the larval stage of development (Simmons et al. 2007; Carsten-Conner et al.2010). It would thus appear that type of reproductive tactic the male adult beetle willemploy must be determined during larval development. Because nutritional resourcesprovided by the brood ball are finite (Moczek 2011), resources allocated to testesmust be traded-off against those which could be allocated to the development of someother organ or structure. For example in Onthophagus spp. (Scarabaeidae: Scarabaei-nae) the growth of horns is known to come at the expense of the development of otherstructures such as eyes or antennae (Emlen 2001; Emlen et al. 2005). In O. taurusSchreber larvae, ablation of precursor cells in the larva, which are destined to becomethe testes in the adult, causes the growth of larger horns (Moczek and Nijhout 2004),while in O. nigriventris D’Orbigny male larvae with ablated horn precursors grewlarger and had larger testes (Simmons and Emlen 2006). Smaller C. bacchus malesmay also face a resource allocation trade-off post-emergence whereby food resourceswhich could be directed towards weight or organ maintenance in larger individualsare used instead for larger ejaculates.
The evolution of large testes in response to male-male contest competition hasbeen observed in multiple taxa (Simmons et al. 2007; Simmons and Emlen 2006;Longair 2004; Byrne et al. 2002; Birkhead 1998) and now includes C. bacchus whichimplies an alternative reproductive tactic. In addition to evidence for sperm compe-tition, C. bacchus demonstrates alternative male phenotypes suggesting that thiscould occur in other ball rolling dung beetles where males have difficulty monopo-lizing access to females in above-ground courtship encounters with many males.Favilla et al. (2005) found that female Canthon cyanellus cyanellus LeConte (Scar-abaeinae; Canthonini) were receptive during the underground ball-making period,when sneaking males may have a better chance of gaining the attentions of a female.
Female mate choice in C. bacchus remains unexplored, and because of its longlarval developmental period and low fecundity, seems destined to remain that way.
J Insect Behav

Nevertheless, male quality is unlikely to be assessed by the female solely in terms ofbody size, which has a low heritability because it is dictated by the brood ball size.Nevertheless, this preference will be maintained as females of other species have beenfound to produce more offspring when paired with large males (Cook, 1988).Canthon c. cyanellus males achieve sperm precedence by repeated matings whichlead to sperm replacement (Favilla et al. 2005) and favor the evolution of large testeswhen competition occurs between the ejaculates of two or more males. In addition,Simmons and Kotiaho (2002) found testis weight and ejaculate volume contributed tomale fertilization success in O. taurus, and that these were condition dependent -condition being defined as the pool of resources available for production or mainte-nance of traits that enhance fitness (Rowe and Houle 1996). Interestingly, male C.bacchus show a negative or neutral relationship between overall body condition (size/weight) and testes size, which would be expected to moderate rather than reinforcethe selection pressures on the opposing forces of pre-mating female preference for largemales versus the post-copulatory advantage of small males with larger testes. Never-theless, the selective advantage of either male phenotype will change through the courseof several, often unpredictable, seasons for these long-lived beetles, and may includecryptic female choice at some stage. Canthon c. cyanellus males also produce largespermatophores and an anti-fungal agent which protects the brood ball (Favilla et al.2005), so male contributions may extend beyond just self-serving resources.
Tunneling dung beetles with horned males have made major contributions to ourknowledge about the systems and evolutionary outcomes of sexual selection inanimals. Further examination of the circumstances which govern the adoption andadvantages of alternative reproductive strategies in C. bacchus can broaden ourunderstanding of such mechanisms of sexual selection for relatively long-livedanimals in variable environments.
Acknowledgements This research formed a part of work undertaken by the Organization for TropicalStudies (OTS) South Africa Program Fall 2011. We would like to thank OTS for this opportunity and aregreatly appreciative of the staff and students who assisted in data collection. In particular we would like tothank Melissa Manus, Gabriella Chateau, and Natasha Kohanzadeh for their invaluable help with thedissections and preliminary data analysis. We would also like to thank the two anonymous reviewers whocommented on the manuscript and whose advice helped to improve our manuscript.
References
Andersson M (1994) Sexual selection. Princeton University Press, PrincetonBirkhead TR (1998) Sperm competition in birds. Rev Reprod 3:123–129Birkhead TR, Atkin L, Møller AP (1987) Copulation behavior of birds. Behavior 101:101–138Byrne PG, Roberts JD, Simmons LW (2002) Sperm competition selects for increased testes mass in
Australian frogs. Science 15:347–355Carsten-Conner LD, Papaj DR, O’Brien DM (2010) Resource allocation to testes in walnut flies and
implications for reproductive strategy. J Insect Physiol 56:1523–1539Chown SL, Scholtz CH, Klok CJ, Joubert FJ, Coles KS (1995) Ecophysiology, range contraction and survival
of a geographically restricted African dung beetle (Coleoptera: Scarabaeidae). Funct Ecol 9:30–39Cook D (1988) Sexual selection in dung beetles. II. Female fecundity as an estimate of male reproductive
success in relation to horn size, and alternative behavioural strategies in Onthophagus binodis Thun-berg (Scarabaeidae: Onthophagini). Aust J Zool 36:521–523
J Insect Behav

Darwin C (1871) The descent of man and the selection in relation to sex. John Murry, LondonDeutsch JC (1997) Color diversification in Malawi cichlids: evidence for adaptation, reinforcement or
sexual selection? Biol J Linn Soc 62:1–14Diamond JM (1986) Biology of birds of paradise and bowerbirds. Annu Rev Ecol Syst 17:17–37Eberhard WD, Huber BA, Rodriguez RL, Briceño RD, Salas I, Rodriquez V (1998) One size fits all?
Relationships between the size and degree of variation in genetalia and other body parts in twentyspecies of insects and spiders. Evolution 52:415–431
Eberhard WD, Huber BA, Rodriguez RL (1999) Don’t forget the biology: a reply to Green. Evolution53:1624–1627
Emlen DJ (1997) Alternative reproductive tactics and male-dimorphism in the horned beetle Onthophagusacuminatus (Coleoptera: Scarabaeidae). Behav Ecol Sociobiol 41:335–341
Emlen DJ (2001) Costs and the diversification of exaggerated animal structures. Science 291:1534–1536
Emlen DJ (2008) The evolution of animal weapons. Annu Rev Ecol Evol Syst 39:387–413Emlen DJ, Nijhout HF (2001) Hormonal control of male horn length dimorphism in Onthophagus taurus
(Coleoptera: Scarabaeidae): a second critical period of sensitivity to juvenile hormone. J Insect Physiol47:1045–1054
Emlen ST, Oring LW (1977) Ecology, sexual selection, and the evolution of mating systems. Science197:215–223
Emlen DJ, Marangelo J, Ball B, Cunningham CW (2005) Diversity in the weapons of sexual selection:Horn evolution in the beetle genus Onthophagus (Coleoptera: Scarabaeidae). Evolution 59:1060–1084
Fabricius JC (1781) Species Insectorum, Vol 2. Hamburg, KilioniFavilla MF, Nolasco J, Florescano IC, Equihua M (2005) Sperm competition and evidence of sperm
fertilization patters in the carrion ball-roller beetle Canthon cyanellus cyanellus LeConte (Scarabaei-dae: Scarabaeinae). Behav Ecol Sociobiol 59:38–43
Flower J, Cohen L, Jarvis P (1998) Practical statistics for field biology. Wiley, West SussexGage MJG (1995) Associations between body size, mating pattern, testis size and sperm lengths across
butterflies. Proc R Soc Lond B Biol Sci 258:247–254Gilliard ET (1969) Birds of paradise and bower birds. Weidenfeld and Nicholson, LondonGreen AJ (1999) Allometry of genetalia in insects: one size does not fit all. Evolution 53:1621–1624Harcourt AH, Purvis A, Liles L (1995) Sperm competition: mating system, not breeding season, affects
testes size of primates. Funct Ecol 9:468–476Knell R (2011) Male contest competition and the evolution of weapons. In: Simmons LW, Ridsill-
Smith TJ (eds) Ecology and evolution of dung beetles. Blackwell Publishing Ltd, London, pp47–66
Knell RJ, Simmons LW (2010) Mating tactics determine patterns of condition dependence in a dimorphichorned beetle. Proc R Soc B 277:2347–2353
Kruuk LEB, Slate J, Pemberton JM, Brotherstone S, Guinness F, Clutton-Brock T (2002) Antler size in reddeer: heritability and selection but no evolution. Evolution 56:1683–1695
Kryger U, Cole KS, Tukker R, Scholtz CH (2006) Biology and ecology of Circellium bacchus (Fabricius1781) (Coleoptera Scarabaeidae), a South African dung beetle of conservation concern. Trop Zool19:187–207
Lawrence WS (1986) Male choice and competition in Tetraopes tetraophthalmus: effects of local sex rationvariation. Behav Ecol Sociobiol 18:289–296
Le Roux E, Schlotz C, Kinahan H, Bateman PW (2008) Pre and Post-copulatory mate selection mecha-nisms in an African dung beetle, Circellium bacchus (Coleoptera: Scarabaeidae). J Insect Behav21:111–122
Longair RW (2004) Tusked males, male dimorphism and nesting behavior in a subsocial Afrotropical wasp,Synagris cornuta, and weapons and dimorphism in the Genus (Hymenoptera/: Vespidae/: Eumeninae).J Kans Entomol Soc 77:528–557
Moczek A (2011) Evolution and development: onthophagus beetles and the evolutionary developmentalgenetics of innovation, allometry and plasticity. In: Simmons LW, Ridsill-Smith TJ (eds) Ecology andevolution of dung beetles. Blackwell Publishing Ltd, London, pp 126–151
Moczek AP, Nijhout HF (2004) Trade-offs during the development of primary and secondary sexual traitsin a horn dimorphic beetle. Am Nat 163:184–191
Møller AP (1991) Sperm competition, sperm depletion, paternal care, and relative testis size in birds. AmNat 137:882–906
Parker GA (1970) Sperm competition and its evolutionary consequences in the insects. Biol Rev 45:525–567
J Insect Behav

Parker GA (1990) Sperm competition games: sneaks and extra-pair copulations. Proc R Soc Long B BiolSci 22:127–133
Parker GA (1998) Sperm competition and the evolution of ejaculates: towards a theory base. In: BirkheadTR, Moller AP (eds) Sperm competition and sexual selection. Academic Press, California, pp 3–54
Pitcher TE, Dunn PO, Whittingham LA (2005) Sperm competition and the evolution of testes size in birds.J Evol Biol 18:557–567
Pomfret JC, Knell RJ (2006) Sexual selection and horn allometry in the dung beetle Euoniticellusintermedius. Anim Behav 71:567–576
Rasmussen JL (1994) The influence of horn and body size on the reproductive behavior of the hornedrainbow scarab beetle Phanaeus difformis (Coleoptera: scarabaeidae). J Insect Behav 7:67–82
Rebelo T (1998) Limestone Fynbos. In: Low AB, Rebelo AG (eds) Vegetation of South Africa, Lesothoand Swaziland. Department of Environmental Affairs and Tourism, Pretoria
Rowe L, Houle D (1996) The lek paradox and the capture of genetic variance by condition dependent traits.Proc R Soc B 263:1415–1421
Sato H, Hiramatsu K (1993) Mating behaviour and sexual selection in the African ball-rolling scarabKhepher platynotus (Bates) (Coleoptera: Scarabaeidae). J Nat Hist 27:657–668
Scholtz CH (2009) Pre-copulatory behavior, sexual attraction, mate selection and co- operative breeding.In: Scholtz CH, Davis ALV, Kryger U (eds) Evolutionary biology and conservation of dung beetles.Pensoft Publishers, Sofia, pp 62–68
Simmons LW (2011) Sexual selection after mating: the evolutionary consequences of sperm competitionand cryptic female choice in Onthophagines. In: Simmons LW, Ridsill-Smith TJ (eds) Ecology andevolution of dung beetles. Blackwell Publishing Ltd, London, pp 66–86
Simmons LW, Emlen DJ (2006) Evolutionary trade-off between weapons and testes. Proc Nat Acad SciUSA 103:16346–16351
Simmons LW, García-Conzález F (2008) Evolutionary reduction in testes size and competitive fertilizationsuccess in response to the experimental removal of sexual selection in dung beetles. Evolution62:2580–2591
Simmons LW, Kotiaho JS (2002) Evolution of ejaculates: patterns of phenotypic and genotypic variationand condition dependence in sperm competition traits. Evolution 56:1622–1631
Simmons LW, Kotiaho JS (2007) The effects of reproduction on courtship, fertility and longevity withinand between alternative male mating tactics of the horned beetle Onthophagus binodis. Proc Nat AcadSci USA 104:16604–16608
Simmons LW, Ridsill-Smith TJ (eds) (2011) Ecology and evolution of dung beetles. Blackwell PublishingLtd, London
Simmons LW, Emlen DJ, Tomkins JL (2007) Sperm competition games between sneaks and guards: acomparative analysis using dimorphic male beetles. Evolution 61:2684–2692
Statistica Version 8.0. (2006). Statsoft Inc., Tulsa, USATrivers RL (1972) Parental investment and sexual selection. In: Campbell B (ed) Sexual selection and the
descent of man. Aldine, Chicago, pp 1871–1971Turner GF (1994) Speciation mechanisms in Lake Malawi cichlids: a critical review. Ergeb Limnol 44:139–
160Tynedale-Biscoe M (1978) Physiological age-grading in females of the dung beetle Euoniticellus inter-
medius (Reiche) (Coleoptera; Scarabaeidae). Bull Ent Res 68:207–217Vahed K, Parker DJ (2011) The evolution of large testes: sperm competition or male mating rate? Ethology
118:107–117van Oppen MJH, Turner GF, Rico C, Robinson RL, Deutsh JC, Genner M, Hewitt GM (1998) Assortative
mating among rock dwelling cichlid fishes supports high estimates of species richness from LakeMalawi. Mol Ecol 7:991–1001
Watson NL, Simmons LW (2010) Reproductive competition promotes the evolution of female weaponry.Proc R Soc B 277:2035–2040
J Insect Behav




















