
Areas of cat auditory cortex as defined by neurofilament proteins expressing SMI32
Upload
independentCategory
view
0download
0
The Neurofilament Infrastructure of aDeveloping Presynaptic Calyx
JACQUES PAYSAN, WILLIAM G. CONROY, JAY S. COGGAN,
AND DARWIN K. BERG*
Department of Biology, University of California, San Diego, La Jolla,California 92093-0357
ABSTRACTCalyx-type synapses appear to be specifically designed to support fast, reliable,
high-frequency excitatory transmission. In the chick ciliary ganglion, calyx terminalsfrom preganglionic neurons in the midbrain form early in development on ciliary neurons.We find that labeling the calyx membranes with a lipophilic dye delivered by diffusiondown the preganglionic nerve reveals a large membrane structure engulfing the postsyn-aptic cell by the end of embryogenesis. In contrast, labeling the calyces with a water-soluble dye by diffusion through the preganglionic nerve suggests large discontinuities inthe calyx. A similar pattern of discontinuities is seen when presynaptic neurofilamentsare labeled with antibodies selective for highly phosphorylated neurofilaments. Theneurofilament infrastructure of the calyx first appears as a single thick bundle, whichsubsequently bifurcates during development and eventually generates a fine meshwork offilaments subdivided by several large neurofilament bundles encircling the postsynapticcell body. The large bundles probably produce protruding ridges in the otherwise thincalyx cup, accounting for the disparity in staining patterns observed with membrane andcytosolic dyes. The postsynaptic membrane also undergoes restructuring during develop-ment with the appearance of large folded mats of somatic spines heavily invested withnicotinic receptors. The large presynaptic neurofilament bundles do not overlap thepostsynaptic receptor clusters but do codistribute with large tracks of presynaptic micro-tubules. The neurofilament bundles may act as girders to provide structural supportwhile at the same time defining conduits for microtubule-dependent transport of mate-rials and rapid propagation of electrical signals throughout the extended calyx. J. Comp.Neurol. 425:284 –294, 2000. © 2000 Wiley-Liss, Inc.
Indexing terms: nicotinic; receptor; ciliary; ganglion; acetylcholine; synapse; terminal
The vertebrate neuromuscular junction has long pro-vided a useful model for analyzing the development ofsynaptic structure and function (Sanes and Lichtman,1999). In two respects, however, the neuromuscular junc-tion is atypical of synapses in the nervous system. First, itis designed to deliver high-frequency, reliable suprath-reshold stimulation whereas most interneuronal synapsesgenerate unreliable (high failure rate) subthresholdpostsynaptic signals (Smetters and Zador, 1996). Second,the neuromuscular junction is a large structure, madepossible in part by the postsynaptic muscle fiber, whichprovides machinery for synaptic stabilization and signaltransduction. Many interneuronal synapses, in contrast,involve single presynaptic boutons juxtaposed to smallpostsynaptic densities either on dendritic spines or ondiscrete patches of dendritic or somal membrane (Peterset al., 1976). The postsynaptic spines, moreover, are dy-
namic in an activity-dependent manner (Matus, 1999;Toni et al., 1999).
An interesting class of hybrid vertebrate synapses isprovided by those having a large presynaptic calyx engulf-ing the postsynaptic neuronal soma. Examples includecalyx synapses in the mammalian trapezoid body (Held,
Grant sponsor: NIH; Grant numbers: NS12601 and NS35469; Grantsponsor: the Tobacco-Related Diseases Research Program; Grant number:6RT-0050; Grant sponsor: the Swiss National Science Foundation; Grantsponsor: the Deutsche Forschungs Gemeinschaft.
Jacques Paysan’s current address is: the Institute of Physiology (230),University of Hohenheim, 70593 Stuttgart, Germany.
*Correspondence to: Darwin K. Berg, Department of Biology, Universityof California, San Diego, La Jolla, California 92093-0357.E-mail: [email protected]
Received 7 December 1999; Revised 1 May 2000; Accepted 2 June 2000
THE JOURNAL OF COMPARATIVE NEUROLOGY 425:284–294 (2000)
© 2000 WILEY-LISS, INC.

1893), the vestibular organ (Ramon y Cajal, 1908), and theavian ciliary ganglion (von Lenhossek, 1911). Like theneuromuscular junction, calyx synapses appear to becharged with the responsibility of generating reliablehigh-frequency suprathreshold excitation (Dryer, 1994),but the design of the synapse is constrained by the geom-etry of the postsynaptic neuron soma. An extended pre-synaptic calyx terminal overlaying multiple postsynapticsites of signal transduction appears to be a common strat-egy for meeting this challenge.
The development of functional transmission at a calyxsynapse has been described in some detail in the chickciliary ganglion. Two classes of neurons are found in theganglion in about equal numbers: choroid neurons thatinnervate smooth muscle in the choroid layer and ciliaryneurons that innervate striated muscle in the iris andciliary body (Landmesser and Pilar, 1974a). Both classesof neurons are innervated by preganglionic axons arisingfrom the accessory oculomotor nucleus of the midbrain,but only the ciliary neurons receive calyces. The initialcontacts involve bouton-like termini and achieve func-tional nicotinic transmission by embryonic day 7 (E7;Landmesser and Pilar, 1972). Over the next few days ofembryonic life the boutons are replaced by a calyx struc-ture covering much of the ciliary neuron soma, and theneurons acquire two kinds of nicotinic acetylcholine recep-tors (nAChRs) in increasing numbers. One species (a3*-nAChRs) contains multiple kinds of subunits (Vernallis etal., 1993; Conroy and Berg, 1995) and is found at postsyn-aptic densities as well as perisynaptic sites on the neurons(Jacob et al., 1984; Loring and Zigmond, 1987; WilsonHorch and Sargent, 1995). The other (a7-nAChRs) con-tains a7 subunits (Vernallis et al., 1993), is much moreabundant (Chiappinelli and Giacobini, 1978; Corriveauand Berg, 1994) and excluded from postsynaptic densities(Jacob and Berg, 1983; Wilson Horch and Sargent, 1995),but is highly concentrated on somatic spines tightly foldedinto discrete clusters on the neuron (Shoop et al., 1999).Near the end of embryogenesis the synapse develops anelectrical component that features prominently in trans-mission (Martin and Pilar, 1963; Landmesser and Pilar,1972). At maturation, the synapses are capable of sustain-ing transmission at rates in excess of 100 Hz (Dryer,1994).
Little is known about the molecular infrastructure thatforms and stabilizes the presynaptic calyx terminal. Wehave used confocal microscopy and immunofluorescence toexamine the distribution of neurofilament (NF) bundles inthe calyx. Comparisons with a7-nAChR distributions onthe postsynaptic cell, as well as costaining with space-filling and lipid-soluble dyes, have provided additionalinformation about the structural development of calyxsynapses.
MATERIALS AND METHODS
Neurobiotin and DiI staining
DiI, biocytin, and neurobiotin staining was performedas previously described (Coggan et al., 1997). Briefly, ca-lyces in ciliary ganglia of various developmental stageswere filled with 1,19-dihexadecyl-3,3,39,39-tetramethyl-indocarbo-cyanine (DiI; Molecular Probes, Eugene, OR) byplacing small crystals of the dye on the preganglionicnerve root of ganglia fixed in 4% phosphate-buffered form-
aldehyde solution, pH 7.4, and waiting 5 days for diffusionto occur along the nerve. Alternatively, either small bio-cytin (Sigma, St. Louis, MO) or neurobiotin (Vector, Bur-lingame, CA) crystals were placed on the preganglionicnerve root of fresh ganglia and maintained for 1 hour atroom temperature in extracellular recording solution (120mM NaCl, 4 mM KCl, 10 mM glucose, 2 mM CaCl2, 1 mMMgSO4, 1 mM NaH2PO4, and 25 mM NaHCO3, pH 7.4)gassed with 95% O2/5% CO2. Biocytin and neurobiotinwere visualized by fixing the labeled ganglia in freshlyprepared formaldehyde solution (2% (w/v) formaldehydesolution, 150 mM sodium phosphate, pH 7.4) for 30 min-utes on ice, rinsing with ice-cold phosphate-buffered sa-line (PBS; 150 mM sodium chloride plus 10 mM sodiumphosphate, pH 7.4), and then incubating for 30 minutes atroom temperature in PBS containing 10% (v/v) donkeyserum (Sigma), 0.3% (v/v) Triton X-100, and a 1:500 dilu-tion of Cy3-conjugated streptavidin (Molecular Probes).The staining patterns were examined with a Noran Odys-sey confocal laser scanning microscope.
Immunostaining and a7-nAChR labeling
Immunofluorescent localization of nAChRs was con-ducted as previously described (Kassner et al., 1998).Freshly dissected embryonic chick ciliary ganglia weredissected and fixed with 2% paraformaldehyde in 150 mMsodium phosphate, pH 7.4, for 20 minutes at room tem-perature. After rinsing in PBS, the ganglia were incubatedfor 2 hours at room temperature or overnight at 4°C withprimary antibodies in PBS containing 0.2% Triton X-100and 10% donkey serum, rinsed again with PBS, incubatedwith secondary antibodies for 1 hour at room temperature,and rinsed with PBS. Coverslips were mounted on glassslides with Vectashield (Vector), and images were usuallyanalyzed by the Noran Odyssey (Middleton, WI) confocalmicroscope with a 603, 1.3 NA oil immersion objectivelens. In some cases images were obtained with a Bio-Rad(Hercules, CA) confocal laser scanning microscope or aZeiss Axiovert equipped with Intelligence Imaging Inno-vations (Denver, Co) deconvoluting software, in both caseswith a 633, 1.4 NA oil immersion objective lens. Individ-ual optical sections were taken at 0.75–1-mm steps, andthe final volume was assembled digitally by using NIHimage software for the Noran, Lasersharp software (Bio-Rad) for the Bio-Rad, and the deconvoluting software forthe Zeiss. In all cases the final images were configuredwith Adobe Photoshop software (Mountain View, CA).
The primary antibodies used to detect a7-nAChRs weremAb 306 and goat anti-a7 antiserum (Santa Cruz Biotech-nology, Santa Cruz, CA). For neurofilament heavy chain(NF-H), the primary antibodies were mAbs NE14 and N52and antiserum N4142. NE14 was raised against highlyphosphorylated NF-H from pig spinal cord and is reportedto show no cross-reactivity against dephosphorylatedNF-H (Sigma, #N5389). N52 was raised against enzymat-ically dephosphorylated pig NF-H and purportedly recog-nizes both phosphorylated and nonphosphorylated NF-H(Sigma, #N0142). The polyclonal antiserum N4142 raisedagainst bovine HF-H is reported to recognize phosphory-lated NFs preferentially (Sigma, #N4142). For microtu-bules, the antibodies were either mAb DM1A directedagainst a-tubulin (Sigma) or mAb TUJ1 directed againstb3-tubulin (Covance, Princeton, NJ). Secondary antibod-ies included Oregon Green- (Molecular Probes) and Cy3-
285NEUROFILAMENT BUNDLES IN A PRESYNAPTIC CALYX
conjugated (Jackson ImmunoResearch, West Grove, PA)donkey and goat antisera.
Materials
White leghorn chick embryos were obtained locally andmaintained at 37°C in a humidified incubator. mAb 306was kindly provided by Dr. Jon Lindstrom (University ofPennsylvania, Philadelphia). aBgt was purchased fromBiotoxins (St. Cloud, Florida). All other reagents werepurchased from Sigma unless otherwise indicated.
RESULTS
Calyx structure
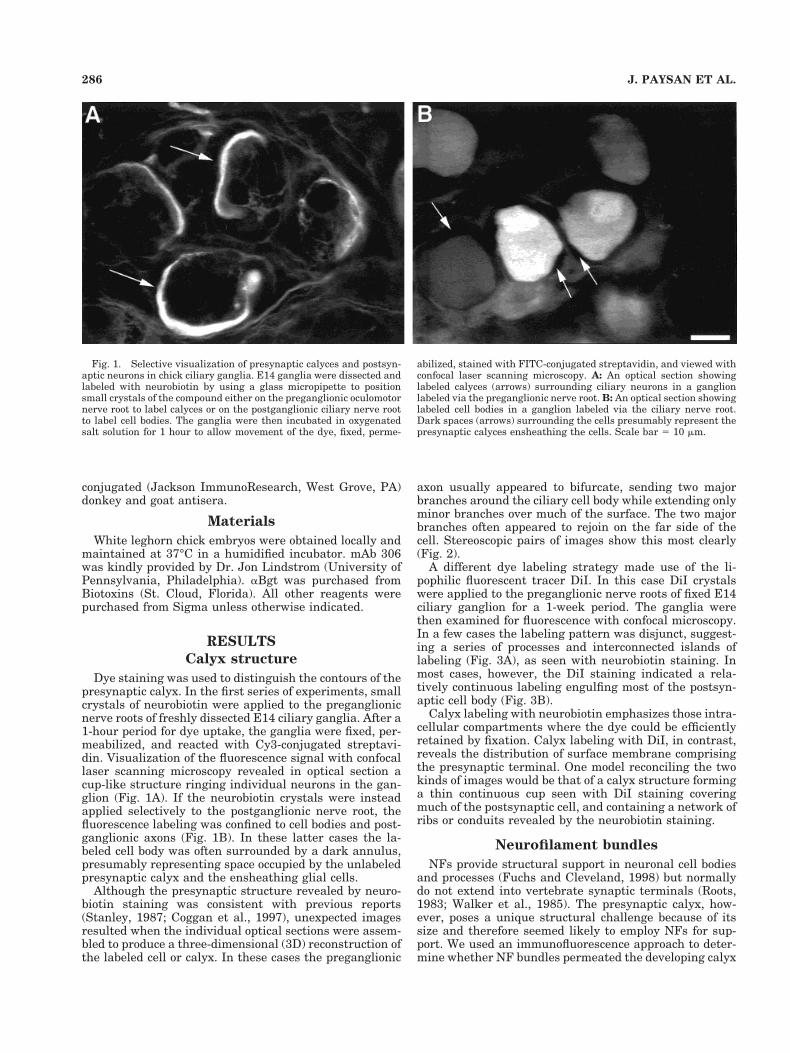
Dye staining was used to distinguish the contours of thepresynaptic calyx. In the first series of experiments, smallcrystals of neurobiotin were applied to the preganglionicnerve roots of freshly dissected E14 ciliary ganglia. After a1-hour period for dye uptake, the ganglia were fixed, per-meabilized, and reacted with Cy3-conjugated streptavi-din. Visualization of the fluorescence signal with confocallaser scanning microscopy revealed in optical section acup-like structure ringing individual neurons in the gan-glion (Fig. 1A). If the neurobiotin crystals were insteadapplied selectively to the postganglionic nerve root, thefluorescence labeling was confined to cell bodies and post-ganglionic axons (Fig. 1B). In these latter cases the la-beled cell body was often surrounded by a dark annulus,presumably representing space occupied by the unlabeledpresynaptic calyx and the ensheathing glial cells.
Although the presynaptic structure revealed by neuro-biotin staining was consistent with previous reports(Stanley, 1987; Coggan et al., 1997), unexpected imagesresulted when the individual optical sections were assem-bled to produce a three-dimensional (3D) reconstruction ofthe labeled cell or calyx. In these cases the preganglionic
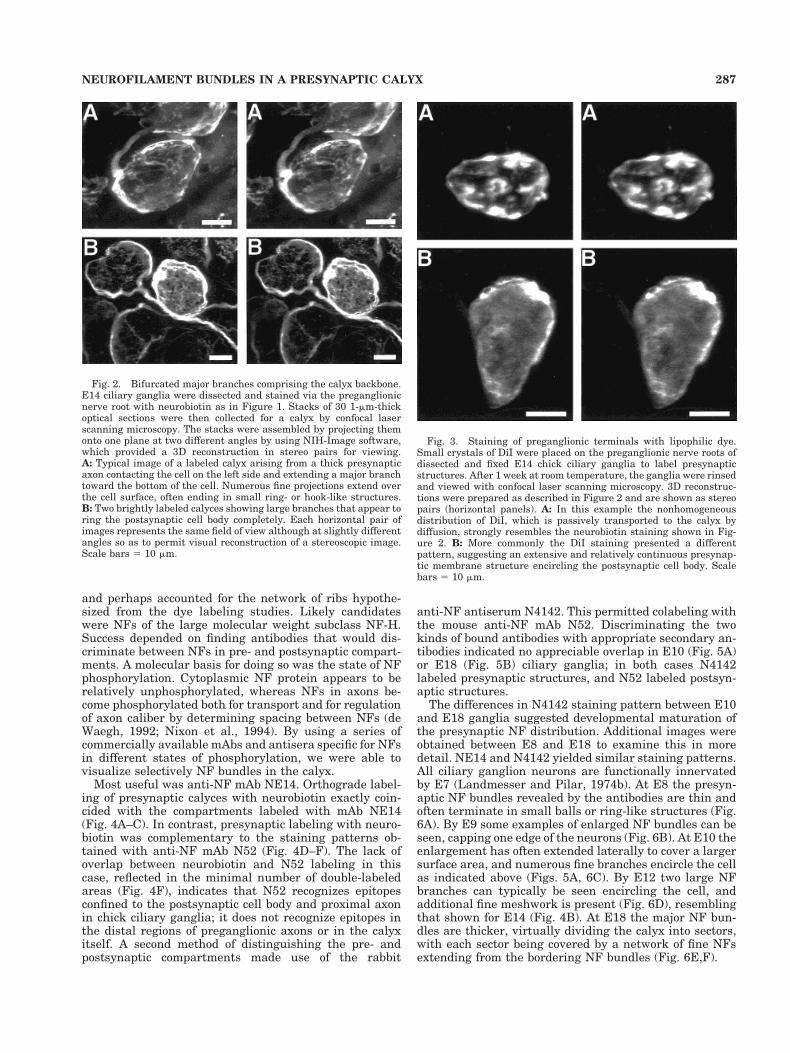
axon usually appeared to bifurcate, sending two majorbranches around the ciliary cell body while extending onlyminor branches over much of the surface. The two majorbranches often appeared to rejoin on the far side of thecell. Stereoscopic pairs of images show this most clearly(Fig. 2).
A different dye labeling strategy made use of the li-pophilic fluorescent tracer DiI. In this case DiI crystalswere applied to the preganglionic nerve roots of fixed E14ciliary ganglion for a 1-week period. The ganglia werethen examined for fluorescence with confocal microscopy.In a few cases the labeling pattern was disjunct, suggest-ing a series of processes and interconnected islands oflabeling (Fig. 3A), as seen with neurobiotin staining. Inmost cases, however, the DiI staining indicated a rela-tively continuous labeling engulfing most of the postsyn-aptic cell body (Fig. 3B).
Calyx labeling with neurobiotin emphasizes those intra-cellular compartments where the dye could be efficientlyretained by fixation. Calyx labeling with DiI, in contrast,reveals the distribution of surface membrane comprisingthe presynaptic terminal. One model reconciling the twokinds of images would be that of a calyx structure forminga thin continuous cup seen with DiI staining coveringmuch of the postsynaptic cell, and containing a network ofribs or conduits revealed by the neurobiotin staining.
Neurofilament bundles
NFs provide structural support in neuronal cell bodiesand processes (Fuchs and Cleveland, 1998) but normallydo not extend into vertebrate synaptic terminals (Roots,1983; Walker et al., 1985). The presynaptic calyx, how-ever, poses a unique structural challenge because of itssize and therefore seemed likely to employ NFs for sup-port. We used an immunofluorescence approach to deter-mine whether NF bundles permeated the developing calyx
Fig. 1. Selective visualization of presynaptic calyces and postsyn-aptic neurons in chick ciliary ganglia. E14 ganglia were dissected andlabeled with neurobiotin by using a glass micropipette to positionsmall crystals of the compound either on the preganglionic oculomotornerve root to label calyces or on the postganglionic ciliary nerve rootto label cell bodies. The ganglia were then incubated in oxygenatedsalt solution for 1 hour to allow movement of the dye, fixed, perme-
abilized, stained with FITC-conjugated streptavidin, and viewed withconfocal laser scanning microscopy. A: An optical section showinglabeled calyces (arrows) surrounding ciliary neurons in a ganglionlabeled via the preganglionic nerve root. B: An optical section showinglabeled cell bodies in a ganglion labeled via the ciliary nerve root.Dark spaces (arrows) surrounding the cells presumably represent thepresynaptic calyces ensheathing the cells. Scale bar 5 10 mm.
286 J. PAYSAN ET AL.
and perhaps accounted for the network of ribs hypothe-sized from the dye labeling studies. Likely candidateswere NFs of the large molecular weight subclass NF-H.Success depended on finding antibodies that would dis-criminate between NFs in pre- and postsynaptic compart-ments. A molecular basis for doing so was the state of NFphosphorylation. Cytoplasmic NF protein appears to berelatively unphosphorylated, whereas NFs in axons be-come phosphorylated both for transport and for regulationof axon caliber by determining spacing between NFs (deWaegh, 1992; Nixon et al., 1994). By using a series ofcommercially available mAbs and antisera specific for NFsin different states of phosphorylation, we were able tovisualize selectively NF bundles in the calyx.
Most useful was anti-NF mAb NE14. Orthograde label-ing of presynaptic calyces with neurobiotin exactly coin-cided with the compartments labeled with mAb NE14(Fig. 4A–C). In contrast, presynaptic labeling with neuro-biotin was complementary to the staining patterns ob-tained with anti-NF mAb N52 (Fig. 4D–F). The lack ofoverlap between neurobiotin and N52 labeling in thiscase, reflected in the minimal number of double-labeledareas (Fig. 4F), indicates that N52 recognizes epitopesconfined to the postsynaptic cell body and proximal axonin chick ciliary ganglia; it does not recognize epitopes inthe distal regions of preganglionic axons or in the calyxitself. A second method of distinguishing the pre- andpostsynaptic compartments made use of the rabbit
anti-NF antiserum N4142. This permitted colabeling withthe mouse anti-NF mAb N52. Discriminating the twokinds of bound antibodies with appropriate secondary an-tibodies indicated no appreciable overlap in E10 (Fig. 5A)or E18 (Fig. 5B) ciliary ganglia; in both cases N4142labeled presynaptic structures, and N52 labeled postsyn-aptic structures.
The differences in N4142 staining pattern between E10and E18 ganglia suggested developmental maturation ofthe presynaptic NF distribution. Additional images wereobtained between E8 and E18 to examine this in moredetail. NE14 and N4142 yielded similar staining patterns.All ciliary ganglion neurons are functionally innervatedby E7 (Landmesser and Pilar, 1974b). At E8 the presyn-aptic NF bundles revealed by the antibodies are thin andoften terminate in small balls or ring-like structures (Fig.6A). By E9 some examples of enlarged NF bundles can beseen, capping one edge of the neurons (Fig. 6B). At E10 theenlargement has often extended laterally to cover a largersurface area, and numerous fine branches encircle the cellas indicated above (Figs. 5A, 6C). By E12 two large NFbranches can typically be seen encircling the cell, andadditional fine meshwork is present (Fig. 6D), resemblingthat shown for E14 (Fig. 4B). At E18 the major NF bun-dles are thicker, virtually dividing the calyx into sectors,with each sector being covered by a network of fine NFsextending from the bordering NF bundles (Fig. 6E,F).
Fig. 2. Bifurcated major branches comprising the calyx backbone.E14 ciliary ganglia were dissected and stained via the preganglionicnerve root with neurobiotin as in Figure 1. Stacks of 30 1-mm-thickoptical sections were then collected for a calyx by confocal laserscanning microscopy. The stacks were assembled by projecting themonto one plane at two different angles by using NIH-Image software,which provided a 3D reconstruction in stereo pairs for viewing.A: Typical image of a labeled calyx arising from a thick presynapticaxon contacting the cell on the left side and extending a major branchtoward the bottom of the cell. Numerous fine projections extend overthe cell surface, often ending in small ring- or hook-like structures.B: Two brightly labeled calyces showing large branches that appear toring the postsynaptic cell body completely. Each horizontal pair ofimages represents the same field of view although at slightly differentangles so as to permit visual reconstruction of a stereoscopic image.Scale bars 5 10 mm.
Fig. 3. Staining of preganglionic terminals with lipophilic dye.Small crystals of DiI were placed on the preganglionic nerve roots ofdissected and fixed E14 chick ciliary ganglia to label presynapticstructures. After 1 week at room temperature, the ganglia were rinsedand viewed with confocal laser scanning microscopy. 3D reconstruc-tions were prepared as described in Figure 2 and are shown as stereopairs (horizontal panels). A: In this example the nonhomogeneousdistribution of DiI, which is passively transported to the calyx bydiffusion, strongly resembles the neurobiotin staining shown in Fig-ure 2. B: More commonly the DiI staining presented a differentpattern, suggesting an extensive and relatively continuous presynap-tic membrane structure encircling the postsynaptic cell body. Scalebars 5 10 mm.
287NEUROFILAMENT BUNDLES IN A PRESYNAPTIC CALYX
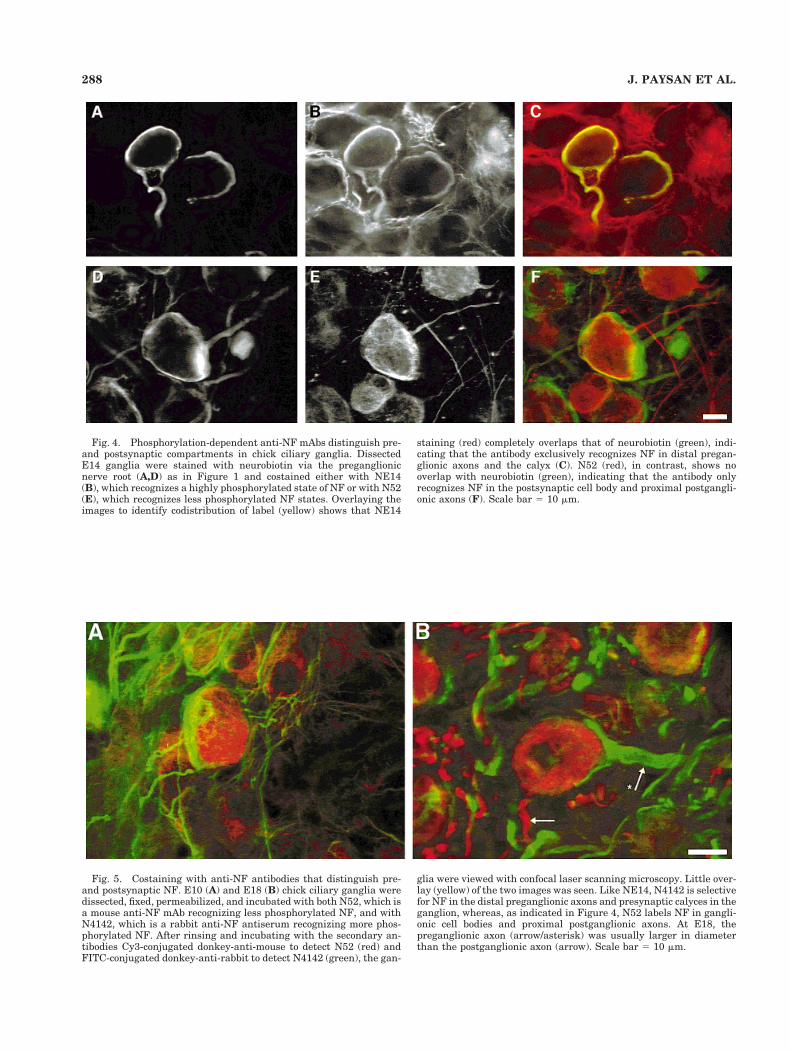
Fig. 4. Phosphorylation-dependent anti-NF mAbs distinguish pre-and postsynaptic compartments in chick ciliary ganglia. DissectedE14 ganglia were stained with neurobiotin via the preganglionicnerve root (A,D) as in Figure 1 and costained either with NE14(B), which recognizes a highly phosphorylated state of NF or with N52(E), which recognizes less phosphorylated NF states. Overlaying theimages to identify codistribution of label (yellow) shows that NE14
staining (red) completely overlaps that of neurobiotin (green), indi-cating that the antibody exclusively recognizes NF in distal pregan-glionic axons and the calyx (C). N52 (red), in contrast, shows nooverlap with neurobiotin (green), indicating that the antibody onlyrecognizes NF in the postsynaptic cell body and proximal postgangli-onic axons (F). Scale bar 5 10 mm.
Fig. 5. Costaining with anti-NF antibodies that distinguish pre-and postsynaptic NF. E10 (A) and E18 (B) chick ciliary ganglia weredissected, fixed, permeabilized, and incubated with both N52, which isa mouse anti-NF mAb recognizing less phosphorylated NF, and withN4142, which is a rabbit anti-NF antiserum recognizing more phos-phorylated NF. After rinsing and incubating with the secondary an-tibodies Cy3-conjugated donkey-anti-mouse to detect N52 (red) andFITC-conjugated donkey-anti-rabbit to detect N4142 (green), the gan-
glia were viewed with confocal laser scanning microscopy. Little over-lay (yellow) of the two images was seen. Like NE14, N4142 is selectivefor NF in the distal preganglionic axons and presynaptic calyces in theganglion, whereas, as indicated in Figure 4, N52 labels NF in gangli-onic cell bodies and proximal postganglionic axons. At E18, thepreganglionic axon (arrow/asterisk) was usually larger in diameterthan the postganglionic axon (arrow). Scale bar 5 10 mm.
288 J. PAYSAN ET AL.
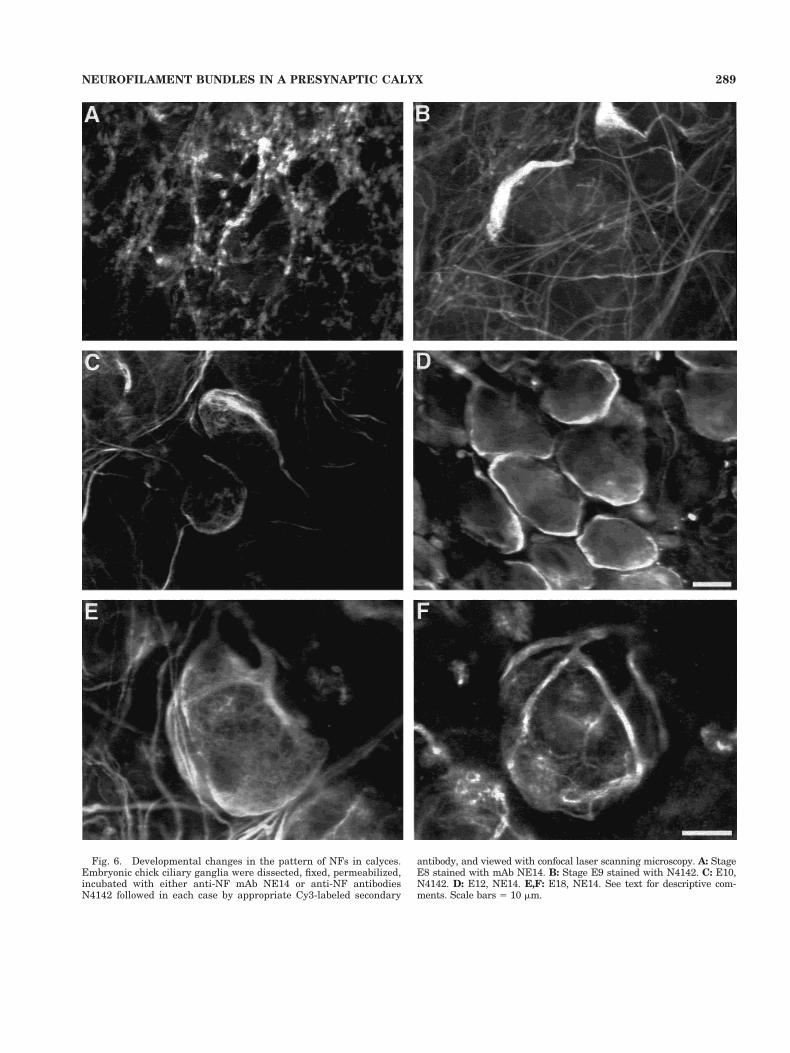
Fig. 6. Developmental changes in the pattern of NFs in calyces.Embryonic chick ciliary ganglia were dissected, fixed, permeabilized,incubated with either anti-NF mAb NE14 or anti-NF antibodiesN4142 followed in each case by appropriate Cy3-labeled secondary
antibody, and viewed with confocal laser scanning microscopy. A: StageE8 stained with mAb NE14. B: Stage E9 stained with N4142. C: E10,N4142. D: E12, NE14. E,F: E18, NE14. See text for descriptive com-ments. Scale bars 5 10 mm.
289NEUROFILAMENT BUNDLES IN A PRESYNAPTIC CALYX

Distribution of a7-nAChRs on ciliaryneurons
The developmental changes in NF pattern seen presyn-aptically raised the question of whether postsynapticstructures change in parallel. A prominent structural fea-ture of the postsynaptic cell near the end of embryogenesisis the existence of discrete mats of tightly folded somaticspines containing large numbers of a7-nAChRs (Shoop etal., 1999). The macroclusters of a7-nAChRs become appar-ent after E8 (Blumenthal et al., 1999). To determine howthis postsynaptic pattern comes about and whether itparallels presynaptic changes in NF bundles, ganglia werestained with the anti-a7-nAChR mAb 306 followed byCy3-conjugated secondary antibody. The mAb was raisedagainst the major cytoplasmic loop of a7-nAChR and,therefore, recognizes an intracellular epitope (Schoepferet al., 1990).
At E8 only a faint level of staining was detectable, andfor the most part was spread diffusely over the cell bodies;receptor clusters were sometimes distinguishable butwere rudimentary (Fig. 7A). The signal represented spe-cific binding as demonstrated by the negative controls inwhich the primary mAb was replaced with an equivalentamount of normal mouse IgG (Fig. 7F). By E10 a numberof large a7-nAChR clusters appeared on the cells (Fig. 7B).In addition, some of the label was distributed in a filamen-tous fashion over the cells at both E8 and E10. From E12through E18 the number of a7-nAChR clusters per cellincreased in number and in brightness, and filamentousstaining was relatively insignificant (Fig. 7C–E). Similarresults were obtained when a7-nAChRs were stained ei-ther with a goat anti-a7-nAChR antiserum or with fluo-rescently labeled aBgt (data not shown).
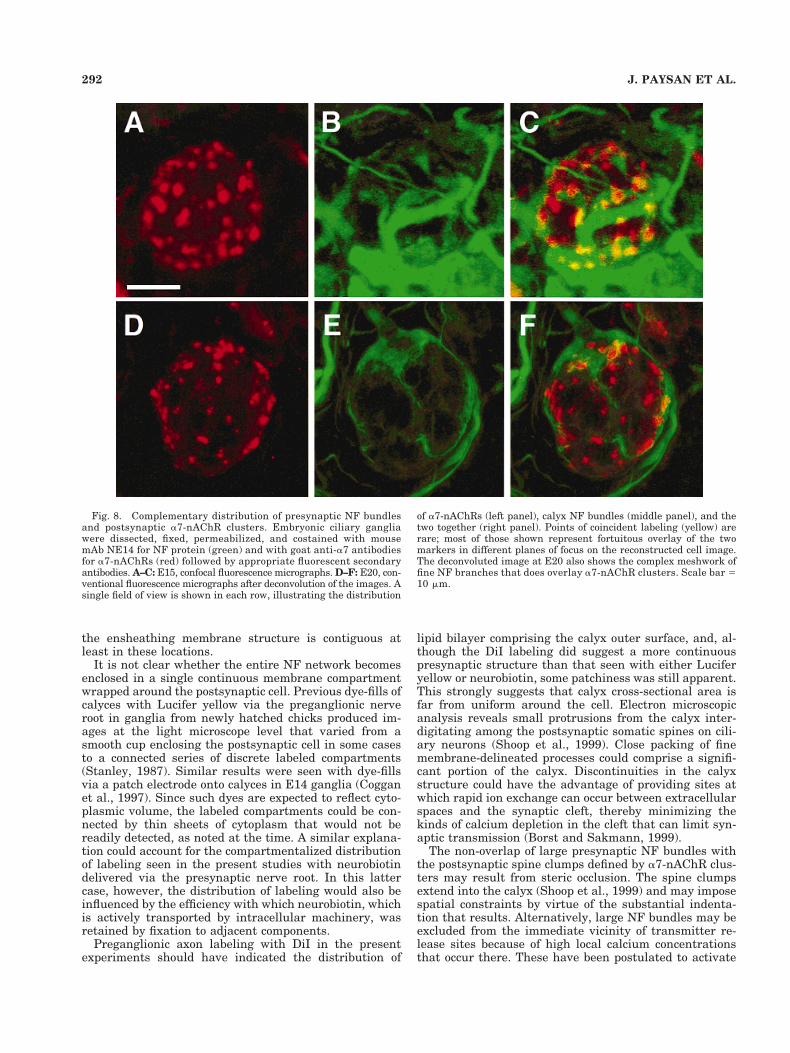
Costaining with markers for presynaptic NFs andpostsynaptic a7-nAChRs was used to examine the spatialrelationship between the receptor clusters and the largeNF bundles. Numerous large a7-nAChR clusters could bedistinguished on the cell bodies of neurons in E15 gangliaby using anti-a7 antibodies (Fig. 8A–C). Examining thesame cells stained with NE14 revealed the large presyn-aptic NF bundles with smaller branches extendingthroughout the calyx. Combining the two images showed aconsistent pattern suggesting complementary but largelyexclusive staining distributions. A similar pattern wasfound at E20, immediately prior to hatching (Fig. 8D–F).Thus the large NF bundles did not appear to terminate onor systematically overlay the receptor clusters but insteadskirted them, occupying adjacent space in the 2D projec-tion. Often the bundles appeared in close proximity to theclusters, defining cluster rows on the postsynaptic cell.The results indicate a nonrandom spatial relationship be-tween the large presynaptic NF bundles and the promi-nent postsynaptic mats of somatic spines containing a7-nAChRs. A complex meshwork of fine NF branches couldalso be distinguished, particularly at late embryonictimes, and was found to traverse the regions between thelarge NF bundles; the meshwork was not excluded fromcalyx regions overlying the postsynaptic spine clumps.
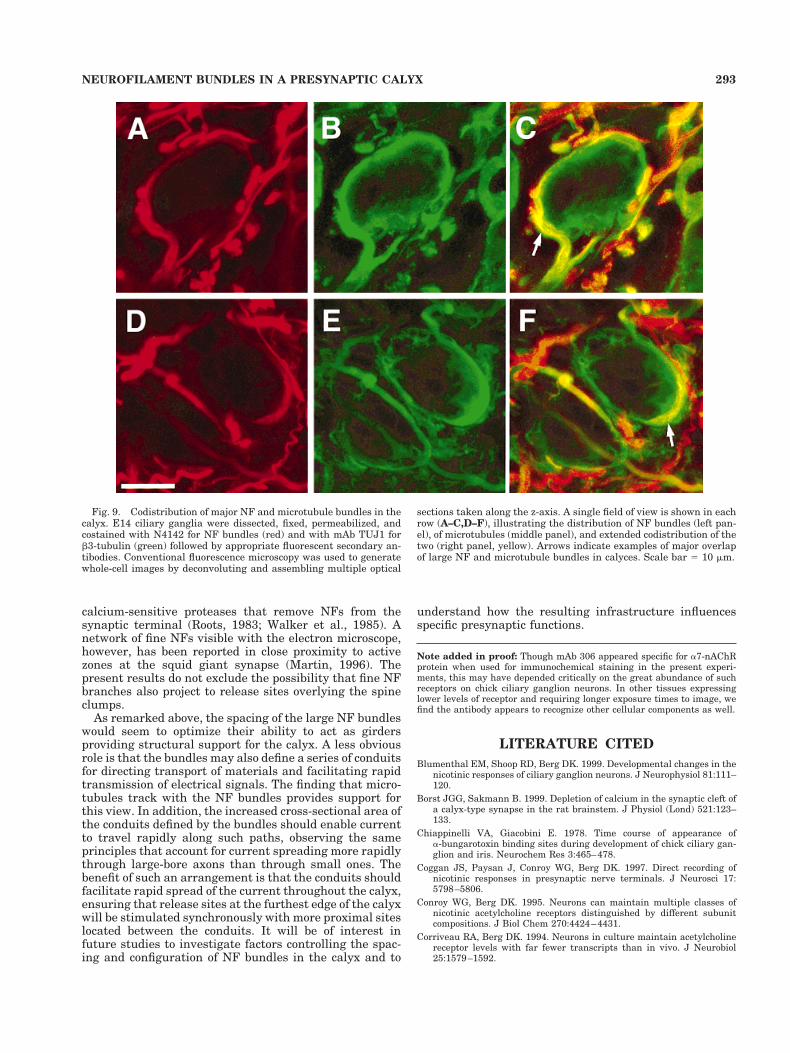
Antitubulin antibodies were used to determine the re-lationship between NF bundles and microtubules in thepresynaptic calyx. Costaining indicated prominent micro-tubule bundles tracking with the large NF bundles in thecalyx (Fig. 9). In addition, a fine array of microtubules wasseen spreading out between the NF bundles, and some
microtubule staining was present in the postsynaptic cellas well (appearing as diffuse staining in this case). Theresults support the hypothesis that the large NF bundlesnot only provide structural support for the calyx, but alsoprovide routes for microtubule-dependent traffic of mate-rials to distal calyx surfaces.
DISCUSSION
The major findings reported here are that the presyn-aptic calyx appears to be largely a continuous thin mem-branous structure covering most of the ciliary neuron bylate embryogenesis, and that it is penetrated by large NFbundles that create prominent ridges through the calyxbefore spreading out into a network of fine filaments. Thepresynaptic NF bundles do not overlay the large postsyn-aptic clumps of somatic spines as defined by a7-nAChRclusters, but instead the two display largely complemen-tary distributions. The presynaptic NF bundles do, how-ever, codistribute with prominent bundles of microtubulesin the calyx. This latter finding, together with the spacingof the large NF bundles, suggests a dual function: thebundles may serve both a structural role in which they actas girders to secure the scaffold of fine filaments support-ing the extended calyx, and they may serve to defineconduits through the calyx for microtubule-dependenttransport of materials.
The ability of the anti-NF antibodies to discriminatebetween NFs in the distal preganglionic axons and pre-synaptic calyx on the one hand from those in the postgan-glionic cell bodies and proximal postganglionic axons onthe other is probably the result of differences in the stateof NF phosphorylation. Previous studies with phosphory-lation state-dependent anti-NF antibodies have shownthat NFs in distal axons are much more phosphorylatedthan those found in the cell body and proximal axon(Sternberger and Sternberger, 1983; Lee et al., 1987; Dahlet al., 1988). The antibodies used in the present studieswere in all cases raised against either phosphorylated NFsor against phosphorylated and unphosphorylated NFscombined. The fact that mAb NE14 is reported not torecognize dephosphorylated NF whereas mAb N52 can doso is consistent with the results obtained here showingpreferential calyx labeling by NE14 and preferential cellbody labeling by N52. The anti-NF-H antiserum N4142,which also prefers phosphorylated NF, showed a labelingpattern here similar to that of NE14.
Use of the anti-NF antibodies to visualize NF distribu-tion in the calyx during development revealed a progres-sive maturation of the cytoskeleton infrastructure definedby the large NF bundles. Initially a single major NF bun-dle representing the incoming axon was apparent, fol-lowed developmentally by a bifurcation in which a secondmajor branch began circling the postsynaptic cell in theopposite direction. Often a lateral spreading of the majorNF bundle could also be seen at the site of the bifurcation,and numerous fine NF branches were beginning to extendover the cell body. This early pattern is consistent withvariations in the 3D structure inferred from the develop-ing calyx from scanning electron microscopy on E10 gan-glia (Fujiwara and Nagakuro, 1989). By late development,most of the postsynaptic cell surface becomes covered bythe fine branches, and several large NF bundles extendover the surface as well. Often the two largest branchesappear to rejoin on the far side of the cell, suggesting that
290 J. PAYSAN ET AL.
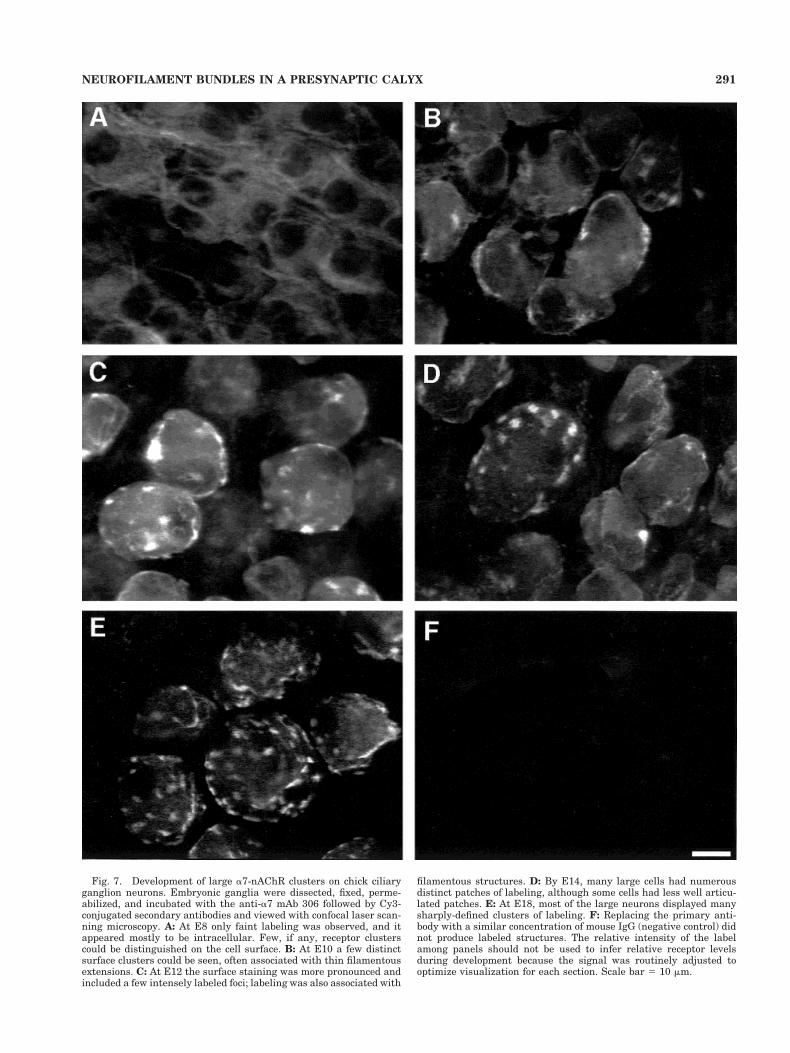
Fig. 7. Development of large a7-nAChR clusters on chick ciliaryganglion neurons. Embryonic ganglia were dissected, fixed, perme-abilized, and incubated with the anti-a7 mAb 306 followed by Cy3-conjugated secondary antibodies and viewed with confocal laser scan-ning microscopy. A: At E8 only faint labeling was observed, and itappeared mostly to be intracellular. Few, if any, receptor clusterscould be distinguished on the cell surface. B: At E10 a few distinctsurface clusters could be seen, often associated with thin filamentousextensions. C: At E12 the surface staining was more pronounced andincluded a few intensely labeled foci; labeling was also associated with
filamentous structures. D: By E14, many large cells had numerousdistinct patches of labeling, although some cells had less well articu-lated patches. E: At E18, most of the large neurons displayed manysharply-defined clusters of labeling. F: Replacing the primary anti-body with a similar concentration of mouse IgG (negative control) didnot produce labeled structures. The relative intensity of the labelamong panels should not be used to infer relative receptor levelsduring development because the signal was routinely adjusted tooptimize visualization for each section. Scale bar 5 10 mm.
291NEUROFILAMENT BUNDLES IN A PRESYNAPTIC CALYX
the ensheathing membrane structure is contiguous atleast in these locations.
It is not clear whether the entire NF network becomesenclosed in a single continuous membrane compartmentwrapped around the postsynaptic cell. Previous dye-fills ofcalyces with Lucifer yellow via the preganglionic nerveroot in ganglia from newly hatched chicks produced im-ages at the light microscope level that varied from asmooth cup enclosing the postsynaptic cell in some casesto a connected series of discrete labeled compartments(Stanley, 1987). Similar results were seen with dye-fillsvia a patch electrode onto calyces in E14 ganglia (Cogganet al., 1997). Since such dyes are expected to reflect cyto-plasmic volume, the labeled compartments could be con-nected by thin sheets of cytoplasm that would not bereadily detected, as noted at the time. A similar explana-tion could account for the compartmentalized distributionof labeling seen in the present studies with neurobiotindelivered via the presynaptic nerve root. In this lattercase, however, the distribution of labeling would also beinfluenced by the efficiency with which neurobiotin, whichis actively transported by intracellular machinery, wasretained by fixation to adjacent components.
Preganglionic axon labeling with DiI in the presentexperiments should have indicated the distribution of
lipid bilayer comprising the calyx outer surface, and, al-though the DiI labeling did suggest a more continuouspresynaptic structure than that seen with either Luciferyellow or neurobiotin, some patchiness was still apparent.This strongly suggests that calyx cross-sectional area isfar from uniform around the cell. Electron microscopicanalysis reveals small protrusions from the calyx inter-digitating among the postsynaptic somatic spines on cili-ary neurons (Shoop et al., 1999). Close packing of finemembrane-delineated processes could comprise a signifi-cant portion of the calyx. Discontinuities in the calyxstructure could have the advantage of providing sites atwhich rapid ion exchange can occur between extracellularspaces and the synaptic cleft, thereby minimizing thekinds of calcium depletion in the cleft that can limit syn-aptic transmission (Borst and Sakmann, 1999).
The non-overlap of large presynaptic NF bundles withthe postsynaptic spine clumps defined by a7-nAChR clus-ters may result from steric occlusion. The spine clumpsextend into the calyx (Shoop et al., 1999) and may imposespatial constraints by virtue of the substantial indenta-tion that results. Alternatively, large NF bundles may beexcluded from the immediate vicinity of transmitter re-lease sites because of high local calcium concentrationsthat occur there. These have been postulated to activate
Fig. 8. Complementary distribution of presynaptic NF bundlesand postsynaptic a7-nAChR clusters. Embryonic ciliary gangliawere dissected, fixed, permeabilized, and costained with mousemAb NE14 for NF protein (green) and with goat anti-a7 antibodiesfor a7-nAChRs (red) followed by appropriate fluorescent secondaryantibodies. A–C: E15, confocal fluorescence micrographs. D–F: E20, con-ventional fluorescence micrographs after deconvolution of the images. Asingle field of view is shown in each row, illustrating the distribution
of a7-nAChRs (left panel), calyx NF bundles (middle panel), and thetwo together (right panel). Points of coincident labeling (yellow) arerare; most of those shown represent fortuitous overlay of the twomarkers in different planes of focus on the reconstructed cell image.The deconvoluted image at E20 also shows the complex meshwork offine NF branches that does overlay a7-nAChR clusters. Scale bar 510 mm.
292 J. PAYSAN ET AL.
calcium-sensitive proteases that remove NFs from thesynaptic terminal (Roots, 1983; Walker et al., 1985). Anetwork of fine NFs visible with the electron microscope,however, has been reported in close proximity to activezones at the squid giant synapse (Martin, 1996). Thepresent results do not exclude the possibility that fine NFbranches also project to release sites overlying the spineclumps.
As remarked above, the spacing of the large NF bundleswould seem to optimize their ability to act as girdersproviding structural support for the calyx. A less obviousrole is that the bundles may also define a series of conduitsfor directing transport of materials and facilitating rapidtransmission of electrical signals. The finding that micro-tubules track with the NF bundles provides support forthis view. In addition, the increased cross-sectional area ofthe conduits defined by the bundles should enable currentto travel rapidly along such paths, observing the sameprinciples that account for current spreading more rapidlythrough large-bore axons than through small ones. Thebenefit of such an arrangement is that the conduits shouldfacilitate rapid spread of the current throughout the calyx,ensuring that release sites at the furthest edge of the calyxwill be stimulated synchronously with more proximal siteslocated between the conduits. It will be of interest infuture studies to investigate factors controlling the spac-ing and configuration of NF bundles in the calyx and to
understand how the resulting infrastructure influencesspecific presynaptic functions.
Note added in proof: Though mAb 306 appeared specific for a7-nAChRprotein when used for immunochemical staining in the present experi-ments, this may have depended critically on the great abundance of suchreceptors on chick ciliary ganglion neurons. In other tissues expressinglower levels of receptor and requiring longer exposure times to image, wefind the antibody appears to recognize other cellular components as well.
LITERATURE CITED
Blumenthal EM, Shoop RD, Berg DK. 1999. Developmental changes in thenicotinic responses of ciliary ganglion neurons. J Neurophysiol 81:111–120.
Borst JGG, Sakmann B. 1999. Depletion of calcium in the synaptic cleft ofa calyx-type synapse in the rat brainstem. J Physiol (Lond) 521:123–133.
Chiappinelli VA, Giacobini E. 1978. Time course of appearance ofa-bungarotoxin binding sites during development of chick ciliary gan-glion and iris. Neurochem Res 3:465–478.
Coggan JS, Paysan J, Conroy WG, Berg DK. 1997. Direct recording ofnicotinic responses in presynaptic nerve terminals. J Neurosci 17:5798–5806.
Conroy WG, Berg DK. 1995. Neurons can maintain multiple classes ofnicotinic acetylcholine receptors distinguished by different subunitcompositions. J Biol Chem 270:4424–4431.
Corriveau RA, Berg DK. 1994. Neurons in culture maintain acetylcholinereceptor levels with far fewer transcripts than in vivo. J Neurobiol25:1579–1592.
Fig. 9. Codistribution of major NF and microtubule bundles in thecalyx. E14 ciliary ganglia were dissected, fixed, permeabilized, andcostained with N4142 for NF bundles (red) and with mAb TUJ1 forb3-tubulin (green) followed by appropriate fluorescent secondary an-tibodies. Conventional fluorescence microscopy was used to generatewhole-cell images by deconvoluting and assembling multiple optical
sections taken along the z-axis. A single field of view is shown in eachrow (A–C,D–F), illustrating the distribution of NF bundles (left pan-el), of microtubules (middle panel), and extended codistribution of thetwo (right panel, yellow). Arrows indicate examples of major overlapof large NF and microtubule bundles in calyces. Scale bar 5 10 mm.
293NEUROFILAMENT BUNDLES IN A PRESYNAPTIC CALYX

Dahl D, Labkovsky B, Bignami A. 1988. Neurofilament phosphorylation inaxons and perikarya: immunofluorescence study of the rat spinal cordand dorsal root ganglia with monoclonal antibodies. J Comp Neurol271:445–450.
de Waegh SM, Lee VM, Brady ST. 1992. Local modulation of neurofilamentphosphorylation, axonal caliber, and slow axonal transport by myeli-nating Schwann cells. Cell 68:451–463.
Dryer S. 1994. Functional development of the parasympathetic neurons ofthe avian ciliary ganglion: a classic model system for the study ofneuronal differentiation and development. Prog Neurobiol 43:281–322.
Fuchs E, Cleveland DW. 1998. A structural scaffolding of intermediatefilaments in health and disease. Science 279:514–519.
Fujiwara T, Nagakuro C. 1989. Three-dimensional structure of the presyn-aptic nerve ending in the ciliary ganglion of the chick embryo: a scan-ning electron microscopic study. Neurosci Lett 98:125–128.
Held H. 1983. Die zentrale Hoerleitung. Arch Anat Physiol p 201.Jacob MH, Berg DK. 1983. The ultrastructural localization of
a-bungarotoxin binding sites in relation to synapses on chick ciliaryganglion neurons. J Neurosci 3:260–271.
Jacob MH, Berg DK, Lindstrom JM. 1984. Shared antigenic determinantbetween the Electrophorus acetylcholine receptor and a synaptic com-ponent on chicken ciliary ganglion neurons. Proc Natl Acad Sci USA81:3223–3227.
Kassner PD, Conroy WG, Berg DK. 1998. Organizing effects of rapsyn onneuronal nicotinic acetylcholine receptors. Mol Cell Neurosci 10:258–270.
Landmesser L, Pilar G. 1972. The onset and development of transmissionin the chick ciliary ganglion. J Physiol (Lond) 222:691–713.
Landmesser L, Pilar G. 1974a. Synapse formation during embryogenesison ganglion cells lacking a periphery. J Physiol (Lond) 241:715–736.
Landmesser L, Pilar G. 1974b. Synaptic transmission and cell death dur-ing normal ganglionic development. J Physiol (Lond) 241:737–749.
Lee VM-Y, Carden M, Schlaepfer W, Trojanski J. 1987. Monoclonal anti-bodies distinguish several differentially phosphorylated states of thetwo largest rat neurofilament subunits (NFH and NFM) and demon-strate their existence in the normal nervous system of adult rats.J Neurosci 7:3474–3488.
Loring RH, Zigmond RE. 1987. Ultrastructural distribution of 125I-toxin Fbinding sites on chick ciliary neurons: synaptic localization of a toxinthat blocks ganglionic nicotinic receptors. J Neurosci 7:2153–2162.
Martin R. 1996. The structure of the neurofilament cytoskeleton in thesquid giant axon and synapse. J Neurocytol 25:547–554.
Martin AR, Pilar G. 1963. Dual mode of synaptic transmission in the avianciliary ganglion. J Physiol (Lond) 168:443–463.
Matus A. 1999. Postsynaptic actin and neuronal plasticity. Curr OpinNeurobiol 9:561–565.
Nixon RA, Paskevich PA, Sihag RK, Thayer CY. 1994. Phosphorylation ofcarboxyl terminus domains of neurofilament proteins in retinal gan-glion neurons in vivo: influences on regional neurofilament accumula-tion, interneurofilament spacing, and axon caliber. J Cell Biol 126:1031–1046.
Peters A, Palay SL, Webster H. 1976. The fine structure of the nervoussystem. Philadelphia: WB Saunders.
Ramon y Cajal S. 1908. Sur un noyau special du nerf vestibulaire despoissons et des oiseaux. Travaux du laboratoire de recherches bi-ologiques de l’Universite de Madrid. T VI p 1.
Roots BI. 1983. Neurofilament accumulation induced in synapses by leu-peptin. Science 221:971–972.
Sanes JR, Lichtman JW. 1999. Development of the vertebrate neuromus-cular junction. Annu Rev Neurosci 22:389–442.
Schoepfer R, Conroy WG, Whiting P, Gore M, Lindstrom J. 1990. Braina-bungarotoxin binding protein cDNAs and mAbs reveal subtypes ofthis branch of the ligand-gated ion channel gene superfamily. Neuron5:35–48.
Shoop RD, Martone ME, Yamada N, Ellisman MH, Berg DK. 1999. Neu-ronal acetylcholine receptors with a7 subunits are concentrated onsomatic spines for synaptic signaling. J Neurosci 19:692–704.
Smetters DK, Zador A. 1996. Synaptic transmission: noisy synapses andnoisy neurons. Curr Biol 6:1217–1218.
Stanley EF. 1987. Light microscopic visualisation of the presynaptic nerveterminal calyx in dissociated chick ciliary ganglion neurons. Brain Res421:367–369.
Sternberger LA, Sternberger NH. 1983. Monoclonal antibodies distinguishphosphorylated and nonphosphorylated forms of neurofilaments insitu. Proc Natl Acad Sci USA 80:6126–6130.
Toni N, Buchs P-A, Nikonenko I, Bron CR, Muller D. 1999. LTP promotesformation of multiple spine synapses between a single axon terminaland a dendrite. Nature 402:421–425.
Vernallis AB, Conroy WG, Berg DK. 1993. Neurons assemble acetylcho-line receptors with as many as three kinds of subunits while main-taining subunit segregation among receptor subtypes. Neuron 10:451– 464.
von Lenhossek M. 1911. Das ganglion ciliare der voegel. Arch MikroskAnat 76:745–769.
Walker JH, Boustead CM, Witzemann V, Shaw G, Weber K, Osborn M.1985. Cytoskeletal proteins at the cholinergic synapse: distribution ofdesmin, actin, fodrin, neurofilaments, and tubulin in Torpedo electricorgan. Eur J Cell Biol 38:123–133.
Wilson Horch HL, Sargent PB. 1995. Perisynaptic surface distribution ofmultiple classes of nicotinic acetylcholine receptors on neurons in thechicken ciliary ganglion. J Neurosci 15:7778–7795.
294 J. PAYSAN ET AL.
Copyright © 2022 FDOKUMEN







![Effect of different electrolytes on the swelling properties of calyx[4]pyrrole-containing polyacrylamide membranes](https://static.fdokumen.com/doc/165x107/631f4fc8d10f1687490fbd44/effect-of-different-electrolytes-on-the-swelling-properties-of-calyx4pyrrole-containing.jpg)












