
Diel separation of Afrotropical dung beetle guilds - avoiding competition and neglecting resources
25
Diel separation of Afrotropical dung beetle guilds—avoiding competition and neglecting resources (Coleoptera: Scarabaeoidea) SYLVIA KRELL-WESTERWALBESLOH{{, FRANK-THORSTEN KRELL{* and K. EDUARD LINSENMAIR{ {Lehrstuhl fu ¨r Tiero ¨kologie und Tropenbiologie, Biozentrum, Universita ¨t Wu ¨rzburg, Am Hubland, D-97074 Wu ¨rzburg, Germany {Soil Biodiversity Programme, Department of Entomology, The Natural History Museum, Cromwell Road, London SW7 5BD, UK (Accepted 15 July 2003) Dung beetle guilds of different competitive level coexist at high abundances in the forest–savanna mosaic of Ivory Coast. A total of 25 875 specimens was recorded from 90 samples of fresh buffalo dung exposed for 4-h periods over the day in the Parc National de la Comoe ´. Redundancy analyses show different patterns of the guild structure determined by time of day, and positive correlations of guild abundance with temperature. The competitively superior telecoprids (rollers) have their abundance peak at midday when the high temperatures presumably enable them to perform their energetically costly rolling behaviour at greater speed. The competitively intermediate paracoprids (tunnellers) and the competitively inferior endocoprids (dwellers) have their peak around dusk when: (1) the superior competitors are not active, and (2) they can easily synchronize their flight activity using light intensity changes. During the two peaks of flight activity of the dung beetles, their abundance at the resource is very high, causing obvious competition. On the other hand, the resource is hardly used between 22:00 h and 10:00 h. This is probably due to physiological constraints (high optimum temperature required for the diurnal species and dependence on light intensity patterns as a flight trigger of the nocturnal species, respectively). Shifting flight activity to these periods of low competition does not occur, resulting in a unequal level of competition over the day, thus periods of considerable competition within dung beetle assemblages alternate with vacant temporal refuges. KEYWORDS: Coleoptera, Comoe ´ National Park, Co ˆ te d’Ivoire, diel separation, dung beetles, Guinea savanna, temporal resource partitioning, Scarabaeidae. Journal of Natural History ISSN 0022-2933 print/ISSN 1464-5262 online # 2004 Taylor & Francis Ltd http://www.tandf.co.uk/journals DOI: 10.1080/00222930310001618921 *Author to whom correspondence is addressed; e-mail: [email protected] JOURNAL OF NATURAL HISTORY, 2004, 38, 2225–2249
Transcript of Diel separation of Afrotropical dung beetle guilds - avoiding competition and neglecting resources

Diel separation of Afrotropical dung beetle guilds—avoidingcompetition and neglecting resources (Coleoptera: Scarabaeoidea)
SYLVIA KRELL-WESTERWALBESLOH{{, FRANK-THORSTEN
KRELL{* and K. EDUARD LINSENMAIR{
{Lehrstuhl fur Tierokologie und Tropenbiologie, Biozentrum, Universitat
Wurzburg, Am Hubland, D-97074 Wurzburg, Germany
{Soil Biodiversity Programme, Department of Entomology, The Natural
History Museum, Cromwell Road, London SW7 5BD, UK
(Accepted 15 July 2003)
Dung beetle guilds of different competitive level coexist at high abundances inthe forest–savanna mosaic of Ivory Coast. A total of 25 875 specimens wasrecorded from 90 samples of fresh buffalo dung exposed for 4-h periods over theday in the Parc National de la Comoe. Redundancy analyses show differentpatterns of the guild structure determined by time of day, and positivecorrelations of guild abundance with temperature. The competitively superiortelecoprids (rollers) have their abundance peak at midday when the hightemperatures presumably enable them to perform their energetically costlyrolling behaviour at greater speed. The competitively intermediate paracoprids(tunnellers) and the competitively inferior endocoprids (dwellers) have their peakaround dusk when: (1) the superior competitors are not active, and (2) they caneasily synchronize their flight activity using light intensity changes. During thetwo peaks of flight activity of the dung beetles, their abundance at the resource isvery high, causing obvious competition. On the other hand, the resource ishardly used between 22:00 h and 10:00 h. This is probably due to physiologicalconstraints (high optimum temperature required for the diurnal species anddependence on light intensity patterns as a flight trigger of the nocturnal species,respectively). Shifting flight activity to these periods of low competition does notoccur, resulting in a unequal level of competition over the day, thus periods ofconsiderable competition within dung beetle assemblages alternate with vacanttemporal refuges.
KEYWORDS: Coleoptera, Comoe National Park, Cote d’Ivoire, diel separation,dung beetles, Guinea savanna, temporal resource partitioning, Scarabaeidae.
Journal of Natural HistoryISSN 0022-2933 print/ISSN 1464-5262 online # 2004 Taylor & Francis Ltd
http://www.tandf.co.uk/journalsDOI: 10.1080/00222930310001618921
*Author to whom correspondence is addressed; e-mail: [email protected]
JOURNAL OF NATURAL HISTORY, 2004, 38, 2225–2249

Introduction
‘‘Quel est celui-ci qui trottine vers le monceau, craignant d’arriver trop tard?’’
(Jean-Henri Fabre, Le Scarabee Sacre, 1879)
If a resource is an unpredictable, ephemeral patch, it is crucial for the success of
any potential user to be in the right place at the right time. This is the more so if
the ephemeral nature of the resource is caused by the users themselves, as in rapid
degradative successions (Begon et al., 1996: 693). Interspecific competition is
inevitable on ephemeral resource patches that are used by different species in high
abundance. Dung pats in humid Afrotropical savannas during the rainy season
are a good model system with which to study strategies of exploiting ephemeral
resource patches. The exact time and place of the appearance of faeces is
unpredictable, in natural habitats they are patchily distributed. Furthermore, they
are used up by dung beetles mostly within less than 24 h. The succession of dung
beetle assemblages in temperate regions (at least in large droppings) generally
follows the facilitation model of Connell and Slayter (1977) (Beaver, 1984; Hanski,
1991): resource exploitation by later users is facilitated by the action of the
first users. In contrast, in humid African savannas the early occupants of a dung
pat tend to use the resource more or less completely (figures 1, 2). Hence, the
succession seems to follow the inhibition model (Connell and Slayter, 1977): the
action of first users inhibits exploitation by later arrivals, because after the first
users have exploited the substrate, only a small fraction is left, which is relatively
unproductive for any later arriving beetles. Possible successful strategies in such a
FIG. 1. Typical buffalo dung pat exposed during the day well before dusk. Users are mostlytelecoprids.
2226 S. Krell-Westerwalbesloh et al.

system are either to be competitively superior (effectively monopolizing parts of the
resource) or to avoid competition (using the resource when superior competitors are
not active).
Dung beetles
Coprophagous scarab beetles (Scarabaeidae) use their resource in different
ways, through which they can be classified into guilds (Bornemissza, 1969;
Cambefort and Hanski, 1991). The guilds show a dominance hierarchy in the
competitive ability of their species (Doube, 1990). Species of the roller guild
(telecoprids) rapidly form balls from the faeces, roll them away from the food
source, and deposit them in or on the soil to ensure the exclusive use of this part
of the resource. Formation of a ball generally takes much less than 1 h, and in the
small Sisyphini only a few minutes (Halffter and Matthews, 1966: 103; Doube,
1990; personal observation). The tunnellers (paracoprids) make nests directly under
the food source and transport dung into the nest where they form dung balls. This
is relatively time-consuming and takes at least a few hours (Doube, 1990). Dwellers
(endocoprids) feed and reproduce directly in the dung pat. Kleptoparasites use
faeces portions monopolized by species of telecoprids or paracoprids. They
penetrate dung balls made by telecoprids, or the dung mass in the subterranean
nests of paracoprids.
If the beetles of all guilds are abundant, these modes of resource utilization are
mutually exclusive: when the dung is rolled away by rapid telecoprids (figure 1),
FIG. 2. Typical buffalo dung pat exposed just before dusk or early at night. Users aremostly paracoprids. The big soil mounds were produced by Coprini (large paracoprids).
Diel separation of dung beetle guilds 2227

paracoprids do not have enough left to bury. When the dung is rolled away or
buried (figure 2), the endocoprids have nothing to lay their eggs in. The com-
petitively superior species are, therefore, the fast telecoprids, the least competitive
species are the endocoprids (Doube, 1990). The success of kleptoparasites depends
on the success of their hosts. Dung relocation (rolling and tunnelling) is burdened
with high energetic costs whereas endocoprid behaviour and kleptoparasitism is
much less energetically costly.
Because dung usually persists for less than 24 h as a usable resource in
the humid African savannas during the rainy season, success of any species is
determined mostly by its early arrival at the resource (Doube, 1987; Hanski, 1989:
‘extreme priority effect’), especially because telecoprid and paracoprid species are
able to monopolize a part of the resource. Therefore, the diel activity of the species
is an important parameter determining their success (Hanski, 1990). The rapidity of
colonization of a freshly emerged resource is enormous. Anderson and Coe (1974)
found that 16 000 beetles had completely removed a 1.5 kg pile of elephant faeces
within 2 h in the Tsavo National Park, Kenya. A normal portion of human faeces
is generally removed completely after 3 h in our study area (Parc National de la
Comoe, Cote d’Ivoire) if it has been deposited during the day in the savanna
parkland (personal observation). Despite this strong competition between rapid,
abundant users, species of different competitive potency occur syntopically in
African savanna parklands.
Did competitively inferior guilds evolve mechanisms for avoiding competition
or do they depend on random success to coexist with superior competitors?
Temporal resource partitioning
Although temporal resource partitioning is considered to be less common
compared to spatial separation and trophic specialization (Schoener, 1986), it is
a widespread mechanism to avoid competition between closely related species or
phylogenetically distant groups. The most common type of temporal separation is a
different phenology, especially seasonally different activity times in using a resource
(Wolda, 1988). Diel resource partitioning within animal assemblages, however, has
rarely been analysed (Remmert, 1976; Southwood, 1978; Loser, 1980; Dennison
and Hodkinson, 1983; Jaksic, 1983). Diel resource partitioning within dung beetle
assemblages has been studied a few times at species level (e.g. Caveney et al., 1995:
paracoprid Onitis spp.; Hanski, 1986: endocoprid Aphodius spp.; Walter, 1985:
not quantitatively) or at genus level (e.g. Peck and Forsyth, 1982: dung beetle
assemblages in Ecuador). Diel separation between guilds has only once been
quantitatively examined (Doube, 1991), but in those assemblages, endocoprids were
extremely rare.
In a parallel study (Krell et al., 2003), we found that guild structures of beetle
assemblages in buffalo dung differ between day and night. We also found that the
abundance (and biomass, unpublished results) of dung beetles is at an obviously
non-competitive level all day and night in the gallery forest and during the day in
the grassland of the river valley. In the savanna parkland, however, the number of
dung beetles seems to be very high at night and during the day. Does competition
always control the structure of coprocenoses (animal assemblages in dung) in the
savanna habitat or are our former results artefacts of low temporal resolution?
In the present study we increased resolution by studying the dung beetle
2228 S. Krell-Westerwalbesloh et al.

assemblages that have populated dung portions during defined 4-h periods to
determine the diel patterns of flight activity and resource partitioning of the
different dung beetle guilds.
Having analysed pooled data from different habitats, Krell et al. (2003) found
that the abundance of telecoprids and their kleptoparasites is positively correlated
with the temperature of faeces and soil, whereas the number of endocoprids
increases with decreasing temperature during the exposure period. Thus, the
competitively most inferior group seems to be tolerant to unfavourable climatic
conditions. The higher temporal resolution of the present study will show whether
these climatic parameters influence the abundance of guilds in the savanna or
whether they are irrelevant in influencing guild structure within one habitat.
Study area
We conducted our experiments in the southern part of the Parc National de la
Comoe in north-eastern Ivory Coast (~Cote d’Ivoire, West Africa) around the
research camp of the University of Wurzburg (Lola-Camp; see Krell et al., 2003:
figure 1). The study area is at the border between the Guinea and Subsudan
savanna attributed by Porembski (1991) to the former and by Poilecot (1991),
amongst others, to the latter. Savanna parkland takes up the largest area, followed
by forest islands, gallery forest and minor habitat types. Since in the savanna
parkland the abundance of dung beetles is high both at night and during the day
(Krell et al., 2003), we studied diel activity patterns on a site in this habitat. It is the
same savanna site used for the study of habitat patterns by Krell et al. (2003:
figure 4; 3‡48’58@W, 8‡45’02@N).
Methods
Experiments
From 7 May to 12 July 1997 (beginning of the rainy season to the beginning
of the ‘Little Dry Season’ in the middle of the rainy season; Ojo, 1977: 97) we
deposited portions of 1 kg (900 ml) of faeces of buffalo (Syncerus caffer (Sparrman))
on the soil during 4-h periods starting 2:00 h, 6:00 h, 10:00 h, 14:00 h, 18:00 h
and 22:00 h (15 samples each; see Appendix). The exposure periods were spread
randomly over the study period with two to four dung portions exposed
simultaneously. After the exposure period, the faeces, and soil beneath that had
been populated by coprophagous beetles, were floated in a bucket of water. In
addition, we searched the complete sediment in the bucket for remaining beetles. A
detailed description and discussion of the method is given by Krell et al. (2003).
We did not use traps because we wanted to record established coprocenoses after a
fixed period of exposure of the resource, not all of the beetles visiting the resource
during a period of time. Some beetles may approach the dung pat but fly away after
a short exploration (personal observations) or when the pat is already highly
populated (Landin, 1961: 207). With our method we register only the actual users
of a resource whereas traps collect both users and tourists. Traps overestimate
the portion of some groups containing many tourists. These groups cannot be
unambiguously identified among the trapped beetles. However, with our method we
underestimate the proportion of telecoprids in the coprocenoses since many of them
have already left the pat before we collected the sample. Although we obviously got
Diel separation of dung beetle guilds 2229

a biased pattern, we are able to identify the bias, whereas in pitfall trap sampling
the overestimated groups are not recognizable.
The abundance of scarab beetles increases strongly after heavy rainfall (Walter,
1985; personal observation). During a period of heavy rain (11–23 June) we did not
expose any dung (see Appendix).
The collected material (25 875 specimens) will be incorporated into the
collection of The Natural History Museum, London. The specimens were identified
by F.-T.K.
Microclimatic change during the day
To find possible correlations of guild abundances with microclimatic parameters
we measured:
(1) air temperature at about 1 m above ground at the beginning and end of the
exposure period;(2) soil temperature at a depth of 1 and 10 cm at the beginning and end of the
exposure period;(3) dung temperature just before collecting the sample (1–3: microcomputer
thermocouple thermometer HI 9063, Hanna Instruments);(4) relative air humidity (RH) at about 1 m above the ground at start and end
of the exposure period (electronic hygrometer HI 9064, Hanna Instruments).
Since the instrument is reliable up to 95% RH, all measurements above 95%
are counted as 95%. This reduces the arithmetic means of RH during the
night periods (table 3), which is certainly closer to 100%.
Guild classification
We treat each species as belonging to exactly one guild. The term ‘guild’ is used
as in its original definition as a group of species which use the same resource in
a similar way regardless of their phylogenetic relationships (cf. Simberloff and
Dayan, 1991). However, we confine this analysis to the dung beetles of the family
Scarabaeidae which are by far the major users of the dung resource. Other
coprophilous beetles (Histeridae, Hydrophilidae, Carabidae, Staphylinidae) are
either predators, occur in relatively low numbers in the studied assemblages or use
the resource in different ways. We follow the rough classification of coprophagous
members of the coprocenoses into the guilds of telecoprids (rollers), paracoprids
(tunnellers), endocoprids (dwellers) and kleptoparasites (Bornemissza, 1969;
Cambefort and Hanski, 1991) but we divide the last group into kleptoparasites
of paracoprids and those of telecoprids (Cambefort, 1991). As usual, there is no
simple correlation between higher taxa and guilds. In the studied assemblages, the
guilds consist of the following members (for sources and more detailed discussion
of the guild classification see Krell et al., 2003):
. telecoprids (rollers): all Gymnopleurini, all Sisyphini, all Canthonini (only
Anachalcos spp.) (Scarabaeini were not present and are generally very rare in
the study area; personal observation);. paracoprids (tunnellers): all Coprini (including Dichotomiini), all Onitini,
most Oniticellini, most Onthophagini, Aphodius (Neocolobopterus) maculi-
collis Reiche;. endocoprids (dwellers): all Aphodiinae (except Aphodius maculicollis),
Oniticellus formosus Chevrolat;
2230 S. Krell-Westerwalbesloh et al.

. obligatory kleptoparasites:(a) of paracoprids: Pedaria spp., Onthophagus juvencus Klug;
(b) of telecoprids: Cleptocaccobius spp., Hyalonthophagus nigroviolaceus
(d’Orbigny), Onthophagus lioides d’Orbigny, O. tersipennis d’Orbigny.
Statistics
For the statistical analyses we used the computer programs Canoco for
Windows 4.0 (Centre for Biometry, Wageningen, The Netherlands), SsS 1.1a
(Rubisoft Software GmbH, Eichenau, Germany) and XLSTAT 4.4 (Thierry
Fahmy, Paris, France).
Correlations between the abundance of guilds are shown by a standardized
principal component analysis (PCA) (Canoco) using log-transformed guild
abundance data. PCA requires a normal distribution for each guild which is not
present in all our data, even when log-transformed (telecoprids and both groups
of kleptoparasites; Kolmogoroff-Smirnoff test, SsS), but for merely descriptive
purposes, larger departures from ideal data structure are tolerable (Greig-Smith,
1980; Gauch, 1982: 137). Significance of the correlations was tested with the non-
parametric Spearman Rank Correlation Coefficient (XLSTAT) using the critical
values given by Zar (1996).
A standardized redundancy analysis (RDA) (Canoco; Jongman et al., 1995;
Legendre and Legendre, 1998) using log-transformed guild abundance data shows
the relationship between environmental parameters and guild abundances. The
highest increase of temperature during the exposure period was re-scaled to 0; the
highest decrease becomes the highest value.
Results
Diel differences in abundance and guild structure of coprocenoses
Both the ratios of dung beetle guilds and the overall abundance of dung beetles
was extremely unequally distributed over the day. The two peaks of overall
abundance were at midday (48%; 829 individuals per kg dung) and around dusk
(25%; 429 ind. per kg dung) (figure 3; table 1). Since we underestimate the number
of telecoprids (Krell et al., 2003), the real peak at midday is higher than shown in
figure 3 and, hence, much higher than the peak at dusk. It is even possible that there
is no local minimum at 14:00–18:00 h. The dung beetles have their lowest flight
activity during the night until dawn. Less than 4.5% of all dung beetles in our
samples flew between 22:00 h and 6:00 h (38 ind. per kg dung). The guild structure
of dung beetle assemblages over the day is given in table 1.
Diel separation and correlation of guilds
The PCA (figure 4) shows graphically the distribution of samples in a two-
dimensional space considering the highest possible proportion of overall variability.
Day and night samples are clearly separated with only a slight overlap by samples
from the period just after dawn. All samples from the periods of highest flight
activity [around dusk (18:00–22:00 h) and midday (10:00–14:00 h)] are isolated with
only a slight overlap of the latter with other daytime periods. The 6:00–10:00 h and
14:00–18:00 h samples and the 22:00–2:00 h and 2:00–6:00 h samples are mixed,
respectively.
Diel separation of dung beetle guilds 2231
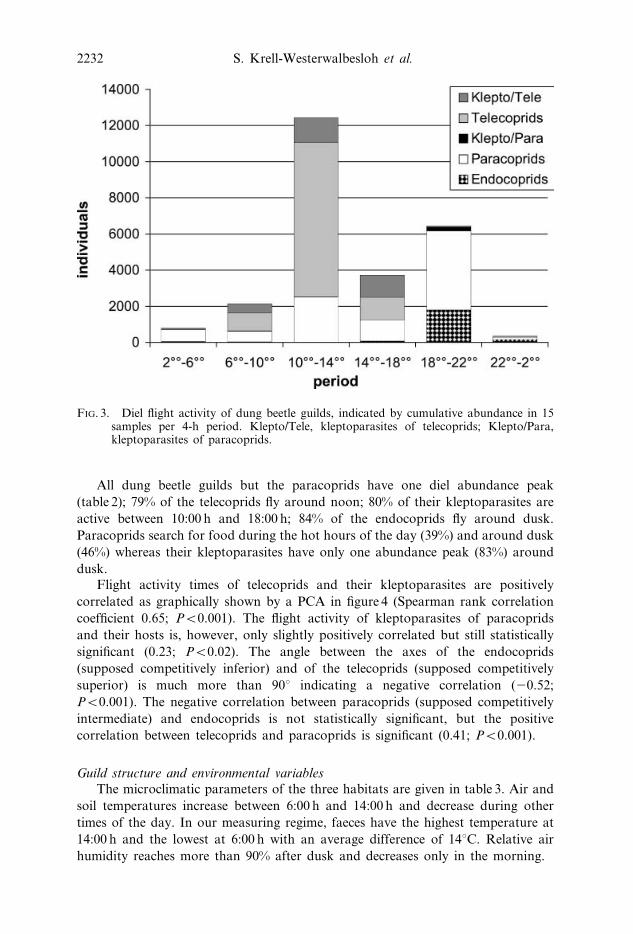
All dung beetle guilds but the paracoprids have one diel abundance peak
(table 2); 79% of the telecoprids fly around noon; 80% of their kleptoparasites are
active between 10:00 h and 18:00 h; 84% of the endocoprids fly around dusk.
Paracoprids search for food during the hot hours of the day (39%) and around dusk
(46%) whereas their kleptoparasites have only one abundance peak (83%) around
dusk.
Flight activity times of telecoprids and their kleptoparasites are positively
correlated as graphically shown by a PCA in figure 4 (Spearman rank correlation
coefficient 0.65; Pv0.001). The flight activity of kleptoparasites of paracoprids
and their hosts is, however, only slightly positively correlated but still statistically
significant (0.23; Pv0.02). The angle between the axes of the endocoprids
(supposed competitively inferior) and of the telecoprids (supposed competitively
superior) is much more than 90‡ indicating a negative correlation (20.52;
Pv0.001). The negative correlation between paracoprids (supposed competitively
intermediate) and endocoprids is not statistically significant, but the positive
correlation between telecoprids and paracoprids is significant (0.41; Pv0.001).
Guild structure and environmental variables
The microclimatic parameters of the three habitats are given in table 3. Air and
soil temperatures increase between 6:00 h and 14:00 h and decrease during other
times of the day. In our measuring regime, faeces have the highest temperature at
14:00 h and the lowest at 6:00 h with an average difference of 14‡C. Relative air
humidity reaches more than 90% after dusk and decreases only in the morning.
FIG. 3. Diel flight activity of dung beetle guilds, indicated by cumulative abundance in 15samples per 4-h period. Klepto/Tele, kleptoparasites of telecoprids; Klepto/Para,kleptoparasites of paracoprids.
2232 S. Krell-Westerwalbesloh et al.
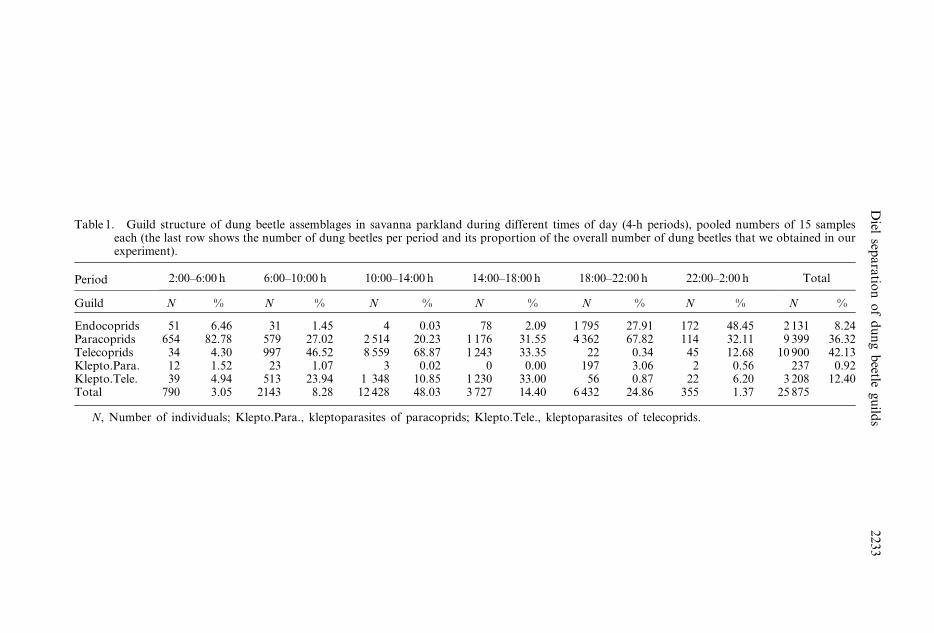
Table 1. Guild structure of dung beetle assemblages in savanna parkland during different times of day (4-h periods), pooled numbers of 15 sampleseach (the last row shows the number of dung beetles per period and its proportion of the overall number of dung beetles that we obtained in ourexperiment).
Period 2:00–6:00 h 6:00–10:00 h 10:00–14:00 h 14:00–18:00 h 18:00–22:00 h 22:00–2:00 h Total
Guild N % N % N % N % N % N % N %
Endocoprids 51 6.46 31 1.45 4 0.03 78 2.09 1 795 27.91 172 48.45 2 131 8.24Paracoprids 654 82.78 579 27.02 2 514 20.23 1 176 31.55 4 362 67.82 114 32.11 9 399 36.32Telecoprids 34 4.30 997 46.52 8 559 68.87 1 243 33.35 22 0.34 45 12.68 10 900 42.13Klepto.Para. 12 1.52 23 1.07 3 0.02 0 0.00 197 3.06 2 0.56 237 0.92Klepto.Tele. 39 4.94 513 23.94 1 348 10.85 1 230 33.00 56 0.87 22 6.20 3 208 12.40Total 790 3.05 2143 8.28 12 428 48.03 3 727 14.40 6 432 24.86 355 1.37 25 875
N, Number of individuals; Klepto.Para., kleptoparasites of paracoprids; Klepto.Tele., kleptoparasites of telecoprids.
Diel
sepa
ratio
no
fd
un
gb
eetleg
uild
s2
23
3

Relationships between environmental parameters and guild abundances over
the day are shown by an RDA (figure 5). Endocoprids and kleptoparasites of
paracoprids are clearly associated with the period around dusk and obviously not
negatively affected by cooling of air and soil during that period. Telecoprids and
their kleptoparasites prefer the hottest time of the day (10:00–14:00 h) and are
closely related to the temperature of faeces, soil and air at the end of the exposure
period. The paracoprids are more related to soil and air temperature at the
beginning of the exposure period. The relative humidity is negatively correlated
with the abundance of paracoprids and kleptoparasites of telecoprids and rather
uncorrelated with the other guilds.
FIG. 4. Biplot of PCA of log-transformed absolute numbers of individuals within each guild[mapping of the data (samples) simultaneously with the mapping of the initial of thevariables (guilds) on the factorial axes]. Eigenvalues: component 1, 0.639; component2, 0.200. Endo, endocoprids (dwellers); Klepto.P, kleptoparasites of paracoprids;Klepto.T, kleptoparasites of telecoprids; Para, paracoprids; Tele, telecoprids (rollers).(#) 6:00–10:00 h samples; (%) 10:00–14:00 h samples; (e) 14:00–18:00 h samples; (.)18:00–22:00 h samples; (&) 22:00–2:00 h samples; (¤) 2:00–6:00 h samples.
2234 S. Krell-Westerwalbesloh et al.
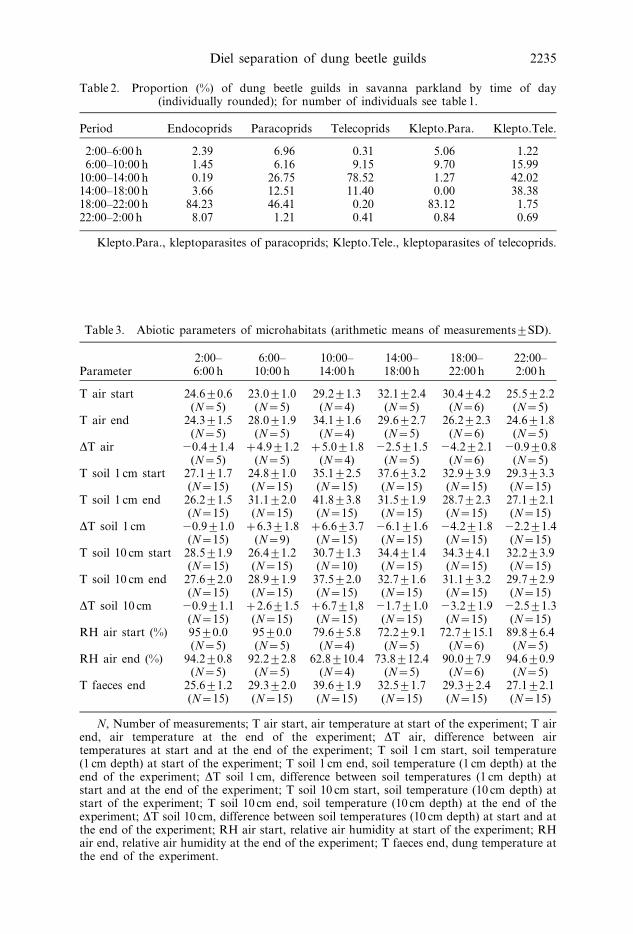
Table 2. Proportion (%) of dung beetle guilds in savanna parkland by time of day(individually rounded); for number of individuals see table 1.
Period Endocoprids Paracoprids Telecoprids Klepto.Para. Klepto.Tele.
2:00–6:00 h 2.39 6.96 0.31 5.06 1.226:00–10:00 h 1.45 6.16 9.15 9.70 15.99
10:00–14:00 h 0.19 26.75 78.52 1.27 42.0214:00–18:00 h 3.66 12.51 11.40 0.00 38.3818:00–22:00 h 84.23 46.41 0.20 83.12 1.7522:00–2:00 h 8.07 1.21 0.41 0.84 0.69
Klepto.Para., kleptoparasites of paracoprids; Klepto.Tele., kleptoparasites of telecoprids.
Table 3. Abiotic parameters of microhabitats (arithmetic means of measurements¡SD).
Parameter2:00–6:00 h
6:00–10:00 h
10:00–14:00 h
14:00–18:00 h
18:00–22:00 h
22:00–2:00 h
T air start 24.6¡0.6 23.0¡1.0 29.2¡1.3 32.1¡2.4 30.4¡4.2 25.5¡2.2(N~5) (N~5) (N~4) (N~5) (N~6) (N~5)
T air end 24.3¡1.5 28.0¡1.9 34.1¡1.6 29.6¡2.7 26.2¡2.3 24.6¡1.8(N~5) (N~5) (N~4) (N~5) (N~6) (N~5)
DT air 20.4¡1.4 z4.9¡1.2 z5.0¡1.8 22.5¡1.5 24.2¡2.1 20.9¡0.8(N~5) (N~5) (N~4) (N~5) (N~6) (N~5)
T soil 1 cm start 27.1¡1.7 24.8¡1.0 35.1¡2.5 37.6¡3.2 32.9¡3.9 29.3¡3.3(N~15) (N~15) (N~15) (N~15) (N~15) (N~15)
T soil 1 cm end 26.2¡1.5 31.1¡2.0 41.8¡3.8 31.5¡1.9 28.7¡2.3 27.1¡2.1(N~15) (N~15) (N~15) (N~15) (N~15) (N~15)
DT soil 1 cm 20.9¡1.0 z6.3¡1.8 z6.6¡3.7 26.1¡1.6 24.2¡1.8 22.2¡1.4(N~15) (N~9) (N~15) (N~15) (N~15) (N~15)
T soil 10 cm start 28.5¡1.9 26.4¡1.2 30.7¡1.3 34.4¡1.4 34.3¡4.1 32.2¡3.9(N~15) (N~15) (N~10) (N~15) (N~15) (N~15)
T soil 10 cm end 27.6¡2.0 28.9¡1.9 37.5¡2.0 32.7¡1.6 31.1¡3.2 29.7¡2.9(N~15) (N~15) (N~15) (N~15) (N~15) (N~15)
DT soil 10 cm 20.9¡1.1 z2.6¡1.5 z6.7¡1,8 21.7¡1.0 23.2¡1.9 22.5¡1.3(N~15) (N~15) (N~15) (N~15) (N~15) (N~15)
RH air start (%) 95¡0.0 95¡0.0 79.6¡5.8 72.2¡9.1 72.7¡15.1 89.8¡6.4(N~5) (N~5) (N~4) (N~5) (N~6) (N~5)
RH air end (%) 94.2¡0.8 92.2¡2.8 62.8¡10.4 73.8¡12.4 90.0¡7.9 94.6¡0.9(N~5) (N~5) (N~4) (N~5) (N~6) (N~5)
T faeces end 25.6¡1.2 29.3¡2.0 39.6¡1.9 32.5¡1.7 29.3¡2.4 27.1¡2.1(N~15) (N~15) (N~15) (N~15) (N~15) (N~15)
N, Number of measurements; T air start, air temperature at start of the experiment; T airend, air temperature at the end of the experiment; DT air, difference between airtemperatures at start and at the end of the experiment; T soil 1 cm start, soil temperature(1 cm depth) at start of the experiment; T soil 1 cm end, soil temperature (1 cm depth) at theend of the experiment; DT soil 1 cm, difference between soil temperatures (1 cm depth) atstart and at the end of the experiment; T soil 10 cm start, soil temperature (10 cm depth) atstart of the experiment; T soil 10 cm end, soil temperature (10 cm depth) at the end of theexperiment; DT soil 10 cm, difference between soil temperatures (10 cm depth) at start and atthe end of the experiment; RH air start, relative air humidity at start of the experiment; RHair end, relative air humidity at the end of the experiment; T faeces end, dung temperature atthe end of the experiment.
Diel separation of dung beetle guilds 2235

Nine of the parameters (environmental variables and times of the day) are
potentially relevant for the interpretation of diel guild patterns since their
eigenvalues in the RDA are higher than 0.2 (table 4). The most important
parameters that possibly affect guild structure are the temperatures of air and soil
(1 cm) at the end of the exposure period, closely followed by the faecal temperature.
FIG. 5. RDA ordination biplot of guilds and environmental variables based on log-transformed guild abundance data of all samples. Eigenvalues: axis 1, 0.577; axis 2,0.162. (,) Centroids of times of the day (categorical variables); Endo, endocoprids(dwellers); Klepto.P, kleptoparasites of paracoprids; Klepto.T, kleptoparasites oftelecoprids; Tele, telecoprids (rollers); diffTair, difference between air temperatures atstart and at the end of the experiment; diffTsoil, difference between soil temperatures(1 cm depth) at start and at the end of the experiment; RHend, relative air humidity atthe end of the experiment; RHstart, relative air humidity at start of the experiment;Tair end, air temperature at the end of the experiment; Tair start, air temperature atstart of the experiment; Tfaeces, dung temperature at the end of the experiment;Tsoil1end, soil temperature (1 cm depth) at the end of the experiment; Tsoil1start, soiltemperature (1 cm depth) at start of the experiment; Tsoil10end, soil temperature(10 cm depth) at the end of the experiment; Tsoil10start, soil temperature (10 cmdepth) at start of the experiment.
2236 S. Krell-Westerwalbesloh et al.

The strong intercorrelation of most of the variables causes a very low conditional
effect of 15 of 17 parameters, but eight of them, including air, faecal and soil
temperature, have a statistically significant effect.
The two periods of highest flight activity are analysed separately (10:00–14:00 h:
figure 6; 18:00–22:00 h: figure 7) to identify possible correlations between micro-
climate and abundance of guilds at a higher temporal resolution.10:00–14:00 h. During their peak flight period telecoprids show no close
correlation with any microclimatic parameter, and only a slightly positive
correlation with the air temperature at the end of the exposure period (i.e.
during the collecting time). The abundance of their kleptoparasites, however, is
related to all absolute temperatures. Absolute numbers of paracoprids are related
to a lower increase of soil temperature. The most important parameters that can
affect guild structure are the temperature of the soil in 1 cm depth at the end of
the exposure period, the relative air humidity and the soil temperatures in 10 cm
depth. However, only the soil temperature in 1 cm depth at the end of the expo-
sure period and the air temperature at 10:00 h have a significant effect in the
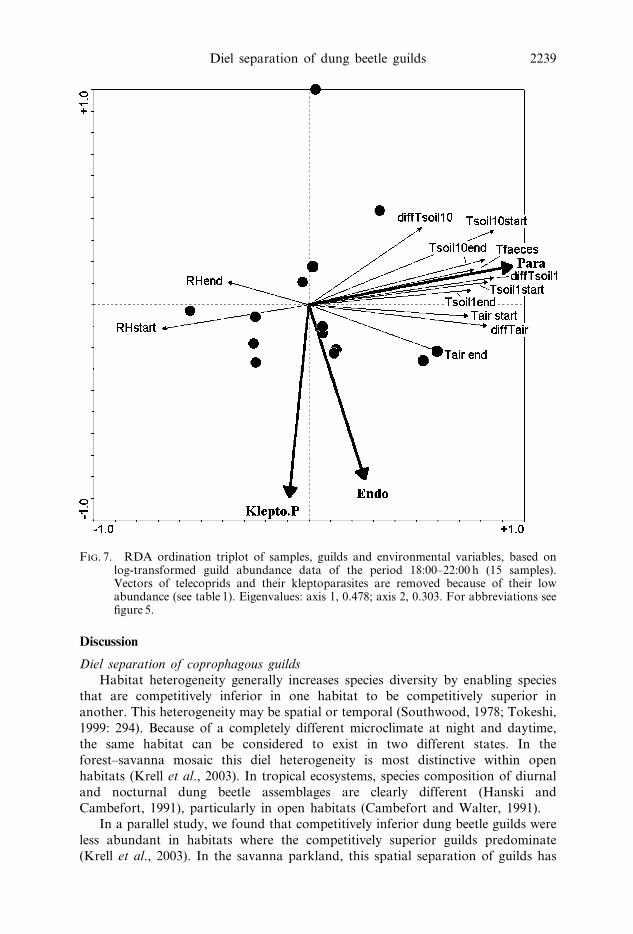
interplay of all variables (table 5).18:00–22:00 h. Around dusk the endocoprids and the kleptoparasites of para-
coprids have their single peak (figure 3). They are not closely correlated with any
of the environmental parameters that we measured. Endocoprids may prefer a
higher air temperature. The paracoprids, however, are strongly positively corre-
lated with all absolute temperatures and negatively correlated with relative air
humidity. Most of these parameters are possibly affecting the guild structure
(table 6). Only the relative air humidity at 22:00 h is negligibly correlated which
approximates to saturation at every night in the rainy season (table 3). In the
Table 4. Eigenvalues of the environmental variables in the RDA (all samples considered).
Marginal effects Conditional effects
Variable l1 Variable lA P
Tair end 0.46 Tair end 0.46 0.001Tsoil1end 0.46 18:00–22:00 h 0.20 0.001Tfaeces 0.45 RHend 0.03 0.001Rhend 0.40 22:00–2:00 h 0.04 0.001diffTsoil10 0.36 6:00–10:00 h 0.02 0.001Tsoil10end 0.33 Tfaeces 0.03 0.00110:00–14:00 h 0.32 Tsoil10end 0.01 0.020diffTair 0.28 diffTair 0.00 0.02618:00–22:00 h 0.26 14:00–18:00 h 0.01 0.010diffTsoil1 0.18 Tsoil1start 0.01 0.350Tsoil1start 0.14 Tair start 0.00 0.49322:00–2:00 h 0.14 2:00–6:00 h 0.00 0.371Rhstart 0.11 Tsoil1end 0.00 0.531Tair start 0.11 diffTsoil1 0.00 0.6572:00–6:00 h 0.07 Tsoil10start 0.00 0.79914:00–18:00 h 0.04 diffTsoil10 0.01 0.891Tsoil10start 0.04 RHstart 0.00 0.6836:00–10:00 h 0.02
The significance of the variables is tested by a Monte Carlo Permutation Test (Canoco;999 permutations).
For abbreviations see figure 5.
Diel separation of dung beetle guilds 2237

interplay of all microclimatic variables, only the soil temperature in 10 cm depth
at 18:00 h has a major and significant effect, whereas the effects of air tempera-
ture are significant but only minor (table 6).
Differences to previous results
The number of the kleptoparasites of telecoprids is remarkably lower than in
our habitat experiment (Krell et al., 2003) where we exposed the dung in the
savanna from 6:00 h to 16:00 h. We suppose that the number of kleptoparasites
is correlated with the length of deposition time during the activity period of these
beetles. They are likely to accumulate in the dung pat during the day, because the
number of arriving individuals is higher than the number of successful individuals
leaving the pat with their host.
FIG. 6. RDA ordination triplot of samples, guilds and environmental variables, based onlog-transformed guild abundance data of the period 10:00–14:00 h (15 samples).Vectors of endocoprids and kleptoparasites of paracoprids are removed becauseof their low abundance (see table 1). Eigenvalues: axis 1, 0.488; axis 2, 0.164. Forabbreviations see figure 5.
2238 S. Krell-Westerwalbesloh et al.
Discussion
Diel separation of coprophagous guilds
Habitat heterogeneity generally increases species diversity by enabling species
that are competitively inferior in one habitat to be competitively superior in
another. This heterogeneity may be spatial or temporal (Southwood, 1978; Tokeshi,
1999: 294). Because of a completely different microclimate at night and daytime,
the same habitat can be considered to exist in two different states. In the
forest–savanna mosaic this diel heterogeneity is most distinctive within open
habitats (Krell et al., 2003). In tropical ecosystems, species composition of diurnal
and nocturnal dung beetle assemblages are clearly different (Hanski and
Cambefort, 1991), particularly in open habitats (Cambefort and Walter, 1991).
In a parallel study, we found that competitively inferior dung beetle guilds were
less abundant in habitats where the competitively superior guilds predominate
(Krell et al., 2003). In the savanna parkland, this spatial separation of guilds has
FIG. 7. RDA ordination triplot of samples, guilds and environmental variables, based onlog-transformed guild abundance data of the period 18:00–22:00 h (15 samples).Vectors of telecoprids and their kleptoparasites are removed because of their lowabundance (see table 1). Eigenvalues: axis 1, 0.478; axis 2, 0.303. For abbreviations seefigure 5.
Diel separation of dung beetle guilds 2239

its temporal parallel at a diel level: the competitively most inferior endocoprids
are flying to the resource in large numbers only around dusk when the best
competitors, the telecoprids, are not active. Walter (1985) described a similar
pattern from central Africa. Telecoprids generally prefer fresh dung without crust
to make their balls. Dung pats exposed before dusk might be already too old or less
attractive for telecoprids when their next activity period starts. Around dusk, we
found also the highest abundance of the competitively intermediate paracoprids.
Paracoprids, however, do not always use the dung completely (figure 2) so that
endocoprids have a chance to feed and reproduce in dung pats used by paracoprids.
Table 6. Eigenvalues of the environmental variables in the RDA of the samples of theperiod 18:00–22:00 h.
Marginal effects Conditional effects
Variable l1 Variable lA P
Tsoil10start 0.40 Tsoil10start 0.40 0.001diffTsoil1 0.36 diffTair 0.14 0.015Tsoil1start 0.36 RHend 0.07 0.117Tsoil10end 0.35 Tair end 0.12 0.011diffTair 0.35 Tair start 0.07 0.031Tfaeces 0.32 diffTsoil1 0.04 0.094Tsoil1end 0.31 RHstart 0.03 0.231Tair start 0.29 Tsoil1start 0.02 0.355Rhstart 0.24 Tsoil10end 0.02 0.600Tair end 0.22 Tfaeces 0.03 0.115diffTsoil10 0.17 diffTsoil10 0.03 0.153Rhend 0.08 Tsoil1end 0.00 0.731
The significance of the variables is tested by a Monte Carlo Permutation Test (Canoco;999 permutations).
For abbreviations see figure 5.
Table 5. Eigenvalues of the environmental variables in the RDA of the samples of theperiod 10:00–14:00 h.
Marginal effects Conditional effects
Variable l1 Variable lA P
Tsoil1end 0.32 Tsoil1end 0.32 0.001Rhend 0.31 RHend 0.11 0.054Tsoil10start 0.27 Tair start 0.15 0.010Tsoil10end 0.23 Tair end 0.04 0.322Rhstart 0.21 Tsoil10start 0.03 0.497Tair start 0.19 Tsoil10end 0.03 0.541diffTsoil1 0.18 Tfaeces 0.02 0.710Tair end 0.16 Tsoil1start 0.02 0.624Tfaeces 0.16 diffTsoil10 0.04 0.481Tsoil1start 0.13 diffTsoil1 0.03 0.607diffTair 0.06diffTsoil10 0.04
The significance of the variables is tested by a Monte Carlo Permutation Test (Canoco;999 permutations).
For abbreviations see figure 5.
2240 S. Krell-Westerwalbesloh et al.

As between habitats (Krell et al., 2003) we find a diel separation of competitors
at the guild level.
Abundance peaks and discontinuous competitive level
During the night and in the morning (22:00–10:00 h) the overall abundance of
dung beetles is very low and certainly at a non-competitive level at the resource
(figure 3). Inferior competitors would therefore have had the chance to use the
resource extensively during this time of the day, but they did not.
We may slightly underestimate the utilization of the resource between 22:00 h
and dawn, because the number of Coprini and Heliocopris spp. (big tunnellers)
was very low during the study period, presumably because the rainfall during
the first half of the rainy season was much below average and Coprini emerge
mainly after strong rain. In a preliminary study at the same site in 1996 (a
relatively wet year) we had a much higher number of Coprini in our samples,
some of which came to the dung after 23:00 h (Westerwalbesloh, unpublished
data). Many Coprini are known to have a flight period that is extended until
long after dusk (Walter, 1985; Davis, 1999; personal observation), so they are
able to use the resource during a period without any competitive pressure.
However, they are not very abundant then (Davis, 1999; Westerwalbesloh,
unpublished data) and use only a part of the resource. The greater part (or,
in our study, nearly all) of the resource remains unused between 22:00 h and
dawn.
We assume that the activity peaks of dung beetles are not correlated with the
availability of dung because herbivore dung appears as an available resource all day
long: buffaloes defecate randomly (Leuthold, 1977: 32), there seem to be no diel
preference periods (Mloszewski, 1983: 120).
Our results indicate that there are vacant temporal refuges (Tokeshi, 1999: 303)
of which dung beetles cannot take advantage whether they are competitively
superior or inferior. Remmert (1976) did not know any well-analysed example of
an ecological niche that is available all the day but realized only during a certain
period of the day. The diel activity of savanna dung beetles is such an example.
Why is the dung beetle abundance so unequally distributed over the day with peaks
of high competition and periods when the resource is neglected?
Neglecting resources due to physiological constraints?
The abundance peak at dusk is a global pattern, and we often find the diurnal
peak around midday (Peck and Forsyth, 1982; Walter, 1985; Fincher et al., 1986;
Davis, 1996; Davis, 1999; Feer, 2000). The nocturnal abundance minimum between
22:00 h and just before dawn is also present in temperate regions (Koskela, 1979),
in Borneo (Davis, 1999), South Africa (Davis, 1996), less pronounced in
South America (Feer, 2000) and postponed to after 2:00 h in Kenya (Heinrich
and Bartholomew, 1979). According to Walter (1985), Afrotropical dung beetles
show flight activity throughout the whole day, but he does not give the abundance
data of the species per hour. Therefore, it is not clear when the species have their
activity peaks. Since the activity generally starts and ends with a low number
of active beetles (Carne, 1956; Koskela, 1979; Palmer and Garcıa Ple, 1992), an
undefinable part of the activity time indicated by Walter may be due to singletons
alone.
Diel separation of dung beetle guilds 2241

Using light intensity as activity trigger probably leads to the crepuscular activ-
ity peak. The abundance peak at dusk is most likely to be caused by a mechan-
ism to synchronize the flight by light intensity differences. Light intensity was
found several times to be responsible for the onset of flight of crepuscular dung
beetles (Aphodius: Carne, 1956; Landin, 1968: 23; Onitis: Houston and McIntyre,
1985) and also in other Scarabaeidae (Kosmachevskii, 1941; Wensler, 1974).
Light intensity possibly functions only as a fine trigger near the flight period
(Carne, 1956) or as a trigger for emergence with an endogenous circadian clock
eventually inducing the onset of flight (Evans and Gyrisco, 1958), because the
activity rhythm can be completely determined by an endogenous clock, but
activity can be prohibited by too much light (Vico et al., 1986). Since light
intensity changes regularly and periodically only at dusk and dawn, species that
use this factor for regulation (i.e. synchronization) of their activity periods have
to be crepuscular. They are able to approach the resource only during a very
limited period of the day and can, consequently, use only a limited number of
the resource patches that are daily deposited in their activity range: only those
that are deposited just before or during their flight period. This habit is certainly
a trade-off between the advantage of the ease of finding a sexual partner (by
synchronization of flight periods) and the disadvantage of a restriction of the
use of resources. A longer activity period with more possibilities to use newly
appeared resources seems to be less advantageous than temporally limited flight
activity.Paracoprids show a positive correlation with temperature of faeces, soil and air
during their activity peaks at dusk (figure 7), but a negative correlation with all
temperature parameters during their diurnal peak (figure 6). They seem to avoid
high diurnal temperatures, which may probably lead to lethal body temperatures
particularly in large black beetles flying in the sun at midday. Lethal body
temperatures for insects are between 40 and 50‡C. A big dung beetle may already
be 44‡C at take-off (Bartholomew and Heinrich, 1978). The competitive superiority
of thermophilous telecoprids is unlikely to be the reason for this pattern, because
their abundance is not negatively correlated with the number of paracoprids during
their activity peak (figure 6).
Since crepuscular paracoprids are positively correlated with temperature
parameters during their activity peak (figure 7), their low number at dawn is as
expected: although dawn is as suitable as dusk for synchronizing flight activity, late
night and dawn are the coldest hours of the day (table 3).Correlation of guild abundance and temperature: the inevitability of a midday
peak. A positive correlation of temperature with flight activity of dung beetles
has been shown in the paracoprid Onitis alexis (Houston and McIntyre, 1985:
soil temperature in 3 cm depth) and in Aphodius (Carne, 1956: air temperature).
Martın-Piera et al. (1994) found that the diurnal Palaearctic roller Sisyphus
schaefferi (L.) is active all day and night if the temperature is at least 29‡C. In
a parallel study comparing habitat types, we found a positive correlation
between abundance of endocoprids, telecoprids and their kleptoparasites with
temperature (Krell et al., 2003). From the present study also a positive correla-
tion between temperature and the abundance of paracoprids, telecoprids and
their kleptoparasites results, if we pool the samples from all over the day
(figure 5). As in the habitat study, numbers of telecoprids and their kleptopara-
sites are strongly correlated with dung, soil and air temperatures at the end of
2242 S. Krell-Westerwalbesloh et al.

the exposure period (i.e. when the temperatures have their maximum). This cor-
roborates our former hypothesis (Krell et al., 2003), that telecoprids prefer
higher temperatures to perform their energetically costly rolling behaviour effec-
tively. Bartholomew and Heinrich (1978) found that the velocity of beetles
increases with the body temperature. For the success of the kleptoparasites of
rollers, high temperature is likely to be only indirectly necessary by increasing
the number of telecoprid hosts.For the (diurnal) telecoprids it seems only to be crucial to have their activity
period during the warmest time of the day. It does not matter how warm it is,
because their numbers show no correlation with any temperature parameter during
their activity peak (figure 6) (or the sample size is too low to show any such
patterns).
Summary
‘Temporal partitioning is, in theory, even less likely. No energetic gain can be
derived from not feeding during most time periods…’ This consideration of
Schoener (1986) does not apply if organisms are forced to a limited activity period
by energetic constraints or at least temporal energetic optima. The diel activity
peaks of dung beetles depend on the climatic optima for the most abundant groups:
the competitively superior telecoprids have their peak at midday when maximum
heat enables them to perform their energetically costly rolling behaviour at the
highest possible speed. The competitively intermediate paracoprids and the com-
petitively inferior endocoprids have their peak around dusk (1) when they can easily
synchronize their flight activity using light intensity changes and (2) when the
superior competitors are not active. During these two activity peaks at midday and
around dusk, the abundance of dung beetles at the resource is very high, causing
to a certain degree obvious competition. On the other hand, between 22:00 h and
10:00 h the resource is hardly used and would offer inferior competitors good
opportunities. However, shifting or prolonging their flight periods to other times
of the day is not a realized strategy to avoid competition. This is probably due to
physiological constraints: the need for high optimum temperature and dependence
on light intensity patterns, respectively.
AcknowledgementsOur employees in the Lola Camp, Koffi Kouadjo, Lakado, Abdoulaye Dabila
Kouadjo and the late Adoulae Ada created a pleasant environment for living and
working in the field. We thank Paul Eggleton and Dorothy Newman, The Natural
History Museum London, for helpful comments. The study was supported by a
grant of the Deutsche Forschungsgemeinschaft (DFG) (Li 150/18-1,2,3, K.E.L.)
and was initially a part of the DFG programme ‘Mechanismen der Aufrechter-
haltung tropischer Diversitat’. Fieldwork was permitted by the Ministere de
l’Agriculture et des Ressources Animales de Cote d’Ivoire, Abidjan.
Appendix
Numbers of individuals per guild per sample
Sample numbers are as written on the specimen labels; Klepto.Para. are
the kleptoparasites of paracoprids; Klepto.Tele. are the kleptoparasites of
telecoprids.
Diel separation of dung beetle guilds 2243

2:00–6:00 h
Sample no. SC20 SC21 SC22 SC44 SC45 SC46 SC55 SC56 SC57 SC58 SC84 SC85 SC86 SC87 SC88
Total Mean SDDate ofcollection
26May
26May
26May
5June
5June
5June
8June
8June
8June
8June
11July
11July
11July
12July
12July
Endocoprids 0 3 5 6 2 7 6 5 6 3 2 0 0 4 2 51 3.4 2.38Paracoprids 44 30 40 40 56 130 27 28 28 28 28 67 46 21 41 654 43.6 26.93Telecoprids 3 3 2 0 2 3 4 6 5 6 0 0 0 0 0 34 2.3 2.25Klepto.Para. 0 1 0 2 1 0 1 0 1 1 1 1 1 0 2 12 0.8 0.68Klepto.Tele. 12 5 8 1 0 1 1 3 2 3 1 0 1 1 0 39 2.6 3.38Total 59 42 55 49 61 141 39 42 42 41 32 68 48 26 45 790 52.7 26.79
6:00–10:00 h
Sample no. SC5 SC6 SC7 SC28 SC29 SC30 SC59 SC60 SC61 SC62 SC72 SC73 SC74 SC89 SC90
Total Mean SDDate ofcollection
8May
8May
8May
28May
28May
28May
8June
8June
8June
8June
12June
12June
12June
12July
12July
Endocoprids 0 1 1 1 3 3 1 0 0 1 1 6 4 6 3 31 2.1 2.02Paracoprids 50 31 67 19 20 28 34 23 57 63 34 22 72 25 34 579 38.6 18.23Telecoprids 49 21 79 1 0 2 130 54 152 113 28 78 288 2 0 997 66.5 79.43Klepto.Para. 0 0 0 4 7 4 0 0 0 0 1 2 1 1 3 23 1.5 2.10Klepto.Tele. 0 1 1 26 63 59 10 13 82 63 14 22 148 3 8 513 34.2 41.58Total 99 54 148 51 93 96 175 90 291 240 78 130 513 37 48 2 143 142.9 125.29
22
44
S.
Krell-W
esterwa
lbeslo
het
al.

10:00–14:00 h
Sample no. SC10 SC11 SC12 SC13 SC31 SC32 SC33 SC34 SC51 SC52 SC53 SC54 SC69 SC70 SC71
Total Mean SDDate ofcollection
18May
18May
18May
18May
30May
30May
30May
30May
7June
7June
7June
7June
10June
10June
10June
Endocoprids 0 0 0 2 0 0 0 0 0 0 0 0 0 2 0 4 0.3 0.70Paracoprids 140 148 90 231 125 239 89 154 140 246 165 192 223 207 129 2 514 167.6 51.95Telecoprids 460 445 369 463 781 1 427 799 730 360 728 377 368 578 402 272 8 559 570.6 292.87Klepto.Para. 0 2 0 0 0 0 0 0 0 0 0 0 0 1 0 3 0.2 0.56Klepto.Tele. 59 97 81 251 107 49 109 225 41 107 79 83 14 27 19 1 348 89.9 68.25Total 659 692 540 947 1 013 1 715 997 1 109 541 1 081 621 643 815 635 420 12 428 828.5 327.33
14:00–18:00 h
Sample no. SC1 SC2 SC3 SC4 SC23 SC24 SC47 SC48 SC49 SC50 SC63 SC64 SC65 SC80 SC81
Total Mean SDDate ofcollection
7May
7May
7May
7May
27May
27May
6June
6June
6June
6June
9June
9June
9June
8July
8July
Endocoprids 0 0 3 0 2 1 5 2 2 3 6 7 2 23 22 78 5.2 7.33Paracoprids 63 90 93 52 66 85 30 59 44 70 139 116 116 50 102 1 176 78.4 30.87Telecoprids 8 31 57 26 88 0 71 93 32 71 197 156 158 112 143 1 243 82.9 60.09Klepto.Para. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0.0 0.00Klepto.Tele. 0 0 0 0 119 48 10 13 6 81 38 21 72 254 568 1 230 82.0 150.40Total 71 121 154 78 275 134 116 167 84 225 380 300 348 438 835 3 727 248.5 200.27
Diel
sepa
ratio
no
fd
un
gb
eetleg
uild
s2
24
5
18:00–22:00 h
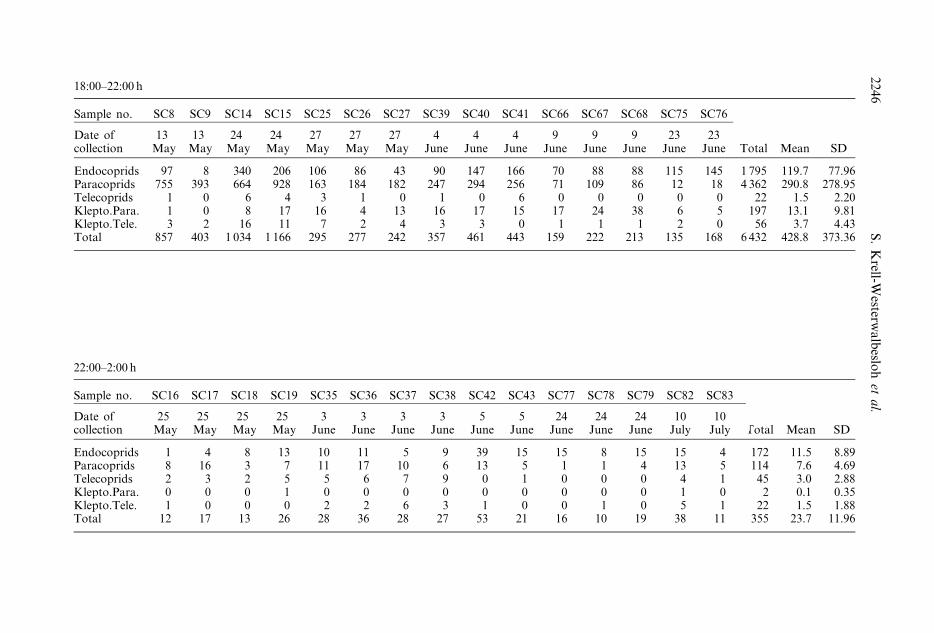
Sample no. SC8 SC9 SC14 SC15 SC25 SC26 SC27 SC39 SC40 SC41 SC66 SC67 SC68 SC75 SC76
Total Mean SDDate ofcollection
13May
13May
24May
24May
27May
27May
27May
4June
4June
4June
9June
9June
9June
23June
23June
Endocoprids 97 8 340 206 106 86 43 90 147 166 70 88 88 115 145 1 795 119.7 77.96Paracoprids 755 393 664 928 163 184 182 247 294 256 71 109 86 12 18 4 362 290.8 278.95Telecoprids 1 0 6 4 3 1 0 1 0 6 0 0 0 0 0 22 1.5 2.20Klepto.Para. 1 0 8 17 16 4 13 16 17 15 17 24 38 6 5 197 13.1 9.81Klepto.Tele. 3 2 16 11 7 2 4 3 3 0 1 1 1 2 0 56 3.7 4.43Total 857 403 1 034 1 166 295 277 242 357 461 443 159 222 213 135 168 6 432 428.8 373.36
22:00–2:00 h
Sample no. SC16 SC17 SC18 SC19 SC35 SC36 SC37 SC38 SC42 SC43 SC77 SC78 SC79 SC82 SC83
Total Mean SDDate ofcollection
25May
25May
25May
25May
3June
3June
3June
3June
5June
5June
24June
24June
24June
10July
10July
Endocoprids 1 4 8 13 10 11 5 9 39 15 15 8 15 15 4 172 11.5 8.89Paracoprids 8 16 3 7 11 17 10 6 13 5 1 1 4 13 5 114 7.6 4.69Telecoprids 2 3 2 5 5 6 7 9 0 1 0 0 0 4 1 45 3.0 2.88Klepto.Para. 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 2 0.1 0.35Klepto.Tele. 1 0 0 0 2 2 6 3 1 0 0 1 0 5 1 22 1.5 1.88Total 12 17 13 26 28 36 28 27 53 21 16 10 19 38 11 355 23.7 11.96
22
46
S.
Krell-W
esterwa
lbeslo
het
al.

References
ANDERSON, J. M. and COE, M. J., 1974, Decomposition of elephant dung in an arid, tropicalenvironment, Oecologia, 14, 111–125.
BARTHOLOMEW, G. A. and HEINRICH, B., 1978, Endothermy in African dung beetles duringflight, ball making, and ball rolling, Journal of Experimental Biology, 73, 65–83.
BEAVER, R. A., 1984, Insect exploitation of ephemeral habitats, South Pacific Journal ofNatural Science, 6, 3–47.
BEGON, M., HARPER, J. L. and TOWNSEND, C. R., 1996, Ecology. Individuals, Populationsand Communities, 3rd edn (Oxford: Blackwell Science), xiiz1068 pp.
BORNEMISSZA, G. F., 1969, A new type of brood care observed in the dung beetle Oniticelluscinctus (Scarabaeidae), Pedobiologia, 9, 223–225.
CAMBEFORT, Y., 1991, Dung beetles in tropical savannas, in I. Hanski and Y. Cambefort(eds) Dung Beetle Ecology (Princeton, NJ: Princeton University Press), pp. 156–178,388–400.
CAMBEFORT, Y. and HANSKI, I., 1991, Dung beetle population biology, in I. Hanski andY. Cambefort (eds) Dung Beetle Ecology (Princeton, NJ: Princeton University Press),pp. 36–50.
CAMBEFORT, Y. and WALTER, P., 1991, Dung beetles in tropical forests in Africa, inI. Hanski and Y. Cambefort (eds) Dung Beetle Ecology (Princeton, NJ: PrincetonUniversity Press), pp. 198–210.
CARNE, P. B., 1956, An ecological study of the pasture scarab Aphodius howitti Hope,Australian Journal of Zoology, 4, 259–318, pls 1, 2.
CAVENEY, S., SCHOLTZ, C. H. and MCINTYRE, P., 1995, Patterns of daily flight activity inonitine dung beetles (Scarabaeinae: Onitini), Oecologia, 103, 444–452.
CONNELL, J. H. and SLAYTER, R. O., 1977, Mechanisms of succession in naturalcommunities and their role in community stability and organization, AmericanNaturalist, 111, 1119–1144.
DAVIS, A. J., 1999, Species packing in tropical forests: diel flight activity of rainforest dung-feeding beetles (Coleoptera: Aphodiidae, Scarabaeidae, Hybosoridae) in Borneo,Raffles Bulletin of Zoology, 47, 473–486.
DAVIS, A. L. V., 1996, Diel and seasonal community dynamics in an assemblageof coprophagous, Afrotropical, dung beetles (Coleoptera: Scarabaeidae s. str.,Aphodiidae, and Staphylinidae: Oxytelinae), Journal of African Zoology, 110,291–308.
DENNISON, D. F. and HODKINSON, I. D., 1983, Structure of the predatory beetle commu-nity in a woodland soil ecosystem II. Diurnal activity rhythms, Pedobiologia, 25,169–174.
DOUBE, B. M., 1987, Spatial and temporal organization in communities associated with dungpads and carcasses, in J. H. R. Gee and P. S. Giller (eds) Organization ofCommunities. Past and Present (Oxford: Blackwell), pp. 255–280.
DOUBE, B. M., 1990, A functional classification for analysis of the structure of dung beetleassemblages, Ecological Entomology, 15, 371–383.
DOUBE, B. M., 1991, Dung beetles of Southern Africa, in I. Hanski and Y. Cambefort (eds)Dung Beetle Ecology (Princeton, NJ: Princeton University Press), pp. 133–155.
EVANS, W. G. and GYRISCO, G. G., 1958, The influence of light intensity on the nocturnalemergence of the European chafer, Ecology, 39, 761–763.
FABRE, J.-H., 1879, Souvenirs entomologiques (Premiere serie). Etudes sur l’instinct et lesmoeurs des insectes (Paris: Delagrave).
FEER, F., 2000, Les coleopteres coprophages et necrophages (Scarabaeidae s. str. etAphodiidae) de la foret de Guyane Francaise: composition specifique et structure despeuplements, Annales de la Societe Entomologique de France NS, 36, 29–43.
FINCHER, G. T., BLUME, R. R., HUNTER, J. S. and BEERWINKLE, K. R., 1986, Seasonaldistribution and diel flight activity of dung-feeding scarabs in open and woodedpasture in east-central Texas, Southwestern Entomologist Supplement, 10, 1–35.
GAUCH, H. G., 1982, Multivariate Analysis in Community Ecology (Cambridge: CambridgeUniversity Press), xz298 pp.
GREIG-SMITH, P., 1980, The development of numerical classification and ordination,Vegetatio, 42, 1–9.
Diel separation of dung beetle guilds 2247

HALFFTER, G. and MATTHEWS, E. G., 1966, The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera, Scarabaeidae), Folia Entomologica Mexicana,12–14, 1–312.
HANSKI, I., 1986, Individual behaviour, population dynamics and community structure ofAphodius (Scarabaeidae) in Europe, Acta Oecologica Oecologia Generalis, 7, 171–187.
HANSKI, I., 1989, Dung beetles, in H. Lieth and M. J. A. Werger (eds) Tropical Rain ForestEcosystems. Biogeographical and Ecological Studies. Ecosystems of the World 14B(Amsterdam: Elsevier), pp. 489–511.
HANSKI, I., 1990, Dung and carrion insects, in B. Shorrocks and I. R. Swingland (eds) Livingin a Patchy Environment (Oxford: Oxford University Press), pp. 127–145.
HANSKI, I., 1991, The dung insect community, in I. Hanski and Y. Cambefort (eds) DungBeetle Ecology (Princeton, NJ: Princeton University Press), pp. 5–21.
HANSKI, I. and CAMBEFORT, Y., 1991, Resource partitioning, in I. Hanski and Y. Cambefort(eds) Dung Beetle Ecology (Princeton, NJ: Princeton University Press), pp. 330–349.
HEINRICH, B. and BARTHOLOMEW, G. A., 1979, Roles of endothermy and size in inter- andintraspecific competition for elephant dung in an African dung beetle, Scarabaeuslaevistriatus, Physiological Zoology, 52, 484–496.
HOUSTON, W. W. and MCINTYRE, P., 1985, The daily onset of flight in the crepuscular dungbeetle Onitis alexis, Entomologia Experimentalis et Applicata, 39, 223–232.
JAKSIC, F. M., 1983, The trophic structure of sympatric assemblages of diurnal andnocturnal birds of prey, American Midland Naturalist, 109, 152–162.
JONGMAN, R. H. G., TER BRAAK, C. J. F. and vAN TONGEREN, O. F. R., 1995, Data Analysisin Community and Landscape Ecology (Cambridge: Cambridge University Press),xxiz299 pp.
KOSKELA, H., 1979, Patterns of diel flight activity in dung-inhabiting beetles: an ecologicalanalysis, Oikos, 33, 419–439.
KOSMACHEVSKII, A. S., 1941, Sumerechnyi let nekotoryck plastinchatousykh zhukov (Thetwilight flight of Lamellicornia beetles), Zoologicheskii Zhurnal, 20, 246–251 [inRussian].
KRELL, F.-T., KRELL-WESTERWALBESLOH, S., WEISS, I., EGGLETON, P. and LINSENMAIR,K. E., 2003, Spatial separation of Afrotropical dung beetle guilds: a trade-offbetween competitive superiority and energetic constraints (Coleoptera: Scarabaeidae),Ecography, 26, 210–222.
LANDIN, B.-O., 1961, Ecological studies on dung-beetles, Opuscula Entomologica Supplement,19, 228 pp.
LANDIN, B.-O., 1968, The diel flight activity of dung-beetles (Coleoptera Scarabaeidae). Astudy of the influence of environmental factors, with particular reference to the light,Opuscula Entomologica Supplement, 32, 172 pp.
LEGENDRE, P. and LEGENDRE, L., 1998, Numerical Ecology, 2nd English edn (Amsterdam:Elsevier).
LEUTHOLD, W., 1977, African Ungulates. A Comparative Review of Their Ethology andBehavioral Ecology (Berlin: Springer), xiiiz307 pp.
LOSER, S., 1980, Zur tageszeitlichen Aktivitatsverteilung von Arthropoda der Bodenstreu(Coleoptera, Diplopoda, Isopoda, Opiliones, Araneae) eines Buchen-Eichen-Waldes(Fago-Quercetum), Entomologia Generalis, 6, 169–180.
MARTIN-PIERA, F., SANMARTIN, I. and LOBO, J. M., 1994, Observaciones sobre el ritmo deactividad diaria en Escarabeidos telecopridos (Coleoptera, Scarabaeidae), Bulletin dela Societe Entomologique de France, 99, 463–470.
MLOSZEWSKI, M. J., 1983, The Behavior and Ecology of the African Buffalo (Cambridge:Cambridge University Press), ixz256 pp.
OJO, O., 1977, The Climates of West Africa (London: Heinemann), xviiz219 pp.PALMER, M. and GARCIA PLE, C., 1992, Relacion entre ciclo de actividad diaria y estructura
de una comunidad: primeros resultados con Aphodius Illiger (Coleoptera:Scarabaeoidea: Aphodiidae), Elytron, 5(Suppl.), 167–175.
PECK, S. B. and FORSYTH, A., 1982, Composition, structure, and competitive behaviour in aguild of Ecuadorian rain forest dung beetles (Coleoptera: Scarabaeidae), CanadianJournal of Zoology, 60, 1624–1634.
2248 S. Krell-Westerwalbesloh et al.

POILECOT, P. (ed.), 1991, Un ecosysteme de savane soudanienne: Le Parc National de laComoe (Cote d’Ivoire), Projet UNESCO/PNUD n‡IVC/87/007, Note technique, 2, 346 pp.
POREMBSKI, S., 1991, Beitrage zur Pflanzenwelt des Comoe-Nationalparks (Elfenbeinkuste),Natur und Museum, 121, 61–83.
REMMERT, H., 1976, Gibt es eine tageszeitliche okologische Nische?, Verhandlungen derDeutschen Zoologischen Gesellschaft, 69, 29–45.
SCHOENER, T. W., 1986, Resource partitioning, in J. Kikkawa and D. J. Anderson (eds)Community Ecology: Pattern and Process (Melbourne: Blackwell), pp. 91–126.
SIMBERLOFF, D. and DAYAN, T., 1991, The guild concept and the structure of ecologicalcommunities, Annual Review of Ecology and Systematics, 22, 115–143.
SOUTHWOOD, T. R. E., 1978, The components of diversity, in L. A. Mound and N. Waloff(eds) Diversity of Insect Faunas. Symposia of the Royal Entomological SocietyLondon 9 (Oxford: Blackwell), pp. 19–40.
TOKESHI, M., 1999, Species Coexistence. Ecological and Evolutionary Perspectives (Oxford:Blackwell Science), xz454 pp.
VICO, L., GROSFILLEY, A. and BUISSON, B., 1986, L’influence de la lumiere sur le rythmelocomoteur circadien endogene d’un insects coprophage (Geotrupes stercorarius L.),Bulletin de la Societe Zoologique de France, 110, 413–419.
WALTER, P., 1985, Diurnal and nocturnal flight activity of scarabaeine coprophages intropical Africa, Geo-Eco-Trop, 9, 67–87.
WENSLER, R. J., 1974, Crepuscular activity of adult Sericesthis geminata (Coleoptera:Scarabaeidae): influence of circadian rhythmicity and light intensity, New ZealandJournal of Zoology, 1, 197–204.
WOLDA, H., 1988, Insect seasonality: why?, Annual Review of Ecology and Systematics, 19,1–18.
ZAR, J. H., 1996, Biostatistical Analysis, 3rd edn (Upper Saddle River, NJ: Prentice Hall),xz662 pp.
Diel separation of dung beetle guilds 2249




















