
Arp2/3 Complex from Acanthamoeba Binds Profilin and Cross-links Actin Filaments
12
Molecular Biology of the Cell Vol. 9, 841– 852, April 1998 Arp2/3 Complex from Acanthamoeba Binds Profilin and Cross-links Actin Filaments R. Dyche Mullins,* Joseph F. Kelleher, ‡ James Xu, § and Thomas D. Pollard* *The Salk Institute for Biological Studies, La Jolla California 92037; ‡ Johns Hopkins University School of Medicine, Baltimore Maryland 21205; and § Johns Hopkins University, Baltimore Maryland 21218 Submitted July 2, 1997; Accepted January 20, 1998 Monitoring Editor: David Drubin The Arp2/3 complex was first purified from Acanthamoeba castellanii by profilin affinity chromatography. The mechanism of interaction with profilin was unknown but was hypothesized to be mediated by either Arp2 or Arp3. Here we show that the Arp2 subunit of the complex can be chemically cross-linked to the actin-binding site of profilin. By analytical ultracentrifugation, rhodamine-labeled profilin binds Arp2/3 complex with aK d of 7 mM, an affinity intermediate between the low affinity of profilin for barbed ends of actin filaments and its high affinity for actin monomers. These data suggest the barbed end of Arp2 is exposed, but Arp2 and Arp3 are not packed together in the complex exactly like two actin monomers in a filament. Arp2/3 complex also cross-links actin filaments into small bundles and isotropic networks, which are mechanically stiffer than solutions of actin filaments alone. Arp2/3 complex is concentrated at the leading edge of motile Acanthamoeba, and its localization is distinct from that of a-actinin, another filament cross-linking protein. Based on localization and actin filament nucleation and cross-linking activities, we propose a role for Arp2/3 in determining the structure of the actin filament network at the leading edge of motile cells. INTRODUCTION Active motility and passive mechanical properties of eukaryotic cells depend on the actin cytoskeleton. Mo- tile cells move across a substrate using actin polymer- ization to drive membrane protrusion at the leading edge (Tilney et al., 1981; Wang, 1985). Adherent cells transmit forces between sites of attachment via bun- dles and orthogonal networks of cross-linked actin filaments. Even nonmotile cells such as Saccharomyces cerevisiae and Schizosaccharomyces pombe rely on actin filaments to transport cellular components and to complete cell division. Despite our detailed under- standing of actin biochemistry and the impressive cat- alog of known actin-binding proteins, a fundamental question remains unanswered: How exactly do eu- karyotic cells control the organization of the actin cytoskeleton? To move or even to change shape, cells must somehow regulate, both spatially and tempo- rally, the de novo nucleation of actin filaments as well as their organization into higher order structures such as bundles or isotropic networks. One potentially important regulator of cytoskeletal organization is the Arp2/3 complex, a seven-subunit protein complex first identified by profilin affinity chromatography (Machesky et al., 1994). The complex is composed of two actin-related proteins, Arp2 and Arp3, and five novel proteins, p40, p35, p19, p18, and p14. Five lines of evidence implicate Arp2/3 complex in regulation of the actin cytoskeleton. (1) The complex is ubiquitous and essential. The genes for Arp2 and Arp3 are highly conserved across eukaryotic phyla, appar- ently as ancient as actin and essential in both budding and fission yeast (for recent reviews see Frankel and Mooseker, 1996; Mullins et al., 1996; Machesky, 1997). This is not true for many proteins associated with the actin cytoskeleton, which appear to have overlapping or nonessential functions. (2) The Arp2/3 complex is directly associated with the actin cytoskeleton. In Acanthamoeba, Arp2/3 complex binds to and decorates ² Corresponding author. © 1998 by The American Society for Cell Biology 841
Transcript of Arp2/3 Complex from Acanthamoeba Binds Profilin and Cross-links Actin Filaments

Molecular Biology of the CellVol. 9, 841–852, April 1998
Arp2/3 Complex from Acanthamoeba Binds Profilinand Cross-links Actin FilamentsR. Dyche Mullins,*† Joseph F. Kelleher,‡ James Xu,§ andThomas D. Pollard*
*The Salk Institute for Biological Studies, La Jolla California 92037; ‡Johns Hopkins University Schoolof Medicine, Baltimore Maryland 21205; and §Johns Hopkins University, Baltimore Maryland 21218
Submitted July 2, 1997; Accepted January 20, 1998Monitoring Editor: David Drubin
The Arp2/3 complex was first purified from Acanthamoeba castellanii by profilin affinitychromatography. The mechanism of interaction with profilin was unknown but washypothesized to be mediated by either Arp2 or Arp3. Here we show that the Arp2subunit of the complex can be chemically cross-linked to the actin-binding site of profilin.By analytical ultracentrifugation, rhodamine-labeled profilin binds Arp2/3 complex witha Kd of 7 mM, an affinity intermediate between the low affinity of profilin for barbed endsof actin filaments and its high affinity for actin monomers. These data suggest the barbedend of Arp2 is exposed, but Arp2 and Arp3 are not packed together in the complexexactly like two actin monomers in a filament. Arp2/3 complex also cross-links actinfilaments into small bundles and isotropic networks, which are mechanically stiffer thansolutions of actin filaments alone. Arp2/3 complex is concentrated at the leading edge ofmotile Acanthamoeba, and its localization is distinct from that of a-actinin, anotherfilament cross-linking protein. Based on localization and actin filament nucleation andcross-linking activities, we propose a role for Arp2/3 in determining the structure of theactin filament network at the leading edge of motile cells.
INTRODUCTION
Active motility and passive mechanical properties ofeukaryotic cells depend on the actin cytoskeleton. Mo-tile cells move across a substrate using actin polymer-ization to drive membrane protrusion at the leadingedge (Tilney et al., 1981; Wang, 1985). Adherent cellstransmit forces between sites of attachment via bun-dles and orthogonal networks of cross-linked actinfilaments. Even nonmotile cells such as Saccharomycescerevisiae and Schizosaccharomyces pombe rely on actinfilaments to transport cellular components and tocomplete cell division. Despite our detailed under-standing of actin biochemistry and the impressive cat-alog of known actin-binding proteins, a fundamentalquestion remains unanswered: How exactly do eu-karyotic cells control the organization of the actincytoskeleton? To move or even to change shape, cellsmust somehow regulate, both spatially and tempo-rally, the de novo nucleation of actin filaments as well
as their organization into higher order structures suchas bundles or isotropic networks.
One potentially important regulator of cytoskeletalorganization is the Arp2/3 complex, a seven-subunitprotein complex first identified by profilin affinitychromatography (Machesky et al., 1994). The complexis composed of two actin-related proteins, Arp2 andArp3, and five novel proteins, p40, p35, p19, p18, andp14.
Five lines of evidence implicate Arp2/3 complex inregulation of the actin cytoskeleton. (1) The complex isubiquitous and essential. The genes for Arp2 and Arp3are highly conserved across eukaryotic phyla, appar-ently as ancient as actin and essential in both buddingand fission yeast (for recent reviews see Frankel andMooseker, 1996; Mullins et al., 1996; Machesky, 1997).This is not true for many proteins associated with theactin cytoskeleton, which appear to have overlappingor nonessential functions. (2) The Arp2/3 complex isdirectly associated with the actin cytoskeleton. InAcanthamoeba, Arp2/3 complex binds to and decorates† Corresponding author.
© 1998 by The American Society for Cell Biology 841

the sides of actin filaments (Mullins et al., 1997), and insea urchin egg extracts the complex binds F-actin af-finity columns and cosediments with actin filaments(Terasaki et al., 1997). The complex also sedimentswith the cytoskeletal fraction after lysis of humanplatelets (Welch et al., 1997a). In vivo Arp2/3 complexis localized to actin-rich regions, especially the leadingedges of motile cells and the actin-rich comet tails ofListeria monocytogenes (Machesky et al., 1994; Kelleheret al., 1995; Mullins et al., 1997, Welch et al., 1997a,1997b) as well as to actin-rich cortical patches in S.pombe and S. cerevisiae (Balasubramanian et al., 1996;Moreau et al., 1996; Winter et al., 1997). (3) Arp2/3 com-plex interacts with the actin-regulatory protein, profilin.In vitro the complex binds profilin affinity columns andin S. pombe, homologs of Arp3 and p40 interact geneti-cally with profilin (Machesky et al., 1994; Balasubrama-nian et al., 1996; McCollum et al., 1996). (4) In S. cerevisiaeArp3 function is required for proper organization andmotility of cortical actin patches (Moreau et al., 1996;Winter et al., 1997) and for the internalization step ofendocytosis (Moreau et al., 1997). (5) Along with partiallypurified mammalian cytoskeletal extracts, Arp2/3 com-plex is sufficient to reconstitute actin-based Listeria mo-tility (Welch et al., 1997a).
Based on structural arguments, Kelleher et al. (1995)suggested that the Arp2/3 complex might nucleate denovo assembly of actin filaments. Welch et al. (1997a)invoked such a nucleation activity to account for theability of Arp2/3 complex to induce the formation ofactin clouds around Listeria cells. And recently wedemonstrated that Arp2/3 complex from Acan-thamoeba tightly caps the pointed ends of actin fila-ments and nucleates formation of filaments that growfrom their barbed ends (Mullins and Pollard, unpub-lished data). These data suggest that the Arp2/3 com-plex plays an important role in initiating polymeriza-tion of actin at the leading edge of motile cells. Herewe show that profilin binds the Arp2/3 complex bydirect interaction with the Arp2 subunit. This is thefirst demonstration of direct binding between an actin-related protein and a conventional actin-binding pro-tein, and it supports the idea of Kelleher et al. (1995)that surfaces on the barbed end of conventional actinare conserved on Arp2 and exposed at the surface ofthe Arp2/3 complex. We also show that in vitroArp2/3 organizes actin filaments into small bundlesand orthogonal networks of the type seen at the lead-ing edges of motile cells, suggesting that the Arp2/3complex not only regulates actin polymerization butalso filament organization at the leading edge.
MATERIALS AND METHODS
Protein PurificationWe purified Arp2/3 complex from Acanthamoeba: extract by bothpoly-L-proline affinity chromatography and by conventional col-
umn chromatography essentially as previously described (Ma-chesky et al., 1994; Mullins et al., 1997) with some modifications.Cells were homogenized in either pyrophosphate extraction bufferas described before (Lynch et al., 1991; Machesky et al., 1994) orsucrose extraction buffer (10 mM Tris-HCl, pH 8.0, 11.6% sucrose, 1mM EGTA, 1 mM ATP, 1 mM dithiothreitol (DTT), 0.1 mM benza-midine, 2 mg/l leupeptin, 20 mg/l soybean trypsin inhibitor, and 10mg/l pepstatin A). For preparations using sucrose extraction, theextract was not dialyzed against a low-salt pyrophosphate bufferbut was loaded directly onto diethylaminoethyl (DEAE)-celluloseequilibrated with 10 mM Tris-HCl, pH 8.0, 0.2 mM CaCl2, 0.5 mMDTT, 0.5 mM ATP, 0.5 mM phenylmethylsulfonylfluoride, and 0.1mM benzamidine (Tseng et al., 1984). For both sucrose and phos-phate extraction procedures, flow-through from the DEAE-cellulosecolumn was collected and loaded onto poly-L-proline Sepharose(Kaiser et al., 1989). Arp2/3 complex was eluted from the poly-L-proline Sepharose with 0.4 M MgCl2 in 10 mM Tris-HCl (pH 7.5),100 mM NaCl, 100 mM glycine, and 1 mM DTT. Profilin wassubsequently eluted with 8 M urea.
Purification of Arp2/3 complex by ion exchange chromatographywas modified from the method originally used by Machesky et al.(1994). Briefly, the DEAE flow-through was adjusted to pH 6.0 bythe addition of solid 2-[N-morpholino]ethanesulfonic acid (MES)and loaded onto P-11 phosphocellulose (PC) (Whatman, Maidstone,England) equilibrated with 25 mM MES, pH 6.0. The PC columnwas washed with 300 mM NaCl and eluted with 500 mM NaCl.Peak fractions were pooled, concentrated, and chromatographed in300 mM NaCl on a Sephacryl S-300 gel filtration column. TheArp-containing pool from this column was then diluted to a finalNaCl concentration of 100 mM and loaded onto a Mono-S FPLCcolumn (Pharmacia Biotech, Piscataway, NJ). The flow-throughfrom this column, containing the majority of the Arp2/3 complex,was loaded onto hydroxyapatite Bio-Gel HT (Bio-Rad, Hercules,CA), and the complex was eluted with 100 mM sodium pyrophos-phate and 100 mM NaCl.
For storage, purified Arp2/3 complex was dialyzed into 10 mMimidazole (pH 7.5), 150 mM NaCl, 0.2 mM MgCl2, 0.2 mM ATP, and1.0 mM dithiothreitol, concentrated by dialysis against solid su-crose, and stored on ice.
Acanthamoeba actin was purified from DEAE column fractions bypolymerization-depolymerization and gel filtration (Pollard, 1984).Rabbit skeletal muscle actin was purified from acetone powder bythe method of Spudich and Watt (1971) followed by gel filtration(MacLean-Fletcher and Pollard, 1980).
Chemical Cross-linkingThe zero-length chemical cross-linker 1-ethyl-3-(3-dimethylamin-opropyl)-carbodiimide hydrochloride (EDC), along with N-hy-droxysuccinimide (NHS) were obtained from Pierce (Rockford, IL).Stock solutions (203) were made fresh in dry dimethylsulfoxide(DMSO) immediately before use. Final concentration of DMSO in allreactions was #10%. Reactions were carried out for 1 h at roomtemperature.
Preparation of Fluorescently-labeled ProteinsAcanthamoeba actin was labeled with pyrene iodoacetamide (Pol-lard, 1984). Acanthamoeba profilin-II mutants N58C and S38C werelabeled with rhodamine maleimide (Vinson, De La Cruz, Kaiser,Higgs, Pollard, unpublished data).
Profilin Binding by Equilibrium UltracentrifugationWe did equilibrium analytical ultracentrifugation in a Beckmanmodel XL-A ultracentrifuge. We loaded samples containing rhoda-mine-labeled profilin-II into six-hole, charcoal-filled Epon center-pieces and centrifuged them to equilibrium (at least 20 h at eachspeed) in a Beckman model An60ti rotor. We monitored absorption
R.D. Mullins et al.
Molecular Biology of the Cell842

at 550 nm, a wavelength specific for the rhodamine-label and col-lected data sets every hour. To monitor the approach to equilibriumwe calculated total root mean square (RMS) deviation of each dataset from the final data set collected using the program MATCH (JeffLary, National Analytical Ultracentrifuge Facility, Storrs CT). Weconsidered experiments to have reached equilibrium when therewas no change in RMS deviation of consecutive data sets. Weparsed the data from individual samples into separate data setsusing the program REEDIT (Jeff Lary, National Analytical Ultracen-trifuge Facility).
Absorption optics allow us to selectively monitor labeled profilinin the presence of other proteins, to determine directly the distribu-tions of free and bound profilin, and to calculate equilibrium bind-ing constants (Schachman, 1960; Schachman et al., 1962). A similarmethod has been used previously to measure equilibrium bindingof pyrene-labeled actobindin to actin (Bubb et al., 1994). First we fiteach data set to a sum of two exponential functions by linear leastsquares methods
P0espSr2
2 2rm
2
2D 1 P1espaSr2
2 2rm
2
2Dwhere Po and P1 are the fitting constants to be determined, r is radialposition, rm is the radius of the meniscus, and sp and spa are theeffective reduced molecular weights of profilin alone and profilinbound to Arp2/3 complex or to actin monomers. The effectivereduced molecular weight is defined as
s 5Mv2(12vr)
RT
where M is molecular mass (g/mol), v is the angular velocity(rad/sec), v is the partial specific volume (cm3/gm), r is the solventdensity (gm/cm3), R is the gas constant, and T is the absolutetemperature (K). From these fits we determined the total amounts offree and bound profilin by
Pfree 5Erm
rb
P0espSr2
2 2rm
2
2Ddr Pbound 5Erm
rb
P1espaSr2
2 2rm
2
2D dr
Where rm and rb are the radii of the meniscus and the bottom of thesample cell. The total amount of profilin is simply the sum of thebound and free. The total amount of Arp2/3 complex or actin can becalculated from the relative loading concentration.
Ptotal 5 Pfree 1 PboundAtotal 5 CrPtotal
Where Cr is the ratio of the initial Arp2/3 complex or actin concen-tration to the initial profilin concentration. For a stoichiometry of1:1, the amount of bound profilin will equal the amount of boundArp2/3 complex or actin and we calculate the distribution of freeArp2/3 or actin
A0esaSr2
2 2rm
2
2Dfrom
A0 5 (Atotal 2 Pbound) 3Erm
rb
esaSr2
2 2rm
2
2D dr421
The dissociation equilibrium constant is then given by
Kd 5A0P0
P1
It is apparent from the formulas above that the accuracy of thismethod depends on accurate determination of both the fitting pa-rameters P0 and P1 as well as the positions of the meniscus and baseof the sample cell. The position of the meniscus is marked by a spikein the data set and can be assigned with confidence. We added asmall amount of a dense imiscible liquid (usually carbon tetrachlo-ride) to each sample chamber to form a flat base, perpendicular tothe direction of centrifugation, so the exact position of the bottom ofthe sample column was less obvious. For our calculation we chosethe base of each sample chamber to satisfy conservation of massaccording to the following equation
[P]0 5Erm
rb
P0espSr2
2 2rm
2
2D 1 P1espaSr2
2 2rm
2
2D dr
Where [P]0 is the initial baseline concentration of labeled profilin inthe sample cell at time t 5 0.
RheometryRheological measurements were made with a cone and plate Rheo-metrics rheometer (Rheometrics Inc., Piscataway, NJ) in the smallamplitude(strain # 2%), forced oscillation mode (Sato et al., 1984).Arp2/3 complex in KME buffer (50 mM KCl, 10 mM imidazole, pH7.0, 1 mM EGTA, and 1 mM MgCl2) was mixed with actin in BufferG [2 mM Tris-HCl (pH 8.0), 0.2 mM CaCl2, 0.2 mM ATP, 0.5 mMDTT, and 1 mM NaN3], polymerized by adding one-tenth volume of10 3 KME and immediately placed between the metal plates of therheometer to polymerize at 25°C. The plates were kept in a water-saturated chamber to prevent sample dehydration. Measurementsof G9 and G0 were made every 30 s using time sweep mode toobserve the gel formation. After G9 and G0 reached a plateau,frequency sweep mode was used to measure the rheological param-eters: the value of complex modulus uG*u, where uG*u5(G’21G“2)1/2;and the phase shift d, where d5tan–1(G”/G’) (Ferry, 1980).
Electron MicroscopyAcanthamoeba actin filaments in the presence and absence of Arp2/3complex were negatively stained with freshly prepared 1% uranylformate (Aebi et al., 1981). Other samples were fixed, embedded,and sectioned (Maciver et al., 1991). Acanthamoeba actin (10 mM) waspolymerized in the presence or absence of 1 mM Arp2/3 complex ina 50 ml volume on a porcelain tray in a humid chamber for 1 h at24°C. Samples were overlaid with 1% glutaraldehyde and 2 mg/mltannic acid in 100 mM sodium phosphate, pH 7.0, 50 mM KCl, and5 mM MgCl2 for 30 min at 24°C, washed with 100 mM sodiumphosphate, pH 7.0, and then with 100 mM sodium phosphate, pH6.0, fixed again with 0.1% OsO4 in 100 mM sodium phosphate, pH6.0, at 24°C for 30 min, and then dehydrated with an ethanol seriesand embedded in Epon for thin sectioning. All micrographs weremade at 80 kV with a JEOL JEM100CX or a Zeiss 10A electronmicroscope (Carl Zeiss, Thornwood, NY) at nominal magnificationsbetween 23,0003 and 46,0003. Actual magnification was deter-mined by calibration with tropomyosin paracrystals. Negative im-ages were scanned digitally and prepared for presentation usingAdobe Photoshop.
Monoclonal AntibodiesMouse monoclonal antibodies against Acanthamoeba profilin (Kaiserand Pollard, 1996) and Acanthamoeba a-actinin (Kaiser, Sato, Karaki,and Pollard, unpublished) were prepared by standard methods(Kiehart et al., 1984).
Arp2/3 Complex
Vol. 9, April 1998 843

Fluorescence MicroscopyAcanthamoeba were maintained and fixed as previously described(Yonemura and Pollard, 1992). Cells were incubated with rabbitpolyclonal antisera specific for Acanthamoeba Arp2, Arp3 (Kelleher,et al., 1995), p40, or p35 (Mullins et al., 1997) or mouse monoclonalantibodies specific for Acanthamoeba a-actinin, for 1 h at 24°C. Pre-absorbed (Yonemura and Pollard, 1992) Cy-3 conjugated goat anti-rabbit IgG secondary antibody (Amersham, Arlington Heights, IL)was applied for 30 min. BODIPY-FL phallicidin (Molecular Probes,
Sunnyvale, CA) was used to label filamentous actin. Cells weremounted in 50% glycerol containing 20 mM DTT and 5.6 mMp-phenylenediamine. We observed and photographed labeled cellsusing both phase-contrast and fluorescence microscopy as describedpreviously (Fujiwara and Pollard, 1976). Figures were preparedusing Adobe Photoshop from digitally scanned 35-mm negatives.
RESULTS
Profilin Binds the Arp2 Subunit of the Arp2/3ComplexBy optimizing conditions we were able to cross-linkboth profilin-I and profilin-II to the Arp2/3 complexusing the zero-length cross-linker EDC with NHS. At20 mM profilin and 3 mM Arp2/3, EDC/NHS pro-duces two new bands on SDS-PAGE of 56 kDa and 59kDa that react with antibodies against Acanthamoebaprofilin (Figure 1A). Profilin-I and profilin-II producedidentical sets of bands. Both bands react with anti-Arp2 antibodies but not with antibodies to Arp3 orany other member of the complex (Figure 1B). Weconclude that the two bands are two different het-erodimers of Arp2 and profilin, cross-linked at differ-ent residues and with slightly different mobilities onSDS- PAGE gels. Under other conditions, chemicalcross-linking agents that covalently cross-link actin toprofilin fail to cross-link profilin to any subunit ofArp2/3 complex (Mullins et al., 1997).
A panel of monoclonal antibodies to profilin revealsthat the 56-kDa product is similar to the profilin-actincomplex cross-linked by EDC/NHS. EDC/NHS cross-links E364 of Acanthamoeba actin to K115 of Acan-thamoeba profilin-I or profilin-II (Vandekerchove et al.,1989). This binding site has been studied with the aidof four epitope-mapped monoclonal anti-profilin an-tibodies, P4, P5, P6 and P7 (Kaiser and Pollard, 1996).P5 and P7 bind profilin near K115, and cross-linkingprofilin to actin by EDC/NHS blocks binding of bothantibodies (Kaiser and Pollard, 1996). Antibodies P4and P6 bind to different regions of profilin and stillrecognize profilin cross-linked to actin. All four anti-bodies recognize the upper band, but only P4 and P6react with the 56-kDa band (Figure 2). P5 and P7 both
Figure 1. EDC with NHS cross-links Acanthamoeba profilin to theArp2 subunit of the Arp2/3 complex. (A) SDS-PAGE of proteinscross-linked with EDC/NHS and stained with Coomassie blue (cb)or immunoblotted (ib) with monoclonal antiprofilin antibodies.Samples, as indicated by the horizontal bars: 3 mM Arp2/3 complex,Arp2/3 plus 20 mM profilin-II, profilin-II alone. (B) Immunoblot ofArp2/3 complex and profilin-II cross-linked with EDC/NHS andprobed with antibodies against profilin (lanes 1 and 4), Arp2 (lanes2 and 5), and Arp3 (lanes 3 and 6). Lanes 1–3, 3 mM Arp2/3complex; lanes 4–6, 3 mM Arp2/3 and 20 mM profilin-II. Asteriskmarks the two profilin/Arp2 cross-linked products. Identical resultswere obtained with Acanthamoeba profilin-I. Conditions: Cross-link-ing was carried out for 1 h at 24°C with 5 mM EDC and 5 mM NHS.Buffer: 150 mM NaCl, 0.2 mM MgCl2, 0.1 mM EGTA, 0.2 mM ATP,1 mM DTT, 10 mM imidazole, pH 7.5. Arp2/3 complex was purifiedby profilin:poly-L-proline affinity chromatography.
Figure 2. Arp2 binds at or near the actin-binding site on profilin.Immunoblot of SDS-PAGE of profilin-II cross-linked to Arp2/3complex with EDC/NHS and probed with four epitope-mapped,monoclonal anti-profilin antibodies, P4, P5, P6, and P7. AntibodiesP4 and P6 bind free profilin and profilin cross-linked to actin byEDC/NHS. Both also react with the two Arp2-profilin cross-linkedproducts. P5 and P7 bind free profilin but not profilin cross-linkedto actin and do not bind the smaller of the two Arp2-profilincross-linked products. Conditions as in Figure 1.
R.D. Mullins et al.
Molecular Biology of the Cell844

fail to recognize this band. We conclude that the lowerband is Arp2 cross-linked to profilin at or near K115,in the actin-binding site of profilin. The upper band isa distinct cross-linked product of Arp2 and profilinwith a different mobility on SDS-PAGE in which Arp2is probably cross-linked to a residue of profilin otherthan K115. The presence of this product does notnecessarily suggest two binding sites for profilin onArp2 but rather the proximity of another cross-link-able residue of profilin to a residue on Arp2 that is notconserved on conventional actin.
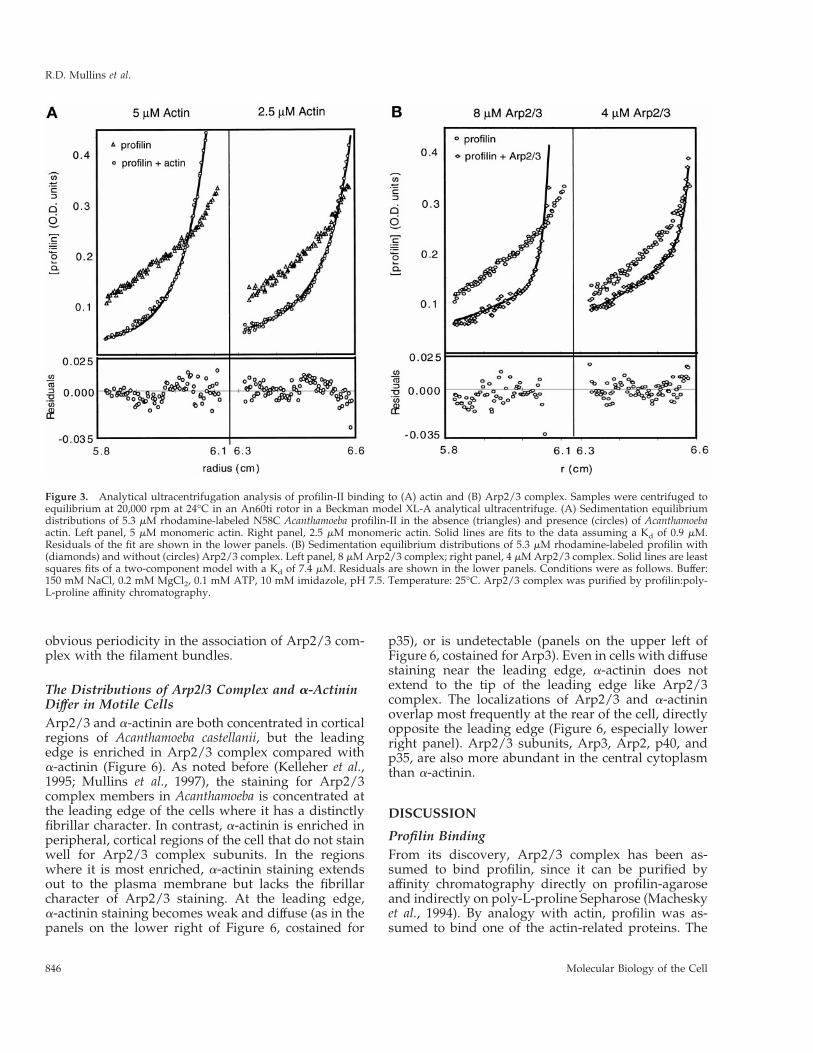
Profilin Binds Actin More Strongly than the Arp2/3ComplexWe used sedimentation equilibrium ultracentrifuga-tion to determine the affinity of profilin-II for actin andArp2/3 complex. To measure the concentration ofprofilin in the presence of actin or Arp2/3 complex,we labeled profilin-II mutants in which eitherserine-38 or asparagine-58 was replaced by cysteine(S38C and N58C profilin-II) with rhodamine maleim-ide. Actin and poly-L-proline binding of these rhoda-mine-labeled constructs is indistinguishable from thatof unlabeled profilin (Vinson, De La Cruz, Kaiser,Higgs, and Pollard, unpublished data). At equilib-rium, both rhodamine-labeled mutants form exponen-tial gradients characteristic of single, homogeneous,and thermodynamically ideal species. For N58C wecalculated a native molecular weight of 12,930 Da,close to the value of 13,045 Da deduced from aminoacid sequence.
Measuring the distribution of labeled profilin in thepresence of actin or Arp2/3 complex by absorbance at550 nm gave a signal equal to the sum of the absor-bances of bound and free profilin. We determinedequilibrium dissociation constants for profilin bindingactin and Arp2/3 complex by least-squares fitting ofthe equilibrium distribution of labeled profilin to asum of two distributions, one determined by the ef-fective reduced molecular weight of profilin alone (sp)and the other by that of profilin bound to the largerprotein (spa). We measured the effective reduced mo-lecular weights of profilin, actin, and Arp2/3 complexindividually, at the same speeds and in the samebuffers used in the binding experiments and usedthese values in curve fitting. We independently calcu-lated and averaged equilibrium dissociation constantsfrom six data sets at three different protein concentra-tions spun at two speeds.
By this method N58C profilin-II binds Ca21-ATPAcanthamoeba actin monomers with a Kd of 0.92 6 1.0mM (n 5 6, Figure 3A) and Arp2/3 complex with a Kdof 7.4 6 0.5 mM (n 5 6, Figure 3B). The conditionsdiffered in these two experiments. A low-ionicstrength buffer was used in the actin binding experi-ments to keep actin monomers from polymerizing,
while 150 mM NaCl was used in the Arp2/3 bindingexperiment to keep the complex from aggregating.The large uncertainty in the actin experiment arisesfrom the fact that the concentration of profilin neededfor detection (5 mM) is well above the Kd of the inter-action. Similar results were obtained with the S38Cmutant, indicating that the specific cysteine mutationsand rhodamine labeling do not alter the binding toArp2/3 complex.
Arp2/3 Complex Cross-links and Bundles ActinFilamentsPreviously we found that Arp2/3 complex binds tothe sides of actin filaments (Mullins et al., 1997) andhypothesized that it could cross-link filaments intonetworks or bundles. Preliminary experimentsshowed that Arp2/3 complex pellets with actin fila-ments under centrifugal forces where actin filamentsalone do not pellet (our unpublished results). We char-acterized these higher order structures by quantitativerheometry and electron microscopy.
By quantitative rheometry, Arp2/3 complex in-creases the stiffness of solutions of actin filaments in aconcentration-dependent manner (Figure 4). Additionof Arp2/3 complex increases the complex modulus,uG*u—a measure of stiffness, and decreases the phaseshift, d—a measure of fluid- or solid-like behavior(Figure 4B). In these assays, d 5 0 radians for a solidand d 5 1.6 radians for a Newtonian fluid. Viscoelasticmaterials like actin filaments have intermediate valuesof d.
In the presence of Arp2/3 complex, actin filamentsform networks of individual filaments and bundlesof filaments that are visible in electron micrographs ofthin sections of samples imbedded in Epon and ofsamples adsorbed to EM grids and negatively stainedwith 1% uranyl formate (Figure 5). Pure actin fila-ments have a low tendency to associate, and the den-sity of individual filaments in thin sections of actinfilaments alone is quite uniform. Actin filament net-works in the presence of Arp2/3 complex are quiteheterogeneous in thin sections cut from a single sam-ple. Individual filaments are visible, but many areaggregated into bundles containing (within the thick-ness of the section) two to eight filaments. Individualfilaments appear to fray off the ends of these bundles,and some appear to run from one bundle to another.Bundles tend to associate over a wide range of anglesfrom nearly parallel to perpendicular and are studdedwith 10-nm particles not seen in samples of pure actinfilaments or in bundles induced by a-actinin (Wach-sstock et al., 1993b). The size of these particles is con-sistent with the low resolution structure of the com-plex determined by electron microscopy of rotaryshadowed samples (Mullins et al., 1997). There was no
Arp2/3 Complex
Vol. 9, April 1998 845
obvious periodicity in the association of Arp2/3 com-plex with the filament bundles.
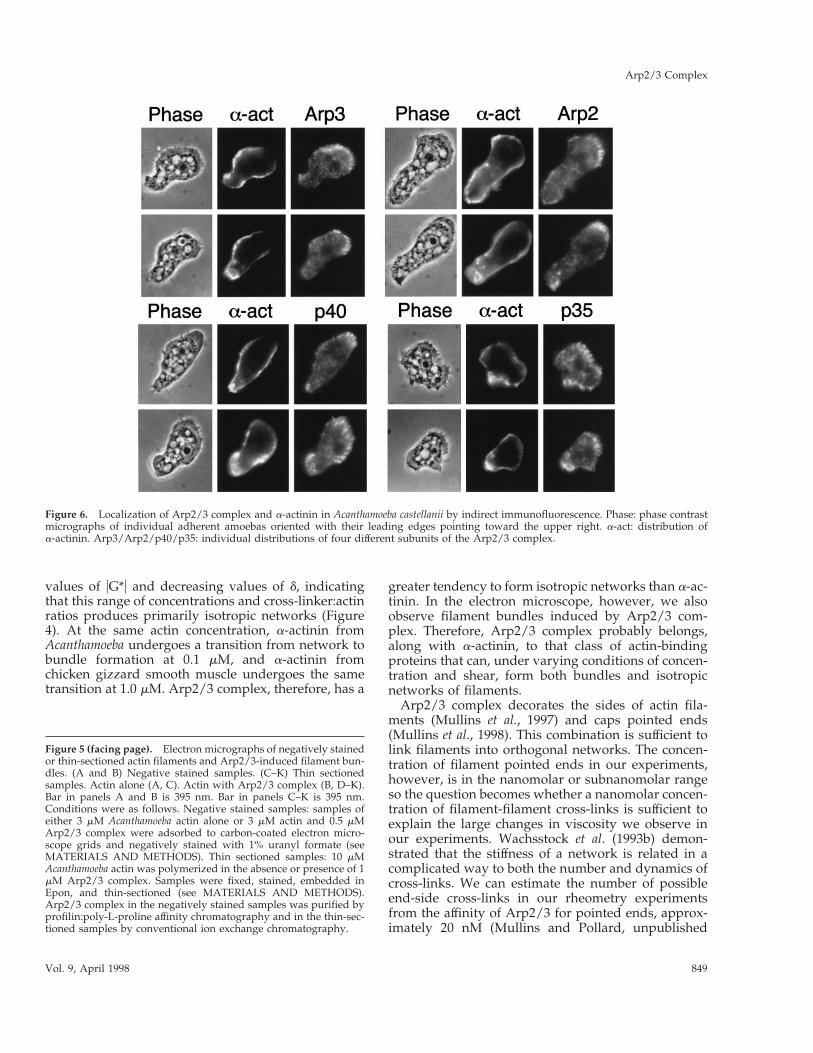
The Distributions of Arp2/3 Complex and a-ActininDiffer in Motile CellsArp2/3 and a-actinin are both concentrated in corticalregions of Acanthamoeba castellanii, but the leadingedge is enriched in Arp2/3 complex compared witha-actinin (Figure 6). As noted before (Kelleher et al.,1995; Mullins et al., 1997), the staining for Arp2/3complex members in Acanthamoeba is concentrated atthe leading edge of the cells where it has a distinctlyfibrillar character. In contrast, a-actinin is enriched inperipheral, cortical regions of the cell that do not stainwell for Arp2/3 complex subunits. In the regionswhere it is most enriched, a-actinin staining extendsout to the plasma membrane but lacks the fibrillarcharacter of Arp2/3 staining. At the leading edge,a-actinin staining becomes weak and diffuse (as in thepanels on the lower right of Figure 6, costained for
p35), or is undetectable (panels on the upper left ofFigure 6, costained for Arp3). Even in cells with diffusestaining near the leading edge, a-actinin does notextend to the tip of the leading edge like Arp2/3complex. The localizations of Arp2/3 and a-actininoverlap most frequently at the rear of the cell, directlyopposite the leading edge (Figure 6, especially lowerright panel). Arp2/3 subunits, Arp3, Arp2, p40, andp35, are also more abundant in the central cytoplasmthan a-actinin.
DISCUSSION
Profilin BindingFrom its discovery, Arp2/3 complex has been as-sumed to bind profilin, since it can be purified byaffinity chromatography directly on profilin-agaroseand indirectly on poly-L-proline Sepharose (Macheskyet al., 1994). By analogy with actin, profilin was as-sumed to bind one of the actin-related proteins. The
Figure 3. Analytical ultracentrifugation analysis of profilin-II binding to (A) actin and (B) Arp2/3 complex. Samples were centrifuged toequilibrium at 20,000 rpm at 24°C in an An60ti rotor in a Beckman model XL-A analytical ultracentrifuge. (A) Sedimentation equilibriumdistributions of 5.3 mM rhodamine-labeled N58C Acanthamoeba profilin-II in the absence (triangles) and presence (circles) of Acanthamoebaactin. Left panel, 5 mM monomeric actin. Right panel, 2.5 mM monomeric actin. Solid lines are fits to the data assuming a Kd of 0.9 mM.Residuals of the fit are shown in the lower panels. (B) Sedimentation equilibrium distributions of 5.3 mM rhodamine-labeled profilin with(diamonds) and without (circles) Arp2/3 complex. Left panel, 8 mM Arp2/3 complex; right panel, 4 mM Arp2/3 complex. Solid lines are leastsquares fits of a two-component model with a Kd of 7.4 mM. Residuals are shown in the lower panels. Conditions were as follows. Buffer:150 mM NaCl, 0.2 mM MgCl2, 0.1 mM ATP, 10 mM imidazole, pH 7.5. Temperature: 25°C. Arp2/3 complex was purified by profilin:poly-L-proline affinity chromatography.
R.D. Mullins et al.
Molecular Biology of the Cell846

profilin-binding site of actin is more conserved instructural models of Arp2 than of Arp3 (Kelleher et al.,1995). In line with these predictions, we find thatEDC/NHS cross-links both profilin-I and profilin-II toArp2 but not to Arp3.
With either profilin-I or profilin-II EDC/NHS pro-duces two cross-linked profilin-Arp2 conjugates withslightly different mobilities on SDS-PAGE gels. Reac-tion of these cross-linked products with a panel ofepitope-mapped, anti-profilin monoclonal antibodiesshows that the 56-kDa product is similar to profilincross-linked to actin-E364 by EDC, because cross-linked Arp2 blocks binding of two antibodies with
epitopes in the actin-binding site, just like cross-linkedactin (Kaiser and Pollard, 1996). We conclude thatArp2 contacts the actin-binding site of profilin. Incross-linking experiments there was no detectable dif-ference between profilin-I and profilin-II. Arp1 fila-ments are thought to bind spectrin (Holleran et al.,1996), but ours is the first direct experimental evidencethat an Arp binds to the actin-binding site of a con-ventional actin-binding protein. Since the profilinbinding site on actin is located at the barbed end ofthe subunit, the barbed end of Arp2 is probablyexposed on the surface of the complex.
By sedimentation equilibrium ultracentrifugation,rhodamine-profilin-II binds Arp2/3 with a Kd of 7.4mM. This method yielded a Kd for profilin binding toactin of 0.9 mM. Under similar conditions, using fluo-rescence anisotropy, Vinson and Pollard (unpublisheddata) measured a Kd of 0.2 mM for the same rhoda-mine-labeled profilin mutant binding to AcanthamoebaCa21-ATP-actin. By both measurements the affinity ofprofilin for conventional actin monomers is at least 10times higher than for Arp2/3 complex. The dissocia-tion rate constant of profilin from actin is 4 s21 (Vin-son, DeLaCruz, Kaiser, Higgs, and Pollard, unpub-lished data), so we predict that profilin is in a rapidequilibrium with Arp2 at a rate of at least 50 s21.
Arp2/3 Complex Cross-links Actin FilamentsFilament cross-linking proteins are traditionallyclassed as either bundling or network-forming pro-teins, depending on their tendency to form either par-allel bundles or randomly oriented isotropic networksof filaments. However, many actin-binding proteins,such as a-actinin, induce the formation of both bun-dles and isotropic networks. Wachsstock et al. (1993a)proposed a model to describe the dependence of themechanical properties of cross-linked actin filamentnetworks on the concentrations of actin and cross-linker. At low concentrations of cross-linker, filamentsare bound in a random isotropic network that behaveslike a single elastic solid. In this region, the complexmodulus uG*u increases and the phase shift d decreasesas a function of cross-linker concentration. Above athreshold concentration that depends on the affinity ofthe cross-linker, actin filaments become organized intobundles. These bundles, although long and stiff, canslip past each other in solution and behave more likea very viscous fluid. In this region uG*u begins to de-crease, and d begins to increase as a function of cross-linker concentration.
To determine whether Arp2/3 complex preferen-tially forms bundles or isotropic networks in solutionwe performed quantitative rheometry on actin fila-ments cross-linked with Arp2/3 complex. With 15 mMfilamentous actin, concentrations of Arp2/3 complexfrom 0.1–3.0 mM produced monotonically increasing
Figure 4. Rheological properties of actin filaments in the presenceof Arp2/3 complex. (A) Frequency dependence of complex modu-lus, uG*u. (B) Frequency dependence of the phase shift. Samplescontained 15 mM rabbit skeletal muscle actin in the absence (F) orpresence of Arp2/3 complex at 0.1 mM (E), 0.5 mM (f), 1.5 mM (M),or 3 mM (Œ). Conditions were as follows. Cone and plate rheometerin the small amplitude (strain # 2%), forced oscillation mode.Buffer: 50 mM KCl, 10 mM imidazole, pH 7.0, 1 mM EGTA, and 1mM MgCl2. Arp2/3 complex was purified by conventional ionexchange chromatography.
Arp2/3 Complex
Vol. 9, April 1998 847

Figure 5.
R.D. Mullins et al.
Molecular Biology of the Cell848
values of uG*u and decreasing values of d, indicatingthat this range of concentrations and cross-linker:actinratios produces primarily isotropic networks (Figure4). At the same actin concentration, a-actinin fromAcanthamoeba undergoes a transition from network tobundle formation at 0.1 mM, and a-actinin fromchicken gizzard smooth muscle undergoes the sametransition at 1.0 mM. Arp2/3 complex, therefore, has a
greater tendency to form isotropic networks than a-ac-tinin. In the electron microscope, however, we alsoobserve filament bundles induced by Arp2/3 com-plex. Therefore, Arp2/3 complex probably belongs,along with a-actinin, to that class of actin-bindingproteins that can, under varying conditions of concen-tration and shear, form both bundles and isotropicnetworks of filaments.
Arp2/3 complex decorates the sides of actin fila-ments (Mullins et al., 1997) and caps pointed ends(Mullins et al., 1998). This combination is sufficient tolink filaments into orthogonal networks. The concen-tration of filament pointed ends in our experiments,however, is in the nanomolar or subnanomolar rangeso the question becomes whether a nanomolar concen-tration of filament-filament cross-links is sufficient toexplain the large changes in viscosity we observe inour experiments. Wachsstock et al. (1993b) demon-strated that the stiffness of a network is related in acomplicated way to both the number and dynamics ofcross-links. We can estimate the number of possibleend-side cross-links in our rheometry experimentsfrom the affinity of Arp2/3 for pointed ends, approx-imately 20 nM (Mullins and Pollard, unpublished
Figure 6. Localization of Arp2/3 complex and a-actinin in Acanthamoeba castellanii by indirect immunofluorescence. Phase: phase contrastmicrographs of individual adherent amoebas oriented with their leading edges pointing toward the upper right. a-act: distribution ofa-actinin. Arp3/Arp2/p40/p35: individual distributions of four different subunits of the Arp2/3 complex.
Figure 5 (facing page). Electron micrographs of negatively stainedor thin-sectioned actin filaments and Arp2/3-induced filament bun-dles. (A and B) Negative stained samples. (C–K) Thin sectionedsamples. Actin alone (A, C). Actin with Arp2/3 complex (B, D–K).Bar in panels A and B is 395 nm. Bar in panels C–K is 395 nm.Conditions were as follows. Negative stained samples: samples ofeither 3 mM Acanthamoeba actin alone or 3 mM actin and 0.5 mMArp2/3 complex were adsorbed to carbon-coated electron micro-scope grids and negatively stained with 1% uranyl formate (seeMATERIALS AND METHODS). Thin sectioned samples: 10 mMAcanthamoeba actin was polymerized in the absence or presence of 1mM Arp2/3 complex. Samples were fixed, stained, embedded inEpon, and thin-sectioned (see MATERIALS AND METHODS).Arp2/3 complex in the negatively stained samples was purified byprofilin:poly-L-proline affinity chromatography and in the thin-sec-tioned samples by conventional ion exchange chromatography.
Arp2/3 Complex
Vol. 9, April 1998 849

data). With 5 nM filament ends in a solution of 15 mMfilamentous actin, 0.5 mM Arp2/3 will cap 4.8 nM ofthe pointed ends, and at 1.5 mM it will cap 4.9 nM.Over the same concentration range, however, the stiff-ness of the filament network increases by more than afactor of 2, suggesting that the network formationactivity cannot be completely explained by end-sideassociations but is also produced by side-side cross-links. The existence of two filament side-binding sitesin addition to the pointed end binding site is alsoconsistent with the fact that three subunits can bechemically cross-linked to actin filaments (Mullins etal., 1997).
Subcellular Localizations of Arp2/3 and a-ActininArp2/3 complex cross-links actin filaments in a man-ner similar to a-actinin, so we wondered if the cellularfunctions of these proteins overlap. One way to ad-dress this question is to determine the extent to whichthe subcellular distributions of the two proteins over-lap. By immunofluorescence microscopy, we find bothArp2/3 and a-actinin enriched in peripheral regionsof cells, but the staining patterns do not overlap com-pletely, particularly at the leading edge (Figure 10).The staining of Arp2/3 subunits is enriched at theleading edge of adherent Acanthamoeba and fibrillar incharacter, while a-actinin staining is diffuse or unde-tectable at the leading edge (Figure 6). We consistentlyobserve that, even in cells with a-actinin staining nearthe leading edge, the staining does not reach theplasma membrane while Arp3, Arp2, p40, and p35staining always extends to the tip of the leading edge.
The fact that Arp2/3 localizes to the tip of the lead-ing edge suggests that it does not passively diffuseinto the region but is involved in the initial events ofleading edge formation. This pattern is consistent withthe recent observation that pseudopodia produced byrapidly polymerizing actin filaments in the tails ofmotile Listeria monocytogenes also do not contain a-ac-tinin (Sechi et al., 1997). One explanation for the ab-sence of a-actinin in such actin-rich structures is thatthey form more rapidly than a-actinin can diffuse intothem. The cellular concentrations of filamentous actinand a-actinin in Acanthamoeba are approximately 100and 5 mM (Gordon et al., 1976; Pollard et al., 1986) andthe Kd of a-actinin binding to actin filaments is 5 mMso at a given time more than 95% of the cellulara-actinin is bound to existing filaments. The dissocia-tion rate constant of Acanthamoeba a-actinin is 5 s21
(Wachsstock et al., 1993a) while the rate of addition ofactin monomers to existing filaments in vivo can be ashigh 1000 s21, so it is possible in a cell for filamentformation to outpace a-actinin redistribution, espe-cially in regions where actin polymerization is drivingmembrane protrusion.
Role of Profilin in Arp2/3 FunctionHow does profilin, which interacts with both actin andArp2, affect the interaction of actin with Arp2/3 com-plex? Arp2/3 complex nucleates barbed end growthof filaments and remains tightly attached to thepointed ends of filaments. Profilin-bound actin mono-mers add to the barbed end of an actin filament withthe same kinetics as free monomers so we would notexpect profilin to affect the ability of actin to bindArp2/3 complex. Arp2/3 binds filament ends withnanomolar affinity (Mullins et al., 1998) and profilinwith micromolar affinity so we would also not expectprofilin to significantly inhibit binding of Arp2/3 toactin filament ends.
The affinity of profilin for its ligands varies consid-erably: submicromolar for actin monomers, an orderof magnitude weaker for Arp2/3 complex, and yetanother order of magnitude weaker for both actindimers and the ends of actin filaments. For barbedends the Kd is estimated at 50 mM (Cooper and Pol-lard, 1985; Kaiser et al., 1986), and for cross-linkedactin dimers it is estimated at 70 mM (Mockrin andKorn, 1983). Cooper and Pollard (1985) speculated thatsteric hindrance from adjacent subunits in a filamentreduces the affinity for profilin. The low affinity ofprofilin for cross-linked actin dimers (Mockrin andKorn, 1983) indicates that a single additional mono-mer could produce such hindrances. If steric con-straints are indeed responsible for the low affinity ofactin filaments and dimers for profilin, then theseconstraints are not as severe in the Arp2/3 complex,suggesting that Arp2 and Arp3 are not packed exactlylike actin subunits at the end of a filament.
Given a Kd of 7 mM, is there enough free profilin inAcanthamoeba to bind a substantial fraction of Arp2/3complex? The concentrations of profilin (Tseng et al.,1984) and unpolymerized actin (Gordon et al., 1976)are about 100 mM each and that of Arp2/3 complex is2 mM (Kelleher et al., 1995). Calculations based onequilibrium binding constants and results from cellfractionation studies (Perelroizen et al., 1994; Vinson,DeLaCruz, Kaiser, Higgs, and Pollard, in preparation;Kaiser, unpublished observations) indicate that only70–80% of cellular profilin is bound to actin. Thus, inthe absence of other competing ligands, a cellular poolof 20–30 mM profilin is available to bind Arp2/3 com-plex. At these concentrations about 75% of the avail-able Arp2/3 complex would be saturated with profi-lin. So, despite its relative weakness, the interactionmay have physiological significance. Arp2 and Arp3bind and hydrolyze ATP, but the exchange of nucle-otide from the Arp2/3 complex is very slow (Mullinsand Kelleher, unpublished observations) so one pos-sible function of profilin binding is to facilitate nucle-otide exchange in the complex.
R.D. Mullins et al.
Molecular Biology of the Cell850

Biological Implications of Filament Cross-linkingActivityAt the leading edge of motile cells we expect Arp2/3complex to nucleate filament formation, cap the fila-ment pointed ends (Mullins and Pollard, unpublisheddata), and bind the sides of filaments (Mullins et al.,1997). These activities are sufficient to form orthogonalnetworks of actin filaments in which the pointed endsare attached to the sides of other filaments in T junc-tions of the type observed at the leading edges of fishkeratocytes (Small, et al., 1995; Svitkina et al., 1997). Asdiscussed above, one side binding and one pointedend binding site are probably insufficient to producethe stiff orthogonal networks observed in our rheo-metric experiments, so there may be an additionalfilament binding site.
Arp2/3 complex is thus capable of promoting poly-merization at the leading edge and attaching newlyformed filaments to a rigid structure, a requirementfor the “Elastic Brownian ratchet” model of polymer-ization-driven motility (Mogilner and Oster, 1996). Asthe actin filaments grow at their membrane-proximalends and drive forward motility (Tilney et al., 1981;Wang, 1985), the bound complex will be translocatedaway from the membrane. This would explain theuniform localization of the complex throughout theactin-rich zone of the leading edge and all along theactin filament tails of motile Listeria (Welch, et al.,1997a). The partitioning of Arp2/3 complex to theleading edge and a-actinin to other parts of theamoeba cortex suggests that other cross-linking pro-teins (with different properties) replace the complex atthe lateral margins of protrusive zones.
Our observations raise several questions for futurestudy: Are the nucleation and filament cross-linkingactivities separable? If so, are they regulated sepa-rately in the cell? Which activity of the complex isessential for cell polarity and motility—nucleation,filament organization, or both? The answers to thesequestions will provide us with a more complete pic-ture of how living cells regulate motility and shapechanges.
ACKNOWLEDGMENTS
We thank members of the Pollard laboratory for technical assis-tance, advice, and helpful discussion, particularly Pam Maupin whoprepared thin sections, Don Kaiser who supplied us with monoclo-nal antibodies, and Mike Ostap who participated in almost everyamoeba preparation. We thank Cynthia Wolberger for use of heranalytical ultracentrifuge and Tom Laue for checking our math. Thiswork was supported by NIH research grant GM-26338 to T.D.P., afellowship to R.D.M. from the Jane Coffin Childs Fund for CancerResearch, and a Jenkins Fellowship to J.X.
REFERENCES
Aebi, U., Fowler, W.E., Isenberg, G., Pollard, T.D., and Smith, P.R.(1981). Crystalline actin sheets: their structure and polymorphism.J. Cell Biol. 91, 340–351.
Balasubramanian, M.K., Feoktistova, A., McCollum, D., and Gould,K.L. (1996). Fission yeast Sop2p: a novel and evolutionarily con-served protein that interacts with Arp3p and modulates profilinfunction. EMBO J. 15, 6426–6437.
Bubb, M.R., Lewis, M.S., and Korn, E.D. (1994). Actobindin bindswith high affinity to a covalently cross-linked actin dimer. J. Biol.Chem. 269, 25587–25591.
Cooper, J.A., and Pollard, T.D. (1985). Effects of capping protein onthe kinetics of actin polymerization. Biochemistry 24, 793–799.
Ferry, J.D. (1980). Viscoelestic Properties of Polymers, 3rd ed., NewYork: John Wiley and Sons.
Frankel, S., and Mooseker, M.S. (1996). The actin-related proteins.Curr. Opin. Cell Biol. 8, 30–37.
Fujiwara, K., and Pollard, T.D. (1976). Fluorescent antibody local-ization of myosin in the cytoplasm, cleavage furrow, and mitoticspindle of human cells. J. Cell Biol. 71, 848–875.
Gordon, D.J., Eisenberg, E., and Korn, E.D. (1976). Characterizationof a cytoplasmic actin isolated from Acanthamoeba castellanii by anew method. J. Biol. Chem. 251, 4778–4786.
Holleran, E.A., Tokito, M.K., Karki, S., Holzbaur, E.L.F. (1996) Cen-tractin (Apr1) associates with spectrin revealing a potential mecha-nism to link dynactin to intracellular organelles. J. Cell Biol. 135,1815–1830.
Kaiser, D.A., Goldschmidt-Clermont, P.J., Levine, B.A., and Pollard,T.D. (1989). Characterization of renatured profilin purified by ureaelution from poly-L-proline agarose columns. Cell Motil. 14, 251–262.
Kaiser, D.A., and Pollard, T.D. (1996). Characterization of actin andpoly-L-proline binding sites of Acanthamoeba profilin with monoclo-nal antibodies and by mutagenesis. J. Mol. Biol. 255, 89–107.
Kaiser, D.A., Sato, M., Ebert, R., and Pollard, T.D. (1986). Purifica-tion and characterization of two isoforms of Acanthamoeba profilin.J. Cell Biol. 102, 221–226.
Kelleher, J.F., Atkinson, S.J., and Pollard., T.D. (1995). Sequences,structural models, and cellular localization of the actin-related pro-teins Arp2 and Arp3 from Acanthamoeba. J. Cell Biol. 131, 385–397.
Kiehart, D.P., Kaiser, D.A., and Pollard, T.D. (1984). Monoclonalantibodies demonstrate limited structural homology between myo-sin isozymes from Acanthamoeba. J. Cell Biol. 99, 1002–1014.
Lynch, T.J., Brzeska, H., Baines, I.C., and Korn, E.D. (1991). Purifi-cation of myosin-I and myosin-I heavy chain kinase. Methods En-zymol. 196, 12–23.
Machesky, L.M. (1997). Cell motility: complex dynamics at theleading edge. Curr. Biol. 7, R164–R167.
Machesky, L.M., Atkinson, S.J., Ampe, C., Vandekerckhove, J., andPollard, T.D. (1994). Purification of a cortical complex containingtwo unconventional actins from Acanthamoeba by affinity chroma-tography on profilin agarose. J. Cell Biol. 127, 107–115.
Maciver, S.K., Wachsstock, D., Schwarz, W.H., and Pollard, T.D.(1991). The actin filament severing protein actophorin catalyzes theformation of rigid bundles of actin filaments crosslinked with alpha-actinin. J. Cell Biol. 115, 1621–1628.
MacLean-Fletcher, S., and Pollard, T.D. (1980). Identification of afactor in conventional muscle actin preparation which inhibits actinfilament self-association. Biochem. Biophys. Res. Commun. 96, 18–27.
Arp2/3 Complex
Vol. 9, April 1998 851

McCollum, D., Feoktistova, A., Morphew, M., Balasubramanian, M.,Gould, K.L. (1996). The Schizosaccharomyces pombe actin-related pro-tein, Arp3, is a component of the cortical actin cytoskeleton andinteracts with profilin. EMBO J. 15, 6438–6446.
Mockrin, S.C., and Korn, E.D. (1983). Kinetics of polymerization andATP hydrolysis by covalently cross-linked actin dimers. J. Biol.Chem. 258, 3215–3221.
Moreau, V., Galan, J.M., Devilliers, G., Haguenauer-Tsapis, R., andWinsor, B. (1997). The yeast actin-related protein Arp2p is requiredfor the internalization step of endocytosis. Mol. Biol. Cell 8, 1361–1375.
Moreau, V., Madania, A., Martin, R.P., and Winsor, B. (1996). TheSaccharomyces cerevisiae actin-related protein Arp2 is involved in theactin cytoskeleton. J. Cell Biol. 134, 117–132.
Mullins, R.D., Kelleher, J.F., and Pollard, T.D. (1996). Actin’ likeactin? Trends Cell Biol. 6, 208–212.
Mullins, R.D., Stafford, W.F., and Pollard, T.D. (1997). Structure,subunit topology, and actin-binding activity of the Arp2/3 complexfrom Acanthamoeba. J. Cell Biol. 136, 331–343.
Mullins, R.D., Heuser, J.A., Pollard, T.D. (1998). The interaction ofArp2/3 complex with actin: nucleation, high-affinity pointed endcapping, and formation of branching networks of filaments. Proc.Natl. Acad. Sci. USA (in press).
Perelroizen, I., Marchand, J.B., Blanchoin, L., Didry, D., and Carlier,M.F. (1994). Interaction of profilin with G-actin and poly(L-proline).Biochemistry 33, 8472–8478.
Pollard, T.D. (1984). Purification of a high molecular-weight actinfilament gelation protein from Acanthamoeba that shares antigenicdeterminants with vertebate spectrins. J. Cell Biol. 99, 1970–1980.
Pollard, T.D., Tseng, P.C.-H., Rimm, D.L., Bichell, D.P., Williams,R.C., and Sinard, J. (1986). Characterization of alpha-actinin fromAcanthamoeba. Cell Motil. 6, 649–661.
Sato, M., Wong, T.Z., Brown, D., and Allen, R.D. (1984). Rheologicalproperties of living cytoplasm: a preliminary investigation of squidaxoplasm. Cell Motil. 4, 7–23.
Schachman, H.K. (1960). Deductions from hydrodynamic and ther-modynamic measurements. Brookhaven Symp. Biol. 13, 49–70.
Schachman, H.K., Gropper, L., Hanlon, S., and Putney, F. (1962).Ultracentrifuge studies with absorption optics. II. Incorporation of amonochrometer and its application to the study of proteins andinteracting systems. Arch. Biochem. Biophys. 99, 175–190.
Sechi, A.S., Wehland, J., Small, J.V. (1997). The isolated comet tailpseudopodium of Listeria monocytogenes: a tail of two actin filament
populations, long and axial and short and random. J. Cell Biol. 137,155–167.
Small, J.V., Herzog, M., and Anderson, K. (1995). Actin filamentorganization in the fish keratocyte lamellipodium. J. Cell Biol. 129,1275–1286.
Spudich, J.A., and Watt, S. (1971). The regulation of rabbit skeletalmuscle contraction. I. Biochemical studies of the interaction of thetropomyosin-troponin complex with actin and the proteolytic frag-ments of myosin. J. Biol. Chem. 246, 4866–4871.
Terasaki, A.G., Ohnuma, M., and Mabuchi, I. (1997). Identificationof actin-binding proteins from sea urchin eggs by F-actin affinitycolumn chromatography. J. Biochem. 122, 226–236.
Tilney, L.G., Bonder, E.M., and DeRosier, D.J. (1981). Actin filamentselongate from their membrane-associated ends. J. Cell Biol. 90,485–494.
Tseng, P.C. H., Runge, M.S., Cooper, J.A., Williams, R.C., Jr., andPollard, T.D. (1984). Physical, immunochemical, and functionalproperties of Acanthamoeba profilin. J. Cell Biol. 98, 214–221.
Vandekerchove, J., Kaiser, D.A., and Pollard, T.D. (1989). Acan-thamoeba actin and profilin can be crosslinked between glutamic acid364 of actin and lysine 115 of profilin. J. Cell Biol. 109, 619–626.
Wachsstock, D.H., Schwarz, W.H., and Pollard, T.D. (1993a). Affin-ity of alpha-actinin for actin filaments determines the structure andmechanical properties of actin filament gels. Biophys. J. 65, 205–214.
Wachsstock, D.H., Schwarz, W.H., Pollard, T.D. (1993b). Structureand mechanical properties of actin filament gels. Biophys. J. 65,205–214.
Wang, Y. (1985). Exchange of actin subunits at the leading edge ofliving fibroblasts: Possible role of treadmilling. J. Cell Biol. 101,597–602.
Welch, M.D., Iwamatsu, A., and Mitchison, T.J. (1997a). Actin po-lymerization is induced by Arp2/3 protein complex at the surface ofListeria monocytogenes. Nature 385, 265–269.
Welch, M.D., Iwamatsu, A., and Mitchison, T.J. (1997b). The humanArp2/3 complex is composed of evolutionarily conserved subunitsand is localized to cellular regions of dynamic actin filament assem-bly. J. Cell Biol. 138, 375–384.
Winter, D., Podtelejnikov, A.V., Mann, M., and Li, R. (1997). Thecomplex containing actin-related proteins Arp2 and Arp3 is re-quired for the motility and integrity of yeast actin patches. Curr.Biol. 7, 519–529.
Yonemura, S.Y., and Pollard, T.D. (1992). Localization of myosin-Iand myosin-II in Acanthamoeba. J. Cell Sci. 102, 629–642.
R.D. Mullins et al.
Molecular Biology of the Cell852




















