
High-resolution Structural Analysis of Mammalian Profilin 2a Complex Formation with Two...
21
High-resolution Structural Analysis of Mammalian Profilin 2a Complex Formation with Two Physiological Ligands: The Formin Homology 1 Domain of mDia1 and the Proline-rich Domain of VASP Petri Kursula 1,2 ⁎, Inari Kursula 1 , Marzia Massimi 3 , Young-Hwa Song 1 Joshua Downer 3 , Will A. Stanley 1 , Walter Witke 3 and Matthias Wilmanns 1 1 EMBL Hamburg Outstation, DESY, Notkestrasse 85, D-22607 Hamburg, Germany 2 Department of Biochemistry, University of Oulu, Oulu, Finland 3 EMBL Monterotondo Outstation, Monterotondo, Italy Received 25 June 2007; received in revised form 15 October 2007; accepted 17 October 2007 Available online 24 October 2007 Profilins are small proteins capable of binding actin, poly-L-proline and other proline-rich sequences, and phosphatidylinositol (4,5)-bisphosphate. A number of proline-rich ligands for profilin have been characterised, including proteins of the Ena/VASP and formin families. We have deter- mined the high-resolution crystal structures of mouse profilin 2a in complex with peptides from two functionally important ligands from different families, VASP and mDia1. The structures show that the binding mode of the peptide ligand is strongly affected by the non-proline residues in the sequence, and the peptides from VASP and mDia1 bind to profilin 2a in distinct modes. The high resolution of the crystallographic data allowed us to detect conserved CH-π hydrogen bonds between the peptide and profilin in both complexes. Furthermore, both peptides, which are shown to have micromolar affinity, induced the dimerisation of profilin, potentially leading to functionally different ligand-profilin–actin complexes. The peptides did not significantly affect actin polymerisation kinetics in the presence or in the absence of profilin 2a. Mutant profilins were tested for binding to poly-L-proline and the VASP and mDia1 peptides, and the F139A mutant bound proline-rich ligands with near-native affinity. Peptide blotting using a series of designed peptides with profilins 1 and 2a indicates differences between the two profilins towards proline-rich peptides from mDia1 and VASP. Our data provide structural insights into the mechanisms of mDia1 and VASP regulated actin polymerisation. © 2007 Elsevier Ltd. All rights reserved. Edited by I. Wilson Keywords: profilin; actin; crystal structure; mDia1; VASP Introduction The actin microfilament system has a central role in a large number of cellular processes, such as lamel- lipodial protrusion and the formation of filopodia. A number of proteins interacting with the barbed ends of microfilaments regulate the elongation and stab- ility of the filaments, and thus, are involved in cell motility. Profilins are a family of small 15 kDa proteins that sequester monomeric actin and function in processes related to microfilament nucleation and polymerisation. 1 Profilins bind monomeric actin, and they interact with phosphatidylinositol (4,5)- *Corresponding author. E-mail address: [email protected]. Present addresses: I. Kursula, Department of Medical Biochemistry and Biophysics, Karolinska Institute, Stockholm, Sweden; W. Stanley, ARC COE Plant Energy Biology, University of Western Australia, Crawley, Australia; J. Downer, Irrational Games Boston, Quincy, MA, USA. Abbreviations used: mDia1, mammalian homologue 1 of Drosophila diaphanous; VASP, vasodilator-stimulated phosphoprotein; PtdIns(4,5)P 2 , phosphatidylinositol (4,5)-bisphosphate; EVH, Ena/VASP homology; WH, WASPhomology; WASP, Wiscott-Aldrich syndrome protein; FH, formin homology; ITC, isothermal titration calorimetry; DID, Diaphanous inhibitory domain; DAD, Diaphanous autoinhibitory domain. doi:10.1016/j.jmb.2007.10.050 J. Mol. Biol. (2008) 375, 270–290 Available online at www.sciencedirect.com 0022-2836/$ - see front matter © 2007 Elsevier Ltd. All rights reserved.
-
Upload
uni-hamburg -
Category
Documents
-
view
0 -
download
0
Transcript of High-resolution Structural Analysis of Mammalian Profilin 2a Complex Formation with Two...

doi:10.1016/j.jmb.2007.10.050 J. Mol. Biol. (2008) 375, 270–290
Available online at www.sciencedirect.com
High-resolution Structural Analysis of MammalianProfilin 2a Complex Formation with Two PhysiologicalLigands: The Formin Homology 1 Domain of mDia1and the Proline-rich Domain of VASP
Petri Kursula1,2⁎, Inari Kursula1, Marzia Massimi3, Young-Hwa Song1
Joshua Downer3, Will A. Stanley1, Walter Witke3
and Matthias Wilmanns1
1EMBL Hamburg Outstation,DESY, Notkestrasse 85,D-22607 Hamburg, Germany2Department of Biochemistry,University of Oulu, Oulu,Finland3EMBL MonterotondoOutstation, Monterotondo, Italy
Received 25 June 2007;received in revised form15 October 2007;accepted 17 October 2007Available online24 October 2007
*Corresponding author. E-mail [email protected] addresses: I. Kursula, Depa
Biochemistry and Biophysics, KarolinStockholm, Sweden; W. Stanley, ARCBiology, University ofWestern AustraJ. Downer, Irrational Games Boston, QAbbreviations used: mDia1, mam
of Drosophila diaphanous; VASP, vasphosphoprotein; PtdIns(4,5)P2, phos(4,5)-bisphosphate; EVH, Ena/VASPWASP homology; WASP, Wiscott-Aprotein; FH, formin homology; ITC,calorimetry; DID, Diaphanous inhibDiaphanous autoinhibitory domain.
0022-2836/$ - see front matter © 2007 E
Profilins are small proteins capable of binding actin, poly-L-proline andother proline-rich sequences, and phosphatidylinositol (4,5)-bisphosphate.A number of proline-rich ligands for profilin have been characterised,including proteins of the Ena/VASP and formin families. We have deter-mined the high-resolution crystal structures of mouse profilin 2a in complexwith peptides from two functionally important ligands from differentfamilies, VASP and mDia1. The structures show that the binding mode ofthe peptide ligand is strongly affected by the non-proline residues in thesequence, and the peptides from VASP and mDia1 bind to profilin 2a indistinct modes. The high resolution of the crystallographic data allowed usto detect conserved CH-π hydrogen bonds between the peptide and profilinin both complexes. Furthermore, both peptides, which are shown to havemicromolar affinity, induced the dimerisation of profilin, potentiallyleading to functionally different ligand-profilin–actin complexes. Thepeptides did not significantly affect actin polymerisation kinetics in thepresence or in the absence of profilin 2a. Mutant profilins were tested forbinding to poly-L-proline and the VASP and mDia1 peptides, and the F139Amutant bound proline-rich ligands with near-native affinity. Peptideblotting using a series of designed peptides with profilins 1 and 2a indicatesdifferences between the two profilins towards proline-rich peptides frommDia1 and VASP. Our data provide structural insights into the mechanismsof mDia1 and VASP regulated actin polymerisation.
© 2007 Elsevier Ltd. All rights reserved.
Edited by I. Wilson
Keywords: profilin; actin; crystal structure; mDia1; VASPess:
rtment of Medicalska Institute,COE Plant Energylia, Crawley, Australia;uincy, MA, USA.
malian homologue 1odilator-stimulatedphatidylinositolhomology; WH,
ldrich syndromeisothermal titrationitory domain; DAD,
lsevier Ltd. All rights reserve
Introduction
The actinmicrofilament systemhas a central role ina large number of cellular processes, such as lamel-lipodial protrusion and the formation of filopodia. Anumber of proteins interacting with the barbed endsof microfilaments regulate the elongation and stab-ility of the filaments, and thus, are involved in cellmotility.Profilins are a family of small 15 kDa proteins
that sequester monomeric actin and function inprocesses related to microfilament nucleation andpolymerisation.1 Profilins bind monomeric actin,and they interact with phosphatidylinositol (4,5)-
d.

271Profilin 2a Complexed with VASP and mDia1
bisphosphate (PtdIns(4,5)P2).2–5 An intensively stu-
died group of ligands for profilins is comprised ofpoly-L-proline6 and sequences in natural ligandshaving proline-rich segments.1 A growing numberof profilin ligands have been characterised that har-bour one or more profilin-binding segments.1,7–13
Such proteins include, for example, members of theEna/VASP and formin families. Two well charac-terised profilin ligands from these families are thevasodilator-stimulated phosphoprotein (VASP)12
and the mammalian homologue 1 of Drosophila dia-phanous (mDia1), respectively (Figure 1).13 WhileVASP functions in focal adhesions, lamellipodia,filopodia, and immune receptor signalling,14,15 theformins are involved in the formation of filopodia,the determination of cell polarity, and cytokinesis.16
The two best studied profilin isoforms, profilin 1and profilin 2, are generated from different genes. Inaddition, two splice forms of profilin 2, termed pro-filins 2a and 2b, were described.17,18 Profilin 2a isexpressed at high levels in the brain in bothmice andhumans.19 Differences in ligand binding specificityhave been observed between the different pro-filins.1,18,20,21 A common feature of profilin-bindingmotifs is the presence of at least five consecutive Proresidues, often flanked by hydrophobic residues.The polyproline binding site of profilin consists
of conserved aromatic residues22,23 that lie on theopposite face from that interacting with actin.24,25 Inline with this, profilin is able to bind both actin andproline-rich ligands simultaneously.26 Polyprolinebinding involves mostly residues from the first andlast α helix of profilin, which form a specific, pre-formed, surface that can accommodate a type IIpolyproline helical segment from the ligand. Both
sequence of the FH1 domain of mDia1, which has 13 highly hpeptide used in this study, containing two binding motifs, is u
hydrophobic interactions and hydrogen bondsbetween profilin and the ligand are importantfor binding.27 In yeast, a functional polyprolinebinding site in profilin is required for cell viability.28
VASP is a homologue of the Drosophila proteinEnabled that binds to profilin via its proline-richregion.9 It is a tetrameric protein having a proline-rich profilin-binding domain and flanking Ena/VASP homology (EVH) domains 1 and 2.29 TheEVH2 domain can further be divided into threesubdomains: a G-actin binding Wiscott-Aldrich syn-drome protein (WASP) homology (WH) 2 domain,an F-actin binding domain, and a C-terminaltetramerisation domain. The structure of the VASPC-terminal tetramerisation motif30 is a right-handedcoiled coil based on a 15 residue repeat.31 VASPbinds with high affinity to profilin 2a, and does notinteract strongly with profilin 1.20 While the profilin-binding region is dispensable for normal fibroblastmovement,32 and thus is not required for VASPfunction within lamellipodia, it seems to be impor-tant for the formation of filopodia in Dictyostelium.33
Formation of a complex between VASP and profilinwas found in the movement of the intracellularbacteria Listeria.34–38 The bacteria bind VASP ontheir surface, which in turn interacts with profilin,suggesting that profilin recruits actin to the bacterialsurface. Profilin recruitment to the bacterial surfaceis proportional to intracellular Listeria speed.36
mDia1 belongs to the formin family and, inaddition to the formin homology (FH) domains 1and 2, which are common to all formins, it containsan N-terminal region, which has been referred to asan FH3 domain, and a C-terminal diaphanous auto-inhibitory domain (DAD). The so-called FH3 do-
Figure 1. The profilin ligandsused in the current study. (a) Adiagram of the structures of theprofilin 2a ligands, mDia1 andVASP. For mDia1, the domains areas follows: GBD, Rho-GTPase-bind-ing domain; DID, diaphanous inhi-bitory domain; DD, dimerisationdomain; CC, coiled-coil domain;FH1, formin homology domain 1;FH2, formin homology domain 2;DAD, diaphanous autoinhibitorydomain. The regions N-terminal tothe FH1 domain have been referredto in the literature as the FH3 do-main. For VASP, the EVH1 domain,the proline-rich domain, and theEVH2 domain are indicated. TheEVH2 domain contains a WH2domain (dark grey), an F-actinbinding domain (light grey), and atetramerisation domain (white). Thesequences of the peptides used inthis study are shown below theprofilin-binding domains. (b) The
omologous proline-rich repeats arranged in tandem. Thenderlined.
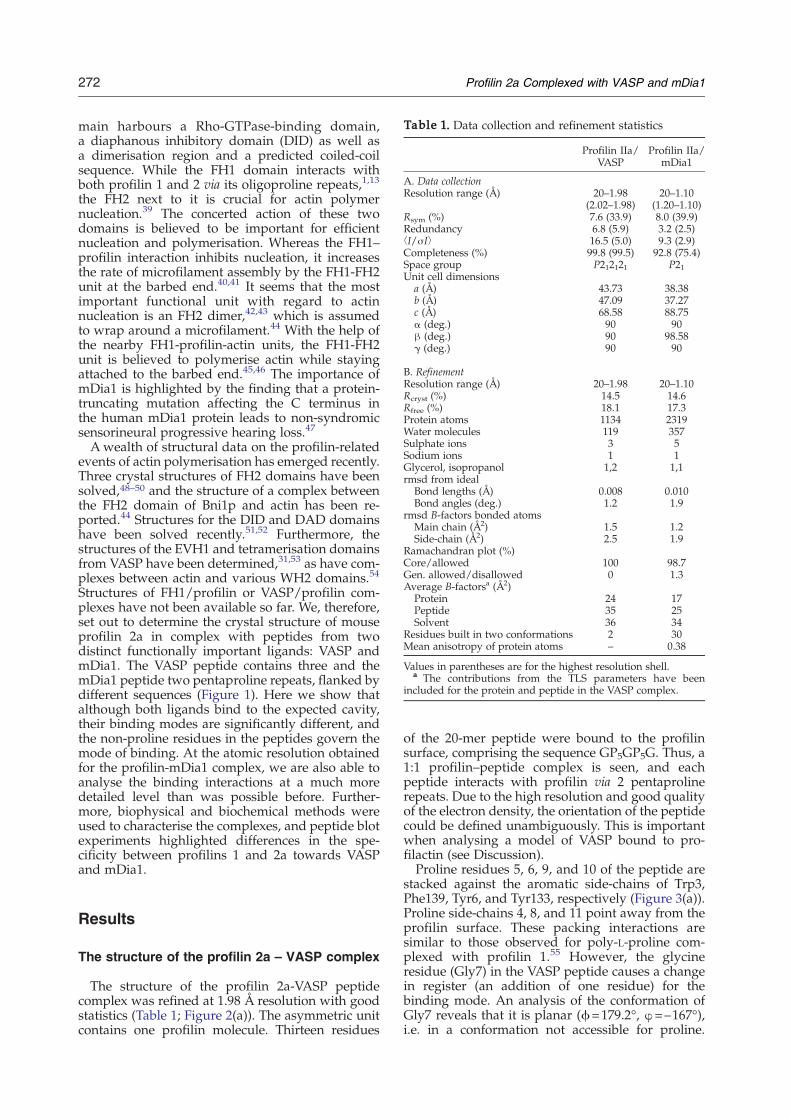
Table 1. Data collection and refinement statistics
Profilin IIa/VASP
Profilin IIa/mDia1
A. Data collectionResolution range (Å) 20–1.98
(2.02–1.98)20–1.10
(1.20–1.10)Rsym (%) 7.6 (33.9) 8.0 (39.9)Redundancy 6.8 (5.9) 3.2 (2.5)⟨I/σI⟩ 16.5 (5.0) 9.3 (2.9)Completeness (%) 99.8 (99.5) 92.8 (75.4)Space group P212121 P21Unit cell dimensions
a (Å) 43.73 38.38b (Å) 47.09 37.27c (Å) 68.58 88.75α (deg.) 90 90β (deg.) 90 98.58γ (deg.) 90 90
B. RefinementResolution range (Å) 20–1.98 20–1.10Rcryst (%) 14.5 14.6Rfree (%) 18.1 17.3Protein atoms 1134 2319Water molecules 119 357Sulphate ions 3 5Sodium ions 1 1Glycerol, isopropanol 1,2 1,1rmsd from ideal
Bond lengths (Å) 0.008 0.010Bond angles (deg.) 1.2 1.9
rmsd B-factors bonded atomsMain chain (Å2) 1.5 1.2Side-chain (Å2) 2.5 1.9
Ramachandran plot (%)Core/allowed 100 98.7Gen. allowed/disallowed 0 1.3Average B-factorsa (Å2)
Protein 24 17Peptide 35 25Solvent 36 34
Residues built in two conformations 2 30Mean anisotropy of protein atoms – 0.38
Values in parentheses are for the highest resolution shell.a The contributions from the TLS parameters have been
included for the protein and peptide in the VASP complex.
272 Profilin 2a Complexed with VASP and mDia1
main harbours a Rho-GTPase-binding domain,a diaphanous inhibitory domain (DID) as well asa dimerisation region and a predicted coiled-coilsequence. While the FH1 domain interacts withboth profilin 1 and 2 via its oligoproline repeats,1,13
the FH2 next to it is crucial for actin polymernucleation.39 The concerted action of these twodomains is believed to be important for efficientnucleation and polymerisation. Whereas the FH1–profilin interaction inhibits nucleation, it increasesthe rate of microfilament assembly by the FH1-FH2unit at the barbed end.40,41 It seems that the mostimportant functional unit with regard to actinnucleation is an FH2 dimer,42,43 which is assumedto wrap around a microfilament.44 With the help ofthe nearby FH1-profilin-actin units, the FH1-FH2unit is believed to polymerise actin while stayingattached to the barbed end.45,46 The importance ofmDia1 is highlighted by the finding that a protein-truncating mutation affecting the C terminus inthe human mDia1 protein leads to non-syndromicsensorineural progressive hearing loss.47
Awealth of structural data on the profilin-relatedevents of actin polymerisation has emerged recently.Three crystal structures of FH2 domains have beensolved,48–50 and the structure of a complex betweenthe FH2 domain of Bni1p and actin has been re-ported.44 Structures for the DID and DAD domainshave been solved recently.51,52 Furthermore, thestructures of the EVH1 and tetramerisation domainsfrom VASP have been determined,31,53 as have com-plexes between actin and various WH2 domains.54
Structures of FH1/profilin or VASP/profilin com-plexes have not been available so far. We, therefore,set out to determine the crystal structure of mouseprofilin 2a in complex with peptides from twodistinct functionally important ligands: VASP andmDia1. The VASP peptide contains three and themDia1 peptide two pentaproline repeats, flanked bydifferent sequences (Figure 1). Here we show thatalthough both ligands bind to the expected cavity,their binding modes are significantly different, andthe non-proline residues in the peptides govern themode of binding. At the atomic resolution obtainedfor the profilin-mDia1 complex, we are also able toanalyse the binding interactions at a much moredetailed level than was possible before. Further-more, biophysical and biochemical methods wereused to characterise the complexes, and peptide blotexperiments highlighted differences in the spe-cificity between profilins 1 and 2a towards VASPand mDia1.
Results
The structure of the profilin 2a – VASP complex
The structure of the profilin 2a-VASP peptidecomplex was refined at 1.98 Å resolution with goodstatistics (Table 1; Figure 2(a)). The asymmetric unitcontains one profilin molecule. Thirteen residues
of the 20-mer peptide were bound to the profilinsurface, comprising the sequence GP5GP5G. Thus, a1:1 profilin–peptide complex is seen, and eachpeptide interacts with profilin via 2 pentaprolinerepeats. Due to the high resolution and good qualityof the electron density, the orientation of the peptidecould be defined unambiguously. This is importantwhen analysing a model of VASP bound to pro-filactin (see Discussion).Proline residues 5, 6, 9, and 10 of the peptide are
stacked against the aromatic side-chains of Trp3,Phe139, Tyr6, and Tyr133, respectively (Figure 3(a)).Proline side-chains 4, 8, and 11 point away from theprofilin surface. These packing interactions aresimilar to those observed for poly-L-proline com-plexed with profilin 1.55 However, the glycineresidue (Gly7) in the VASP peptide causes a changein register (an addition of one residue) for thebinding mode. An analysis of the conformation ofGly7 reveals that it is planar (ϕ=179.2°, φ=–167°),i.e. in a conformation not accessible for proline.

Figure 2. Overall structures of the profilin-peptide complexes. (a) VASP. (b) mDia1. Top, structure of the profilin-peptide complex; middle, 2Fo–Fc electron density at 1σ of the bound peptide; bottom, surface of the binding cavity. Thearomatic residues of the binding surface are coloured as follows: Trp, green; Phe, magenta; Tyr, yellow.
273Profilin 2a Complexed with VASP and mDia1
Furthermore, the proline residue next to this glycine,Pro6, is stacked against the C-terminal Phe139 ofprofilin, and is the only Pro residue in the peptidethat does not have a polyproline type II helicalconformation (Table 2).Four direct hydrogen bonds are seen between the
peptide and profilin (Figure 3(a)). All of them are
formed between side-chains of conserved residuesin profilin and main-chain oxygen atoms of thepeptide. The carbonyl oxygen atoms of Pro3, Gly7,Pro8, and Pro10 from the peptide form hydrogenbonds to the profilin residues Trp3, Tyr6, Tyr133,and Asn9, respectively. What is more, no watermolecules lie between the peptide and the protein.

Figure 3. Binding site determinants. (a) Stereo view of the binding site for the VASP peptide, showing hydrogen bondinteractions and several residues discussed in the text. (b) Superposition of the peptide binding modes from the VASP(proline, orange; glycine, red) and mDia1 (proline, green; non-proline, blue) complexes. The N termini of the peptidespoint right in the Figure.
274 Profilin 2a Complexed with VASP and mDia1
The peptide in the complex also interacts with asymmetry-related profilin molecule, as well as withthe peptide bound to the latter. For example, Pro11 isstacked against Arg107 from the symmetry-relatedmolecule, which forms a neutralising salt-bridgewith Asp26. A hydrogen bond is present betweenArg107 and the carbonyl oxygen from Pro9. Bothpeptides interact with Phe139 of one monomer, sug-gesting a role for this residue in peptide-mediateddimerisation.
The structure of the mDia1 complex
The profilin 2a-mDia1 peptide complex structurewas determined at atomic (1.10 Å) resolution (Figure2(b)). The asymmetric unit contains two profilinmolecules, and one pentaproline repeat of the pep-tide is well defined in both monomers. In monomerA, the sequence IPPPPPL is seen in the electrondensity, and in the slightly better defined B mono-mer, the visible peptide is IPPPPPLPG. Five residues(PGVAS) connecting the two halves were not builtdue to weak electron density. The distance betweenthe Cα atoms of Leu7 and Ile13 of the peptide is 12 Å,a distance easily spanned by a disordered patchof five residues. This part of the peptide is not incontact with either of the protein subunits nor is itinvolved in crystal contacts, and is therefore veryflexible. The terminal carboxyl group is very well
defined in the electron density for the glycineresidue (Figure 2(b)), proving that it is the secondrepeat of the peptide that is seen in monomer B. Thedetailed analysis below relates to the repeat seen inmonomer B; the binding mode in monomer A ishighly similar.The Ile and Leu residues of the mDia1 repeat act as
anchors at both ends of the pentaproline segment.The side-chain of the Ile packs tightly against Tyr31,while the side-chain of the Leu packs against Tyr6.Thus, the mDia1 pentaproline repeat is actuallypositioned differently from the pentaproline in theVASP peptide (Figure 3(b)). Intriguingly, only two ofthe Pro residues of the pentaproline in the mDia1peptide are stacked against the aromatic residues inthe binding surface. This involves the third andfourth proline of the repeat and profilin residuesTrp3 and Phe139.Three hydrogen bonds exist between the peptide
and profilin (Figure 4(a)); these are formed betweenthe side-chains of Trp3, Tyr6, and Tyr133, and thebackbone oxygen atoms of Pro2/14, Pro5/17, andPro6/18, respectively. Asn9 does not form a hydro-gen bond to the peptide in this case. Again, water isexcluded from the binding interface.In the putative peptide-mediated dimer (see
below), the interactions between the profilin mono-mers do not seem specific, with few direct protein–protein contacts via hydrogen bonds. The buried

Table 2. Phi/Psi angles of the peptides and theirinteractions with profilin
Peptide Residue ϕ (°) φ (°) H-bond CH-π bond
A. VASPGly1 −55.4 166.1Pro2 −81.6 167.2 Trp31Pro3 −64.4 147.6 Trp3 Trp31Pro4 −67.3 158.6Pro5 −65.3 155.4 Trp3 (2)Pro6 −68.5 −23.3 Phe139Gly7 179.2 −167.0 Tyr6Pro8 −66.6 156.1 Tyr133Pro9 −66.4 171.9 Tyr6 (Asn9)Pro10 −66.2 162.7 Asn9 Tyr133Pro11 −58.7 141.3Pro12 −67.3 148.0Gly13 −89 145
B. mDia1Ile1 51.3 140.9 Trp31Pro2 −76.1 157.7 Trp3 (Trp31)Pro3 −71.4 146.0Pro4 −80.0 162.6 Trp3 (2)Pro5 −71.4 157.1 Tyr6 (Phe139)Pro6 −65.4 148.5Leu7 −87.0 −41.5 Tyr6Ile13 119.8 120.1 Trp31Pro14 −74.8 150.5 Trp3 Trp31Pro15 −60.4 140.0Pro16 −76.0 164.1 Trp3 (2)Pro17 −75.9 160.1 Tyr6 Phe139Pro18 −65.0 148.4 Tyr133Leu19 −84.7 168.0 Tyr6Pro20 −60.1 152.9Gly21 101.2 44.8
H-bonds concern the backbone oxygen of the peptide and aprofilin side-chain. CH-π bonds concern CH groups from thepeptide and aromatic systems from profilin.
275Profilin 2a Complexed with VASP and mDia1
surface area is small, and nearly all of the contactsare water-mediated. It, thus, seems feasible that bothprofilin monomers bind their respective repeatsfrom the peptide rather independently.
Details of profilin–peptide interaction: CH-πhydrogen bonding and anisotropic rubbingagainst the aromatic planes
The very high resolution of the crystallographicdata for the mDia1 complex allowed accurate refine-ment of the peptide conformation and the subse-quent addition of hydrogen atoms to their expectedpositions. Thus, the putative occurrence of CH-πhydrogen bonds,56,57 involving CH groups from thepeptide and the π systems of the aromatic residuesin profilin, in the binding interface was analysed.Indeed, such hydrogen bonds can be detected for allof the aromatic residues involved in peptide bind-ing; furthermore, in tryptophan residues, suchbonds are observed for both of the ring structuresin the side-chain (Figure 4(a)). Interestingly, the Ileand Leu residues stacked against Trp31 and Tyr6also form these bonds.A similar analysis was carried out for the 1.98 Å
structure of the VASP complex; all of the peptide–aromatic stacking interactions seem to involve CH-π
bonds (Figure 4(b)). Although the energy for onesuch bond is relatively low, the clustering of theseinteractions together with the hydrogen bondsformed between the peptide backbone carbonylsand the profilin side-chains is likely to provide animportant portion of the binding energy. In spite ofthe different binding modes for the two peptides, asseen in the crystal structures, the conservation ofthese weak interactions for both peptides gives anexplanation for the similar energetics of binding, asmeasured by isothermal titration calorimetry (ITC)(see below).Since anisotropic atomic displacement parameters
were refined for the mDia1 complex, the anisotropyof the peptide and the profilin residues interactingwith it could be analysed. It is clear that there is noapparent concerted motion that would involve boththe binding surface and the ligand; the aromaticresidues of profilin are rather isotropic and rigid. Aclear trend exists for those atoms in the peptidethat are involved in CH-π bonding: the longest axisof their anisotropy ellipsoid is always orientedparallel with the plane of the correspondingaromatic side-chain in profilin (Figure 4(c)). Thus,the CH-π distance stays more or less constantdespite the mobility of the peptide atoms. Thisanisotropic rubbing against the aromatic planes bythe CH groups from the peptide provides evidencefor the importance of CH-π interactions in profilin-ligand binding.
Analysis of peptide binding affinity by wild-typeprofilin 2a by isothermal titration calorimetry
The thermodynamic parameters for profilin 2abinding to the VASP and mDia1 peptides weredetermined by ITC (Figure 5(a)). ITC analysis ofprofilin 2a titrated with the peptides gave a strongexothermic response for both peptides. The numberof binding sites corresponds to a molar ratio(peptide:profilin) of 0.5 for both peptides. The deter-mined parameters are given in Table 3. It is note-worthy that the parameters for the two peptides arehighly similar, with favourable enthalpy ΔH andunfavourable entropy ΔS components, despite theirdifferent modes of binding. This reflects the obser-vation that in the two binding modes, the overalltypes and amounts of profilin-peptide interactions,including both conventional H-bonds as well asCH-π bonds, are very similar.
Dimerisation of profilin 2a upon peptide binding
Dimerisation of profilin upon binding a peptidefrom VASP has been observed.20,58 Poly-L-prolineinduces dimerisation,58 but the resulting complexbehaved differently from the VASP peptide complexin a functional assay, suggesting an important func-tional role for the glycine residues within theprofilin-binding domain of VASP.Gel-filtration chromatography (Figure 5(b)) indi-
cated that in addition to the VASP peptide, themDia1 peptide caused a quantitative shift of the

Figure 4. A detailed view of the complexes. (a) Details of peptide binding in the profilin–mDia1 complex. The threeconventional hydrogen bonds between the protein and the peptide are shown by broken green lines. Black lines indicateputative CH-π hydrogen bonds. (b) The profilin-VASP complex. Hydrogen bonds and CH-π interactions as in (a). (c) Astereo view of the anisotropy at the binding site for the mDia1 complex. The thermal ellipsoids are contoured at aprobability of 30%. Note that the apparent major direction of movement for the ligand atoms engaged in CH-πinteractions is parallel with the plane of the corresponding aromatic residue, resulting in the conservation of the distancebetween the anisotropic atoms and the aromatic plane.
276 Profilin 2a Complexed with VASP and mDia1
apparent molecular mass of profilin 2a from 15 kDato approximately 35 kDa, clearly indicative ofdimerisation. Profilin 2a without peptide behavedas a monomeric 15 kDa protein. On the basis of thegel-filtration and ITC experiments, it can be con-cluded that the profilin–peptide complexes have 2:1stoichiometry in solution.Since dimerisation of profilin 2a was observed
with both peptides, the crystal structures werefurther analysed to visualise the peptide-induceddimers. As no large-scale change occurs in theprofilin molecule upon peptide binding, we con-cluded that the dimers are, to a large extent, peptide-mediated. In the VASP complex crystal, it is clearthat one peptide molecule is bound to one profilinmolecule, and thus, a profilin dimer would have tobe a 2:2 complex in the crystal state. A neighbouringsymmetry molecule is found in the crystal so thateach peptide interacts with two profilin molecules;Phe139 seems to be central to this interaction.Another explanation is that the excess peptide andthe crystallisation conditions have broken theVASP–profilin complex to have 1:1 stoichiometry,
instead of the 1:2 complex apparently present insolution.Since a 2:1 complex between profilin 2a and the
mDia1 peptide was observed, possible peptideconnections in the corresponding crystal latticewere sought to resolve the functional dimer. Theonly possible connection is such that the peptidebound in the A monomer comprises the firstpentaproline sequence of the peptide, and thepeptide in the B monomer comprises the secondone. Although the electron density for the linkerregion between the two binding repeats is weak,it is highly likely that the same peptide is boundto both of the profilin monomers in the asym-metric unit. All other connections of peptides inthe lattice would lead either to distances that aretoo long, infinite peptides, or a change of peptidebackbone orientation. In addition, the tight pack-ing of the crystal lattice excludes the possibilitythat two separate peptides would be bound tothe profilin monomers; there is simply no roomfor the extra residues within the lattice in thiscase.

Figure 5. Size-exclusion chromatography and calorimetric analysis of the interaction. (a) ITC analysis of profilin–peptide complex formation. Left, VASP; right, mDia1. (b) Gel-filtration chromatography. In the absence of peptide,profilin 2a has an apparent molecular mass of 15 kDa (thick line), while in the presence of both the VASP (thin line) andmDia1 peptides (broken line), all profilin elutes at 35 kDa.
277Profilin 2a Complexed with VASP and mDia1
Binding assays with profilin mutants
The profilin 2a point mutants Y29S, Y133S, S137D,and F139Awere generated, and their ability to bindpoly-L-proline was assessed by a pull-down assay(Figure 6(a)). While S137D and Y133S did not bind,
the ability of F139A to bind poly-L-proline wasimpaired only slightly. Y29S also bound to poly-L-proline. ITC experiments using the mDia1 and VASPpeptides indicated a significant decrease in bindingaffinity upon the Y133S and S137D mutations, whilethe F139A mutant bound the peptides with a near-

Table 3. ITC analysis of the profilin IIa – peptideinteractions
Peptide/profilinmutant n
Kd(μM)
ΔH(kcal/mol)
−TΔS(kcal/mol)
ΔG(kcal/mol)
VASP/wt 0.5 24.3 −12.4 6.0 −6.4mDia1/wt 0.5 16.5 −13.2 6.6 −6.6VASP/F139A 0.5 23.8 −1.1 −5.3 −6.4mDia1/F139A 0.5 28.1 −4.1 −2.2 −6.3VASP/Y133S 0.5 920 −6.2 2.0 −4.2mDia1/Y133S 0.5 540 −2.3 −2.2 −4.5
The S137D mutant was included in the assay, but no binding ofeither peptide was observed.
278 Profilin 2a Complexed with VASP and mDia1
native affinity (Table 3). However, the enthalpy forbinding was low and the entropy was favourable,which is opposite to the results obtained using thewild-type protein.Phe139 is one of the aromatic residues packing
against the x-Pro units in profilin ligands (see above),and one of the two proline residues in mDia1packing into the binding site is stacked againstPhe139. The observation that the F139A mutantbinds poly-L-proline as well as the VASP and mDia1peptides with nearly the same affinity as the wild-type protein suggests that Phe139 is not crucial forthe interaction. However, the energetics of the inter-
Figure 6. Functional assays. (a) Binding of poly-L-prolinepresence of profilin 2a (P2) and the VASP and mDia1 peptide
action are completely different, in that binding seemsto be driven by large favourable entropy rather thanenthalpy as in wild-type profilin 2a. The increase inentropy when compared to the wild-type complexcan be rationalised, to some extent, by the fact thatPhe139 seems to force the peptide into a strainedconformation in the complexes, and it is likely thatthe peptide is more flexible in the F139A complex. Inthe crystal structure, especially the region of theVASP peptide interactingwith Phe139 is in a strainedconformation, and the difference in energetics islarger for the VASP peptide when using the F139Amutant. It is also possible that hydrogen bondingbetween profilin and the peptides is affected by theF139A mutation indirectly, which would explain thedecreased enthalpy observed in ITC. A minor effectwill surely come from the loss of the CH-π bondbetween the aromatic ring of Phe139 and the pep-tides, and from the entropy related to fixing the con-formation of the large side-chain of Phe139 in thewild-type protein upon peptide binding.Ser137 can be phosphorylated by protein kinase C
in profilin 1.59 In the present structures of profilin 2a,Ser137 is close in space to the bound peptides; e.g.the distances between its Cβ atom and the atomsCβ(Pro8) in mDia1 and Cγ(Pro6) in VASP are 3.7 Åand 3.6 Å, respectively. Furthermore, its Oγ atom is
by the profilin mutants. (b) Actin polymerisation in thes.

279Profilin 2a Complexed with VASP and mDia1
hydrogen bonded to the backbone oxygen of Tyr133,one of the crucial aromatic residues in the bindingsite. Since the S137D mutant does not bind poly-L-proline or the peptides, it is evident that phosphor-ylation at this position should abrogate proline-richligand binding by profilin. In line with this, the samemutant also fails to bind the ligand dynamin 1.8
Since the Y133S mutant, analogous to the well-characterised H133S mutation in profilin 1,60,61 isunable to bind dynamin 1,8 its failure to bind poly-L-proline and the peptides was expected. For bothpeptides, the determined ΔΔG (mutant versus wild-type) is approximately 2 kcal/mol (Table 3). Tyr133is located at the edge of the binding site, and it maybe that its mutation alters the conformation of the C-terminal helix. In the VASP complex, Tyr133 stacksagainst Pro10 of the peptide, and there is a hydrogenbond between its hydroxyl group and the backboneoxygen of Pro9. In the mDia1 complex, the Tyr133hydroxyl group also hydrogen bonds to main-chainoxygen atoms (Pro18 in the B subunit and Pro6 inthe A subunit), but no stacking interaction with thepeptide is observed; in fact, a molecule of isopropa-nol located at a crystal contact area stacks againstTyr133 forming a CH-π interaction.An interesting residue with regard to ligand
binding by profilin 2a is Tyr29, which is serine inprofilin 1. It has been suggested that Tyr29 wouldenable a longer binding surface for ligands.62 Tyr29is not in a central position in the binding site, whichexplains the ability of Y29S, a mutation mimickingprofilin 1 at this position, to bind poly-L-proline inthe pulldown assay. Tyr29 has no direct interactionwith the peptides, but it is in a position to possiblyinteract with the central linker of the mDia1 peptide.In essence, it defines the end of the binding site at theN-terminal side of the peptide without strong inter-actions towards the peptides. The loop leadingtowards Tyr29 (25-CDAKY-29) is the region inprofilin with the greatest difference between thetwo peptide complexes.
Effects of the VASP and mDia1 peptides onin vitro actin polymerisation
We used an in vitro assay to investigate the effectsof the peptides on skeletal muscle α-actin polymer-isation in the presence and absence of profilin 2a(Figure 6(b)). As expected and shown before, in thepresence of profilin 2a, actin monomers are seques-tered and actin polymerisation is inhibited.63 Addi-tion of the mDia1 or VASP peptides does notattenuate the inhibition by profilin 2a, nor do thepeptides by themselves have a significant stimula-tory effect on polymerisation. Previously, the VASPpeptide was shown to overcome the inhibition ofactin nucleation brought about by the concertedaction of profilin and thymosin β4.58
Peptide blotting
The FH1 domain of mDia1 contains 13 tandemlyarranged repeats that are highly similar to each
other in sequence; the obvious question waswhether each repeat is able to bind profilin. A seriesof peptides from the mDia1 FH1 domain weresynthesised on a SPOT membrane to analyse theinteraction of profilin 2a with this repetitive region(Figure 7; Table 4). Another comparison was carriedout between profilin isoforms, including profilin 1 inthe assay.The results using single repeats from the FH1
domain (Figure 7(a)) indicate that both profilins areable to bind most of the repeats; the exceptions con-cern repeats that have less than five consecutiveproline residues. In line with this, shortening theproline stretch further also resulted in no detectablebinding (Figure 7(b)).A series of tandem double repeats was also used
in the assay, corresponding to the 12 possiblearrangements from the mDia1 FH1 domain (Figure7(f)). Again, binding to all double repeats wasobserved with both profilins. However, the specifi-cities of the two profilins are somewhat different:while profilin 1 prefers the double repeats from themiddle region of the FH1 domain, profilin 2a bindsbetter than profilin 1 to double repeats from the Nand C-terminal ends of the FH1 domain.Using the mDia1 peptide as a basis, a series of
mutant peptides were used in which either theN-terminal pentaproline region or the C-terminalpentaproline region, or both, was successivelyshortened. The results clearly indicate that the muta-tion of one pentaproline region in the double repeatpeptide still allows for binding of both profilins,while the mutation of both binding sites abrogatesbinding (Figure 7(c)–(e)).The peptides used for the other experiments were
included in the series (Figure 7(f)), and while profilin1 binds somewhat more strongly to the mDia1peptide, the VASP peptide is a better ligand forprofilin 2a. VASP has been shown to be one of theligands specific for profilin 2a,12 while mDia1 isknown to bind both these profilins.13
Discussion
Profilin is an important regulator of the actinmicrofilament system, linking the control of nuclea-tion and polymerisation of microfilaments to avariety of proline-rich ligands it binds. Profilinbinding to VASP is a key event, for example, inthe regulation of actin networks in filopodia andlamellipodia64 and Listeria intracellular move-ment,34 while by interactions with mDia1, profilinis involved in the regulation of microfilament nu-cleation39 and the acceleration of elongation.41 Pro-filin binding to its polyproline-containing ligandsrequires only a short sequence motif in the ligand,usually comprised of a pentaproline segmentflanked by hydrophobic residues, as indicated byour experimental data.Although a large number of studies have been
carried out with regard to the role of profilintogether with its ligands in actin polymerisation,

Figure 7. Peptide blots with labelled profilins 1 and 2a. (a) Single repeats from the mDia1 FH1 domain. (b) Shortenedproline stretches. (c) Double repeats with the N-terminal repeat mutated. (d) Double repeats with the C-terminal repeatmutated. (e) Double repeats with both mutated. (f) The 12 possible tandem repeats from the mDia1 FH1 domain, plus thepeptides used in the experiments (13, mDia1; 14, VASP). Peptide 15 is identical with peptide 5. Below, a graph is shownwhere the intensities from densitometry have been compared by dividing the intensity for profilin 2a by the profilin 1intensity. It is clear that compared to profilin 1, profilin 2a has a higher affinity towards the terminal repeats of the mDia1FH1 domain, and towards the VASP peptide.
280 Profilin 2a Complexed with VASP and mDia1
the exact mode of action is rather unclear. Profilinalone inhibits polymerisation by sequestering freeG-actin.65,66 It also promotes nucleotide exchange inactin.63 When bound to ligands, it has varyingeffects, e.g. the formin family proteins use profilin asa tool to nucleate and processively polymerise actinmicrofilaments.46 It is likely that the proline-richligands of profilin target profilin–actin to largersignalling assemblies, such as the Arp2/3, mDia1, orWAVE complexes, in order to facilitate actin filamentnucleation and elongation.Profilin 2a-deficient mice display neurological and
behavioural abnormalities. Profilin 2a, being, incontrast to profilin 1, localised also in the presynap-tic matrix, is required for synaptic actin polymerisa-tion, and its absence in mouse leads to increasednovelty-seeking behaviour.67 Recent studies suggestthat one role of profilin is to aid the transport ofactin from the nucleus to the cytoplasm,68 and thatprofilin can be targeted to neuronal nuclei.69
Structural data on profilin–ligand complexes
A number of crystal structures of profilins 1 and 2fromdifferent organisms have been reported.24,62,70,71
These structures include a complex with monomericactin,72 as well as complexes of profilin 1with boundpoly-L-proline peptides.27,55 It has been showncrystallographically that, like SH3 domains, profilinsare able to bind proline-rich substrates in twoopposite backbone orientations.55 However, animportant shortcoming of all structural and mostbiochemical studies on profilin–proline-rich ligandinteractions to date is that the natural ligands ofprofilin contain specific binding sequences, usuallyconsisting of a pentaproline sequence flanked byhydrophobic non-proline residues. On many occa-sions, a number of such binding modules are foundwithin the same protein in close vicinity to eachother. For example, the mouse mDia1 proteincontains 13 tandem repeats arranged within itsFH1 domain (Figure 1). Furthermore, no structurehas been available for profilin 2 in complex withpeptides, while experimental evidence suggests thatprofilin 2 is the isoform with the higher affinity forpolyproline sequences.20 The crystal structure ofhuman profilin 2b has been determined,62 but thisisoform has low affinity towards poly-L-proline.17,18
The only crystal structure of profilin 2a so far is fromAcanthamoeba.71 Our structures represent the first

Table 4. Peptides used in the profilin overlay assay
Position Sequence Notes
A1. IPPPPPPPPLPGGV Single repeat 1A2. VPPSPPLPPGTC Single repeat 2A3. IPPPPPLPGGAC Single repeat 3A4. IPPPPQLPGSAA Single repeat 4A5. IPPPPPLPGVAS Single repeat 5A6. IPPPPPLPGATA Single repeat 6A7. IPPPPPLPGATA Single repeat 7A8. IPPPPPLPGGTG Single repeat 8A9. IPPPPPPLPGSVG Single repeat 9A10. VPPPPPLPGGPG Single repeat 10A11. LPPPPPPFPGAP Single repeat 11A12. GIPPPPPGMG Single repeat 12A13. VPPPPPFGFGV Single repeat 13B1. IPPPLQLPGSAA Single repeat 4
mutatedB2. IPPVLQLPGSAA Single repeat 4
mutatedB3. IPCVLQLPGSAA Single repeat 4
mutatedB4. IPPPPQLVGSAA Single repeat 4
mutatedC1. IPPPPQLPGVASIPPPPPLPG tandem repeatC2. IPPPLQLPGVASIPPPPPLPG N-term mutatedC3. IPPVLQLPGVASIPPPPPLPG N-term mutatedC4. IPCVLQLPGVASIPPPPPLPG N-term mutatedC5. IPPPPPLVGVASIPPPPPLPG N-term mutatedD1. IPPPPPLPGVASILPPPPLPG C-term mutatedD2. IPPPPPLPGVASILQPPPLPG C-term mutatedD3. IPPPPPLPGVASILQVPPLPG C-term mutatedD4. IPPPPPLPGVASILQVGPLPG C-term mutatedD5. IPPPPPLPGVASIPPPPPLVG C-term mutatedE1. IPPPPQLPGVASILPPPPLPG both mutatedE2. IPPPLQLPGVASILQPPPLPG Both mutatedE3. IPPVLQLPGVASILQVPPLPG Both mutatedE4. IPCVLQLPGVASILQVGPLPG Both mutatedE5. IPPPPPLVGVASIPPPPPLVG Both mutatedF1. IPPPPPPPPLPGGVVPPSPPLPPGTC Tandem repeat
1–2F2. VPPSPPLPPGTCIPPPPPLPGGAC Tandem repeat
2–3F3. IPPPPPLPGGACIPPPPQLPGSAA Tandem repeat
3–4F4. IPPPPQLPGSAAIPPPPPLPGVAS Tandem repeat
4–5F5. IPPPPPLPGVASIPPPPPLPGATA Tandem repeat
5–6F6. IPPPPPLPGATAIPPPPPLPGATA Tandem repeat
6–7F7. IPPPPPLPGATAIPPPPPLPGGTG Tandem repeat
7–8F8. IPPPPPLPGGTGIPPPPPPLPGSVG Tandem repeat
8–9F9. IPPPPPPLPGSVGVPPPPPLPGGPG Tandem repeat
9–10F10. VPPPPPLPGGPGLPPPPPPFPGAP Tandem repeat
10–11F11. LPPPPPPFPGAPGIPPPPPGMG Tandem repeat
11–12F12. GIPPPPPGMGVPPPPPFGFGV Tandem repeat
12–13F13. IPPPPPLPGVASIPPPPPLPG mDia1 peptideF14. GPPPPPGPPPPPGPPPPPGL VASP peptideF15. IPPPPPLPGVASIPPPPPLPGATA Identical with F5
Series A, peptides having single repeats from the FH1 domain ofmDia1. Series B, mutated single repeat 4. Series C, double repeatswith the N-terminal site mutated. Series D, double repeats withthe C-terminal site mutated. Series E, double repeats with bothsites mutated. Series F, all possible 12 tandem double repeats frommDia1 and the VASP and mDia1 peptides used in the otherexperiments in this study.
281Profilin 2a Complexed with VASP and mDia1
high-resolution structural data on mammalian pro-filin 2a.A comparison of the binding modes of the VASP
and mDia1 peptides to the poly-L-proline peptidecomplexed with profilin 1 is shown in Figure 8. Thebinding mode of the mDia1 peptide resembles thepolyproline peptide, with the exception that its Ileand Leu residues interact intimately with thebinding site, superimposing well on proline residuesfrom the polyproline peptide. For the VASP peptide,the pentaproline segment is positioned differentlyfrom that in mDia1. In addition, the Gly residuebetween the pentaproline segments causes a changein register with respect to the other peptides; inessence, one more peptide residue is inserted intothe binding site, causing an apparently strainedconformation. Comparing all three peptides, thehydrogen bonds between the peptide backboneoxygen atoms and the profilin side-chains are con-served, as are the CH-π interactions between thearomatic rings and the residues stacking againstthem. The combination of these types of H-bonds iscommon for other domains binding to proline-richligands, such as SH3 domains.73,74
Binding mode and affinity
Here, we have shown that the binding modesbetween profilin 2a and peptides from its twoligands, VASP and mDia1, differ considerably bothfrom each other and from the binding mode of poly-L-proline to profilin 1.27,55 It is clear from our struc-tures that the non-proline residues, specific for eachligand, have a major impact on the fine details of thebinding mode. In this sense, it is rather surprising
Figure 8. Superposition of the current structures andthe earlier structure of profilin 1 complexed with poly-L-Pro. (a) Superposition of the VASP peptide (VASP, green;poly-L-Pro, orange). (b) Superposition of the mDia1 pep-tide (mDia1, cyan; poly-L-Pro, orange).

Figure 9 (legend on next page)
282 Profilin 2a Complexed with VASP and mDia1

283Profilin 2a Complexed with VASP and mDia1
that the thermodynamic data for the interaction ofprofilin 2a with both of the peptides are virtuallyidentical. The dissociation constants are in the rangeobserved previously for profilin and proline-richpeptides75 and, interestingly, for the interactions ofSH3 domains with peptide ligands, where a typicalKd value is around 10 μM. This is related to bothprofilin and SH3 domains being involved indynamic interactions that must not be too tight inthe cell. Furthermore, considering the clear dif-ferences in the peptide-binding modes in our twocomplexes, it is surprising how small the differencesin the protein are in the different liganded andunliganded structures of profilins in general; thebinding site for the proline-rich ligands seems to bepre-arranged.Combining the results of gel-filtration chroma-
tography and ITC, it can be concluded that insolution, both profilin 2a-peptide complexes have a2:1 stoichiometry. The crystal structures presentedhere allow us to propose a model for the complexwhere profilin binds both a proline-rich ligandand actin (Figures 9(a) and 10(a)). Obviously, themodel is similar to that suggested before.27 Con-sidering the locations of the actin-binding surfaces inthe structures, the orientations of actin molecules inthe predicted complexes are significantly different,in the sense that actin molecules modelled on theprofilin-mDia1 dimer are on the same side of theprofilin dimer.For both of the peptide ligands, the energetics of
the interaction were similar, the binding beingdriven by a large favourable enthalpy coupled tounfavourable entropy contributions. We concludethat the favourable thermal energy comes from theformation of three or four strong hydrogen bondsbetween the peptide and profilin. Since it is assumedthat no large-scale rearrangement in profilin occursupon ligand binding, the unfavourable entropy canbe attributed to the folding of the disordered pep-tides into a polyproline type II helix upon binding,and the rigidification of the profilin residues directlyinvolved in binding. Comparing with the resultsobtained for Acanthamoeba profilin 1 bound to dif-ferent peptides,75 the affinity of profilin 2a towardsits specific physiological ligands is slightly higherthan that of Acanthamoeba profilin 1 to L-Pro11. Itshould be noted that the Acanthamoeba profilin 1corresponds more closely to mammalian profilin 2and the Acanthamoeba profilin 2 is more similar tomammalian profilin 1. Replacing the middle Pro inthe L-Pro11 peptide byGly or Ala lowered the affinityof Acanthamoeba profilin 1 towards the peptide toeightfold;75 this peptide is nearly identical with thefraction of the VASP peptide well-defined in our
Figure 9. Models of profilin function via interaction with Vpeptide. The peptide is in green, profilin is in grey, and actin isand (b) and in Figure 10(a). (b) A model showing the binding s(green), and PtdIns(4,5)P2 (sulphate to the left) on profilacSupplementary Data. (c) A drawing of the function of tetramtheir assembly. The EVH1, proline-rich, WH2, F-actin bindingVASP.
complex structure (GP5GP5G). In this respect, it isinteresting to note that VASP binds with higheraffinity to mammalian profilin 2a than to profilin 1,as seen in our peptide blotting experiments andearlier studies.20 Apparently,Acanthamoeba profilin 1does not tolerate a Pro-Gly substitution in themiddleof the binding sequence.
Details of peptide – aromatic residueinteractions
CH-π hydrogen bonds are often detected foraromatic residues in proteins, also in interactionsbetween proline-rich peptides and their bindingsites.74 In the structures described here, eacharomatic residue of the peptide-binding site isinvolved in such an interaction. Although theenergy of a single CH-π hydrogen bond is low,in the range of 1.0 kcal/mol, the combinedfavourable enthalpy effect from the bonds formedupon profilin-peptide complex formation will besignificant compared to the overall enthalpy ofbinding. Looking at the anisotropically refinedatomic displacement parameters from the mDia1complex, it seems clear that the interaction is suchthat an optimal distance for CH-π bonds ismaintained due to the rubbing of the peptide in adirection parallel with each aromatic plane in thebinding site. Such an interaction could be impor-tant also for other proteins binding proline-richsequences.
The PtdIns(4,5)P2 binding site
Profilin 1 binds PtdIns(4,5)P2 with higher affinitythan profilin 2.20 Mutagenesis studies have beenused to map the PtdIns(4,5)P2 binding site(s) onhuman profilin 1.76 Two binding sites wereidentified, around Arg135 and Arg88, but the func-tional binding site is comprised of a larger surfaceharbouring a number of conserved basic residuesand a positive electrostatic potential. Chemicalcrosslinking of phosphatidylinositol analogues toprofilin 1 suggests an interaction of PtdIns(4,5)P2with the N-terminal region of profilin as well.77
Furthermore, PtdIns(4,5)P2 binding seems to com-pete with binding to both actin and proline-rich sequences,4,20 suggesting a large interactionsurface. However, when covalently crosslinked toactin, profilin I is capable of binding to PtdIns(4,5)P2 micelles,5 indicating that the initial bindingbefore the dissociation of profilin from actinmay happen distal from the actin-binding site,and tight binding involving Arg88 takes place onlysubsequently.
ASP. (a) A model of a profilin dimer mediated by the VASPin orange. The subdomains I–IVof actin are labelled in (a)ites for the VASP proline-rich domain (blue), WH2 domaintin. An animated version of the model is available aseric VASP in crosslinking microfilaments and promoting(FAB), and tetramerisation (TM) domains are shown for

284 Profilin 2a Complexed with VASP and mDia1
Interestingly, in the VASP complex, three sul-phate ions are bound to the putative PtdIns(4,5)P2-binding site (Figure 9(b)); binding of sulphate (orphosphate) to the region around Arg88 has beenobserved for both profilin 1 and profilin 2b.62 Inthe VASP complex, the residues coordinating thesulphate groups include Arg104, Arg135, Lys132,Arg88, and Arg74. Also, in both profilin monomers
Figure 10. Models of mDia1 interacting with profilactin. (repeat peptide. The peptide (green, with the missing linker instructure, and actin (orange) has been added by superpositionFH1 domain where each repeat binds one profilactin complexgrey, and actin orange. The arrangement should be feasible forgrowing microfilament. (c) A view of the function of mDia1 FHdimeric FH2 domain moves along the actin filament, while thend. It may not be necessary for the FH2 domain to move aheadbinding sites for profilactin on each FH1 domain.
of the mDia1 complex, a sulphate is boundbetween Arg135 and Arg104, as well as nearArg107. The putative PtdIns(4,5)P2 binding site,filled by these sulphate groups may mimic thethree phosphate groups of PtdIns(4,5)P2. It islocated on the opposite side from the expectedWH2 domain binding site in profilactin (Figure9(b)).
a) A model for a profilactin dimer mediated by a doublemagenta) and profilin (grey) are from the complex crystalfrom the profilactin structure. (b) A model of the mDia1. 4 repeats are shown. The FH1 domain is green, profilinthe rapid incorporation of several actin monomers onto the1 and FH2 domains in microfilament polymerisation. Thee FH1 domain repeats provide profilactin for the growingwith each added actin molecule, due to the presence of 13

Figure 10 (legend on previous page)
285Profilin 2a Complexed with VASP and mDia1
Models for the function of VASP and mDia1
For both VASP and mDia1, structural informationis accumulating that can be used to understand theirfunctional mechanisms in regulating the microfila-ment system. We have now added two significantdetails to this view: the structures of profilin 2acomplexed with its binding sequences in VASP andmDia1. Both structures support the view that onerole for profilin is to provide a high local concen-tration of monomeric actin at the barbed end ofthe microfilament. It is possible that one dimer ofmDia1 is able to recruit a maximum of 26 profilin/profilactin units (Figure 10(b) and (c)), and that onetetramer of VASP can bind eight such units (Figure9(c)). In both mDia1 and VASP, the profilin-bindingdomain is next to a domain binding both monomericand filamentous actin directly (FH2 and EVH2domains, respectively).Our actin polymerisation assays revealed that the
peptides had no significant effect on the kinetics ofactin polymerisation in the presence or absence ofprofilin 2a (Figure 6(b)). This implies that it is notsufficient to bring two profilactin complexes intoclose vicinity to affect actin polymerisation kinetics.In other words, complex formation of actin andprofilin with the peptides is not sufficient to providenucleation activity. Our findings are consistent withthe idea that complex formation of profilactin withthe proline-rich domains of VASP and mDia1 isprimarily a mechanism to recruit and orient actin
monomers in the full-length molecule, while thenucleation activity is provided by domains of theVASP and mDia1 molecule. Once a growing micro-filament has been formed, the accurately orientedVASP and mDia1 proteins, binding to microfila-ments via their respective F-actin binding domains,are then efficiently recruiting profilactin units to thegrowing end in the correct orientation, such thatelongation is supported.The structure of actin complexed with the WH2
domain of WASP has been solved recently,54 and ahomologous globular actin-binding domain residesin VASP C-terminal to the proline-rich region, asa part of its EVH2 domain.78 On the basis of ourVASP–profilin complex structure, the structure ofprofilactin, and the structure of the actin–WH2complex, a model can be built of a complex of pro-filactin with VASP profilin-rich and WH2 domains(Figure 9(b)). The orientation of the peptide inour structure is such that the C terminus of theproline-rich domain is close to the binding site ofthe WH2 domain on actin. The observed sulphategroups in the structure are on the opposite sideof the complex, locating the PtdIns(4,5)P2 bindingsite.The current model of mDia1 action (and formin
action in general) as a processive capper describesmDia1 “walking” along the growing barbed endof the microfilament via its FH2 domain.48,79 Twomonomers of mDia1 are bound to opposite sites ofthe microfilament in this model (Figure 10(c)).44 TheFH1 domain, with 13 adjacent binding sites forprofilactin with a short flexible conserved segmentbetween them, lies immediately N-terminal to theFH2 domain, and thus, a maximum of 26 G-actinmolecules can in theory be brought to the vicinity ofthe growing end by the mDia1 dimer. It has beenshown that the presence of five profilin-binding sitesin the mDia1 FH1 domain results in the sameelongation rate as the full FH1 domain, while thelack of profilin-binding sites reduces the rate sig-nificantly.41 The spacing of one profilactin bindingsite to the next is expected to be around 30 Å on thebasis of our structure, assuming no specific interac-tion between adjacent profilin molecules. Thisarrangement, with a very high local concentrationof monomeric actin present, may allow for the FH2domain to move more rapidly, while keeping closeto the barbed end. It is possible that the interactionsof the profilactin complex with the barbed endgenerate force to help bring the trailing FH2 domainalong the barbed end.
Materials and Methods
Profilin purification
Mouse profilins 1 and 2a, containing the full nativesequence, were produced without tag using the phageT7-polymerase-driven expression vector pMW17280 andpurified by affinity chromatography.81 On the average,10–50 mg of N95% pure protein could be purified from 2 l

286 Profilin 2a Complexed with VASP and mDia1
of Escherichia coli culture after overnight induction with0.5 mM IPTG. The cell pellet was resuspended in 50 ml ofbuffer A (50 mM Tris–HCl (pH 7.3), 5 mM EGTA, 0.1 mMEDTA, 50 mM KCl, 8 M urea, 10 mM DTT), and cells werelysed by sonication. Debris was removed by centrifuga-tion at 100,000g for 1 h, and the cleared supernatant wasdialysed overnight against 2 l of 4 M urea buffer. Theconcentration of urea was successively reduced to zero inbuffer B (50 mM Tris–HCl (pH 7.3), 1 mM EGTA, 0.1 mMEDTA, 50 mM KCl, 1 mM DTT), and finally, the lysatewas dialysed one more time against buffer B. Precipitatewas removed by centrifugation at 100,000g and thesupernatant loaded onto a poly-L-proline column asdescribed.82 The column was washed extensively with1 M urea, and profilin was eluted from the column withbuffer A. Concentrated fractions were pooled anddialysed against decreasing concentrations of urea, andfinally, against several changes of buffer B.
Purification of GST-tagged profilin 2a point mutants
Profilin 2a point mutants were expressed using thepGEX3 vector system (GE Healthcare). Exponentiallygrowing E. coli cultures were induced with 1 mM IPTGfor 6 h. Cells were harvested and resuspended in lysisbuffer (20 mMTris–HCl (pH 7.5), 1 mM EGTA, 1mMDTT,1% (v/v) Triton X-100, 10% (w/v) sucrose). Cells werelysed by sonication, the lysate cleared, and recombinantprofilins purified on gluthatione Sepharose 4B (GEHealthcare), according to the manufacturer's instructions.The GST tag was removed by thrombin cleavage (2 units/mg) overnight at +16 °C, and after removing excessglutathione by dialysis, the recombinant protein wasagain passed over glutathione Sepharose 4B. Mutantprofilins were collected from the flowthrough.
Peptides
The synthetic peptides corresponding to the proline-richdomain of VASP (GPPPPPGPPPPPGPPPPPGL) and atandem double repeat (pentaproline repeats 5 and 6)from the FH1 domain of mDia1 (IPPPPPLPGVA-SIPPPPPLPG) were obtained from Genemed Synthesis.
Crystallisation and data collection
For crystallisation, profilin 2a (10 mg/ml in 10 mMHepes, pH 7.5) was mixed with either the mDia1 orVASP peptide, in a molar ratio of 1:4. Crystals weregrown by the hanging-drop, vapour-diffusion method.The optimised well solution for profilin-VASP crystal-lisation contained 2.4 M ammonium sulphate and 5%(v/v) isopropanol, while that for profilin-mDia1 con-tained 2.2 M ammonium sulphate and 5% isopropanol.Crystals grew to a maximum size of approximately0.2 mm in a few days.Before data collection, the crystals were passed quickly
through a cryoprotectant solution containing 20% (v/v)glycerol in addition to the well solution components. Thecrystals were flash-cooled to 100 K in a stream of nitrogengas. X-ray diffraction data were collected at BESSY(Berlin), on the PSF beamline BL1. For the mDia1 complex,a high-resolution and low-resolution data set werecollected separately. Data processing, scaling, and reduc-tion were performed using XDS83 and XDSi;84 5% (VASP)or 2% (mDia1) of all reflections were randomly set asidefor cross validation.
Crystal structure solution and refinement
Molecular replacement was carried out with AMoRe,85
using the unliganded structure of human profilin 2b (PDBentry 1D1J62) as the model. The profilin–VASP complexwas fully refined using Refmac5,86 applying TLS para-meters.87 An initial atomic-resolution model of theprofilin-mDia1 complex was also refined with Refmac5.Model analysis and manual rebuilding were carried outusing O88 and coot.89 The bound peptides were clearlydefined in the first electron density maps after molecularreplacement in both complexes, and the well-defined partswere built in at an early stage. Water molecules wereadded using ARP/wARP.90 At late stages of refinement,anisotropic B-factors were refined for all atoms in theprofilin-mDia1 structure.91
The profilin-mDia1 structure was further refined usingSHELXL (G. Sheldrick, University of Göttingen), whichresulted in better statistics and electron density maps,showing more details such as extended double conforma-tion networks, and some weak electron density for thelinker region in the mDia1 peptide. The initial model fromthe Refmac5 refinement was first refined isotropically,adding high-resolution data gradually as the refinementproceeded. In the later stages, anisotropic refinement wasperformed and several double conformations, includingextended networks involving both protein and solventatoms, were built. At the end of the refinement, protonswere added at the riding positions. Standard restraintswere applied throughout the refinement. Most parts of theelectron density maps are well defined. Only five residues(PGVAS) in themiddle of themDia1 peptide are not visible,and a profilin loop region from Gln92 to Glu95 (QGGE) inthe A monomer is not clearly defined due to the lack ofcrystal contacts. As a last step, the contents of theasymmetric unit were re-arranged for easier analysis, andanisotropic B-factor refinement was run in phenix.refine.92
Anisotropy analysiswas then carried out usingPARVATI.93
Isothermal titration calorimetry
The interaction between profilin 2a and the mDia1 andVASP peptides was analysed using ITC, on a MicrocalVP-ITC instrument. Before titration, both profilin 2a andthe peptides were dialysed exhaustively against 10 mMHepes (pH 7.5). Then, 0.3 mM profilin 2a was titrated with2 mM peptide at +30 °C, and data analysis was carried outusing the Microcal Origin software package. The mutantsS137D and F139Awere similarly analysed at a concentra-tion of approximately 100 μM. The peptide concentrationin this case was 1 mM. The Y133S mutant was analysed at0.3 mM, using 3 mM peptide in the titration.
Gel-filtration chromatography
Profilin was mixed with tenfold molar excess amountsof peptide and separated on a Sephadex 75 10/30 columnon an Äkta system (GE Healthcare), using 10 mM Hepes(pH 7.5), 100 mM NaCl as the running buffer. A controlexperiment was run in the absence of ligand peptides.Profilin elution was monitored by measuring absorbanceat 280 nm.
Actin polymerisation assays
Actin was purified from rabbit skeletal muscle asdescribed.94 Actin polymerisation assays were performed

287Profilin 2a Complexed with VASP and mDia1
in a 100 μl total reaction volume containing 5 μM G-actinspiked with 10% pyrene-labelled G-actin.63,95 Polymerisa-tion was started by adding a final concentration of 10 mMimidazole, 2 mMMgCl2, and 1mMEGTA. Polymerisationkinetics were monitored at +25 °C in 20 s intervals using afluorescence plate reader (Fluoroskan Ascent II, Labsys-tems) with excitation at 342 nm and emission reading at388 nm. Before the reaction was started, 5 μM profilin 2aand/or 10 μM VASP or mDia1 peptide in G-buffer (2 mMTris–HCl (pH 8.0), 0.2 mM CaCl2, 0.2 mM ATP, 0.01%(w/v) NaN3) was added. The relative fluorescenceincrease was plotted versus time. For each polymerisationcondition (n=8), the standard error was calculated.
Poly-L-proline pulldown assay
Purified wild-type profilin 2a and the mutant profilinsY29S, Y133S, S137D, and F139A (20 μg each) were mixedwith 20 μg of bovine serum albumin as an internal control,brought up to a final volume of 150 μl in 20 mM Tris–HCl(pH 7.5), 1 mM EDTA, 1 mM DTT, and incubated with50 μl of poly-L-proline beads for 1 h. Beads and super-natants were collected after a brief centrifugation at 700g.The beads were washed twice and then boiled in SDSsample buffer. Equivalent amounts of bound andunbound fractions were analyzed by SDS-PAGE in orderto determine the capacity of the profilin mutants to bind topoly-L-proline.
Overlay of peptide membranes with fluorescentlylabelled profilins
Purified profilins 1 and 2awere concentrated to approxi-mately 0.5 mM and dialysed against the labelling buffer(100 mMNa-bicarbonate, pH 8.0, 100 mMNaCl). A 3-foldmolar excess of Alexa Fluor 488 2,3,5,6-tetrafluorophenylester (Invitrogen) in dimethyl formamide was added forlabelling, and the reaction was stopped by separating thelabelled protein from the free dye by passage through asmall desalting column. The labelling efficiency wasestimated to be approximately 50% by recording an ab-sorption spectrum between 250 nm and 650 nm.A series of peptides from the FH1 domain of mDia1 and
the VASP proline-rich region were synthesised on a SPOTmembrane. A list of peptides is given in Table 4. For theassay, the membrane was blocked with 1% bovine serumalbumin in PBS-T for 30 min, after which the membranewas washed twice with PBS-T. Then, Alexa-profilin at afinal concentration of 26 μMwas added and incubated for1 h at room temperature. The membrane was washedthree times with PBS-T, and the signals were detectedusing a fluorescence imager.
Protein Data Bank accession codes
The coordinates and structure factors were deposited inthe PDB under the accession codes 2V8C (the VASPcomplex) and 2V8F (the mDia1 complex).
Acknowledgements
The authors thank the excellent support at theBESSY/PSF beamline BL1, and Frank Lehmann is
acknowledged for technical assistance. P.K. wassupported by a postdoctoral fellowship from the ECgrant CAMKIN (HPRN-CT-2002-00252) to M.W. I.K.was supported by a postdoctoral fellowship fromthe HFSP grant RGP0062/2002-C to M.W. Thisstudy has also been supported financially by theAcademy of Finland (to P.K. and I.K.) and theHelsingin Sanomat 100 Years Jubileum Foundation(to P.K.).
Supplementary Data
Supplementary data associated with this articlecan be found, in the online version, at doi:10.1016/j.jmb.2007.10.050
References
1. Witke, W. (2004). The role of profilin complexes in cellmotility and other cellular processes. Trends Cell Biol.14, 461–469.
2. Lambrechts, A., van Damme, J., Goethals, M., Vande-kerckhove, J. & Ampe, C. (1995). Purification andcharacterization of bovine profilin II. Actin, poly(L-proline) and inositolphospholipid binding. Eur. J.Biochem. 230, 281–286.
3. Machesky, L. M., Goldschmidt-Clermont, P. J. &Pollard, T. D. (1990). The affinities of human plateletand Acanthamoeba profilin isoforms for polyphosphoi-nositides account for their relative abilities to inhibitphospholipase C. Cell Regul. 1, 937–950.
4. Lassing, I. & Lindberg, U. (1985). Specific interactionbetween phosphatidylinositol 4,5-bisphosphate andprofilactin. Nature, 314, 472–474.
5. Skare, P. & Karlsson, R. (2002). Evidence for twointeraction regions for phosphatidylinositol(4,5)-bisphosphate on mammalian profilin I. FEBS Letters,522, 119–124.
6. Tanaka, M. & Shibata, H. (1985). Poly(L-proline)-binding proteins from chick embryos are a profilinand a profilactin. Eur. J. Biochem. 151, 291–297.
7. Boukhelifa, M., Moza, M., Johansson, T., Rachlin, A.,Parast, M., Huttelmaier, S. et al. (2006). The proline-rich protein palladin is a binding partner for profilin.FEBS J. 273, 26–33.
8. Gareus, R., Di Nardo, A., Rybin, V. &Witke, W. (2006).Mouse profilin 2 regulates endocytosis and competeswith SH3 ligand binding to dynamin 1. J. Biol. Chem.281, 2803–2811.
9. Kang, F., Laine, R. O., Bubb, M. R., Southwick, F. S. &Purich, D. L. (1997). Profilin interacts with the Gly-Pro-Pro-Pro-Pro-Pro sequences of vasodilator-stimu-lated phosphoprotein (VASP): implications for actin-based Listeria motility. Biochemistry, 36, 8384–8392.
10. Krebs, A., Rothkegel, M., Klar, M. & Jockusch, B. M.(2001). Characterization of functional domains ofmDia1, a link between the small GTPase Rho andthe actin cytoskeleton. J. Cell Sci. 114, 3663–3672.
11. Miyagi, Y., Yamashita, T., Fukaya, M., Sonoda, T.,Okuno, T., Yamada, K. et al. (2002). Delphilin: a novelPDZ and formin homology domain-containing pro-tein that synaptically colocalizes and interacts withglutamate receptor delta 2 subunit. J. Neurosci. 22,803–814.
12. Reinhard, M., Giehl, K., Abel, K., Haffner, C., Jarchau,T., Hoppe, V. et al. (1995). The proline-rich focal

288 Profilin 2a Complexed with VASP and mDia1
adhesion and microfilament protein VASP is a ligandfor profilins. EMBO J. 14, 1583–1589.
13. Watanabe, N., Madaule, P., Reid, T., Ishizaki, T.,Watanabe, G., Kakizuka, A. et al. (1997). p140mDia, amammalian homolog of Drosophila diaphanous, is atarget protein for Rho small GTPase and is a ligand forprofilin. EMBO J. 16, 3044–3056.
14. Rottner, K., Behrendt, B., Small, J. V. & Wehland, J.(1999). VASP dynamics during lamellipodia protru-sion. Nature Cell Biol. 1, 321–322.
15. Sechi, A. S. &Wehland, J. (2004). ENA/VASP proteins:multifunctional regulators of actin cytoskeletondynamics. Front Biosci. 9, 1294–1310.
16. Faix, J. & Grosse, R. (2006). Staying in shape withformins. Dev. Cell. 10, 693–706.
17. Di Nardo, A., Gareus, R., Kwiatkowski, D. & Witke,W. (2000). Alternative splicing of the mouse profilin IIgene generates functionally different profilin iso-forms. J. Cell Sci. 113, 3795–3803.
18. Lambrechts, A., Braun, A., Jonckheere, V., Aszodi, A.,Lanier, L. M., Robbens, J. et al. (2000). Profilin II isalternatively spliced, resulting in profilin isoformsthat are differentially expressed and have distinctbiochemical properties. Mol. Cell. Biol. 20, 8209–8219.
19. Su, A. I., Cooke, M. P., Ching, K. A., Hakak, Y., Walker,J. R., Wiltshire, T. et al. (2002). Large-scale analysis ofthe human andmouse transcriptomes. Proc. Natl Acad.Sci. USA, 99, 4465–4470.
20. Lambrechts, A., Verschelde, J. L., Jonckheere, V.,Goethals, M., Vandekerckhove, J. & Ampe, C. (1997).The mammalian profilin isoforms display comple-mentary affinities for PIP2 and proline-rich sequences.EMBO J. 16, 484–494.
21. Witke, W., Podtelejnikov, A. V., Di Nardo, A., Suther-land, J. D., Gurniak, C. B., Dotti, C. &Mann, M. (1998).In mouse brain profilin I and profilin II associate withregulators of the endocytic pathway and actinassembly. EMBO J. 17, 967–976.
22. Bjorkegren, C., Rozycki, M., Schutt, C. E., Lindberg, U.& Karlsson, R. (1993). Mutagenesis of human profilinlocates its poly(L-proline)-binding site to a hydro-phobic patch of aromatic amino acids. FEBS Letters,333, 123–126.
23. Archer, S. J., Vinson, V. K., Pollard, T. D. & Torchia,D. A. (1994). Elucidation of the poly-L-proline bindingsite in Acanthamoeba profilin I by NMR spectroscopy.FEBS Letters, 337, 145–151.
24. Cedergren-Zeppezauer, E. S., Goonesekere, N. C.,Rozycki, M. D., Myslik, J. C., Dauter, Z., Lindberg, U.& Schutt, C. E. (1994). Crystallization and structuredetermination of bovine profilin at 2.0 Å resolution.J. Mol. Biol. 240, 459–475.
25. Metzler, W. J., Bell, A. J., Ernst, E., Lavoie, T. B. &Mueller, L. (1994). Identification of the poly-L-proline-binding site on human profilin. J. Biol. Chem. 269,4620–4625.
26. Nyman, T., Page, R., Schutt, C. E., Karlsson, R. &Lindberg, U. (2002). A cross-linked profilin-actinheterodimer interferes with elongation at the fast-growing end of F-actin. J. Biol. Chem. 277, 15828–15833.
27. Mahoney, N. M., Janmey, P. A. & Almo, S. C. (1997).Structure of the profilin-poly-L-proline complexinvolved in morphogenesis and cytoskeletal regula-tion. Nature Struct. Biol. 4, 953–960.
28. Lu, J. & Pollard, T. D. (2001). Profilin binding topoly-L-proline and actin monomers along withability to catalyze actin nucleotide exchange isrequired for viability of fission yeast. Mol. Biol. Cell,12, 1161–1175.
29. Haffner, C., Jarchau, T., Reinhard, M., Hoppe, J.,Lohmann, S. M. & Walter, U. (1995). Molecularcloning, structural analysis and functional expressionof the proline-rich focal adhesion and microfilament-associated protein VASP. EMBO J. 14, 19–27.
30. Zimmermann, J., Labudde, D., Jarchau, T., Walter, U.,Oschkinat, H. & Ball, L. J. (2002). Relaxation, equi-librium oligomerization, and molecular symmetry ofthe VASP (336-380) EVH2 tetramer. Biochemistry, 41,11143–11151.
31. Kuhnel, K., Jarchau, T., Wolf, E., Schlichting, I., Walter,U., Wittinghofer, A. & Strelkov, S. V. (2004). The VASPtetramerization domain is a right-handed coiled coilbased on a 15-residue repeat. Proc. Natl Acad. Sci. USA,101, 17027–17032.
32. Loureiro, J. J., Rubinson, D. A., Bear, J. E., Baltus,G. A., Kwiatkowski, A. V. & Gertler, F. B. (2002).Critical roles of phosphorylation and actin bindingmotifs, but not the central proline-rich region, forEna/vasodilator-stimulated phosphoprotein (VASP)function during cell migration. Mol. Biol. Cell, 13,2533–2546.
33. Han, Y. H., Chung, C. Y., Wessels, D., Stephens, S.,Titus, M. A., Soll, D. R. & Firtel, R. A. (2002).Requirement of a vasodilator-stimulated phospho-protein family member for cell adhesion, the forma-tion of filopodia, and chemotaxis in dictyostelium.J. Biol. Chem. 277, 49877–49887.
34. Auerbuch, V., Loureiro, J. J., Gertler, F. B., Theriot, J. A.& Portnoy, D. A. (2003). Ena/VASP proteins contri-bute to Listeria monocytogenes pathogenesis by con-trolling temporal and spatial persistence of bacterialactin-based motility. Mol. Microbiol. 49, 1361–1375.
35. Geese, M., Loureiro, J. J., Bear, J. E., Wehland, J.,Gertler, F. B. & Sechi, A. S. (2002). Contribution ofEna/VASP proteins to intracellular motility of listeriarequires phosphorylation and proline-rich core butnot F-actin binding or multimerization. Mol. Biol. Cell,13, 2383–2396.
36. Geese, M., Schluter, K., Rothkegel, M., Jockusch, B. M.,Wehland, J. & Sechi, A. S. (2000). Accumulation ofprofilin II at the surface of Listeria is concomitant withthe onset of motility and correlates with bacterialspeed. J. Cell Sci. 113, 1415–1426.
37. Grenklo, S., Geese, M., Lindberg, U., Wehland, J.,Karlsson, R. & Sechi, A. S. (2003). A crucial role forprofilin-actin in the intracellular motility of Listeriamonocytogenes. EMBO Rep. 4, 523–529.
38. Laurent, V., Loisel, T. P., Harbeck, B., Wehman, A.,Grobe, L., Jockusch, B. M. et al. (1999). Role ofproteins of the Ena/VASP family in actin-basedmotility of Listeria monocytogenes. J. Cell Biol. 144,1245–1258.
39. Li, F. & Higgs, H. N. (2003). The mouse Formin mDia1is a potent actin nucleation factor regulated byautoinhibition. Curr. Biol. 13, 1335–1340.
40. Romero, S., Le Clainche, C., Didry, D., Egile, C.,Pantaloni, D. & Carlier, M. F. (2004). Formin is aprocessive motor that requires profilin to accelerateactin assembly and associated ATP hydrolysis. Cell,119, 419–429.
41. Kovar, D. R., Harris, E. S., Mahaffy, R., Higgs, H. N. &Pollard, T. D. (2006). Control of the assembly ofATP- and ADP-actin by formins and profilin. Cell, 124,423–435.
42. Copeland, J. W., Copeland, S. J. & Treisman, R. (2004).Homo-oligomerization is essential for F-actin assem-bly by the formin family FH2 domain. J. Biol. Chem.279, 50250–50256.

289Profilin 2a Complexed with VASP and mDia1
43. Pruyne, D., Evangelista, M., Yang, C., Bi, E., Zigmond,S., Bretscher, A. & Boone, C. (2002). Role of formins inactin assembly: nucleation and barbed-end associa-tion. Science, 297, 612–615.
44. Otomo, T., Tomchick, D. R., Otomo, C., Panchal, S. C.,Machius, M. & Rosen, M. K. (2005). Structural basis ofactin filament nucleation and processive capping by aformin homology 2 domain. Nature, 433, 488–494.
45. Higashida, C., Miyoshi, T., Fujita, A., Oceguera-Yanez,F., Monypenny, J., Andou, Y., Narumiya, S. &Watanabe, N. (2004). Actin polymerization-drivenmolecular movement of mDia1 in living cells. Science,303, 2007–2010.
46. Watanabe, N. & Higashida, C. (2004). Formins: pro-cessive cappers of growing actin filaments. Expt. CellRes. 301, 16–22.
47. Lynch, E. D., Lee, M. K., Morrow, J. E., Welcsh, P. L.,Leon, P. E. & King, M. C. (1997). Nonsyndromicdeafness DFNA1 associated with mutation of ahuman homolog of the Drosophila gene diaphanous.Science, 278, 1315–1318.
48. Xu, Y., Moseley, J. B., Sagot, I., Poy, F., Pellman, D.,Goode, B. L. & Eck, M. J. (2004). Crystal structures of aFormin Homology-2 domain reveal a tethered dimerarchitecture. Cell, 116, 711–723.
49. Shimada, A., Nyitrai, M., Vetter, I. R., Kuhlmann, D.,Bugyi, B., Narumiya, S., Geeves, M. A. & Wittingho-fer, A. (2004). The core FH2 domain of diaphanous-related formins is an elongated actin binding proteinthat inhibits polymerization. Mol. Cell. 13, 511–522.
50. Lu, J., Meng, W., Poy, F., Maiti, S., Goode, B. L. & Eck,M. J. (2007). Structure of the FH2 domain of Daam1:implications for formin regulation of actin assembly.J. Mol. Biol. 369, 1258–1269.
51. Nezami, A. G., Poy, F. & Eck, M. J. (2006). Structure ofthe autoinhibitory switch in formin mDia1. Structure,14, 257–263.
52. Rose, R., Weyand, M., Lammers, M., Ishizaki, T.,Ahmadian, M. R. &Wittinghofer, A. (2005). Structuraland mechanistic insights into the interaction betweenRho and mammalian Dia. Nature, 435, 513–518.
53. Ball, L. J., Kuhne, R., Hoffmann, B., Hafner, A.,Schmieder, P., Volkmer-Engert, R. et al. (2000). Dualepitope recognition by the VASP EVH1 domainmodulates polyproline ligand specificity and bindingaffinity. EMBO J. 19, 4903–4914.
54. Chereau, D., Kerff, F., Graceffa, P., Grabarek, Z.,Langsetmo, K. & Dominguez, R. (2005). Actin-boundstructures of Wiskott-Aldrich syndrome protein(WASP)-homology domain 2 and the implicationsfor filament assembly. Proc. Natl Acad. Sci. USA, 102,16644–16649.
55. Mahoney, N. M., Rozwarski, D. A., Fedorov, E.,Fedorov, A. A. & Almo, S. C. (1999). Profilin bindsproline-rich ligands in two distinct amide backboneorientations. Nature Struct. Biol. 6, 666–671.
56. Brandl, M., Weiss, M. S., Jabs, A., Suhnel, J. &Hilgenfeld, R. (2001). C-H...pi-interactions in proteins.J. Mol. Biol. 307, 357–377.
57. Weiss, M. S., Brandl, M., Suhnel, J., Pal, D. &Hilgenfeld, R. (2001). More hydrogen bonds for the(structural) biologist. Trends Biochem. Sci. 26, 521–523.
58. Jonckheere, V., Lambrechts, A., Vandekerckhove, J. &Ampe, C. (1999). Dimerization of profilin II uponbinding the (GP5)3 peptide from VASP overcomes theinhibition of actin nucleation by profilin II andthymosin beta4. FEBS Letters, 447, 257–263.
59. Singh, S. S., Chauhan, A., Murakami, N., Styles, J.,Elzinga, M. & Chauhan, V. P. (1996). Phosphoinosi-
tide-dependent in vitro phosphorylation of profilin byprotein kinase C. Phospholipid specificity and locali-zation of the phosphorylation site. Recept. SignalTransduct. 6, 77–86.
60. Bjorkegren-Sjogren, C., Korenbaum, E., Nordberg, P.,Lindberg, U. & Karlsson, R. (1997). Isolation andcharacterization of two mutants of human profilin Ithat do not bind poly(L-proline). FEBS Letters, 418,258–264.
61. Pring, M., Evangelista, M., Boone, C., Yang, C. &Zigmond, S. H. (2003). Mechanism of formin-inducednucleation of actin filaments. Biochemistry, 42, 486–496.
62. Nodelman, I. M., Bowman, G. D., Lindberg, U. &Schutt, C. E. (1999). X-ray structure determination ofhuman profilin II: a comparative structural analysis ofhuman profilins. J. Mol. Biol. 294, 1271–1285.
63. Gieselmann, R., Kwiatkowski, D. J., Janmey, P. A. &Witke, W. (1995). Distinct biochemical characteristicsof the two human profilin isoforms. Eur. J. Biochem.229, 621–628.
64. Barzik, M., Kotova, T. I., Higgs, H. N., Hazelwood, L.,Hanein, D., Gertler, F. B. & Schafer, D. A. (2005). Ena/VASP proteins enhance actin polymerization in thepresence of barbed end capping proteins. J. Biol. Chem.280, 28653–28662.
65. Tobacman, L. S., Brenner, S. L. & Korn, E. D. (1983).Effect of Acanthamoeba profilin on the pre-steady statekinetics of actin polymerization and on the con-centration of F-actin at steady state. J. Biol. Chem.258, 8806–8812.
66. Tobacman, L. S. & Korn, E. D. (1982). The regulation ofactin polymerization and the inhibition of monomericactin ATPase activity by Acanthamoeba profilin. J. Biol.Chem. 257, 4166–4170.
67. Pilo Boyl, P., Di Nardo, A., Mulle, C., Sassoe-Pognetto,M., Panzanelli, P., Mele, A. et al. (2007). Profilin2contributes to synaptic vesicle exocytosis, neuronalexcitability, and novelty-seeking behavior. EMBO J.26, 2991–3002.
68. Stuven, T., Hartmann, E. &Gorlich, D. (2003). Exportin6: a novel nuclear export receptor that is specific forprofilin.actin complexes. EMBO J. 22, 5928–5940.
69. Birbach, A., Verkuyl, J. M. & Matus, A. (2006).Reversible, activity-dependent targeting of profilinto neuronal nuclei. Expt. Cell Res. 312, 2279–2287.
70. Eads, J. C., Mahoney, N. M., Vorobiev, S., Bresnick,A. R., Wen, K. K., Rubenstein, P. A., Haarer, B. K. &Almo, S. C. (1998). Structure determination andcharacterization of Saccharomyces cerevisiae profilin.Biochemistry, 37, 11171–11181.
71. Fedorov, A. A., Magnus, K. A., Graupe, M. H.,Lattman, E. E., Pollard, T. D. & Almo, S. C. (1994).X-ray structures of isoforms of the actin-binding pro-tein profilin that differ in their affinity for phospha-tidylinositol phosphates. Proc. Natl Acad. Sci. USA, 91,8636–8640.
72. Schutt, C. E., Myslik, J. C., Rozycki, M. D., Goonese-kere, N. C. & Lindberg, U. (1993). The structure ofcrystalline profilin-beta-actin. Nature, 365, 810–816.
73. Zarrinpar, A., Bhattacharyya, R. P. & Lim, W. A.(2003). The structure and function of proline recogni-tion domains. Sci STKE, RE8.
74. Bhattacharyya, R. & Chakrabarti, P. (2003). Stereo-specific interactions of proline residues in proteinstructures and complexes. J. Mol. Biol. 331, 925–940.
75. Petrella, E. C., Machesky, L. M., Kaiser, D. A. &Pollard, T. D. (1996). Structural requirements andthermodynamics of the interaction of proline peptideswith profilin. Biochemistry, 35, 16535–16543.

290 Profilin 2a Complexed with VASP and mDia1
76. Lambrechts, A., Jonckheere, V., Dewitte, D., Vande-kerckhove, J. & Ampe, C. (2002). Mutational analysisof human profilin I reveals a second PI(4,5)-P2 bindingsite neighbouring the poly(L-proline) binding site.BMC Biochem. 3, 12.
77. Chaudhary, A., Chen, J., Gu, Q. M., Witke, W.,Kwiatkowski, D. J. & Prestwich, G. D. (1998). Probingthe phosphoinositide 4,5-bisphosphate binding site ofhuman profilin I. Chem. Biol. 5, 273–281.
78. Chereau, D. & Dominguez, R. (2006). Understandingthe role of the G-actin-binding domain of Ena/VASPin actin assembly. J. Struct. Biol. 155, 195–201.
79. Goode, B. L. & Eck, M. J. (2007). Mechanism andfunction of formins in control of actin assembly. Annu.Rev. Biochem. 76, 593–627.
80. Way, M., Pope, B., Gooch, J., Hawkins, M. & Weeds,A. G. (1990). Identification of a region in segment 1of gelsolin critical for actin binding. EMBO J. 9,4103–4109.
81. Kaiser, D. A., Goldschmidt-Clermont, P. J., Levine,B. A. & Pollard, T. D. (1989). Characterization ofrenatured profilin purified by urea elution from poly-L-proline agarose columns. Cell Motil. Cytoskel. 14,251–262.
82. Janmey, P. A. (1991). Polyproline affinity method forpurification of platelet profilin and modification withpyrene-maleimide. Methods Enzymol. 196, 92–99.
83. Kabsch, W. (1993). Automatic processing of rotationdiffraction data from crystals of initially unknownsymmetry and cell constants. J. Appl. Crystallog. 26,795–800.
84. Kursula, P. (2004). XDSi - a graphical interface forthe data processing program XDS. J. Appl. Crystallog.37, 347–348.
85. Navaza, J. (2001). Implementation of molecularreplacement in AMoRe. Acta Crystallog. sect. D, 57,1367–1372.
86. Murshudov, G. N., Vagin, A. A. & Dodson, E. J. (1997).Refinement of macromolecular structures by the
maximum-likelihood method. Acta Crystallog. sect. D,53, 240–255.
87. Winn, M. D., Isupov, M. N. & Murshudov, G. N.(2001). Use of TLS parameters to model anisotropicdisplacements in macromolecular refinement. ActaCrystallog. sect. D, 57, 122–133.
88. Jones, T. A., Zou, J. Y., Cowan, S. W. & Kjeldgaard, M.(1991). Improved methods for building proteinmodels in electron density maps and the location oferrors in these models. Acta Crystallog. sect. A, 47,110–119.
89. Emsley, P. & Cowtan, K. (2004). Coot: model-buildingtools for molecular graphics. Acta Crystallog. sect. D,60, 2126–2132.
90. Perrakis, A., Morris, R. & Lamzin, V. S. (1999).Automated protein model building combined withiterative structure refinement. Nature Struct. Biol. 6,458–463.
91. Murshudov, G. N., Vagin, A. A., Lebedev, A., Wilson,K. S. & Dodson, E. J. (1999). Efficient anisotropicrefinement of macromolecular structures using FFT.Acta Crystallog. sect. D, 55, 247–255.
92. Afonine, P. V., Grosse-Kunstleve, R. W. & Adams, P. D.(2005). A robust bulk-solvent correction and aniso-tropic scaling procedure. Acta Crystallog. sect. D, 61,850–855.
93. Merritt, E. A. (1999). Expanding the model: aniso-tropic displacement parameters in protein structurerefinement. Acta Crystallog. sect. D, 55, 1109–1117.
94. Spudich, J. A. & Watt, S. (1971). The regulation ofrabbit skeletal muscle contraction. I. Biochemicalstudies of the interaction of the tropomyosin-troponincomplex with actin and the proteolytic fragments ofmyosin. J. Biol. Chem. 246, 4866–4871.
95. Kouyama, T. & Mihashi, K. (1981). Fluorimetry studyof N-(1-pyrenyl)iodoacetamide-labelled F-actin. Localstructural change of actin protomer both on polymer-ization and on binding of heavy meromyosin. Eur. J.Biochem. 114, 33–38.




















