
The histaminergic tuberomammillary nucleus is critical for motivated arousal

EXPERIMENTAL NEUROLOGY 37, 312-321 (1972)
Activation of Gracile Nucleus:
Time Distribution of Activity in Presynaptic and Postsynaptic Elements
D. WHITEHORN, Al. B. BROMBERG, J. F. HOWE, J. E. PLTTNAM
AND P. R. BURGESS r
Department of Physiology and Biophysics, Unizlersity of Vermont College of Mediciue, Bwlington, Vetwont 05401, and Department of Physiology, Unizfersity
of Utah Collc,c~c of Mrdiciur, Salt Lake City, Utah 84102
Received Jwze 14, 1972
The time required for impulse conduction from the skin to the gracile nucleus has been determined for primary afferent fibers in the sural and accessory sural nerves of the cat. Fibers were classified into types on the basis of their receptor properties. Fibers of a particular type were found to have a characteristic range of conduction times. Fibers of the same type in different nerves were found to have similar skin-to-nucleus conduction times despite differences in conduction distance. These data enable one to predict the temporal characteristics of the input to the nucleus evoked by an abrupt stimulus at the skin. Conduc- tion times from the sciatic nerve to the gracile nucleus were also determined. Comparison was made between the temporal dispersion of the input evoked by electrical stimulation of the sciatic nerve and the first spike latencies of post- synaptic cells in the nucleus excited by the same stimulus. The range of arrival times of the inrut closely matched the distribution of postsynaptic first spike latencies.
Introduction
When an abrupt stimulus is delivered to the skin, the activity evoked at all levels of the somatosensory system is dispersed in time. This dispersion begins at the level of the primary afferent fibers whose conduction veloci- ties range from less than 1 m/set to more than 80 m/set. Several different receptor types are supplied by fibers in a cutaneous nerve and there is a correlation between receptor type and the range of conduction velocities of
r This work supported by Research Grants from US Public Health Service (NB 01576, NS 09472) and NSF (GB 82.53). Dr. Whitehorn was supported during part of this work by a postdoctoral fellowship from Public Health Service (NB 38731). Our thanks to Miss Katharine Fenn for help in preparation of the manuscript. Dr. Howe’s address is Department Neurological Surgery, University of Washington, Seattle, Washington 98105.
312 Copyright 0 1972 by Academic Press, Inc. All rights of reproduction in any form reserved.

GRACILE NUCLEUS 313
its fibers (6). Thus the volley of activity arriving at a central nucleus after an abrupt stimulus will be dispersed in time and arrival time will be re- lated to fiber type. Such dispersion may have several consequences (13). For example, activity arriving relatively early can influence the effects of later inputs, but the reverse cannot occur. In addition, the relationship be- tween fiber types and conduction velocity, and hence arrival time, can be used as a tool in determining the wiring diagram of a nucleus.
In this paper we examine the temporal characteristics of the direct pri- mary afferent input to the nucleus gracilis from hind limb cutaneous nerves. This is compared with the temporal distribution of postsynaptic ac- tivity in the nucle~~s. The relationship between arrival time at the nucleus and fiber type is also described.
Methods
The experiment employed adult cats (approximately 3.5 kg), anesthe- tized with intraperitoneal injection of sodium pentobarbital (Nembutal, Abbot, 35 mg/kg ) , paralyzed with Flaxedil (American Cyanimid) and ar- tificially respired. Blood pressure was measured via a cannula inserted into the carotid artery. In some experiments, expired CO, was measured and end tidal values were maintained between 4 and 5%. Rectal temperature was kept between 37 and 38 C with external heat.
In separate experiments, standard microelectrode techniques were used to record from (a) single myelinated fibers in two cutaneous nerves inner- vating haired skin (accessory sural and main sural) and (b) single post- synaptic elements in the gracile nucleus. Exposed neural tissues were cov- ered with mineral oil and paraffin wax.
In experiments involving primary afferent fibers, a bipolar stimulating electrode was placed on the sciatic nerve at the sciatic notch. Peripheral conduction velocities were determined between the sciatic electrode (0.1 msec duration stimuli) and a more distal recording site in the main or ac- cessory sural nerve. Fibers ranging in conduction velocity from S-80 m/set were sampled. Classification of fibers into fiber types was done on the basis of tests clecribed in Results. Identification of fibers supporting collaterals reaching the gracile nucleus and measurement of antidromic conduction times from the nucleus were accomplished using monopolar stimulation of the exposed dorsal columns at the first cervical level (S j .
In the gracile nucleus, presynaptic elements were distinguished from postsynaptic neurons by their ability to follow electrical shocks to the sciatic nerve at rates greater than 500/set and the appearance of a single evoked spike. Presynaptic recordings in the gracile nucleus were discarded and all data on primary afferent fibers are derived from units isolated in peripheral nerve.

314 WHITEHORN ET AL.
Results
Shin-to-N,ucleus Conduction Times and their Rehtion to Fiber Type. Determination of the temporal characteristics of the primary afferent input to the gracile nucleus after brief (0.1 msec) electrical stimulation at the skin was accomplished by measuring conduction times of individual fibers. Fibers in the accessory sural nerve were used for this study and were clas- sified into types on the basis of their receptor characteristics (2. S-7, 15 ; for recent reviews: 3, 4). For the purpose of this report, only fibers that support a collateral reaching the nucleus gracilis are described (1. S, 12). Four fiber types were defined; all were sensitive mechanoreceptors. Since classification was carried out with the hair clipped, hair and field fibers with similar adequate stimuli were not distinguished.
Type II fibers were the only projecting type with a slowly adapting dis- charge in response to a maintained deformation of the skin. Their proper- ties are well described and include a relatively regular discharge pattern, sensitivity to skin stretch and a localized receptive field.
The G, hair fibers were distinguished by a brief discharge which was evoked only by high velocity movement across the clipped hairs.
The intermediate hair-F, field class responded only to movement. They displayed a relatively good ability to follow 540 Hz vibration in a 1 :l fashion. These fibers were extremely sensitive, developing high-frequency, irregular discharges at the slightest stimulation. Tapping the leg outside the receptive field of the nerve usually evoked one or two spikes from these fibers.
The Gs-field fibers displayed a range of mechanical sensitivities. They detected slower movements than the G, hair or intermediate hair-F, field groups. Some of the less mechanically sensitive G,-field fibers could sup- port a week (.S/sec or less) tonic discharge for a few seconds.
Forty percent of the accessory sural G,-field fibers studied (29 of 72) did not support collaterals reaching the gracile nucleus. In this characteris- tic they differed from type II, intermediate hair-F, field and G, hair fibers, all of which projected to the nucleus.
The conduction time from the skin to the gracile nucleus for a particular fiber was determined by measuring the antidromic latency obtained at the peripheral recording site after stimulation of the dorsal funiculi at C, and adding to this the latency obtained upon stimulating in the center of the fiber’s receptive field on the skin with needle electrodes.
Table 1 shows the results of these experiments on the accessory sural nerve. While a relationship exists between fiber type and skin-to-nucleus conduction time. there is considerable overlap, particularly between G, hair, type II, and intermediate hair-F, field fibers. Among G,-field fibers, a tendency was evident for fibers with clearly “field” characteristics (better

GRACILE NUCLEUS 315
response to actual skin stimulation than hair movement) to have longer conduction times. The longest conduction times (25-35 msec) were ob- tained from fibers with some weak tonic discharge in response to steady pressure.
Antidrowzic and Ortlaodrowic Conduction T&x. The results given in table 1 are antidromic conduction times. This raises the question of the comparability of antidromic and orthodromic conduction times since the latter is the one of interest and the former the one measured. To test for differences, a collision experiment was conducted on fibers of each of the four types. .\n orthodromic impulse was initated by a stimulating electrode placed on the peripheral nerve distal to. and as close as possible to, the mi- croelectrode recording site. (Conduction time from this peripheral stimulus site to the recording site was 0.1-0.2 msec.) At intervals after this, an an- tidromic spike was elicited from the dorsal funiculi at Cl. A critical inter- val was determined at which collision of the orthodromic and antidromic spikes occurred, indicated by a sudden disappearance of the antidromic spike at the recording site. If this value is reduced by the refractory period at the dorsal funiculus-stimulating site, the orthodromic conduction time is obtained. All fibers tested were capable of following lOOO/sec stimulation at the dorsal fuaiculus stimulus site for more than two stimuli. A refrac- tory period of 1 msec was thus assumed. For fibers with short conduction times ( 10 msec or less )) antidromic and orthodromic conduction times dif- fered by less than 1 msec (10% error). Fibers with long conduction times (30 msec ) showed antidromic-orthodromic differences as large as 3 msec (10% error).
The data discussed refer to conduction along primary afferent fibers measured to the caudal border of the gracile nucleus (Cl 1. In a small sample of fibers, conduction times were measured from Cl to the most ros- tral point in the nucleus from which antidromic activation could be oh- tained. Values ranging from 0.4-1.5 msec were found.
TABLE 1
RANGE OF CONDUCTION TIMES FROM SKIN TO CAUDAL BORDER OF GRACILE NUCLEUS FOR FIBERS PROJECTING. DIRECTLY TO THE NUCLEUS FROM
THE ACCESSORY SURAL NERVE
Fiber type Range of conduction times (msec‘,
.~ ~- -~ Intermediate hair-F, field 12-15
G1 hair 12-15
Type II slowly adapting 13-15
G? hair and field 14-35

316 WHITEHORN ET AL
Comparison of Skiwto-Nucleus Conduction Times from Two Nemes. The conduction times shown in Table 1 pertain to fibers originating from the accessory sural nerve. The main sural nerve contains similar fiber types and the contribution of different types to the direct primary afferent input to the gracile nucleus is also similar (12). However, in considering temporal characteristics of the input to the gracile nucleus from these two nerves, an important difference was noted. Fibers in the sural nerve, origi- nating from more distal portions of the hind limb, must traverse a longer distance to the nucleus than do those of the accessory sural. Therefore. we compared skin-to-nucleus conduction times in fibers of the same type in each nerve. Thirty sural nerve fibers were used for this comparison; mem- bers of all the major types being surveyed. For strict comparison both nerves were mounted on the same recording platform so that the conduc- tion distance from Cl to the recording site (popliteal fossa) was virtually the same for both nerves.
The consistent finding of this comparison was that fibers of the same type in each nerve had similar skin-to-nucleus conduction times. However, differences in conduction time over portions of the pathway were observed. Fibers in the sural nerve displayed antidromic latencies from Cl to the re- cording site that were 2-5 msec shorter than those obtained from acces- sory sural fibers. The shorter Cl to recording site conduction time for sural fibers exactly balanced the longer skin to recording site conduction time. Hence, skin-to-nucleus conduction times were the same for fibers of the same type in each nerve and the results shown in Table 1 apply to both nerves.
In the following section, conduction times are reported from the sciatic nerve to the caudal border of the nucleus (Cl). The findings presented above indicate that for fibers of the same type, those originating from nerves innervating more distal portions of the limb will have shorter sciatic-to-nucleus conduction times than will fibers from more proximal nerves.
Comparison of Temporal Characteristics in Presynaptic and Postsynap- tic Populations in the Gracile Nucleus. Figure 1A shows the distribution of conduction times from the sciatic nerve to the first cervical level of the spinal cord for 84 single fibers originating from the sural nerve. The data were obtained by measuring antidromic conduction times for individual primary afferent fibers recorded in the sural nerve in response to supra- maximal stimulation of the surface of the dorsal funiculi at Cl. Since the recording site was distal to the sciatic notch the antidromic latency recorded upon stimulation of the sciatic nerve at the notch was subtracted from the Cl to recording site value to yield a sciatic-to-nucleus conduction time, shown in Fig. IA.
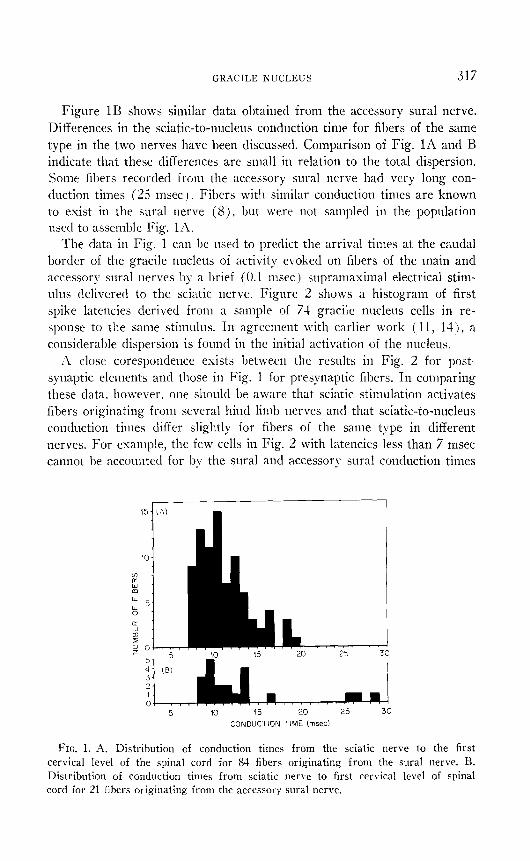
GRACILE KUCLEUS 317
Figure 1B shows similar data obtained from the accessory sural nerve. Differences in the sciatic-to-nucleus conduction time for fibers of the same type in the two nerves have been discussed. Comparison of Fig. 1A and B indicate that these differences are small in relation to the total dispersion. Some fibers recorded from the accessory sural nerve had very long con- duction times (25 msec ) Fibers with similar conduction times are known to exist in the sural nerve (S), but \vere not sampled in the population used to assemble Fig. la.
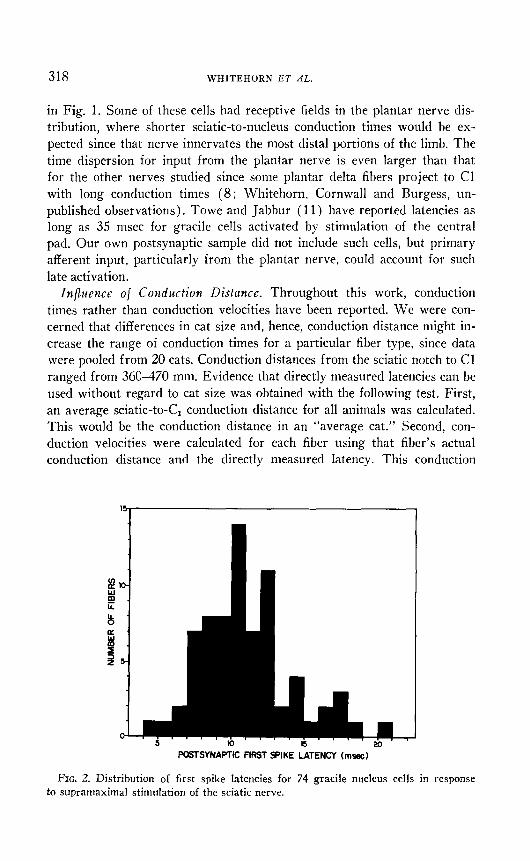
The data in Fig. 1 can he used to predict the arrival times at the caudal border of the gracile nucleus of activity evoked on fibers of the main and accessory sural nerves by a brief ( 0.1 msec ) supramaximal electrical stim- ulus delivered to the sciatic nerve. Figure 2 shows a histogram of first spike latencies derived from a sample of 74 gracile nucleus cells in re- sponse to the same stimulus. In agreement with earlier work ( 11, lit), a considerable dispersion is found in the initial activation of the nucleus.
~1 close corespondence exists between the results in Fig. 2 for post- synaptic elements and those in Fig. 1 for presynaptic fibers. In comparing these data, however, one sl~ould be aware that sciatic stimulation activates fibers originating from several hind limb nerves and that sciatic-to-nucleus conduction times differ slightly for fibers of the same type in different nerves. For example, the few cells in Fig. 2 with latencies less than 7 msec camlot be accounted for hy the sural and accessory sural conduction times
FIG. 1. A. Distribution of conduction times from the sciatic nerve to the first cervical level of the spinal cord for 83 fibers originating from the sural nerve. B. Distribution of conduction times from sciatic nerve to first cervical level of spinal cord for 21 fibers originating from the accessory sural nerve.
318 WHITEHORN ET AL.
in Fig. 1. Some of these cells had receptive fields in the plantar nerve dis- tribution, where shorter sciatic-to-nucleus conduction times would he ex- pected since that nerve innervates the most distal portions of the limb. The time dispersion for input from the plantar nerve is even larger than that for the other nerves studied since some plantar delta fibers project to Cl with long conduction times (8 : Whitehorn, Cornwall and Burgess, un- published observations). Towe and Jabbur (11) have reported latencies as long as 3.5 msec for gracile cells activated by stimulation of the central pad. Our own postsynaptic sample did not include such cells, but primary afferent input, particularly from the plantar nerve, could account for such late activation.
Influence of Conduction Distance. Throughout this work, conduction times rather than conduction velocities have been reported. We were con- cerned that differences in cat size and, hence, conduction distance might in- crease the range of conduction times for a particular fiber type, since data were pooled from 20 cats. Conduction distances from the sciatic notch to Cl ranged from 360370 mm. Evidence that directly measured latencies can be used without regard to cat size was obtained with the following test. First, an average sciatic-to-C, conduction distance for all animals was calculated. This would be the conduction distance in an “average cat.” Second, con- duction velocities were calculated for each fiber using that fiber’s actual conduction distance and the directly measured latency. This conduction
POSTSYNAPTIC FIRST SPIKE LATENCY (mssc)
FIG. 2. Distribution of first spike latencies for 74 gracile nucleus cells in response to supramaximal stimulation of the sciatic nerve.

GRACILE NUCLEUS 319
velocity was then used to calculate the latency that would have occurred in the average cat. These normalized latencies differed from directly meas- ured latencies by less than 1 msec.
Discussion
The present work describes quantitative measurements related to the temporal characteristics of the direct input to the gracile nucleus. The work is based on the view that temporal, as well as spatial, patterns of nerve activity underlie brain function.
Activity initiated by an abrupt stimulus at the skin reaches the gracile nucleus by way of four different fiber types. Two temporal characteristics of such an input have been shown to be related to fiber type. First, the time required for impulse conduction from the skin to the nucleus and sec- ond, the range of conduction times for fibers of a particular type (Table 1). In both of these characteristics, G, hair, intermediate-F, field, and type II fibers have similar properties. Fibers of these types conduct impulses to the nucleus in 12-15 msec. The narrow range of conduction times (3 msec) indicates that relatively little dispersion exists in the activity reach- ing the nucleus via these fiber types after an abrupt stimulus at the skin. The G,-field fibers, on the other hand. have longer conduction times (1635 msec), a larger range of conduction times and, hence, more disper- sion. These differences in dispersion may determine the ability of popula- tions of fibers of different types to convey detailed information about the temporal characteristics of a stimulus to postsynaptic elements. For esam- ple, a rapidly changing stimulus applied to a particular point on the skin will result in discrete changes in the postsynaptic response of cells inner- vated by a fiber population showing little temporal dispersion.
The relative arrival times of activity carried on different fiber types may also have important consequences for information processing in the nu- cleus. If stimulus conditions at the skin change abruptly, the change in input at the nucleus will first be felt from the more rapidlq- conducting fibers. These fibers may then influence cells in the nucleus, altering their response to later arriving activity. Evidence exists that presynaptic changes may also be imposed by the more rapidly conducting fibers on the terminals of slower conducting fibers before the latter’s activity reaches the nucleus (9).
An unexpected result was the finding that fibers of the same type in dif- ferent hind limb nerves had identical skin-to-nucleus conduction times de- spite differences in conduction distance. This has several important conse- quences. First, activity originating from different portions of the limb, simultaneously contacted by a stimulus, will reach the gracile nucleus at the same time. Further the temporal sequence of activity initiated at differ-

320 WHITEHORN ET AL.
ent points along the limb by a moving stimulus will be preserved in the temporal pattern of the input to the gracile nucleus.
Our esperiments have also shown that there is a close correspondence between the time dispersion of the input to the gracile nucleus and the dis- persion in first spike latencies for postsynaptic elements. The dispersion in first spike activation time could be due either to the presence of multi- synaptic connections within the nucleus, or to dispersion in the input. Our results do not resolve this question but do indicate that a relatively long activation latency cannot be used alone as a criterion for identifying multi- synaptically activated neurons,
The relationships between conduction time and fiber type offers aid in determining the fiber types responsible for the initial activation of a partic- ular postsynaptic cell. For example. a cell with an initial latency of 12 msec from the skin must have a significant input from fibers with equal or shorter conduction times. It can also be suggested that a relationship should occur between initial activation latency and the adequate stimulus for gracile neurons.
Finally, the direct measurement of conduction times is important. Ex- trapolations from histological data from peripheral nerve (10) or dorsal funiculi will produce spurious results since changes in conduction velocity occur along the conduction pathway (8, 12).
References
1. BROWN, A. G. 1968. Cutaneous afferent fiber collaterals in the dorsal columns of the cat. E.r). Brain Rcs. 5: 293-305.
2. BROWN, A. G., and A. Icco. 1967. A quantitative study of cutaneous receptors and afferent fibers in the cat and rabbit. J. Ph~&ol. (Lotion) 193: 707-733.
3. BURGESS, P. R. 1972. Cutaneous mechanoreception. In “Handbook of Perception.” Vol. 3, E. C. Carterette and M. P. Friedman [Eds.]. Academic Press (in press).
4. BURGESS, P. R., and E. R. PERL. 1972. Cutaneous mechanoreceptors and noci- ceptors. Ir, “Handbook of Sensory Physiology.” Vol. 2, A. Iggo [Ed.]. Springer (in press).
5. BURGESS, P. R., and E. R. PERL. 1967. Myelinated afferent fibres responding specifically to noxious stimulation of the skin. J. Pltysiol. 190: 541-562.
6. BURGESS, P. R., D. PETIT, and R. M. WARREN. 1968. Receptor types in cat hairy skin supplied by myelinated fibers. J. Newoplzysiol. 31: 833-848.
7. HCNT, C. C. and A. K. MCINTYRE. 1960. An analysis of fiber diameter and receptor characteristics of myelinated cutaneous afferent fibers in cat. J. Physiol. 153 : 99-112.
8. PETIT, D. and P. R. BURGESS. 1968. Dorsal column projection of receptors in cat hairy skin supplied by myelinated fibers. J. Newoplzysiol. 31 : 849-855.
9. PUTNAM, J. E. 1971. “Characteristics of Primary Afferent Depolarization at the Gracile Nucleus.” Thesis, Master of Sciences. University of Vermont.
10. TOWE. A. L. 1968. Neuronal population behavior in the somatosensory systems, pp. 552-574. In “The Skin Senses,” D. R. Kenshalo [Ed.]. Thomas, Springfield, Illinois.

GRACILE NUCLEUS ,321
11. TOWE, A. L., and S. J. JABBUR. 1961. Cortical inhibition of neurons in dorsal column nuclei of cat. J. h'cuvophysiol. 24: 48s-498.
12. WHITEHORN, D., M. C. CORNWALL and P. R. Br-RGESS. 1971. Central course of identified cutaneous afferents from cat hindlimb. I;rd. PI-oc. 30: (No. 2),
433. 13. WILLIAXS, \?I. J. 1972. Transfer Characteristics of Dispersive Nerve Eundles.
IEEE Transactions ou Systems, Man and Cybernetics. Vol. SIIC-I’, No. 1. 14. WINTER, D. L. 1965. N. Gracilis of cat. Functional organization and corticofugal
effects. .I. Nrwopkysiol. 28: 45-70. 15. WITT, I. and H. HENSEL. 1959. Afferent impulse aus der Extremitatenhaut der
Katze bei thermischer und mechanischer Reizung. .irrll. GES. Physiol. 286: 582-596.
Copyright © 2022 FDOKUMEN




















